Abstract
Growth and flower bud initiation (FBI) were studied in single-stem plants of four biennial-fruiting cultivars in a controlled environment and under field conditions at 60°40′ N. Shoot growth varied widely among the cultivars but was significantly enhanced by high temperature (20 °C) in all cultivars, whereas photoperiod had a subordinate growth effect. FBI data from bud dissection after 6 weeks of cultivation in the phytotron were used to calculate FBI indices for the various cultivars and environment conditions. The indices also varied much among the cultivars but were enhanced by elevated temperature, being highest in ‘Natchez’ and ‘Sweet Royalla’, while ‘Natchez’ was the only cultivar in which FBI was significantly enhanced by short days. The non-vigorous and erect growing ‘Ouachita’ remained vegetative at both temperatures but flowered in spring after overwintering at 0.5 °C. The field experiment confirmed the superior growth vigor of ‘Loch Ness’ and ‘Sweet Royalla’ as well as the photoperiodic sensitivity of ‘Natchez’. The results also confirmed that floral initiation starts in lateral buds located 10–20 nodes below the apex, and from there it progresses in both acropetal and basipetal direction. We conclude that temperature is at least as important as the photoperiod for the control of FBI in biennial-fruiting blackberries.
1. Introduction
While the physiology of flowering in red raspberry has been thoroughly investigated and described [1], little is known about the environmental control of flowering in the genetically related blackberry (Rubus subgenus Rubus Watson). In view of the great economic importance of blackberry production [2], it is surprising that so little is known about the specific effects of environmental factors in the control of flower bud initiation (FBI) in blackberry plants. However, flowering phenology has been studied in a range of biennial-fruiting blackberry cultivars under diverse natural environment conditions for identification of the seasonal timing of FBI (e.g., Refs. [3,4,5,6,7,8,9,10]). The results show that the transition from vegetative to generative development takes place during late autumn and winter the year before flowering and fruiting. Because of this seasonal timing, FBI has generally been thought to be a short day (SD) response induced by decreasing daylength in autumn ([7,10], and references therein). However, hardly anything is known about the effects of temperature (which also changes dramatically during autumn and winter) in parallel with the photoperiod. Nor is anything known about the interaction of photoperiod and temperature in the process. That would require physiological experiments conducted under controlled environment conditions. However, experimentation with the bulky and fast-growing blackberry plants requires spacious controlled environment facilities that are not readily available. This is apparently an important reason for the limited knowledge about the environmental factors that are triggering FBI in blackberry.
An experiment of this kind was recently published by Sønsteby and Heide [11] with three contrasting blackberry cultivars grown in daylight phytotron compartments at three temperatures under naturally decreasing autumn daylength. The authors comment on the practical problems involved in experimenting with blackberry plants and report that the plants had to be decapitated after a few weeks of growth at elevated temperature. The growth rate increased with increasing temperature over the 12–20 °C range in all cultivars, and in the erect-growing ‘Ouachita’, early cessation of growth took place at 12 °C. FBI took place at all temperatures in all cultivars except from ‘Ouachita’, which remained vegetative at 12 °C, while the earliest FBI was observed at 16 °C in all cultivars.
It was also found that FBI started in lateral buds located several nodes below the apex and from there it progressed in both acropetal and basipetal direction, as previously reported for biennial-fruiting red raspberry (cf. [12,13]). Sønsteby and Heide [11] suggested that temperature seems to be as important as photoperiod for FBI in blackberry, and that winter chilling tends to enhance flowering and yield potential in partially induced plants.
With the objective of providing more substantial information about the environmental control of FBI in biennial-fruiting blackberries, we have conducted two additional experiments during summer and autumn of 2023 with four cultivars. The main objectives were to identify the specific effects of temperature and photoperiod for FBI in biennial-fruiting blackberries, and secondly, to test out a plant cultivation system that might simplify experimentation with the bulky and complex blackberry plants. For this purpose, we conducted two separate but complementing experiments with single-stem plants of the same four cultivars, one under fully controlled environmental conditions in a phytotron, and the other under field conditions with naturally decreasing autumn temperature and photoperiod. The results from the two experiments are presented and discussed below.
2. Materials and Methods
2.1. General
Single-stem plants of the biennial-fruiting blackberry cultivars ‘Loch Ness’, ‘Natchez’, ‘Ouachita’, and ‘Sweet Royalla’ were used for both experiments. The plants were propagated from root adventitious buds in a greenhouse at 21 °C as described for raspberry plants [13]. From 1 June, when the plants were fully rooted in 0.3 L pots, they were transplanted into 3.5 L pots and raised as single-stem plants in an unheated plastic tunnel under natural day length conditions at the NIBIO Experimental Centre Apelsvoll in South-East Norway (60°40′ N, 10°50′ E). A coarse-textured sphagnum peat medium (NORGRO, Ridabu, Norway) was used as root substrate in both experiments, and the plants were fertigated daily with a complete fertilizer (YARA, Oslo, Norway) solution with conductivity EC = 1.0 mS cm−1. Each fertigation lasted for 5 min, and the frequency was adjusted three to five times per day according to plant size and weather conditions.
Both experiments were factorial with a split-plot design with environment conditions as main plots and cultivars as sub-plots. When appropriate, experimental data were subjected to analysis of variance (ANOVA) by standard procedures using a Minitab® Statistical Software program package (Release 17.2.1 MiniTab, MiniTab Inc., State College, PA, USA). Tukey’s test was used for the separation of means. Percentage values were always subjected to an arc sin transformation before the performance of the ANOVA.
2.2. Phytotron Experiment
The plants used for the phytotron experiment were raised as described above throughout June and July. Then, on 3 August, when the plants had reached an average height of 84, 73, 48, and 62 cm and produced 14, 15, 14, and 17 leaves, respectively, for ‘Loch Ness’, ‘Natchez’, ‘Ouachita’, and ‘Sweet Royalla’, the plants were transferred to the daylight phytotron of the Norwegian University of Life Sciences at Ås, Norway (59°40′, 10°45′ E) and exposed to temperatures of 12 and 20 °C at photoperiods of 10 and 20 h for six weeks. To enable continuous plant growth without tipping, the plants were ‘trailed’ onto supporting sticks during the treatments.
In the phytotron, the temperatures were controlled to ±1 °C, and a water vapor pressure deficit of 530 Pa was maintained at both temperatures. During daytime (08.00–18.00 h), all plants were grown in natural daylight compartments, while at night they were moved into adjacent growth rooms with darkness or low-intensity light from 70 W incandescent lamps (10 µmol quanta m−2 s−1) for photoperiodic manipulation. Whenever the photon flux density in the daylight compartments fell below approximately 150 µmol quanta m−2 s−1 (as on cloudy days), an additional 125 µmol quanta m−2 s−1 was automatically added using Philips HPI-T 400 W lamps (Lysimport.no, Oslo, Norway). Each treatment had three replications, each consisting of three plants of each cultivar (N = 9). The plants were organized on nine trolleys, each with one plant of each cultivar, to a total of nine trolleys. By the twice daily movements of the plant trolleys, their position in the rooms was changed every day.
During the experimental period, plant growth was monitored by weekly observations of shoot height and leaf number of all plants. After six weeks (on 14 September), one plant of each cultivar and replication was cut at the base and all lateral buds along the entire length of the main shoot were dissected under a stereo macroscope for determination of flower development stage. Fresh (non-fixed) buds were used, and the flower development stages were scored according to the scale used by Takeda and Wisniewski [5], where vegetative buds were designated stage 1, buds with early inflorescence bract primordia were designated stage 2, and buds with fully differentiated flowers with both stamen and carpel primordia as stage 7. Nodes with stage 2 or higher were considered generative. For quantification of FBI, an index of FBI was calculated as follows:
FBI index = No. of flowering nodes × Maximal flowering stage/2
The remaining two plants per replication were moved directly into a cold store with a temperature of 5 °C. After 3 weeks, the temperature was lowered to 0.5 °C at which temperature the plants were overwintered. When the risk of night frost was over in the following spring, the plants were moved outdoors and assessed for flowering performance. At the time of anthesis, as well as the plant flowering status, the number of flowering nodes and the total number of flowers were recorded for each plant.
2.3. Field Experiment
The same four cultivars were used in this experiment, and the plants were originally of the same pool as those of the phytotron experiment. On June 29, when the plants had reached a height of approximately 35 cm and produced about 10 leaves, 19 plants of each cultivar were placed outdoors under rain cover at the NIBIO Research Station Apelsvoll where they were grown under natural light conditions and ambient temperature for 14 weeks, as shown in Figure 1.
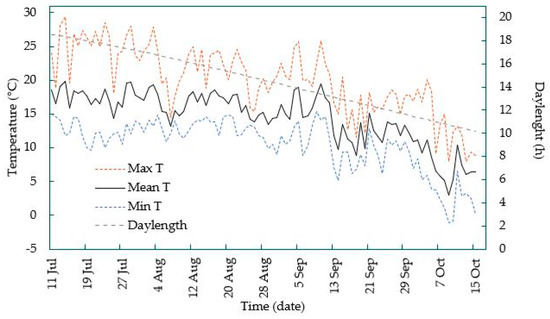
Figure 1.
Daily maximum, minimum, and mean temperatures (°C) and daylength (h) during plant cultivation in the field at NIBIO Apelsvoll (60° 40′ N).
Shoot growth and leaf (node) initiation were monitored by weekly observations on six of the nineteen plants of each cultivar as described for the phytotron plants. After four weeks of field cultivation, one axillary bud from three plants of each cultivar were sampled weekly (from 1 August to 16 October) for dissection and determination of floral development stage as described for the phytotron experiment. The buds were sampled by a shallow longitudinal slit, starting with bud number 10 from the base and moving upwards one position for each subsequent week. On 16 October, the 10 remaining plants of each cultivar were divided into two groups, where five plants were forced directly in a greenhouse at 20 °C and 20 h photoperiod and the other five plants were moved into a cold store and chilled for 16 weeks at 0.5 °C before forcing under the same conditions for recording of flowering behavior.
3. Results
3.1. Phytotron Experiment
While growth rate was strongly enhanced by high temperature, it varied widely between the cultivars, being highest in ‘Loch Ness’ and ‘Sweet Royalla’, followed by ‘Natchez’ and ‘Ouachita’, whereas photoperiod had a subordinate growth effect (Figure 2, Table 1). At 20 °C, all cultivars grew at almost constant but varying rates throughout the 6-week period, while at 12 °C they all ceased growing; ‘Ouachita’ and ‘Natchez’ within two weeks, and ‘Loch Ness’ and ‘Sweet Royalla’ after six weeks of cultivation. The appearance of the plants after six weeks of cultivation under the various conditions are presented in Figure 3, demonstrating that shoot growth was not different during ascending and descending periods of growth of the ‘trailed’ plants, while Table 1 shows an ANOVA of the incremental plant growth data at this stage. The results demonstrate highly significant differences in both plant height and node numbers among the cultivars, and a highly significant overall growth enhancement effect of elevated temperature, as well as a highly significant temperature × cultivar interaction. However, the main effect of photoperiod was subordinate, with no significant interaction with the cultivar (Table 1). Except for a couple of weeks’ delay in the appearance of the newly initiated leaves, the effects on shoot elongation and initiation of new leaves were nearly identical (Figure 2, Table 1).
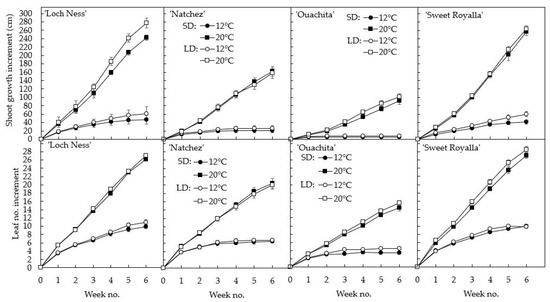
Figure 2.
Time courses of cumulative shoot growth (above) and leaf number increment (below) in four blackberry cultivars during six weeks of cultivation at 12 and 20 °C and 10 and 20 h photoperiod as indicated. The data are the means ± SE of three replications, each with three plants of each cultivar.

Table 1.
Probability levels of significance for main effects and interactions of temperature, photoperiod and cultivar on the incremental shoot growth and leaf number initiation in the blackberry cultivars ‘Loch Ness’, ‘Natchez’, ‘Ouachita’, and ‘Sweet Royalla’ after six weeks of treatment.

Figure 3.
Appearance of representative plants of the four cultivars after six weeks of cultivation under the various temperature and photoperiod conditions as indicated.
The floral development of one plant of each cultivar from each of the three replications is presented in Figure 4, demonstrating that while ‘Ouachita’ had not initiated floral primordia at any of the tested conditions, the other three cultivars had visible floral primordia at various stages of development under all tested conditions after six weeks of cultivation. The FBI indices presented show that the most advanced flower buds were found in ‘Natchez’, which at both temperatures was also advanced by SD conditions. Next in order was ‘Sweet Royalla’, with no specific effects of temperature and photoperiod, and finally, ‘Loch Ness’, in which initiation was generally slow but slightly advanced by elevated temperature in SD. Common for the three cultivars was, however, that at 12 °C, the most advanced buds were located approximately 10 to14 nodes above the shoot base with generative buds occurring almost up to the apex, whereas at 20 °C, 15–20 non-flowering nodes were subtending the apical bud (Figure 4).
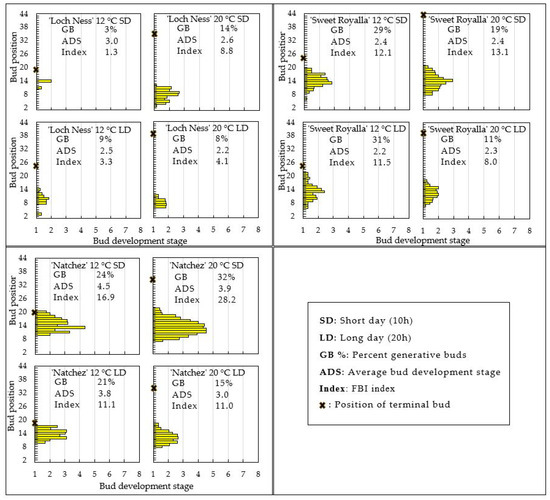
Figure 4.
Profiles of FBI and average stages of flower bud development of lateral buds at the various node positions along the entire shoot length, as well as FBI indices for four blackberry cultivars as affected by six weeks of exposure to temperature and photoperiod as indicated. The data are the means of three plants of each cultivar. No flower buds were observed in ‘Ouachita’.
When the remaining six plants of each cultivar were moved outside in the spring, all plants flowered to some extent, except one half of the ‘Natchez’ plants from 12 °C, which remained non-flowering (Table 2). Interestingly, all the ‘Ouachita’ plants, which remained vegetative in autumn, flowered to some extent in spring. Generally, the earliness of anthesis was consistent with the advancement of FBI recorded in the autumn, except for the ‘Natchez’ plants from 12 °C, which flowered much later than the rest. Surprisingly, all the ‘Natchez’ plants, which had the highest FBI indices in autumn, also had the lowest number of flowers per plant in spring, with values only shared with the ‘Ouachita’ plants, which had remained vegetative in autumn. Across all cultivars and environments, only the main effects of temperature and cultivar were highly significant for the number of flowers per plant, whereas for the percentages of flowering nodes and nonbreaking nodes, there was also a highly significant interaction of cultivar and temperature.
Table 2.
Days to anthesis and the flowering performance of four blackberry cultivars under natural outdoor conditions in the spring as affected by temperature and photoperiod in the previous autumn.
3.2. Field Experiment
As in the phytotron, growth and leaf production were largest in ‘Loch Ness’ and ‘Sweet Royalla’, followed by ‘Natchez’ and finally ‘Ouachita’ (Table 3, Figure 5). However, the small plant size of ‘Ouachita’ was mainly due to very short internodes, while the leaf numbers were only slightly less than in the ‘Natchez’ plants, which were nearly twice as tall. Generally, leaf number increment followed the same pattern as shoot growth, but due to some delay in the appearance of the initiated leaves, leaf appearance usually continued for 1–2 weeks after shoot growth had ceased. The cessation of growth was earliest in ‘Ouachita’ and ‘Natchez’, in which it took place by 1 September, while in ‘Loch Ness’ and ‘Sweet Royalla’, growth continued for another month until approximately 1 October (Figure 5).
Table 3.
Total shoot height and leaf (node) numbers of four blackberry cultivars after 14 weeks of cultivation under rain cover in the field under naturally decreasing autumn temperature and day length at 60°40′ N.
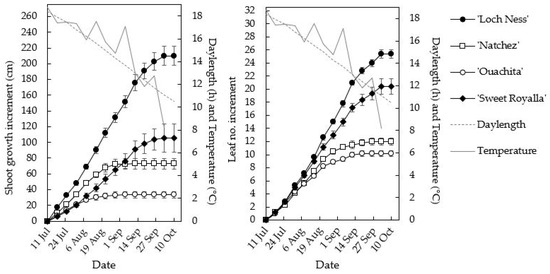
Figure 5.
Time courses of cumulative shoot growth and leaf number increment of four blackberry cultivars during 14 weeks of cultivation in the field under naturally decreasing temperature and daylength at NIBIO Apelsvoll (60°40′ N). The data are means ± SE of six replicate plants of each cultivar.
Initiation of flower buds was first observed in ‘Natchez’ on 1 September, followed by ‘Loch Ness’ and ‘Sweet Royalla’ one week later (Figure 6). ‘Natchez’ also reached the most advanced stages of flower development before the sampling was terminated after 15 weeks in mid-October. On the other hand, ‘Ouachita’ had not initiated any flower buds by that time.
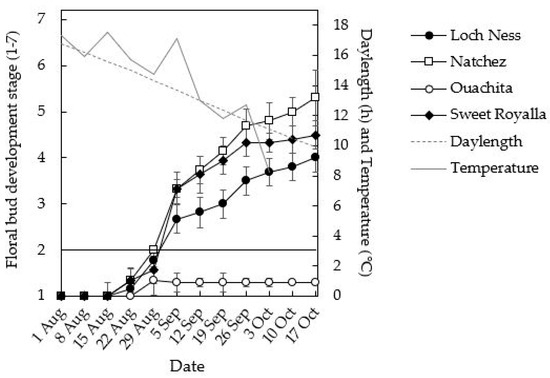
Figure 6.
Time courses of FBI in four blackberry cultivars during 14 weeks of cultivation under naturally decreasing autumn temperature and daylength at NIBIO Apelsvoll (60°40′ N). The data are means ± SD for three replicate plants of each cultivar. The horizontal line at development stage 2 denotes the earliest visible stage of FBI.
The flowering performance of the non-chilled and chilled plants from this experiment is presented in Table 4. It is seen that in the non-chilled plants, hardly any flowering took place in any cultivar after 60 days of forcing, whereas relatively good flowering took place in the chilled plants, apart from those of ‘Ouachita’, of which only a few plants flowered. Parallel effects were obtained for the percentage of flowering nodes per cane for all cultivars.
Table 4.
Effects of 16 weeks of chilling at 0.5 °C on flowering of four blackberry cultivars, which had previously been grown in the field for 14 weeks under naturally decreasing temperature and daylength at NIBIO Apelsvoll (60°40′ N).
4. Discussion
First, the phytotron experiment demonstrates that studies of FBI are problematic in the bulky and fast-growing blackberry plants. Although the results in Figure 2 and Figure 3 show that the trailing plants grew at identical rates during periods of both ascending and descending growth direction, the FBI results in Figure 4 suggest that FBI was suspended during the period of descending growth (see discussion below). In addition, such experiments are rather laborious and require spacious phytotron facilities.
However, as found in earlier experiments [7,8,11], the present phytotron experiment also demonstrated large differences in growth and flowering responses among the cultivars. While growth rate was greatly enhanced by elevated temperature in all cultivars, it increased significantly in order from the erect growing ‘Ouachita’ through the erect to semi-erect ‘Natchez’ and ‘Sweet Royalla’ to the trailing cultivar ‘Loch Ness’, whereas the main growth effect of photoperiod was subordinate (Table 1, Figure 2, Figure 3 and Figure 4). The duration of growth was also greatly different at 12 and 20 °C, continuing at constant but varying rates throughout the six-week period in all cultivars at 20 °C, while it ceased at 12 °C within two weeks in ‘Ouachita’ and ‘Natchez’ and within six weeks in ‘Sweet Royalla’ and ‘Loch Ness’. All these results agree closely with those reported for three of the same cultivars by Sønsteby and Heide [11] who found, however, that in ‘Ouachita’, ample FBI took place also at 16 °C, but not at 12 and 20 °C as in the present experiment. The most advanced floral stages were found at 20 °C in ‘Natchez’, which at both temperatures were advanced by SD, in contrast to ‘Sweet Royalla’, which, as in our earlier experiment [11], had quite advanced FBI at all environmental conditions (Figure 4). ‘Loch Ness’ had generally less advanced flower bud primordia than the other two cultivars under all conditions although it tended to be somewhat advanced by elevated temperature. The SD flowering enhancement of ‘Natchez’ supports the results of Sadojevic [14], who found a critical daylength of 14 h for both growth cessation and FBI at 16 °C, in contrast to ‘Loch Ness’, in which flowering was not affected by photoperiod. The cessation of growth and initiation of flower buds in ‘Natchez’ at approximately 1 September (Figure 5 and Figure 6), when the daylength at NIBIO Apelsvoll was about 14 h, are also in full agreement with these findings.
The flowering performance of the plants in the following spring (Table 2) showed, however, that all plants of ‘Ouachita’ from both temperatures flowered despite their lack of FBI in the fall, with most flowers in plants from 20 °C and LD. Another unexpected result was that the ‘Natchez’ plants, which had the most advanced and richest FBI in the fall, flowered poorly in spring, with only 50% flowering of the plants from 12 °C and SD conditions, and with low flower numbers and percentages of flowering nodes in plants from all conditions. This poor flowering of ‘Natchez’ in spring could possibly be due to low-temperature damage during cold storage of the advanced flower primordia present in this cultivar in the autumn (Figure 4). On the other hand, ‘Ouachita’, which did not initiate any flower buds in autumn, did flower to some extent in spring (Table 2). According to breeders at the University of Arkansas, the two cultivars do not differ markedly in winter hardiness [15,16,17], and therefore, their contrasting flowering behavior in spring seems most likely to be related to their contrasting earliness and advancement of FBI in the autumn. This assumption is also supported by the favorable effect of winter chilling in ‘Loch Ness’, which exhibited a slow and irregular FBI resulting in non-advanced floral primordia in the autumn (cf. Figure 4).
The observation that extended winter chill at near-freezing temperature in darkness not only breaks winter dormancy but also brings about FBI and anthesis in ‘Ouachita’ blackberry, as previously reported for cultivars of red raspberry ([18,19], cf. also [11]), is very interesting. This is a novel flowering response, whose physiological nature is not yet understood (cf. [19]). For blackberry production, however, it may be of large practical importance as a major flowering induction mechanism, which may also explain the commonly observed late FBI in biennial-fruiting blackberries under field conditions, which often does not take place until spring (cf. [3,4,5,6,7,10]). The response will be of particular interest for high tunnel production on overwintered long cane potted plants.
The dissection results also confirmed that FBI always started several nodes below the apex and from there, it progressed in both basipetal and acropetal direction, as also demonstrated for biennial-flowering red raspberries [12,13,20]. This concurs with the findings of Takeda et al. [8] that FBI starts at the mid-section of the shoot, and from there it can continue in both basipetal and acropetal directions. It was furthermore found that FBI in all cultivars started about 10–14 nodes above the ground level at both temperatures. However, while the FBI remained near the terminal bud in all cultivars at 12 °C, the process was terminated approximately 15 to 20 nodes below the apex at 20 °C (Figure 4). These results indicate that while the commencement of FBI takes place at a fixed location approximately 10–14 nodes above the shoot base, the termination of FBI relative to the apex varied with the prevailing growth temperature. When comparing the architecture of the plants after 6 weeks as shown in Figure 3, it is apparent that in all cultivars, the main difference between the plants at 12 and 20 °C corresponded to the descending part of the shoots at 20 °C. Since this also corresponds to the extra 15–20 vegetative nodes subtending the terminal bud in Figure 4, it suggests that no FBI had taken place during the descending part of growth in these long and trailing shoots. In other words, FBI seems to be suspended during periods with descending shoot growth.
The results also confirmed that cessation of growth is not a prerequisite for FBI in biennial-fruiting blackberry (cf. [11]). Thus, the early growth cessation of ‘Ouachita’ at 12 °C was not associated with concurrent FBI, whereas all the other cultivars initiated floral buds in the absence of growth cessation. This is in direct contrast to the strict association of the two processes in the closely related biennial-fruiting red raspberry [1,20].
5. Conclusions
In summary, we conclude that while shoot growth varies widely among biennial-fruiting blackberry cultivars, it was strongly enhanced by elevated temperature and ceased early at low temperature (12 °C), whereas it was less affected by photoperiod. We also conclude that although the control of FBI also varied much between the studied cultivars, temperature seems to be at least as important as photoperiod for the regulation of FBI. Especially important seems to be the flower-inducing effect of winter chill on partially induced plants. ‘Natchez’ was the only cultivar with a significant photoperiodic flowering response, behaving as a quantitative SD plant, whereas the erect growing ‘Ouachita’ was the only cultivar that remained vegetative until spring. The results also suggest that FBI was suspended during periods of descending shoot growth. However, the highly diverse flowering responses observed for the tested cultivars preclude any overall conclusions regarding the flowering control of biennial blackberries in general.
Author Contributions
Conceptualization, A.S. and O.M.H.; methodology, A.S., M.S. and O.M.H.; formal analysis, A.S. and M.S.; investigation, M.S. and A.S.; writing—original draft preparation, O.M.H.; writing—review and editing, A.S. and M.S.; project administration, A.S.; funding acquisition, A.S. All authors have read and agreed to the published version of the manuscript.
Funding
The Authors acknowledge financial support to the research leading to these results from the Norwegian Agricultural Agreement Research Fund/Foundation for Research Levy on Agricultural Products, grant number 326688, and Grofondet, grant number 200033.
Data Availability Statement
The raw data supporting the findings reported in this study are available on request from the corresponding author.
Acknowledgments
We acknowledge the skilful technical assistance of Kari Grønnerød with plant management.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Heide, O.M.; Sønsteby, A. Physiology of flowering and dormancy regulation in annual- and biennial-fruiting red raspberry (Rubus idaeus L.)—A review. J. Hortic. Sci. Biotechnol. 2011, 86, 433–442. [Google Scholar] [CrossRef]
- Strik, B.C.; Clark, J.R.; Finn, C.E.; Banados, P. Worldwide production of blackberries, 1995 to 2005 and prediction for growth. HortTechnology 2007, 17, 205–213. [Google Scholar] [CrossRef]
- Waldo, G.F. Fruit bud formation in brambles. Proc. Am. Soc. Hortic. Sci. 1933, 30, 263–267. [Google Scholar]
- Robertson, M. Further investigation of flower-bud development in the genus Rubus. J. Hortic. Sci. 1957, 32, 265–273. [Google Scholar] [CrossRef]
- Takeda, F.; Wisniewski, M. Organogenesis and patterns of floral bud development in two eastern thornless blackberry cultivars. J. Am. Soc. Hortic. Sci. 1989, 114, 528–531. [Google Scholar] [CrossRef]
- Warmund, M.R.; George, M.F.; Ellersieck, M.R.; Slater, J.V. Susceptibility of blackberry tissue to freezing injury after exposure to 16 °C. J. Am. Soc. Hortic. Sci. 1989, 114, 795–800. [Google Scholar] [CrossRef]
- Takeda, F.; Strik, B.C.; Peacock, D.; Clark, J.R. Cultivar differences and the effect of winter temperature on flower bud development in blackberry. J. Am. Soc. Hortic. Sci. 2002, 127, 495–501. [Google Scholar] [CrossRef]
- Takeda, F.; Strik, B.C.; Peacock, D.; Clark, J.R. Patterns of Floral Bud Development in Canes of Erect and Trailing Blackberries. J. Am. Soc. Hortic. Sci. 2003, 128, 3–7. [Google Scholar] [CrossRef]
- Moore, J.N.; Caldwell, J.D. 1985. Rubus. In Handbook of Flowering; Halevy, A.H., Ed.; CRC Press: Boca Raton, FL, USA, 1985; Volume IV, pp. 226–238. [Google Scholar]
- Strik, B.C. Flowering and fruiting on command in berry crops. Acta Hortic. 2012, 926, 197–214. [Google Scholar] [CrossRef]
- Sønsteby, A.; Heide, O.M. Temperature and daylength effects on growth and floral initiation in biennial-fruiting blackberry. Horticulturae 2023, 9, 1285. [Google Scholar] [CrossRef]
- Williams, I.H. Effects of environment on Rubus idaeus L. IV. Flower formation and development of the inflorescence. J. Hortic. Sci. 1959, 34, 219–228. [Google Scholar] [CrossRef]
- Sønsteby, A.; Heide, O.M. Environmental control of growth and flowering of Rubus idaeus L. cv. Glen Ample. Sci. Hortic. 2008, 117, 249–256. [Google Scholar] [CrossRef]
- Sadojevic, M. Growth and Development of Blackberry Cultivars. Master’s Thesis, Norwegian University of Life Sciences, Ås, Norway, 2024. Available online: https://hdl.handle.net/11250/3148386 (accessed on 21 November 2024).
- Clark, J.R.; Moore, J.N. ‘Ouachita’ thornless blackberry. HortScience 2005, 40, 258–260. [Google Scholar] [CrossRef]
- Clark, J.R.; Moore, J.N. ‘Natchez’ Thornless Blackberry. HortScience 2008, 43, 1897–1899. [Google Scholar] [CrossRef]
- Clark, J.R. ‘Osage’ Thornless Blackberry. HortScience 2013, 48, 909–912. [Google Scholar] [CrossRef]
- Palonen, P.; Pohjola, M.; Karhu, S. Cropping potential of raspberry long cane plants as affected by their growing condition and the duration of storage. J. Hortic. Sci. Biotechnol. 2015, 90, 738–746. [Google Scholar] [CrossRef]
- Sønsteby, A.; Heide, O.M. Estimating the temperature limitations for growth cessation and floral initiation in early- and late-inducing red raspberry cultivars (Rubus idaeus) and the floral-inducing effect of winter chilling. J. Hortic. Sci. Biotech. 2024. [Google Scholar] [CrossRef]
- Hodnefjell, R.; Heide, O.M.; Rivero, R.; Remberg, S.F.; Sønsteby, A. Control of growth cessation and floral initiation in red raspberry (Rubus idaeus L.) cultivars of diverse origin in controlled and natural environments. Sci. Hortic. 2018, 233, 412–420. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).