
Genotype-Specific Response to Silicon Supply in Young Tomato and Unripe Melon Plants Grown in a Floating System

 ,
,  ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growing Conditions
2.2. Biometric Parameters
2.3. Silicon Content
2.4. Experimental Design and Statistical Analyses
3. Results and Discussion
3.1. Biometric Parameters
3.2. Silicon Content
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bouis, H.E. Micronutrient fortification of plants through plant breeding: Can it improve nutrition in man at low cost? Proc. Nutr. Soc. 2003, 62, 403–411. [Google Scholar] [CrossRef] [PubMed]
- White, P.J.; Broadley, M.R. Biofortification of crops with seven mineral elements often lacking in human diets—iron, zinc, copper, calcium, magnesium, selenium and iodine. New Phytol. 2009, 182, 49–84. [Google Scholar] [CrossRef] [PubMed]
- Perez-Granados, A.M.; Vaquero, M.P. Silicon, aluminium, arsenic and lithium: Essentiality and human health implications. J. Nutr. Health Aging 2002, 6, 154–162. [Google Scholar] [PubMed]
- Deshmukh, R.K.; Ma, J.F.; Bélanger, R.R. Editorial: Role of silicon in plants. Front. Plant Sci. 2017, 8, 1858. [Google Scholar] [CrossRef] [PubMed]
- Alves, D.M.R.; de Mello Prado, R.; Barreto, R.F. Silicon and sodium attenuate potassium deficiency in Eruca sativa Mill. Food Chem. 2024, 432, 137225. [Google Scholar] [CrossRef] [PubMed]
- Arnon, D.; Stout, P. The essentiality of certain elements in minute quantity for plants with special reference to copper. Plant Physiol 1939, 14, 371–375. [Google Scholar] [CrossRef] [PubMed]
- Lewin, J.; Reimann, B.E.F. Silicon and Plant Growth. Annu. Rev. Plant Physiol. 1969, 20, 289–304. [Google Scholar] [CrossRef]
- Martos-García, I.; Fernández-Escobar, R.; Benlloch-González, M. Silicon is a non-essential element but promotes growth in olive plants. Sci. Hortic. 2024, 323, 112541. [Google Scholar] [CrossRef]
- Yan, G.C.; Nikolic, M.; Ye, M.J.; Xiao, Z.X.; Liang, Y.C. Silicon acquisition and accumulation in plant and its significance for agriculture. J. Integr. Agric. 2018, 17, 2138–2150. [Google Scholar] [CrossRef]
- Hosseini-Nasr, F.; Etesami, H.; Alikhani, H.A. Silicon Improves Plant Growth-Promoting Effect of Nodule Non-Rhizobial Bacterium on Nitrogen Concentration of Alfalfa Under Salinity Stress. J. Soil Sci. Plant Nutr. 2023, 23, 496–513. [Google Scholar] [CrossRef]
- Ma, J.F.; Yamaji, N. Silicon uptake and accumulation in higher plants. Trends Plant Sci. 2006, 11, 392–397. [Google Scholar] [CrossRef]
- Greger, M.; Landberg, T.; Vaculík, M. Silicon influences soil availability and accumulation of mineral nutrients in various plant species. Plants 2018, 7, 41. [Google Scholar] [CrossRef] [PubMed]
- Olivera-Viciedo, D.; Oliveira, K.S.; de Mello Prado, R.; Habermann, E.; Martínez, C.A.; de Moura Zanine, A. Silicon uptake and utilization on Panicum maximum grass modifies C:N:P stoichiometry under warming and soil water deficit. Soil Tillage Res. 2024, 235, 105884. [Google Scholar] [CrossRef]
- Heine, G.; Tikum, G.; Horst, W.J. Silicon nutrition of tomato and bitter gourd with special emphasis on silicon distribution in root fractions. J. Plant Nutr. Soil Sci. 2005, 168, 600–606. [Google Scholar] [CrossRef]
- Guntzer, F.; Keller, C.; Meunier, J.D. Benefits of plant silicon for crops: A review. Agron. Sustain. Dev. 2012, 32, 201–213. [Google Scholar] [CrossRef]
- López-Pérez, M.C.; Pérez-Labrada, F.; Ramírez-Pérez, L.J.; Juárez-Maldonado, A.; Morales-Díaz, A.B.; González-Morales, S.; García-Dávila, L.R.; García-Mata, J.; Benavides-Mendoza, A. Dynamic modeling of silicon bioavailability, uptake, transport, and accumulation: Applicability in improving the nutritional quality of tomato. Front. Plant Sci. 2018, 9, 647. [Google Scholar] [CrossRef] [PubMed]
- Zellner, W.L. Drastic differences in silicon concentrations between roots and leaves of 10 different Solanum lycopersicum L. varieties. HortScience 2021, 56, 838. [Google Scholar] [CrossRef]
- Haghighi, M.; Pessarakli, M. Influence of silicon and nano-silicon on salinity tolerance of cherry tomatoes (Solanum lycopersicum L.) at early growth stage. Sci. Hortic. 2013, 161, 111–117. [Google Scholar] [CrossRef]
- Li, H.; Zhu, Y.; Hu, Y.; Han, W.; Gong, H. Beneficial effects of silicon in alleviating salinity stress of tomato seedlings grown under sand culture. Acta Physiol. Plant. 2015, 37, 71. [Google Scholar] [CrossRef]
- Hoffmann, J.; Berni, R.; Hausman, J.F.; Guerriero, G. A review on the beneficial role of silicon against salinity in non-accumulator crops: Tomato as a model. Biomolecules 2020, 10, 1284. [Google Scholar] [CrossRef]
- Hernández-Salinas, M.; Valdez-Aguilar, L.A.; Alia-Tejacal, I.; Alvarado-Camarillo, D.; Cartmill, A.D. Silicon enhances the tolerance to moderate NaCl-salinity in tomato grown in a hydroponic recirculating system. J. Plant Nutr. 2022, 45, 413–425. [Google Scholar] [CrossRef]
- Shi, Y.; Zhang, Y.; Han, W.; Feng, R.; Hu, Y.; Guo, J.; Gong, H. Silicon enhances water stress tolerance by improving root hydraulic conductance in Solanum lycopersicum L. Front. Plant Sci. 2016, 7, 196. [Google Scholar] [CrossRef] [PubMed]
- Machado, J.; Fernandes, A.P.G.; Bokor, B.; Vaculík, M.; Heuvelink, E.; Carvalho, S.M.P.; Vasconcelos, M.W. The effect of silicon on the antioxidant system of tomato seedlings exposed to individual and combined nitrogen and water deficit. Ann. Appl. Biol. 2023, 184, 50–60. [Google Scholar] [CrossRef]
- Ali, N.; Schwarzenberg, A.; Yvin, J.C.; Hosseini, S.A. Regulatory role of silicon in mediating differential stress tolerance responses in two contrasting tomato genotypes under osmotic stress. Front. Plant Sci. 2018, 9, 1475. [Google Scholar] [CrossRef] [PubMed]
- Kiirika, L.M.; Stahl, F.; Wydra, K. Phenotypic and molecular characterization of resistance induction by single and combined application of chitosan and silicon in tomato against Ralstonia solanacearum. Physiol. Mol. Plant Pathol. 2013, 81, 1–12. [Google Scholar] [CrossRef]
- Fan, X.Y.; Lin, W.P.; Liu, R.; Jiang, N.H.; Cai, K.Z. Physiological response and phenolic metabolism in tomato (Solanum lycopersicum) mediated by silicon under Ralstonia solanacearum infection. J. Integr. Agric. 2018, 17, 2160–2171. [Google Scholar] [CrossRef]
- Kaya, C.; Tuna, A.L.; Guneri, M.; Ashraf, M. Mitigation effects of silicon on tomato plants bearing fruit grown at high boron levels. J. Plant Nutr. 2011, 34, 1985–1994. [Google Scholar] [CrossRef]
- Hu, A.Y.; Xu, S.N.; Qin, D.N.; Li, W.; Zhao, X.Q. Role of silicon in mediating phosphorus imbalance in plants. Plants 2021, 10, 51. [Google Scholar] [CrossRef]
- Zhang, Y.; Liang, Y.; Zhao, X.; Jin, X.; Hou, L.; Shi, Y.; Ahammed, G.J. Silicon compensates phosphorus deficit-induced growth inhibition by improving photosynthetic capacity, antioxidant potential, and nutrient homeostasis in tomato. Agronomy 2019, 9, 733. [Google Scholar] [CrossRef]
- Epstein, E. Silicon. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 641–664. [Google Scholar] [CrossRef]
- Liang, Y.; Nikolic, M.; Bélanger, R.; Gong, H.; Song, A. Silicon in Agriculture: From Theory to Practice; Springer Science + Business Media Dordrecht, Ed.; Springer: Dordrecht, The Netherlands, 2015; ISBN 9789401799782. [Google Scholar]
- Ma, J.F.; Yamaji, N. A cooperative system of silicon transport in plants. Trends Plant Sci. 2015, 20, 435–442. [Google Scholar] [CrossRef] [PubMed]
- Currie, H.A.; Perry, C.C. Silica in plants: Biological, biochemical and chemical studies. Ann. Bot. 2007, 100, 1383–1389. [Google Scholar] [CrossRef] [PubMed]
- Hodson, M.J.; White, P.J.; Mead, A.; Broadley, M.R. Phylogenetic variation in the silicon composition of plants. Ann. Bot. 2005, 96, 1027–1046. [Google Scholar] [CrossRef] [PubMed]
- D’Imperio, M.; Renna, M.; Cardinali, A.; Buttaro, D.; Santamaria, P.; Serio, F. Silicon biofortification of leafy vegetables and its bioaccessibility in the edible parts. J. Sci. Food Agric. 2016, 96, 751–756. [Google Scholar] [CrossRef] [PubMed]
- Mitani, N.; Jian, F.M. Uptake system of silicon in different plant species. J. Exp. Bot. 2005, 56, 1255–1261. [Google Scholar] [CrossRef] [PubMed]
- Fawe, A.; Menzies, J.G.; Chérif, M.; Bélanger, R.R. Silicon and disease resistance in dicotyledons. In Silicon in Agriculture; Datnoff, L.E., Snyder, G.H., Korndorfe, G.H., Eds.; Elsevier Science B.V.: Amsterdam, The Netherlands, 2001; pp. 159–169. [Google Scholar] [CrossRef]
- Etesami, H.; Jeong, B.R. Silicon (Si): Review and future prospects on the action mechanisms in alleviating biotic and abiotic stresses in plants. Ecotoxicol. Environ. Saf. 2018, 147, 881–896. [Google Scholar] [CrossRef] [PubMed]
- Debona, D.; Rodrigues, F.A.; Datnoff, L.E. Silicon’s Role in Abiotic and Biotic Plant Stresses. Annu. Rev. Phytopathol. 2017, 55, 85–107. [Google Scholar] [CrossRef]
- Buttaro, D.; Bonasia, A.; Minuto, A.; Serio, F.; Santamaria, P. Effect of silicon in the nutrient solution on the incidence of powdery mildew and quality traits in Carosello and barattiere (Cucumis melo L.) grown in a soilless system. J. Hortic. Sci. Biotechnol. 2009, 84, 300–304. [Google Scholar] [CrossRef]
- Liang, Y.; Sun, W.; Zhu, Y.G.; Christie, P. Mechanisms of silicon-mediated alleviation of abiotic stresses in higher plants: A review. Environ. Pollut. 2007, 147, 422–428. [Google Scholar] [CrossRef]
- Somma, A.; Palmitessa, O.D.; Leoni, B.; Signore, A.; Renna, M.; Santamaria, P. Extraseasonal production in a soilless system and characterisation of landraces of Carosello and Barattiere (Cucumis melo L.). Sustainability 2021, 13, 11425. [Google Scholar] [CrossRef]
- Palmitessa, O.D.; Durante, M.; Leoni, B.; Montesano, F.; Renna, M.; Serio, F.; Somma, A.; Santamaria, P. Enhancement of a landrace of Carosello (Unripe Melon) through the Use of light-emitting diodes (LED) and nutritional characterization of the fruit placenta. Sustainability 2021, 13, 11464. [Google Scholar] [CrossRef]
- Dallagnol, L.J.; Rodrigues, F.A.; Tanaka, F.A.O.; Amorim, L.; Camargo, L.E.A. Effect of potassium silicate on epidemic components of powdery mildew on melon. Plant Pathol. 2012, 61, 323–330. [Google Scholar] [CrossRef]
- Pozo, J.; Urrestarazu, M.; Morales, I.; Sánchez, J.; Santos, M.; Dianez, F.; Álvaro, J.E. Effects of silicon in thenutrient solution for three horticultural plant families on the vegetative growth, cuticle, and protection against Botrytis cinerea. HortScience 2015, 50, 1447–1452. [Google Scholar] [CrossRef]
- Preston, H.A.F.; Do Nascimento, C.W.A.; Preston, W.; de Souza Nunes, G.H.; Loureiro, F.L.C.; Mariano, R. de L.R. Silicon slag increases melon growth and resistance to bacterial fruit blotch. Acta Sci.-Agron. 2021, 43. [Google Scholar] [CrossRef]
- Xiao, J.; Li, Y.; Jeong, B.R. Foliar Silicon Spray before Summer Cutting Propagation Enhances Resistance to Powdery Mildew of Daughter Plants. Int. J. Mol. Sci. 2022, 23, 3803. [Google Scholar] [CrossRef] [PubMed]
- Renna, M.; D’Imperio, M.; Maggi, S.; Serio, F. Soilless biofortification, bioaccessibility, and bioavailability: Signposts on the path to personalized nutrition. Front. Nutr. 2022, 9, 2425. [Google Scholar] [CrossRef] [PubMed]
- Pavlovic, J.; Kostic, L.; Bosnic, P.; Kirkby, E.A.; Nikolic, M. Interactions of Silicon with Essential and Beneficial Elements in Plants. Front. Plant Sci. 2021, 12, 697592. [Google Scholar] [CrossRef] [PubMed]
- Powell, J.J.; McNaughton, S.A.; Jugdaohsingh, R.; Anderson, S.H.C.; Dear, J.; Khot, F.; Mowatt, L.; Gleason, K.L.; Sykes, M.; Thompson, R.P.H.; et al. A provisional database for the silicon content of foods in the United Kingdom. Br. J. Nutr. 2005, 94, 804–812. [Google Scholar] [CrossRef]
- Pannico, A.; El-Nakhel, C.; Graziani, G.; Kyriacou, M.C.; Giordano, M.; Soteriou, G.A.; Zarrelli, A.; Ritieni, A.; De Pascale, S.; Rouphael, Y. Selenium biofortification impacts the nutritive value, polyphenolic content, and bioactive constitution of variable microgreens genotypes. Antioxidants 2020, 9, 272. [Google Scholar] [CrossRef]
- Gonnella, M.; Renna, M.; D’imperio, M.; Santamaria, P.; Serio, F. Iodine biofortification of four brassica genotypes is effective already at low rates of potassium iodate. Nutrients 2019, 11, 451. [Google Scholar] [CrossRef]
- D’imperio, M.; Renna, M.; Cardinali, A.; Buttaro, D.; Serio, F.; Santamaria, P. Calcium biofortification and bioaccessibility in soilless “baby leaf” vegetable production. Food Chem. 2016, 213, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Voogt, W.; Holwerda, H.T.; Khodabaks, R. Biofortification of lettuce (Lactuca sativa L.) with iodine: The effect of iodine form and concentration in the nutrient solution on growth, development and iodine uptake of lettuce grown in water culture. J. Sci. Food Agric. 2010, 90, 906–913. [Google Scholar] [CrossRef] [PubMed]
- Tüzel, Y.; Gül, A.; Tüzel, I.H.; Öztekin, G.B. Different soilless culture systems and their management. J. Agric. Food Environ. Sci. 2019, 73, 7–12. [Google Scholar] [CrossRef]
- Karimi, S.; Zare, N. Silicon pretreatment at the transplanting stage, a tool to improve the drought tolerance and subsequent growth of melons in the field. Silicon 2023, 15, 4921–4929. [Google Scholar] [CrossRef]
- Abul-Soud, M.A.; Emam, M.S.A.; Hawash, A.M.H. The potential of soilless culture systems in producing tomato and cucumber under greenhouse conditions. Int. J. Plant Soil Sci. 2021, 33, 67–85. [Google Scholar] [CrossRef]
- Torres Pineda, I.; Lee, Y.D.; Kim, Y.S.; Lee, S.M.; Park, K.S. Review of inventory data in life cycle assessment applied in production of fresh tomato in greenhouse. J. Clean. Prod. 2021, 282, 124395. [Google Scholar] [CrossRef]
- Montesano, F.F.; D’Imperio, M.; Parente, A.; Cardinali, A.; Renna, M.; Serio, F. Green bean biofortification for Si through soilless cultivation: Plant response and Si bioaccessibility in pods. Sci. Rep. 2016, 6, 31662. [Google Scholar] [CrossRef] [PubMed]
- Gomes, F.A.L.; Araújo, R.H.C.R.; Nóbrega, J.S.; de Fátima, R.T.; Santos, M.S.; Santos, A.S.; Teodósio, A.E.M.d.M.; Oliveira, C.J.A. Application of Silicon to Alleviate Irrigation Water Salinity in Melon Growth. J. Exp. Agric. Int. 2018, 25, 1–9. [Google Scholar] [CrossRef]
- George, B.; Kaur, C.; Khurdiya, D.S.; Kapoor, H.C. Antioxidants in tomato (Lycopersium esculentum) as a function of genotype. Food Chem. 2004, 84, 45–51. [Google Scholar] [CrossRef]
- Kaur, H.; Greger, M. A review on si uptake and transport system. Plants 2019, 8, 81. [Google Scholar] [CrossRef]
- Takahashi, E.; Ma, J.F.; Miyake, Y. The possibility of silicon as an essential element for higher plants. Comments Agric. Food Chem. 1990, 2, 99–122. [Google Scholar]
- Sun, H.; Duan, Y.; Mitani-Ueno, N.; Che, J.; Jia, J.; Liu, J.; Guo, J.; Ma, J.F.; Gong, H. Tomato roots have a functional silicon influx transporter but not a functional silicon efflux transporter. Plant Cell Environ. 2020, 43, 732–744. [Google Scholar] [CrossRef] [PubMed]
- Mitani, N.; Yamaji, N.; Ago, Y.; Iwasaki, K.; Ma, J.F. Isolation and functional characterization of an influx silicon transporter in two pumpkin cultivars contrasting in silicon accumulation. Plant J. 2011, 66, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Fei, S.; Xu, Y.; He, Y.; Zhu, Z.; Liu, Y. The structure, function and expression analysis of the nodulin 26-like intrinsic protein subfamily of plant aquaporins in tomato. Sci. Rep. 2022, 12, 9180. [Google Scholar] [CrossRef] [PubMed]
- Nikolic, M.; Nikolic, N.; Liang, Y.; Kirkby, E.A.; Römheld, V. Germanium-68 as an adequate tracer for silicon transport in plants. Characterization of silicon uptake in different crop species. Plant Physiol. 2007, 143, 495–503. [Google Scholar] [CrossRef]
- Liang, Y.; Si, J.; Römheld, V. Silicon uptake and transport is an active process in Cucumis sativus. New Phytol. 2005, 167, 797–804. [Google Scholar] [CrossRef]
- do Nascimento, C.W.A.; de Souza Nunes, G.H.; Preston, H.A.F.; da Silva, F.B.V.; Preston, W.; Loureiro, F.L.C. Influence of Silicon Fertilization on Nutrient Accumulation, Yield and Fruit Quality of Melon Grown in Northeastern Brazil. Silicon 2020, 12, 937–943. [Google Scholar] [CrossRef]
- Raven, J.A. Silicon transport at the cell and tissue level. In Silicon in Agriculture; Datnoff, L.E., Korndorfer, G.H., Eds.; Elsevier Science B.V.: Amsterdam, The Netherlands, 2001; pp. 41–55. [Google Scholar]
- Voogt, W.; Sonneveld, C. Silicon in horticultural crops grown in soilless culture. In Silicon in Agriculture; Datnoff, L.E., Snyder, G.H., Korndorfer, G.H., Eds.; Elsevier Science B.V.: Amsterdam, The Netherlands, 2001; Volume 6, pp. 115–131. [Google Scholar]
- Iler, R.K. The Chemistry of Silica; Wiley-Interscience: Chichester, UK, 1979; ISBN 978-0471024040. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
| Number of Leaves | Leaf FW | Leaf DM | Stem FW | Stem DM | Root FW | Root DM | |
|---|---|---|---|---|---|---|---|
| n. ·plant−1 | g·plant−1 | g·100 g−1 FW | g·plant−1 | g·100 g−1 FW | g·plant−1 | g·100 g−1 FW | |
| Genotypes | |||||||
| Carosello | 4.00 ± 0.17 | 3.98 ± 0.74 | 8.05 ± 0.61 | 0.16 ± 0.01 | 5.25 ± 0.14 | 0.20 ± 0.02 | 5.83 ± 0.31 |
| Tomato ‘Alfa 200’ (TA) | 4.00 ± 0.17 | 2.47 ± 0.19 | 10.78 ± 0.44 | 0.19 ± 0.02 | 6.54 ± 0.49 | 0.23 ± 0.03 | 10.60 ± 0.91 |
| Tomato ‘Versus’ (TV) | 5.00 ± 0.17 | 2.98 ± 0.23 | 10.72 ± 0.45 | 0.20 ± 0.01 | 7.13 ± 0.26 | 0.23 ± 0.04 | 10.53 ± 0.76 |
| Silicon (Si, mg·L−1) | |||||||
| 0 | 4.33 ± 0.17 | 3.03 ± 0.49 | 9.18 ± 0.60 | 0.17 ± 0.02 | 6.00 ± 0.33 | 0.23 ± 0.04 | 7.57 ± 0.65 |
| 50 | 4.33 ± 0.17 | 3.02 ± 0.23 | 9.93 ± 0.45 | 0.20 ± 0.01 | 6.38 ± 0.38 | 0.24 ± 0.03 | 9.17 ± 0.86 |
| 100 | 4.33 ± 0.17 | 3.11 ± 0.22 | 10.44 ± 0.44 | 0.19 ± 0.01 | 6.54 ± 0.53 | 0.25 ± 0.05 | 10.23 ± 1.33 |
| Significance (1) | |||||||
| Tomato vs. Carosello | *** | ns | *** | ns | * | ns | ** |
| TA vs. TV | ns | ** | ns | ns | * | ns | ns |
| Si (linear) | ns | ns | ** | ns | ns | ns | ns |
| Si (quadratic) | ns | ns | ns | ns | ns | ns | ns |
| (Tomato vs. Carosello)*Si (linear) | ns | ns | ns | ns | ns | ns | ns |
| (Tomato vs. Carosello)*Si (quadratic) | ns | ns | ns | ns | ns | ns | ns |
| (TA vs. TV)*Si (linear) | ns | ns | ns | ns | ns | ns | ns |
| (TA vs. TV)*Si (quadratic) | ns | ns | ns | ns | ns | ns | ns |
| Treatment | Silicon |
|---|---|
| mg·kg−1 DW | |
| Plant organ | |
| Stem | 421 ± 93 |
| Leaves | 541 ± 108 |
| Roots | 790 ± 66 |
| Genotypes | |
| Carosello | 908 ± 99 |
| Tomato ‘Alfa 200’ (TA) | 409 ± 80 |
| Tomato ‘Versus’ (TV) | 467 ± 77 |
| Silicon (mg·L−1) | |
| 0 | 299 ± 53 |
| 50 | 635 ± 84 |
| 100 | 838 ± 112 |
| Significance (1) | |
| Shoot vs. roots | *** |
| Leaves vs. stem | ** |
| Tomato vs. Carosello | *** |
| TA vs. TV | *** |
| Si (linear) | *** |
| Si (quadratic) | ns |
| (Shoot vs. roots)*(Tomato vs. Carosello) | ns |
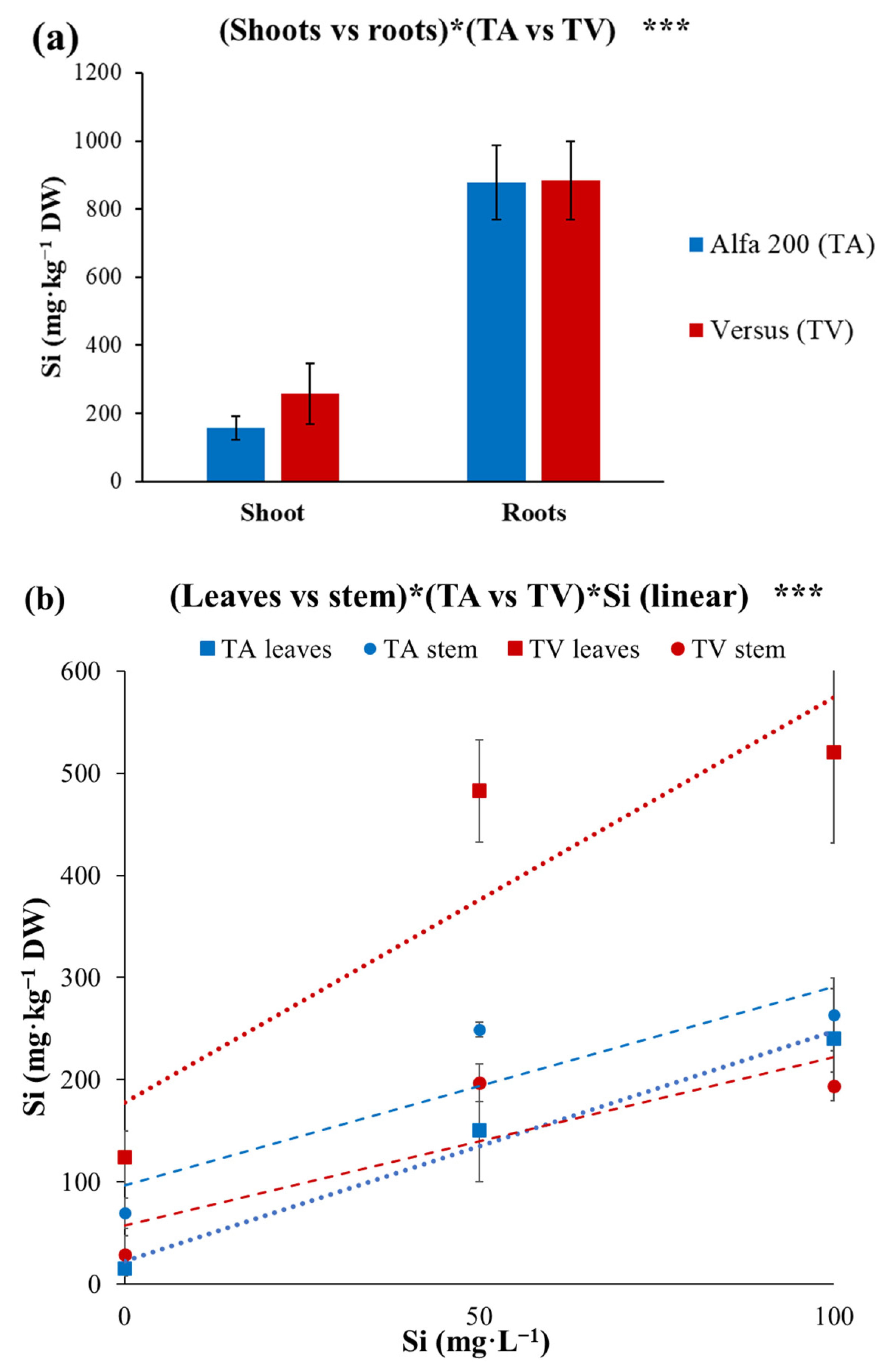
| (Shoot vs. roots)*(TA vs. TV) | *** |
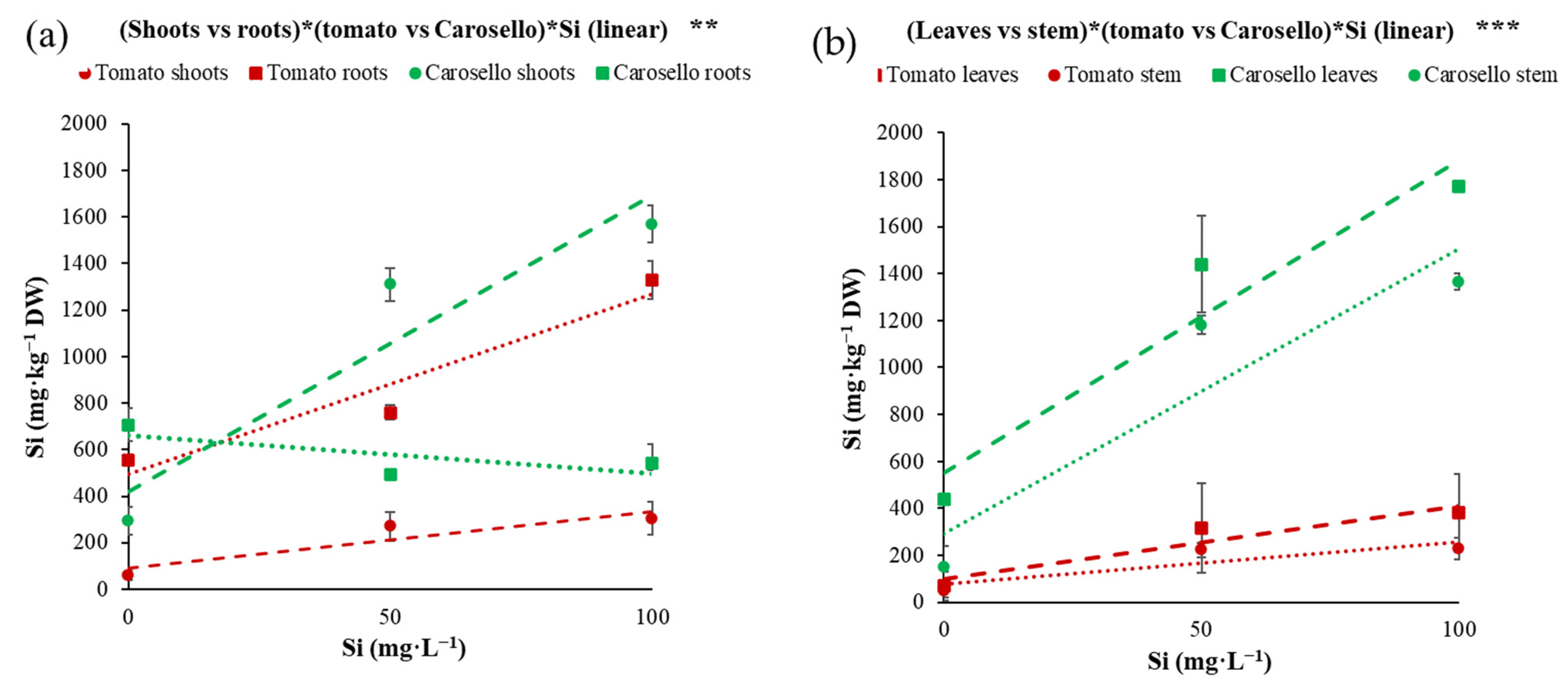
| (Leaf vs. stem)*(Tomato vs. Carosello) | *** |
| (Shoot vs. roots)*Si (linear) | ns |
| (Shoot vs. roots)*Si (quadratic) | ** |
| (Leaves vs. stem)*Si (linear) | ns |
| (Leaves vs. stem)*Si (quadratic) | ns |
| (Tomato vs. Carosello)*Si (linear) | ns |
| (Tomato vs. Carosello)*Si (quadratic) | ns |
| (TA vs. TV)*Si (linear) | *** |
| (TA vs. TV)*Si (quadratic) | *** |
| (Shoot vs. roots)*(Tomato vs. Carosello)*Si (linear) | ** |
| (Shoot vs. roots)*(Tomato vs. Carosello)*Si (quadratic) | ns |
| (Leaves vs. stem)*(Tomato vs. Carosello)*Si (linear) | *** |
| (Leaves vs. stem)*(Tomato vs. Carosello)*Si (quadratic) | ns |
| (Shoot vs. roots)*(TA vs. TV)*Si (linear) | *** |
| (Shoot vs. roots)*(TA vs. TV)*Si (quadratic) | ns |
| (Leaves vs. stem)*(TA vs. TV)*Si (linear) | *** |
| (Leaves vs. stem)*(TA vs. TV)*Si (quadratic) | ns |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Somma, A.; Palmitessa, O.D.; D’Imperio, M.; Serio, F.; Santamaria, P. Genotype-Specific Response to Silicon Supply in Young Tomato and Unripe Melon Plants Grown in a Floating System. Horticulturae 2024, 10, 69. https://doi.org/10.3390/horticulturae10010069
Somma A, Palmitessa OD, D’Imperio M, Serio F, Santamaria P. Genotype-Specific Response to Silicon Supply in Young Tomato and Unripe Melon Plants Grown in a Floating System. Horticulturae. 2024; 10(1):69. https://doi.org/10.3390/horticulturae10010069
Chicago/Turabian StyleSomma, Annalisa, Onofrio Davide Palmitessa, Massimiliano D’Imperio, Francesco Serio, and Pietro Santamaria. 2024. "Genotype-Specific Response to Silicon Supply in Young Tomato and Unripe Melon Plants Grown in a Floating System" Horticulturae 10, no. 1: 69. https://doi.org/10.3390/horticulturae10010069
APA StyleSomma, A., Palmitessa, O. D., D’Imperio, M., Serio, F., & Santamaria, P. (2024). Genotype-Specific Response to Silicon Supply in Young Tomato and Unripe Melon Plants Grown in a Floating System. Horticulturae, 10(1), 69. https://doi.org/10.3390/horticulturae10010069