
Growth and Leaf Color of Coleus under Light Conditions Modified by Translucent Agrivoltaic Panels and Light-Emitting Diodes in a Greenhouse

Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Environmental Conditions
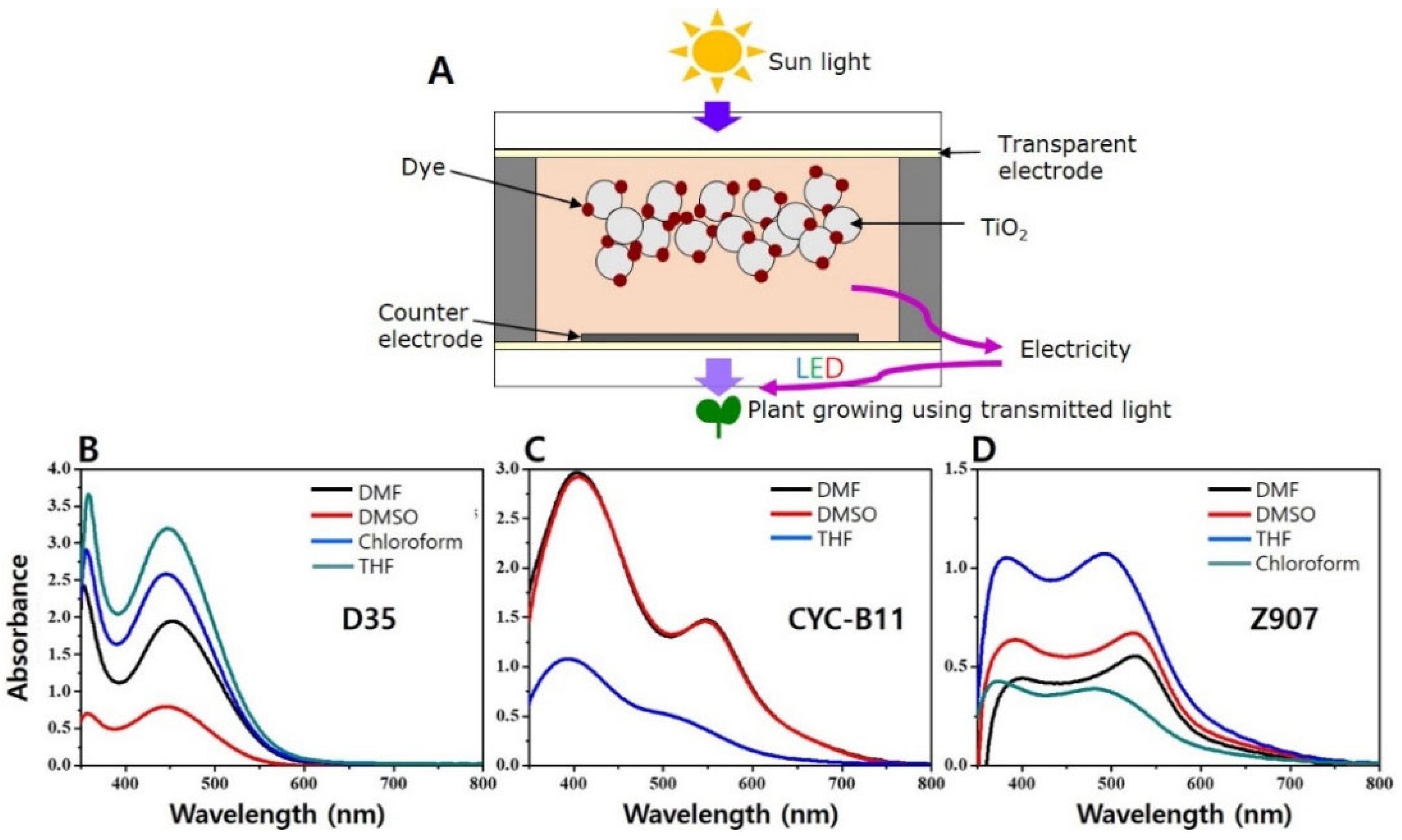
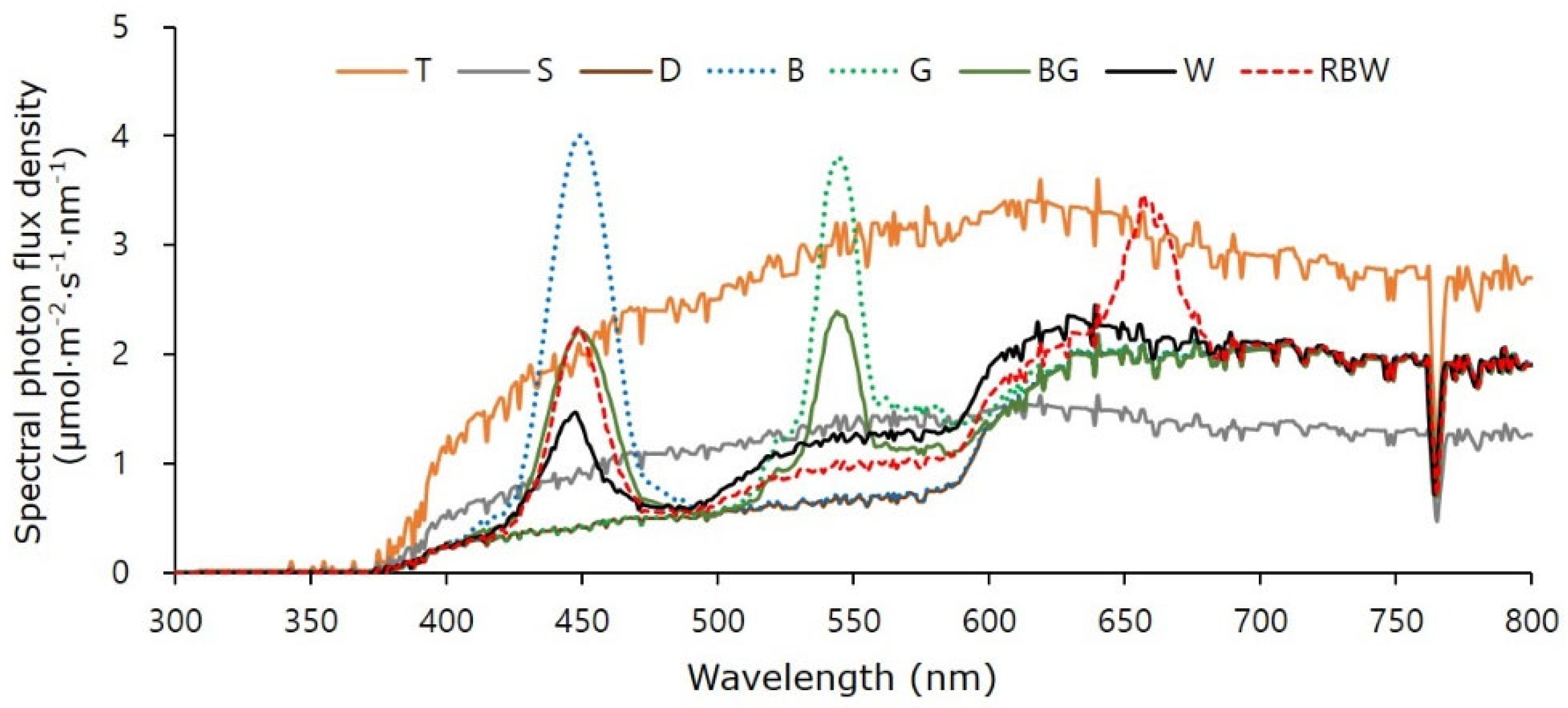
2.2. Light Treatment Using DSSCs and LEDs in a Greenhouse
2.3. Measurements and Statistical Analysis
3. Results and Discussion
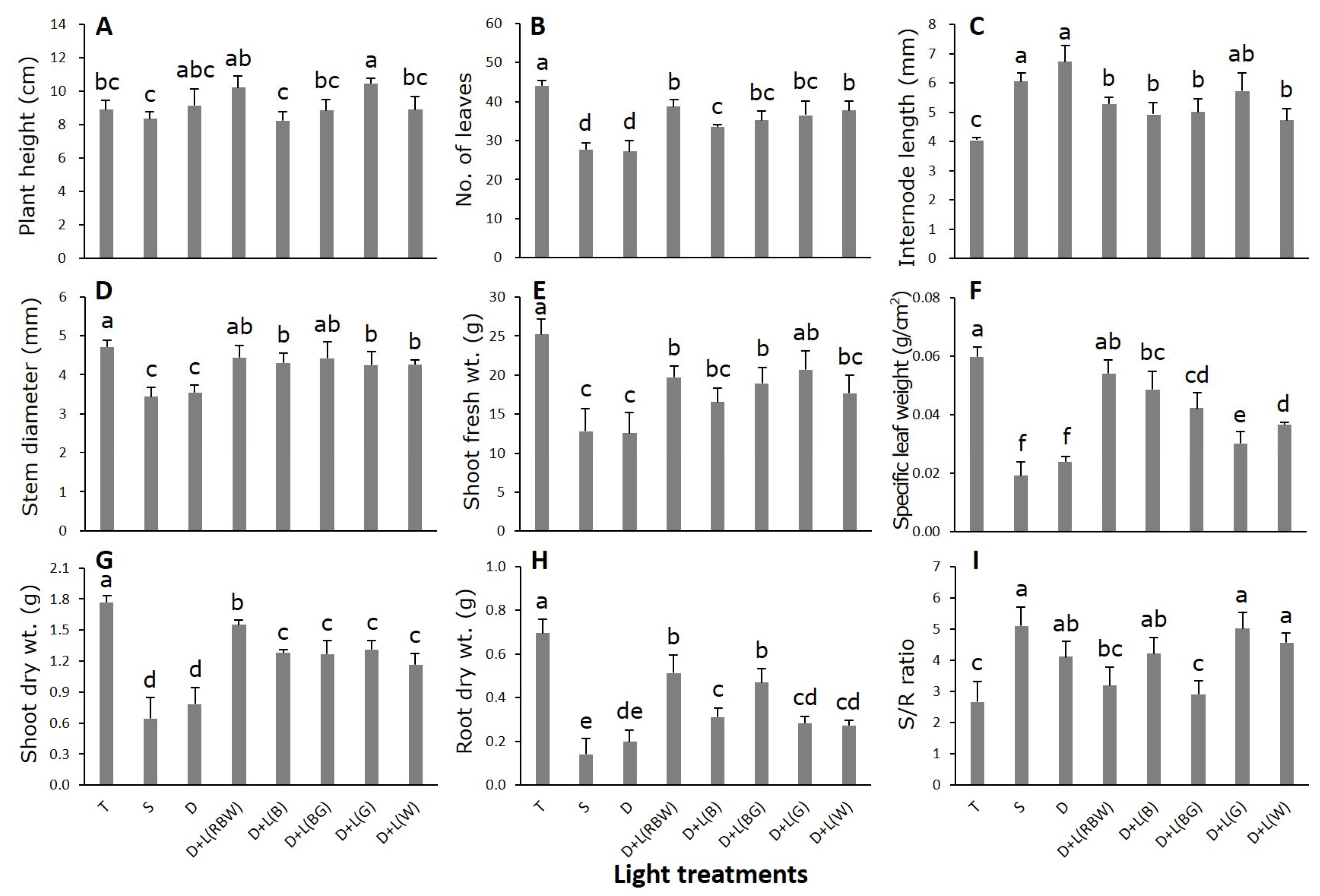
3.1. Growth and Morphological Characteristics
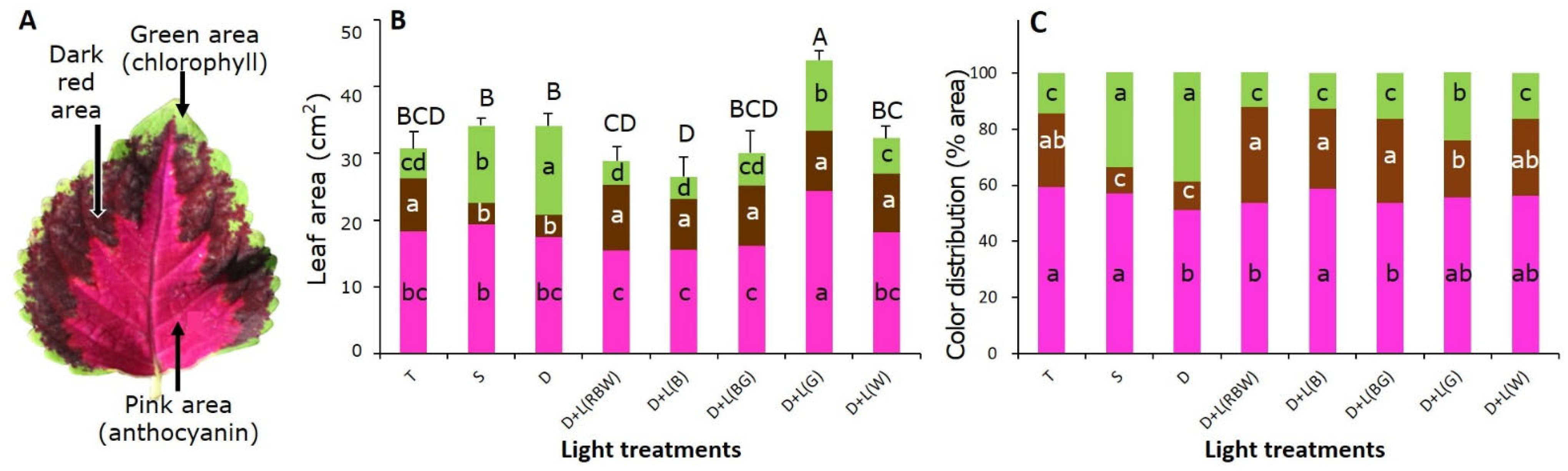
3.2. Leaf Area and Color Distribution
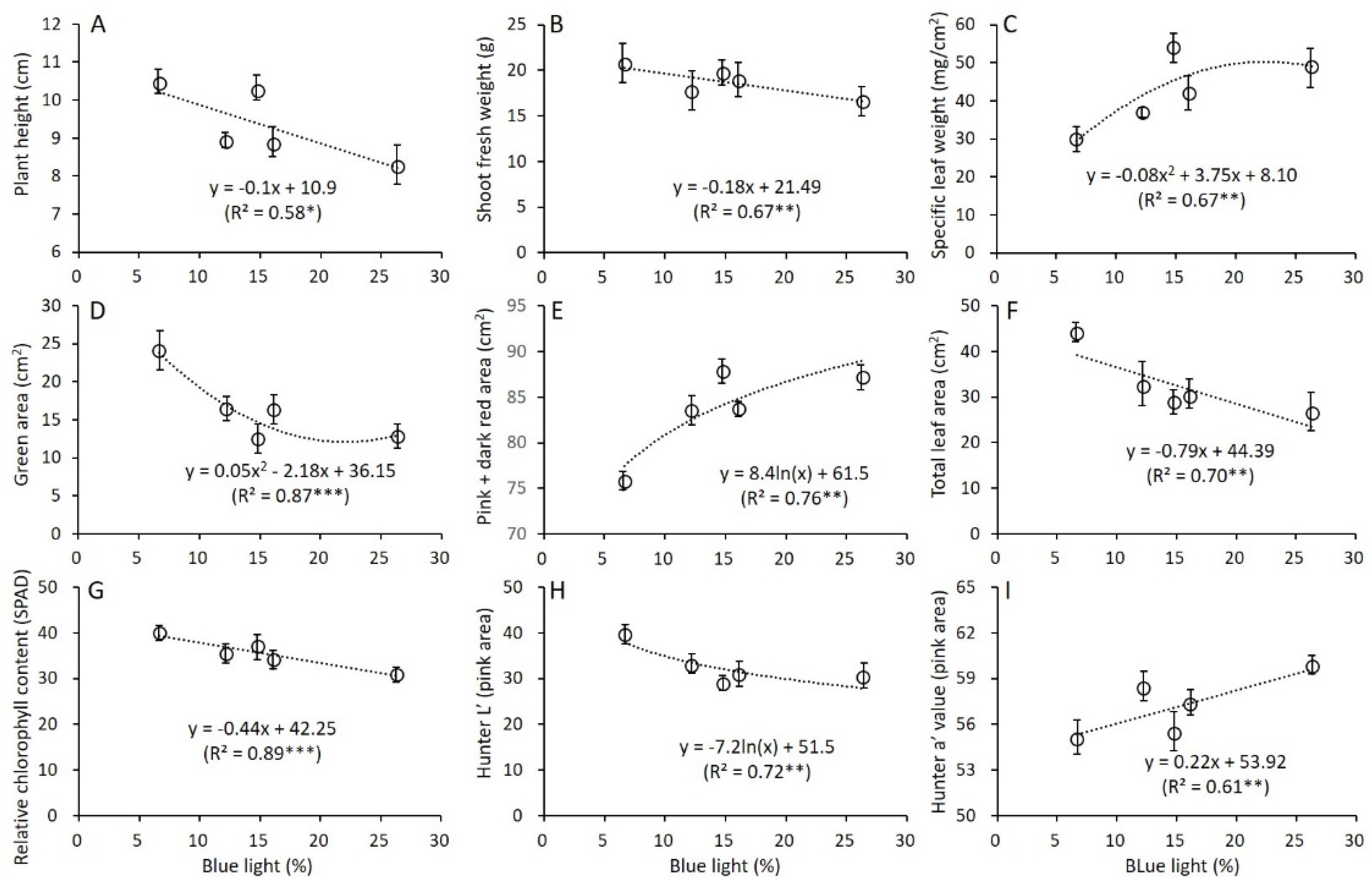
3.3. Correlation between Blue Light and Growth and Leaf Characteristics
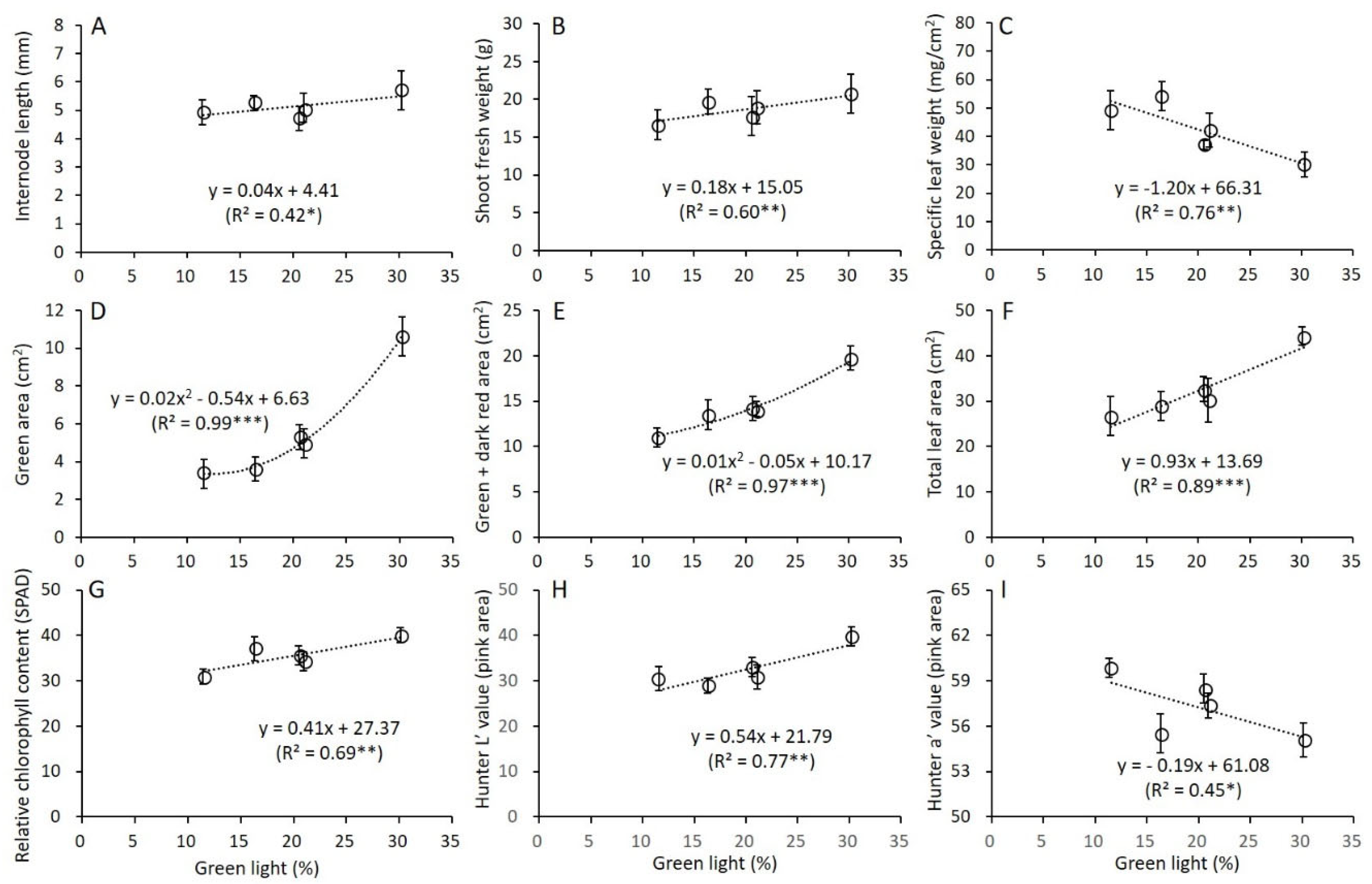
3.4. Correlation between Green Light and Growth and Leaf Characteristics
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Stallknecht, E.J.; Herrera, C.K.; Yang, C.; King, I.; Sharkey, T.D.; Lunt, R.R.; Runkle, E.S. Designing plant—Transparent agrivoltaics. Sci. Rep. 2023, 13, 1903. [Google Scholar] [CrossRef]
- Chalkias, D.A.; Charalampopoulos, C.; Aivali, S.; Andreopoulou, A.K.; Karavioti, A.; Stathatos, E. A di-carbazole-based dye as a potential sensitizer for greenhouse-integrated dye-sensitized solar cells. Energies 2021, 14, 1159. [Google Scholar] [CrossRef]
- Ki, H.C.; Jung, H.Y.; Gu, H.B. A study on the efficiency of dye sensitized solar cell employing TiO2 photoelectrode synthesized using basic catalyst. J. Korean Inst. Electr. Electron. Mater. Eng. 2013, 26, 736–740. [Google Scholar]
- Roslan, N.; Ya’acob, M.E.; Radzi, M.A.M.; Hashimoto, Y.; Jamaludin, D.; Chen, G. Dye sensitized solar cell (DSSC) greenhouse shading: New insights for solar radiation manipulation. Renew. Sustain. Energy Rev. 2018, 92, 171–186. [Google Scholar] [CrossRef]
- Kim, H.S.; Lee, J.; Shin, M. Effective refractive index of dye-sensitized solar cell using transmittance and reflectance measurements. Curr. Photovolt. Res. 2015, 3, 91–96. [Google Scholar]
- Moon, B.E.; Choi, E.G.; Kim, J.G.; Ryou, Y.S.; Yoon, Y.C.; Kim, H.T. Performance analysis of the TiO2 dye-sensitized solar cell according to seasonal changes. Prot. Hortic. Plant Fact. 2014, 23, 221–228. [Google Scholar] [CrossRef]
- Kim, J.J.; Kang, M.; Kwak, O.K.; Yoon, Y.J.; Min, K.S.; Chu, M.J. Fabrication and characterization of dye-sensitized solar cells for greenhouse application. Int. J. Photoenergy 2014, 2014, 376315. [Google Scholar] [CrossRef]
- Statistics Korea (KOSTAT). Agricultural Area Statistics in 2020; KOSTAT: Daejeon, Republic of Korea, 2021; pp. 66–69.
- Jeong, H.K.; Seong, J.H.; Kim, H.J. Agricultural policy tasks to achieve carbon neutrality. In Agricultural Outlook 2022; Korea Rural Economy Institute (KREI): Naju, Republic of Korea, 2022; pp. 109–131. [Google Scholar]
- Song, Y.H.; Jeong, S.R.; Go, E.; Moon, H.D.; Lim, H.J.; Noh, G.W.; Lee, J.P. Korea 2050 Carbon Neutrality Scenario K-Map: System Improvement Tasks and Budget Proposals to Achieve the 2030 K-Map Goal; Green Energy Strategy Research Institute, Green Transition Research Institute, Next Corporation, and Agora Energiewende: Seoul, Republic of Korea, 2023; pp. 5–17. [Google Scholar]
- Lopez, R.; Fisher, P.; Runkle, E. Introduction to specialty crop lighting. In Light Management in Controlled Environments; Lopez, R., Runkle, E., Eds.; Meister Media Worldwide: Willoughby, OH, USA, 2017; pp. 12–20. [Google Scholar]
- Hernandez, R.; Kubota, C. Light quality and photomorphogenesis. In Light Management in Controlled Environments; Lopez, R., Runkle, E., Eds.; Meister Media Worldwide: Willoughby, OH, USA, 2017; pp. 29–37. [Google Scholar]
- Wang, Y.; Folta, K.M. Contributions of green light to plant growth and development. Am. J. Bot. 2013, 100, 70–78. [Google Scholar] [CrossRef]
- Ahmad, M.; Grancher, N.; Heil, M.; Black, R.C.; Giovani, B.; Galland, P.; Lardemer, D. Action spectrum for cryptochrome-dependent hypocotyl growth inhibition in Arabidopsis. Plant Physiol. 2002, 129, 774–785. [Google Scholar] [CrossRef]
- Sng, B.J.R.; Mun, B.; Mohanty, B.; Kim, M.; Phua, Z.W.; Yang, H.; Lee, D.Y.; Jang, I.C. Combination of red and blue light induces anthocyanin and other secondary metabolite biosynthesis pathways in an age-dependent manner in Batavia lettuce. Plant Sci. 2021, 310, 110977. [Google Scholar] [CrossRef]
- Runkle, E.S.; Heins, R.D. Specific functions of red, far red, and blue light in flowering and stem extension of long day plants. J. Am. Soc. Hortic. Sci. 2001, 126, 275–282. [Google Scholar] [CrossRef]
- Runkle, E.S.; Heins, R.D. Stem extension and subsequent flowering of seedling grown under a film creating a far-red deficient environment. Sci. Hortic. 2002, 96, 257–265. [Google Scholar] [CrossRef]
- Park, I.S.; Cho, K.J.; Kim, J.; Cho, J.Y.; Lim, T.J.; Oh, W. Growth and flowering responses of petunia to various artificial light sources with different light qualities. Korean J. Hortic. Sci. Technol. 2016, 34, 55–66. [Google Scholar] [CrossRef]
- Li, M.; Coneva, V.; Robbins, K.R.; Clark, D.; Chitwood, D.; Frank, M. Quantitative dissection of color patterning in the foliar ornamental coleus. Plant Physiol. 2021, 187, 1310–1324. [Google Scholar] [CrossRef]
- Nguyen, P.; Cin, V.D. The role of light on foliage colour development in coleus (Solenostemon scutellarioides (L.) Codd). Plant Physiol. Biochem. 2009, 47, 934–945. [Google Scholar] [CrossRef]
- Rogers, R. Coleus: Rainbow Foliage for Containers and Gardens; Timber Press Inc.: Portland, OR, USA, 2008. [Google Scholar]
- Garland, K.F.; Burnett, S.E.; Stack, L.B.; Zhang, D. Minimum daily light integral for growing high-quality coleus. HortTechnology 2010, 20, 929–933. [Google Scholar] [CrossRef]
- Gusmawan, M.W.A.; Sitawati, S.; Karyawati, A.S. The effect of paclobutrazol concentrations in different shade levels on coleus plant leaves color. J. Agric. Eng. 2022, 11, 647–657. [Google Scholar] [CrossRef]
- Burger, J.; Edwards, G.E. Photosynthetic efficiency, and photodamage by UV and visible radiation, in red versus green leaf coleus varieties. Plant Cell Physiol. 1996, 37, 395–399. [Google Scholar] [CrossRef]
- Park, J.; Lee, P.; Ko, M.J. Design and fabrication of long-term stable dye-sensitized solar cells: Effect of water contents in electrolytes on the performance. Int. J. Precis. Eng. Manuf. Green Technol. 2019, 6, 125–131. [Google Scholar] [CrossRef]
- Yeoh, M.E.; Chan, K.Y. A review on semitransparent solar cells for real-life applications based on dye-sensitized technology. IEEE J. Photovolt. 2021, 11, 354–361. [Google Scholar] [CrossRef]
- Barichello, J.; Vesce, L.; Mariani, P.; Leonardi, E.; Braglia, R.; Di Carlo, A.; Canini, A.; Reale, A. Stable semi-transparent dye-sensitized solar modules and panels for greenhouse application. Energies 2021, 14, 6393. [Google Scholar] [CrossRef]
- Oh, W.; Runkle, E.S.; Warner, R.M. Timing and duration of supplemental lighting during the seedling stage influence quality and flowering in petunia and pansy. HortScience 2010, 45, 1332–1337. [Google Scholar] [CrossRef]
- Franklin, K.A. Shade avoidance. New Phytol. 2008, 179, 930–944. [Google Scholar] [CrossRef]
- Franklin, K.A.; Whitelam, G.C. Phytochromes and shade-avoidance responses in plants. Ann. Bot. 2005, 96, 169–175. [Google Scholar] [CrossRef]
- Zhang, T.; Maruhnich, S.A.; Folta, K.M. Green light induces shade avoidance symptoms. Plant Physiol. 2011, 157, 1528–1536. [Google Scholar] [CrossRef]
- Oh, W. Effects of temperature, photoperiod and light intensity on growth and flowering in Eustoma grandiflorum. Hortic. Sci. Technol. 2015, 33, 349–355. [Google Scholar] [CrossRef]
- Oh, W.; Cheon, I.H.; Kim, K.S.; Runkle, E.S. Photosynthetic daily light integral influences flowering time and crop characteristics of Cyclamen persicum. HortScience 2009, 44, 341–344. [Google Scholar] [CrossRef]
- Tan, T.; Li, S.; Fan, Y.; Wang, Z.; Raza, M.A.; Shafiq, I.; Wang, B.; Wu, X.; Yong, T.; Wang, X.; et al. Far-red light: A regulator of plant morphology and photosynthetic capacity. Crop. J. 2022, 10, 300–309. [Google Scholar] [CrossRef]
- Wilson, P.J.; Thompson, K.E.N.; Hodgson, J.G. Specific leaf area and leaf dry matter content as alternative predictors of plant strategies. New Phytol. 1999, 143, 155–162. [Google Scholar] [CrossRef]
- Schenkels, L.; Saeys, W.; Lauwers, A.; Maurice, P. Green light induces shade avoidance to alter plant morphology and increases biomass production in Ocimum basilicum L. Sci. Hortic. 2020, 261, 109002. [Google Scholar] [CrossRef]
- Johkan, M.; Shoji, K.; Goto, F.; Hashida, S.N.; Yoshihara, T. Blue light-emitting diode light irradiation of seedlings improves seedling quality and growth after transplanting in red leaf lettuce. HortScience 2010, 45, 1809–1814. [Google Scholar] [CrossRef]
- Ilias, I.F.; Rajapakse, N. The effects of end-of-the-day red and far-red light on growth and flowering of Petunia × hybrida ‘Countdown Burgundy’ grown under photoselective films. HortScience 2005, 40, 131–133. [Google Scholar] [CrossRef]
- Beckwith, A.G.; Zhang, Y.; Seeram, N.P.; Cameron, A.C.; Nair, M.G. Relationship of light quantity and anthocyanin production in Pennisetum setaceum cvs. Rubrum and Red Riding Hood. J. Agric. Food Chem. 2004, 52, 456–461. [Google Scholar] [CrossRef]
- Kadomura-Ishikawa, Y.; Miyawaki, K.; Noji, S.; Takahashi, A. Phototropin 2 is involved in blue light-induced anthocyanin accumulation in Fragaria × ananassa fruits. J. Plant Res. 2013, 126, 847–857. [Google Scholar] [CrossRef]
- Zhang, T.; Folta, K.M. Green light signaling and adaptive response. Plant Signal. Behav. 2012, 7, 75–78. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chamber | Light Conditions | Natural Light (%) | Supplemental Lighting (SL) with LEDs * |
|---|---|---|---|
| T | Transparent chamber (control) | 100 | An amount of 1–2 μmol·m−2·s−1 PPFD with scattered light for the same photoperiod (15 h) as LED treatments. |
| S | Chamber shaded by a neutral curtain | 40.9 | |
| D | Chamber covered with DSSCs | 40.4 | |
| D+L(B) | DSSC + blue LED | 41.2 | An amount of 60 μmol·m−2·s−1 PPFD with different colored LEDs in the DSSC chamber. The photoperiod was set to 15 h (05:00–20:00). The LEDs turned on when natural light was less than 300 μmol·m−2·s−1 PPFD. |
| D+L(G) | DSSC + green LED | 40.0 | |
| D+L(BG) | DSSC + blue and green LED | 40.7 | |
| D+L(W) | DSSC + cool white LED | 40.4 | |
| D+L(RBW) | DSSC + red, blue, and white LED | 41.3 |
| Light Chamber | Photon Flux Density (µmol∙m−2∙s−1) | Light Transmittance y (%) | Relative Spectral Distribution (%) | Ratio | DLI x (mol∙m−2∙d−1) | DLI (%) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| B z | G | R | FR | PAR | PAR + FR | PAR | PAR + FR | B | G | R | FR | B:R | R:FR | |||
| Transparent (T) | 196 | 297 | 317 | 269 | 810 | 1079 | 100 | 100 | 19 | 29 | 31 | 21 | 0.62 | 1.18 | 16.52 | 100 |
| Shaded (S) | 89 | 133 | 143 | 126 | 365 | 491 | 45.1 | 45.5 | 19 | 28 | 31 | 22 | 0.62 | 1.13 | 6.75 | 40.9 |
| DSSC (D) | 41 | 70 | 189 | 190 | 300 | 490 | 37.0 | 45.4 | 9 | 16 | 42 | 34 | 0.22 | 0.99 | 6.67 | 40.4 |
| D+L(B) | 162 | 71 | 190 | 192 | 423 | 615 | 52.2 | 57.0 | 26 | 12 | 31 | 31 | 0.85 | 0.99 | 9.47 | 57.3 |
| D+L(G) | 41 | 186 | 191 | 191 | 418 | 609 | 51.6 | 56.4 | 7 | 30 | 31 | 31 | 0.21 | 1.00 | 9.36 | 56.7 |
| D+L(BG) | 99 | 130 | 188 | 189 | 417 | 606 | 51.5 | 56.2 | 16 | 21 | 31 | 31 | 0.53 | 0.99 | 9.44 | 57.1 |
| D+L(W) | 75 | 127 | 218 | 193 | 420 | 613 | 51.9 | 56.8 | 12 | 21 | 35 | 31 | 0.34 | 1.13 | 9.40 | 56.9 |
| D+L(RBW) | 91 | 101 | 238 | 192 | 430 | 622 | 53.1 | 57.6 | 15 | 16 | 39 | 31 | 0.38 | 1.24 | 9.61 | 58.2 |
| Light Chamber z | Hunter’s Value | Relative Chlorophyll Content (SPAD) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Pink Area | Dark-Red Area | Green Area | ||||||||
| L | a | b | L | a | b | L | a | b | ||
| T | 35.86 c y | 59.21 ab | 7.79 a | - x | - | - | 56.92 a | −21.65 a | 40.10 a | 31.2 c |
| S | 41.98 a | 57.44 bc | 2.06 b | - | - | - | 50.51 b | −21.19 a | 33.04 b | 37.6 ab |
| D | 39.95 ab | 56.46 c | 3.04 b | - | - | - | 47.01 c | −22.09 a | 35.17 ab | 39.5 a |
| D+L(RBW) | 28.92 e | 55.44 cd | 9.42 a | 16.86 b | 13.13 a | 2.56 b | - | - | - | 37.1 ab |
| D+L(B) | 30.33 de | 59.83 a | 7.14 a | 21.30 a | 12.34 a | 6.08 a | - | - | - | 30.8 c |
| D+L(BG) | 30.85 de | 57.36 bc | 8.82 a | 17.20 b | 12.38 a | 3.11 b | - | - | - | 34.2 bc |
| D+L(G) | 39.63 ab | 55.06 d | 7.65 a | - | - | - | 53.62 b | −21.74 a | 37.97 ab | 39.9 a |
| D+L(W) | 32.93 d | 58.39 ab | 6.76 a | 20.11 a | 13.37 a | 4.33 ab | - | - | - | 35.5 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, S.; Kim, J.; Oh, W. Growth and Leaf Color of Coleus under Light Conditions Modified by Translucent Agrivoltaic Panels and Light-Emitting Diodes in a Greenhouse. Horticulturae 2024, 10, 115. https://doi.org/10.3390/horticulturae10020115
Park S, Kim J, Oh W. Growth and Leaf Color of Coleus under Light Conditions Modified by Translucent Agrivoltaic Panels and Light-Emitting Diodes in a Greenhouse. Horticulturae. 2024; 10(2):115. https://doi.org/10.3390/horticulturae10020115
Chicago/Turabian StylePark, Suhyun, Jiseon Kim, and Wook Oh. 2024. "Growth and Leaf Color of Coleus under Light Conditions Modified by Translucent Agrivoltaic Panels and Light-Emitting Diodes in a Greenhouse" Horticulturae 10, no. 2: 115. https://doi.org/10.3390/horticulturae10020115
APA StylePark, S., Kim, J., & Oh, W. (2024). Growth and Leaf Color of Coleus under Light Conditions Modified by Translucent Agrivoltaic Panels and Light-Emitting Diodes in a Greenhouse. Horticulturae, 10(2), 115. https://doi.org/10.3390/horticulturae10020115