1. Introduction
Protected cultivation systems such as high tunnels have become increasingly popular due to the opportunity they offer to optimize growing conditions, extend the growing season, and improve crop quality [
1,
2,
3]. Besides the high tunnel structure itself, an additional key element is the covering material. The selection of the covering films can significantly impact the microenvironmental conditions within the high tunnel determining the incidence and severity of pests [
2,
4] and pathogens [
5,
6,
7], crop productivity and produce quality [
8]. Covering films regulate the entry of solar radiation and its component wavelengths, modulate temperature and humidity [
9], and protect crops from pests, pathogens, and adverse weather conditions [
7]. Clear polyethylene films have been extensively used as covering materials. However, advancements have led to the development of alternative covering films with enhanced properties, including varying levels of light diffusion and spectral selective properties which allow the selective transmittance of specific fractions of solar radiation such as ultraviolet (UV) radiation or near-infrared radiation [
10].
Although light diffusive films may slightly reduce the transmittance of the photosynthetically active radiation (PAR), with diffuse radiation the typically shaded vegetation under the canopy can be exposed more uniformly to radiation, because diffused radiation arrives from different directions, reflects within the canopy, and penetrates more deeply into it [
11,
12]. The even distribution of radiation within a plant canopy holds significance because photosynthesis is more effective when most of the leaves receive moderate light levels, thereby improving crop radiation use efficiency [
13,
14]. Shaded leaves, which are typically limited by light availability, exhibit a rapid response to even a slight increase in radiation, thereby enhancing canopy photosynthesis [
15]. Furthermore, diffuse radiation of moderate intensity can increase the canopy’s light-use efficiency compared to direct sunlight [
14]. Nevertheless, the plant’s response to diffused radiation is influenced by various factors related to plant traits, including functional types, leaf physiology, leaf area, leaf orientation, and canopy structure, as well as by specific environmental conditions such as temperature, relative humidity, and vapor pressure deficit [
14]. For these reasons, the effect of diffusive films cannot be assumed and should be evaluated under various conditions.
UV-transparent films permit the majority of UV radiation to reach the plants, potentially influencing their growth, development, and secondary metabolite production. On the other hand, UV-opaque films can limit or block the transmission of UV radiation which may affect not only plant growth and the generation of secondary metabolites but also the activity of insects [
2].
Ultraviolet radiation is an essential component of sunlight and is classified into three subcategories based on the wavelength range, namely UV-A (320–400 nm), UV-B (280–320 nm), and UV-C (below 280 nm) radiation [
16,
17,
18]. Besides PAR with a wavelength range of 400–700 nm, which corresponds to visible light, the radiation relevant for plants includes UV-A, UV-B, and near-infrared radiation (700 nm to 1 mm). Most of the UV radiation reaching the Earth’s surface is UV-A radiation, while nearly all of the UV-C radiation and about 95% of the UV-B radiation is absorbed by the Earth’s stratospheric ozone layer [
19]. Plants specifically have evolved various mechanisms to cope with and utilize UV radiation, as it plays a crucial role in several physiological processes, including photomorphogenesis [
16,
17,
19], and secondary metabolite synthesis [
18]. The effects of UV radiation on plants can be both positive and negative, depending on the energy, intensity, and time of exposure [
17,
20]. Cramer et al. [
2] highlighted that several insect groups are light-sensitive in the UV-A radiation range, and they use this portion of radiation to facilitate their navigation. They found that using plastic covers that blocked more than 90% of the UV-A radiation contributed significantly to decreased Japanese beetle (
Popillia japonica) aggregation and feeding damage [
2]. Other studies have reported that UV-blocking covers are efficient for aphid control [
21] and significantly interrupt the life cycle of fungal pathogens [
7]. Previous studies have shown that using UV-blocking covering films in high tunnels can influence the growth and development of various crops. For instance, studies conducted by Katsoulas et al. [
17] have demonstrated the influence of UV-blocking films on the growth and development of broccoli (
Brassica oleracea var.
italica Plenck), radish (
Raphanus sativus L.), and tomato (
Solanum lycopersicum L.).
Basil (
Ocimum basilicum L.) is a widely cultivated aromatic herb known for its culinary and medicinal uses [
20], making it a valuable crop for high tunnel production. Indeed, basil is a popular and widely grown herb worldwide [
22,
23,
24]. UV radiation has been reported to affect plant growth, leaf morphology, photosynthesis, and antioxidant systems in basil plants [
25]. Similarly, Semenova et al. [
20] investigated the effects of UV radiation on basil plant growth and leaf pigmentation. The controlled transmission of UV radiation can enhance leaf area, shoot, and root biomass, and promote chlorophyll content in basil plants. However, the response may vary depending on the cultivar, UV intensity, and time of exposure.
Basil plants are also known to respond to UV radiation by altering their physiological and metabolic processes. The use of UV-transparent covering films has been found to positively influence the accumulation of secondary metabolites such as phenolics, flavonoids, and anthocyanins in basil [
26], leading to improved flavor, aroma, and pigmentation profiles. Increased levels of certain phenolic compounds, flavonoids, have been reported in basil plants cultivated under UV-transparent films compared to films that block some UV [
20,
27,
28,
29].
High tunnels are a valuable season extension tool to produce fresh basil and the use of alternative covering films characterized by different levels of light diffusion or UV transparency may offer new opportunities for optimizing crop growth and quality. Light-diffusing films may improve radiation use efficiency while UV-transparent films have the potential to enhance growth parameters, induce the synthesis of secondary metabolites, and improve the nutritional profile of basil [
25,
29]. However, further research is needed to determine optimal film properties, and their effects on different basil cultivars. Understanding the mechanisms underlying the response of basil to diffuse light and UV radiation could help to fine-tune high tunnel production systems and improve basil quality.
We hypothesized that different spectra-selective polyethylene covering films (light diffusive or not, UV transparent or UV opaque) would have different effects on the high tunnel microenvironmental conditions (radiation spectrum, temperature), which in turn may affect the plant physiology, yield, and nutritional quality of fresh herbs such as basil.
Thus, this study aimed to evaluate the yield, physiological, and nutritional quality response of purple and green basil to high-tunnel cultivation under alternative covering films characterized by different levels of light diffusion and transparency to UV radiation. Since the response may be modulated by the basil genotype, two varieties of basil with different pigmentation (purple and green) were tested. Moreover, to limit the variability associated with the soil and better isolate the effect of the basil genotype and of the covering films, plants were grown in a soilless system.
2. Materials and Methods
2.1. High Tunnel Research Facilities, Treatments, and Experimental Design
The study was conducted at the High Tunnel Research Facility of the Penn State Russell E. Larson Agricultural Research Center located at Rock Springs, PA (40°71′12.54″ N 77°94′49.58″ W) over the spring-summer of 2019. Twelve research-size (5.18 m × 10.67 m) galvanized metal frame Gothic-style high tunnels (Ledgewood Farms, Moultonboro, NH, USA) were used for the study. High tunnels were oriented about 11 degrees Northeast/Southwest from true North-South. Their fixed wood frame end walls were covered with the same covering materials used for the top and sides of the high tunnels. The tunnels were vented manually with roll-up sides opened to a maximum height of 1.2 m above the baseboard. The high tunnels were covered with four different polyethylene 6-mil films: TuffLite IV® (TIV; Berry Global, Inc., Evansville, IN, USA), KoolLite® Plus (KLP; RKW Hyplast NV, Hoogstraten, Belgium), and two experimental films custom-manufactured that varied in their transparency to UV light, a UV transparent film (UVT) and a UV opaque (UVO) film (BPI-Visqueen, Stevenston, UK, subsequently available through Lightworks Poly, Lancashire, UK). TIV is a clear (non-diffusing) plastic covering marketed as having high levels of transmission of UV, visible, and IR wavelengths, while KLP is marketed as a diffusing “bee-friendly” plastic due to blocking certain wavelengths of UV, mainly in the UV-A range. Both are commonly used by growers in the region. UVT and UVO are diffusing plastics differently from each other in UV transmission. Treatments were arranged according to a split-plot experimental design with three replications. The covering material was randomized in the main plots and two selections of basil (Ocimum basilicum L.), ‘Genovese Compact Improved’ and purple ‘Amethyst Improved’ basil (Johnny Selected Seeds), were randomized as sub-plots.
2.2. Microenvironmental Monitoring and Covering Film Transmittance Properties
Air temperature at a 1.5-m height was measured in each tunnel by WatchDog A125 Temperature/External Temperature dataloggers (Spectrum Technologies, Inc., Aurora, IL, USA) protected by passively vented solar radiation shields. Loggers recorded the ambient air temperature at 10-min intervals for the duration of the experiment.
Solar radiation transmitted into each tunnel was measured within 30 min of solar noon on two clear sky days, 12 July and 24 August, using an Apogee Model PS-300 spectroradiometer equipped with a cosine-corrected detector and leveling fixture (Apogee Instruments, Logan, UT, USA). Measurements were taken in the center of each tunnel at a height of 1 m, immediately preceded and followed by taking a measurement outside of the tunnel in an area clear of obstructions. The average of the two outside measurements was used to calculate the percent transmission of light at each half-wavelength between 300 and 1000 nm into the tunnel through the plastic film covering besides obtaining data on light intensity (photon flux photon density in micromoles/m2/sec) at the same wavelength intervals.
2.3. Plant Growing Conditions and Biometric Assessment
Basil seedlings were seeded on 24 May 2019, in 105-cell nursery trays filled with a peat-perlite growing mix composed of 65–75% Canadian sphagnum peat moss, perlite, dolomite lime (Mix 4, Sun Gro Horticulture, Agawam, MA, USA) and were grown in a polycarbonate greenhouse until transplanting. After germination basil seedlings were fertigated with a 15-3-25 N-P-K water-soluble fertilizer (Peters Professional, Summerville, SC, USA) nutrient solution prepared using a 1 g/L dilution.
On June 18, 2019 basil seedlings were planted in 3.8-L white-on-black plastic grow bags filled with a 2:1 v:v peat:coarse-perlite mix (Pro-Moss Hort Sphagnum Peat Moss, Premier Horticulture LTEE, Rivière-du-Loupe, Quebec, Canada; Industrial Perlite, Thermo-Rock East, New Eagle, PA, USA) with three seedlings per bag. Plants were fertigated with a single dripper per growing bag using a NetafimTM multi-outlet dripper system with 7.6 L/hour drippers split four ways. Once transplanted basil plants were fertigated with a complete nutrient solution prepared using a 20-7-20 N-P-K general purpose soluble fertilizer for alkaline water (Plant Marvel Laboratories, Inc., Chicago Heights, IL, USA) applied at the rate of 0.5 g/L. The nutrient solution contained macro and micro-nutrients at the following concentrations: 100 N (nitrogen), 15 P (phosphorous), 83 K (potassium), 9.5 S, 0.5 Fe (iron) as Fe-EDTA, 0.25 Mn (manganese) as Mn-EDTA, 0.15 B (boron), 0.05 Cu (copper) as Cu-EDTA, 0.05 Mo (molybdenum), and 0.33 Zn (zinc) as Zn-EDTA (µg/L). Calcium (Ca) and magnesium (Mg) were not added to the nutrient solution, because they were already present in adequate quantities in the irrigation water. Six grow bags of each variety were planted as a split of each of two rows (East and West sides) per tunnel for a total of 24 grow bags per tunnel.
2.4. Harvest and Biometric Assessment
Two harvests were conducted, the first at 42 days after planting (DAP) on 30 July 2019, and the second at 78 DAP, on 4 September 2019. Before the two harvests, on 25 July 2019 (between 10:30 and 11:48 a.m. Eastern Daylight Savings Time (EDST)) and 3 September 2019 (between 10:15 a.m. and 12:15 p.m. EDST), basil leaf temperature was measured using an infrared radiometer (MI-220, Apogee Instruments, Logan, UT, USA) with a narrow 18-degree field view. The timing of the leaf temperature measurement was selected considering that the plants had reached good foliar coverage, the sun was high enough to reach the plants through the plastic cover, and the winds were relatively calm to allow stable readings. Basil leaf temperature was measured on the east and west sides of each plant row while the high tunnels roll-up sides were fully open to ensure adequate airflow.
At the first harvest a biometric assessment was conducted measuring leaf area of plants in two grow bags per experimental unit, and plant fresh and dry biomass of stems and leaves separately on plants in four grow bags per experimental unit. Leaf and stem dry biomass were measured after drying a subsample of two plants at 65 °C in a forced-air oven until reaching a constant mass, then samples were weighed to calculate leaves and stem dry matter content. Data collected through the biometric assessment were used to calculate the total plant above-ground dry weight (TPDW = sum of leaf and stem DW), leaf weight fraction (LWF = leaf DW/TPDW), specific leaf area (SLA = LA/leaf DW), and leaf area ratio (LAR = SLA × LWF) as described by Di Gioia et al. [
30]. Dry plant tissue samples were ground and prepared for analysis of the mineral profile. A second subsample of plants in two grow bags per experimental unit was collected in sampling bags and stored in a −80 °C freezer, then freeze-dried and ground for the analysis of the basil chlorophyll and carotenoids content, total phenolics compounds, and antioxidant activity.
At the second harvest, only the total plant fresh biomass was measured on a sample of plants in six grow bags per experimental unit. Given the presence of basil downy mildew (Peronospora belbahrii), a visual assessment was conducted by two of the authors to assess the severity of the foliar disease the day before the second harvest. Plants in six grow bags per experimental unit were rated estimating the percentage of leaf area affected by disease as an indication of severity.
2.5. Basil Mineral Profile and Nutritional Quality Analysis
Ground samples were dried in the oven before being sent to the Penn State Agricultural Analytical Services Laboratory in University Park, PA, for mineral analysis. The analysis consisted of measuring total nitrogen through dry combustion using an Elementar Max Cube in CN mode (Elementar Americas Inc., Ronkonkoma, NY, USA), following the procedure described in Poudel et al. [
31]. In addition, macro-minerals (phosphorus, potassium, calcium, magnesium, sulfur (S), and sodium (Na)) and micro-minerals (manganese, iron, copper, boron, and zinc) were analyzed after acid digestion [
32], using an ICP-OES instrument (Varian 730-ES, Agilent Technologies, Santa Clara, CA, USA).
The total chlorophyll and carotenoid content in the samples were determined as outlined in Poudel et al. [
33]. A freeze-dried ground sample weighing 0.015 g was subjected to extraction with 1.5 mL of 80% acetone for 25 min using a Branson CPX2800H ultrasonic processor (Branson Ultrasonics, Brookfield, CT, USA). After extraction, the mixture was centrifuged at 4000×
g for 5 min. The absorbance of the resulting supernatant was measured at 663 nm (A663), 645 nm (A645), and 470 nm (A470) using a microplate reader (Synergy H1, BioTek, Winooski, VT, USA).
The concentration of total phenolic compounds in the samples was determined using a modified version of the Folin–Ciocalteu method [
34,
35]. For the analysis, the same sample extract used for flavonoid extraction was utilized. A separate microcentrifuge tube was prepared, and the following components were added sequentially: 135 μL of distilled water, 750 μL of Folin–Ciocalteu reagent, 50 μL of the supernatant (obtained from the sample extract), and 600 μL of Na
2CO
3. In the case of the blank sample, 50 μL of 80% acetone was substituted for the supernatant. The mixture was then vortexed for 10
s and incubated in a water bath at 45 °C for 20 min. After cooling the samples to room temperature, the absorbance was measured at 765 nm using a microplate reader (Synergy H1, BioTek, Winooski, VT, USA). A standard curve was prepared using gallic acid, and the total phenol concentration of each sample was expressed as gallic acid equivalent (GAE) per gram of dry weight (mg GAE/g DW).
The 2,2-diphenyl-1-picrylhydrazyl (DPPH) antioxidant assay, originally described by Herald et al. [
36] and Alrifai et al. [
37], was employed with minor modifications to evaluate the overall antioxidant activity. Following the method used for total phenols analysis, the same sample extract utilized for flavonoid extraction was employed as outlined by Poudel et al. [
31]. The initial extraction process was identical to that of total phenols; however, a 1.5-mL portion was stored overnight at −20 °C. On the subsequent day, the samples underwent centrifugation at 12,000×
g for 2 min. Subsequently, 200 μL of the DPPH solution (350 mM in 80% methanol) was mixed with 25 μL of the samples or standard, or 2525 μL of 80% methanol (utilized as a reference to determine the amount of DPPH quenched by the samples or standard). For the blank, 225 μL of 80% methanol was added. The microplate was sealed with parafilm and a lid, and then incubated in the dark at room temperature for 6 h. Finally, the absorbance was measured at 517 nm using a microplate reader (Synergy H1, BioTek, Winooski, VT). Trolox (6-hydroxy-2,5,7,8-tetramethylchroman-2-carboxylic acid) ranging from 25 to 800 μM was employed as the standard for antioxidant activity. The results were expressed as Trolox equivalent antioxidant capacity (TEAC) per gram of dry weight (mM TEAC/g DW).
2.6. Statistical Analysis
Collected data were subject to analysis of variance (ANOVA) as a split-plot experimental design using the GLIMMIX procedure of SAS 9.4 (SAS Institute, Cary, NC, USA) [
38]. When the basil cultivar × plastic film interaction was significant, the SLICEDIFF option was used to compare cultivar means within each plastic, and means for plastics were compared within cultivars. All means were compared using Tukey’s Studentized range test (honestly significant difference) at
p = 0.05. Leaf temperature data were subject to ANOVA as a 2 × 2 × 4 factorial (two sides of each row, two basil genotypes per row, and four plastic treatments) randomized complete block design (RCBD) using PROC GLIMMIX, for which a block was considered a random effect, and row side, cultivar, and plastic cover were considered fixed effects.
3. Results
3.1. Light Spectrum and Transmission Properties of the Covering Films Tested
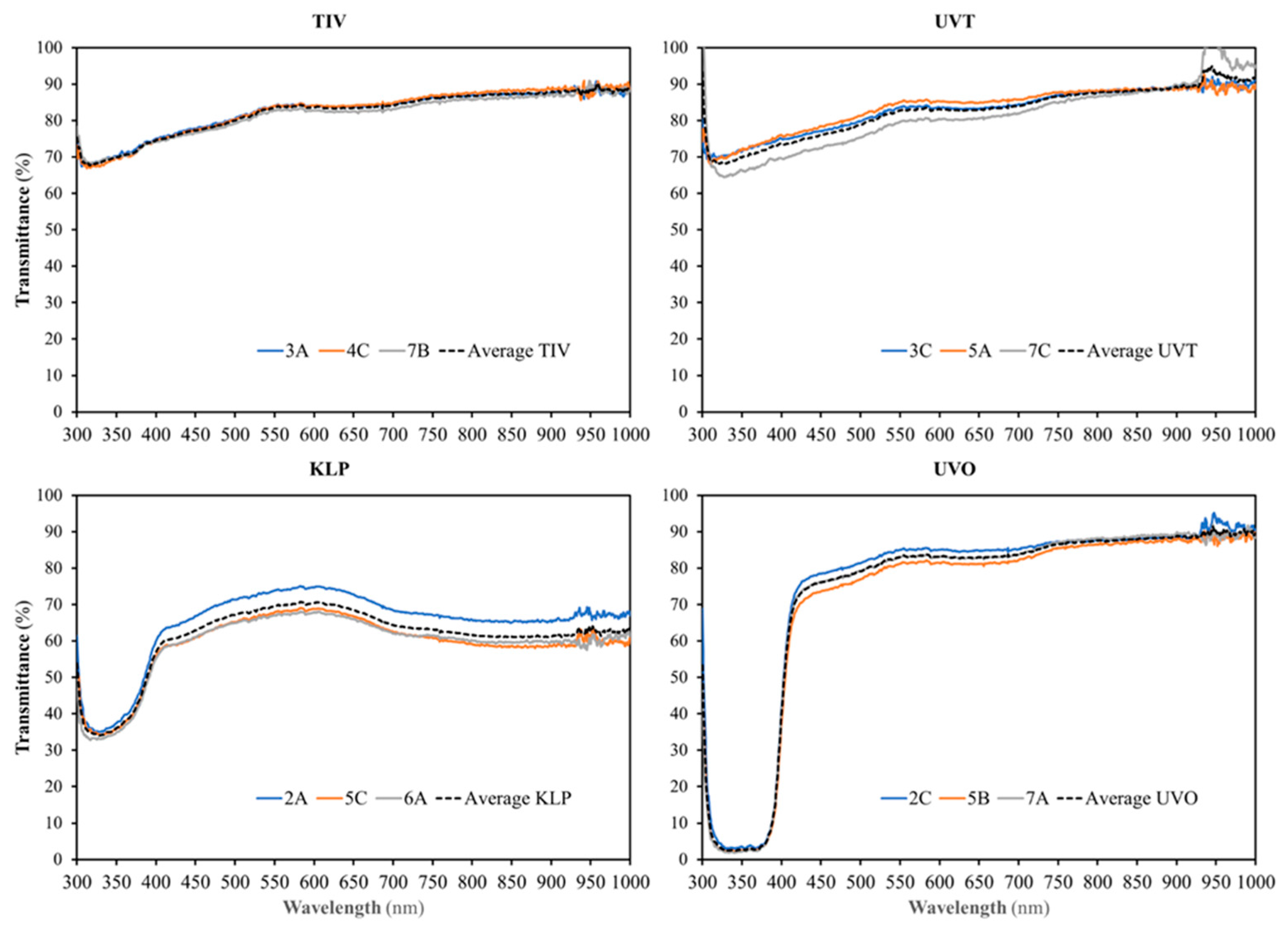
TIV and UVT covers had nearly identical light transmission properties in the UV-A (315–400 nm), visible/PAR (400–700 nm), and far-red (700–780 nm) ranges (
Figure 1 and
Figure 2). However, for UV-B wavelengths between 300 (the shortest wavelength measured) and 306 nm, UVT transmitted 5.12% to 20.85% more light than TIV, with the difference between the plastics increasing as the wavelengths became shorter. UVT also tended to transmit more near-infrared light (NIR, 700–1000 nm) than TIV, with the most apparent difference occurring between 934 and 956 nm, where light intensity was 4.07%–7.02% greater than UVT. UVO and UVT light transmission were nearly identical at wavelengths above 425 nm. However, as indicated by the manufacturer and the film’s designations, differences between the films gradually became more marked below 425 nm with UVO blocking a greater percentage of UV light as wavelengths became shorter.
When comparing UVO and TIV film, light transmittance was nearly identical between 450 and 950 nm, but was slightly greater with UVO above 950 nm, and with TIV in the visible range below 450 nm. KLP reduced light transmission at all wavelengths compared to TIV, with the greatest reductions occurring in the UV-A range, longer wavelengths of the UV-B range, and in the NIR range above 780 nm. KLP also transmitted less light in the NIR range above 780 nm than UVO and UVT. When shown as a percentage of transmitted light relative to incident light from readings taken outdoors (
Figure 2), differences between tunnels covered with the same plastic films were slight, though there was less variability among tunnels covered with TIV than other plastics at that time. Though data were not collected on light diffusion, visible observations indicated that TIV had the greatest clarity and most clearly defined shadows inside of tunnels of all of the films, with UVO and UVT having an intermediate degree of diffusion (haze), and KLP diffusing light the most.
3.2. Effects of Photo-Selective Films on Air Temperature
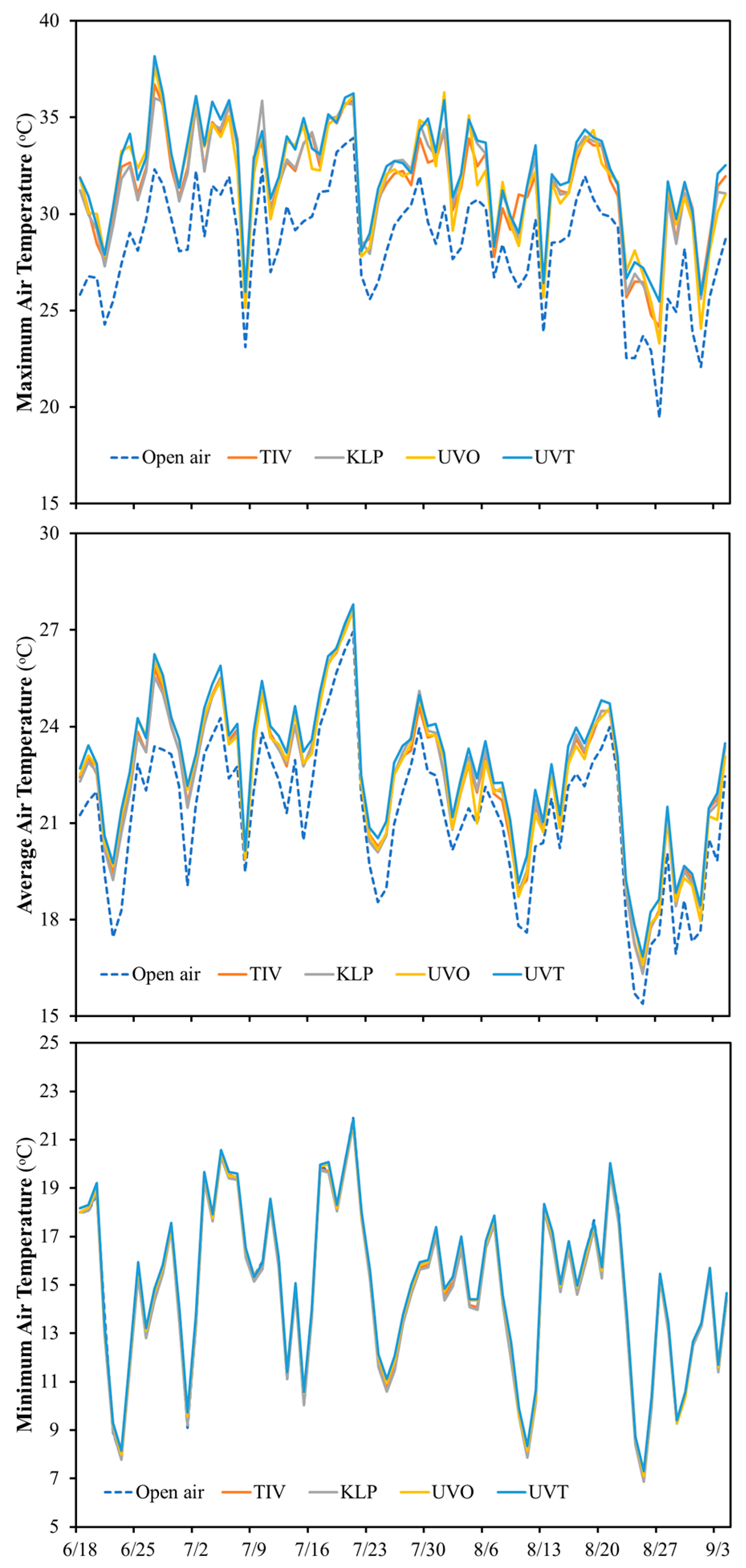
Comparing the effect of the different covering films (TIV, KLP, UVO, UVT) on the high tunnel microenvironment mean, maximum, and minimum air temperatures in relation to open air temperatures, all covering films resulted in slightly higher mean and maximum temperatures compared to the open air (
Figure 3), while limited differences were observed in terms of minimum air temperature. On average, high tunnel mean air temperatures ranged from 22.35 °C (KLP) and 22.70 °C (UVT) and were from 1.12 °C (TIV and KLP) to 1.46 °C (UVT) higher than the mean air temperature in the open air (21.24 °C).
High tunnel maximum temperatures ranged from 36.0 °C (KLP) to 38.17 °C (UVT) and were on average from 2.07 °C (KLP) to 4.23 °C (UVT) higher than the maximum air temperature in the open air (33.93 °C). High tunnels covered with KLP and UVT had the lowest (6.87 °C) and highest (7.30 °C) minimum air temperature and minimal or no differences were observed compared to the minimum air temperature recorded in the open air (7.0 °C).
3.3. Effects on Green and Purple Basil Plant Leaf Temperature, Growth, and Yield
The leaf temperature of green and purple basil measured in correspondence to the two harvest dates was not affected by the four alternative plastic cover films tested (
Table 1). However, it was influenced by the basil genotype and the location of the plant row. Green basil leaves were 3.55% and 4.70% cooler than purple basil leaves before the first and second harvest, respectively. Additionally, the row on the East side of the tunnel exhibited leaf temperatures that were 0.90 and 0.91 °C warmer than the one on the West side before the first and second harvest, respectively.
The four alternative plastic covering films tested influenced the growth of both basil selections and a significant interaction effect was observed between plastic cover and basil genotype for most parameters examined in terms of fresh and dry biomass (
Table 2 and
Table 3). The significant interaction effect observed suggests that while green basil produced higher fresh and dry biomass compared to purple basil, the effect of the alternative plastic covers varied depending on the basil genotype (
Table 2 and
Table 3).
Green basil had the lowest leaf area when grown under TIV plastic covering film and no differences were observed among the other plastic covering films. The purple basil leaf area was higher under the UVO covering film compared to TIV and KLP which had the lowest leaf area. Similar trends were observed in terms of stems, leaves, and total plant fresh biomass.
The green basil variety had a fresh leaf biomass 21.89% higher with the UVO plastic covering film compared to the TIV plastic covering film. However, the KLP and the UVT plastic covering films did not present significant differences for this variable from UVO plastic or each other. Purple basil registered total fresh biomass, respectively, 14.09% and 3.16% higher with the UVO and UVT cover films, compared to the TIV cover film. The green basil leaf fresh biomass was the lowest with the TIV plastic film compared to the other plastic covers, while the purple basil presented the lowest leaf biomass when grown under the KLP plastic film.
Considering the total fresh plant biomass, the green basil had the lowest value with the TIV plastic cover which was 20.68% lower than the UVO cover film. With the purple basil, the lowest values were registered with the KLP and TIV plastic cover and the other plastic covers did not present significant variations for this variable. Thus, green basil had higher fresh biomass compared to purple basil for all plastic covers. Among the plastic covers, UVO and UVT tended to result in higher fresh plant biomass for both green and purple basil.
The leaf dry matter content was higher in green basil compared to purple basil and the covering films had no significant effect on basil dry matter content. However, a significant interaction effect was observed between genotype and covering film for the stem dry matter content. Under TIV the stem dry matter content was similar in green and purple basil, while for all the other treatments green basil had higher stem dry matter compared to purple basil (
Table 2). Examining the treatment effect on the plant dry biomass, green basil had the highest stem dry biomass when grown under UVO plastic film which was 48.68% higher than the TIV plastic film. Purple basil stem dry biomass was the lowest under the KLP plastic cover and was 41.74% lower than the UVO cover film (
Table 3). Compared to the purple basil genotype, the green genotype had higher dry stem biomass under the plastic films UVO, KLP, and UVT. However, green basil grown under the TIV plastic film and purple basil under the KLP plastic film had the lowest leaf and total plant dry biomass. Moreover, green basil had higher leaf and total plant dry biomass than purple basil.
A significant interaction effect was observed for the SLA, LAR, and LWF. The plastic cover had no significant effect on green basil SLA and LAR. However, purple basil grown under the KLP and UVO plastic covers presented the highest values of SLA which were, respectively, 9.43% and 12% higher compared to the TIV cover film. Furthermore, purple basil had higher SLA and LAR compared to green basil, independent of the plastic cover films used. The green basil LWF was 4.41% higher under TIV than under UVO and in purple basil was 9.37% higher under KLP compared to UVO. Thus, with both green and purple basil genotypes, the KLP and TIV plastic cover film presented a higher LWF, while the UVO plastic cover film had the lowest LWF value.
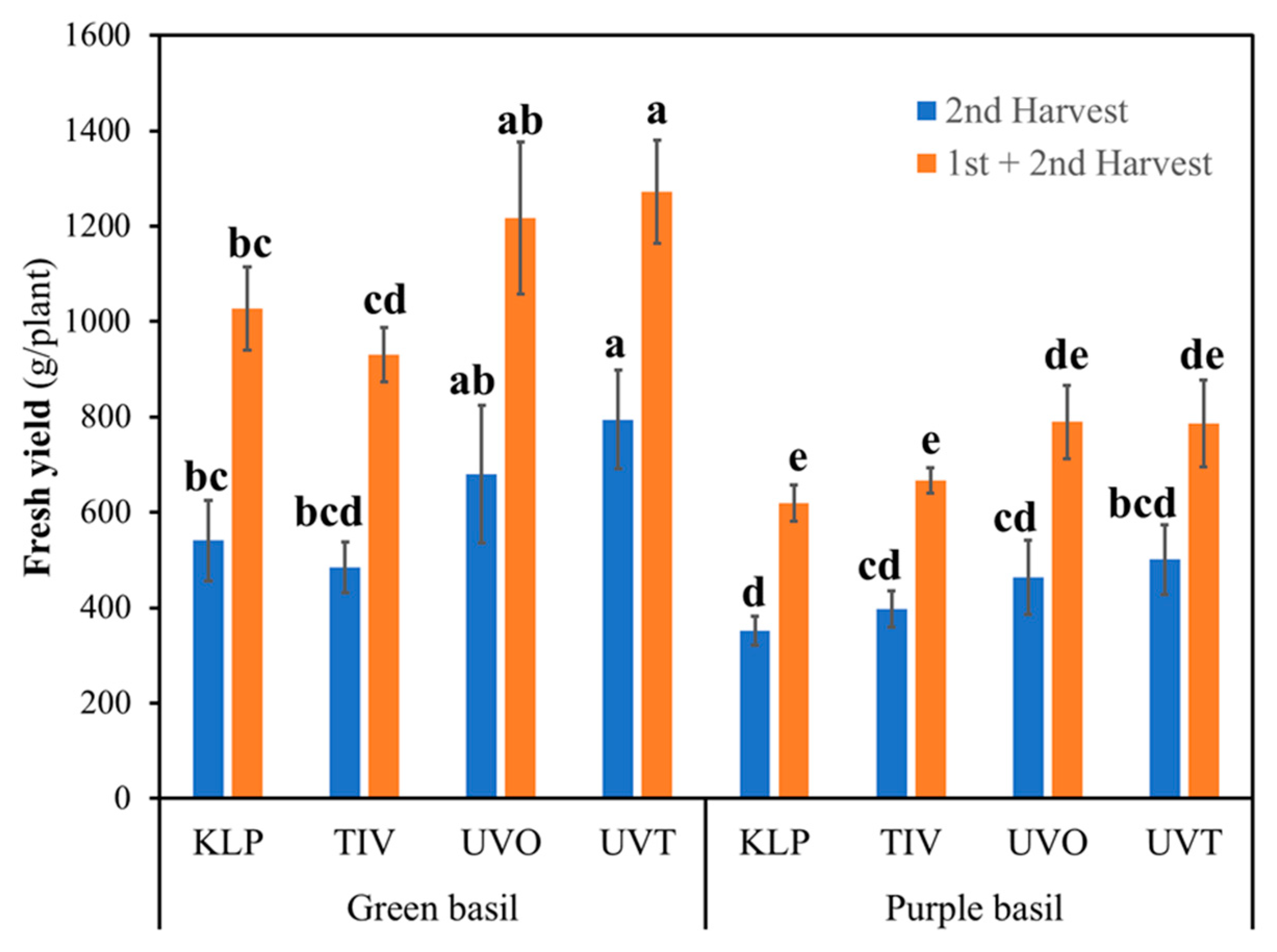
In examining the effects on the second harvest and the total of the first and second harvest fresh yield, a significant film × basil genotype interaction was observed (
Figure 4). Fresh yield for both second and total harvest was generally lower in purple basil and was not affected by the type of covering film used. However, in the case of green basil, the fresh yield was consistently higher in plants grown under UVT and UVO film compared to those grown under TIV. Green basil grown under UVT provided higher total fresh yield even compared to green basil grown under KLP. Green basil provided a higher total fresh yield compared to purple basil, except when grown under TIV film, which provided a total fresh yield comparable to purple basil grown under UVT and UVO.
3.4. Effect of Photo-Selective Films on Basil Downy Mildew Severity
After observing the presence of basil downy mildew (
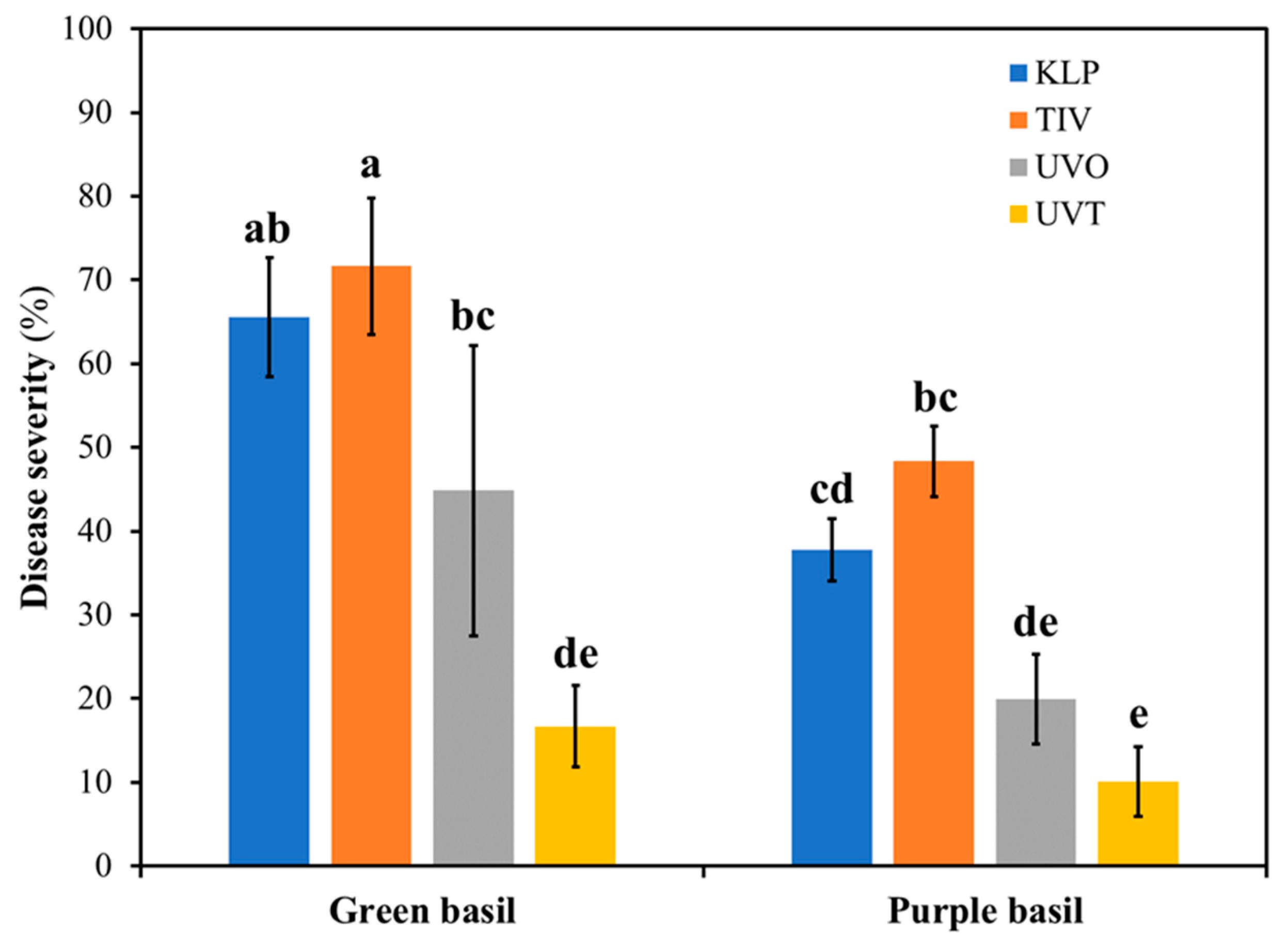
Peronospora belbahrii) in correspondence of the second harvest a visual assessment was conducted to evaluate the severity of the foliar disease as affected by both covering films and basil genotype. The assessment revealed a significant film × basil genotype interaction effect (
Figure 5), suggesting that both basil genotypes and the covering film played a role in determining the extent of downy mildew severity.
Although on average purple basil was less affected by downy mildew compared to green basil, both green and purple basil had the highest and lowest downy mildew severity when grown under the TIV and UVT film, respectively. In both basil genotypes, the second-highest disease severity was observed in plants grown under KLP. However, in green basil plants grown under UVO films the disease severity was higher than in plants grown under UVT. In the case of purple basil, the disease severity in plants grown under UVT and UVO was similar.
3.5. Effects of Photo-Selective Films on Basil Mineral Profile
The photo-selective films tested had limited effects on the mineral concentration of green and purple basil leaves and stems. Instead, the mineral profile of both leaves and stems was influenced by the basil genotype. Nevertheless, a significant film × basil genotype interaction was observed for N, P, K, and S in basil stems, and only for P in basil leaves, indicating that the effect of the films on the concentration of these elements varied depending on the basil genotype (
Table 4 and
Table 5).
For instance, while the covering film had no effect on stem N content in green basil, in the case of purple basil, plants grown under KLP had higher N content compared to those grown under UVT (
Table 6). Similarly, while the film tested had no effect on stem P and K content of green basil, purple basil stem P and K content were higher in plants grown under KLP compared to those grown under TIV, UVT, and TIV, respectively.
Green basil leaves had higher concentrations of N, Ca, Mg, and S compared to purple basil leaves which had higher concentrations of P and K in the leaves (
Table 4). Basil genotype also influenced the leaf concentration of Fe, Zn, and B. Compared to green basil, purple basil leaves had concentrations of Fe, Zn, and B 27.21%, 22.04%, and 2.59% higher, respectively (
Table 5).
3.6. Effects of the Plastic Cover Films on Basil Phytochemical Contents
Both covering films and basil genotype influenced the chlorophyll content, while only genotype influenced total carotenoids, total phenolics, and total antioxidant activity, although a significant film × basil genotype interaction was observed in the case of the total antioxidant activity expressed on a DW basis (
Table 7 and
Table 8).
Compared to the green genotype, purple basil had higher values of chlorophyll (a, b, and a + b) and carotenoid content. When grown under KLP covering films, both basil genotypes exhibited the highest levels of leaf chlorophyll a, b and a + b compared to the other tested covering films, while plants grown under the UVO film had the lowest contents of chlorophyll a, b, and a + b which was, respectively, 8.25, 15.95 and 10.92% lower than the KLP film. No significant difference was observed among the plastic cover films regarding the carotenoid content.
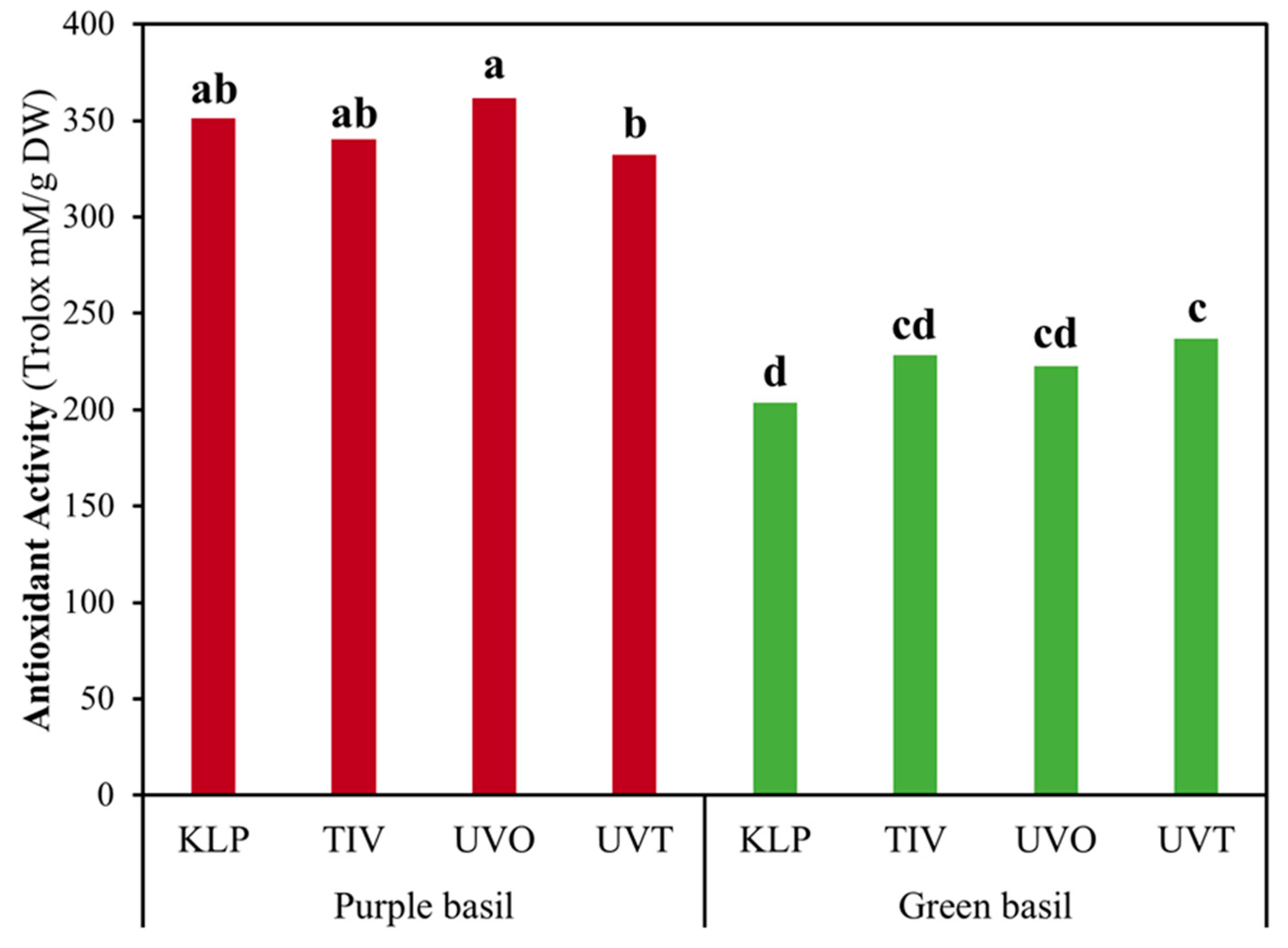
The total phenolics content expressed on a dry weight basis was 15.49% higher in purple basil compared to green basil. However, when considering the total phenolics content on a fresh weight basis, no difference was observed between green and purple basil. Purple basil exhibited values of total antioxidant activity 55.45% and 34.71% higher compared to green basil, on dry and fresh weight, respectively. The different films tested had no effect on total phenolic compounds and total antioxidant activity. However, examining the film × basil genotype interaction it was observed that UVO and UVT determined the highest and lowest total antioxidant activity in purple basil, respectively (
Figure 6). Instead, in green basil, UVT and KLP determined the highest and lowest total antioxidant activity, respectively.
4. Discussion
The plastic cover films had no influence on basil leaf temperature, but this parameter varied with genotype and row side in the high tunnel. However, within any date, the difference between leaf temperature was not greater than 2 °C among both basil genotypes and row sides. In contrast, Cramer et al. [
2] observed notable variations in leaf temperature across different plastic covers used in raspberry cultivation. The temperature difference within a specific date was found to be less than 3 °C, while the variation between different row sides ranged from 2.7 to 5.0 °C within the same date in raspberry plants. This is likely an effect of plant height since raspberry plants are taller in relation to the open sides of the tunnel than basil, which may have experienced a greater influence of outside conditions. Despite the lack of significant differences among plastic film types, there was a tendency for basil plants in tunnels covered with TIV and UVT plastics to have higher leaf temperatures numerically, an effect that has been noted elsewhere and that was ascribed to UV transmittance [
39]. The warmer temperatures of plants located in the row on the East side of the tunnel were likely a warming effect of incident morning sunlight which the row on the West side of the tunnel would not have received. When considering air temperature, the UVT film frequently had the highest maximum and minimum air temperatures observed, perhaps due to the tendency of this film to transmit more NIR light than other films.
The different cover films tested had varying effects on leaf area and fresh biomass as with the findings of Sakalauskaite et al. [
29], while dry matter content was less affected. Additionally, the green basil genotype generally exhibited higher biomass compared to the purple basil genotype, regardless of the plastic cover films tested. This suggests inherent genetic differences between the two genotypes that influence their growth potential and light response since the plastic covers interacted differently with each genotype. The choice of plastic covers plays a crucial role in creating a suitable microenvironment for basil growth. For instance, the UVO and UVT covers generally resulted in higher fresh biomass for both green and purple basil genotypes, while the TIV plastic cover had a negative impact on leaf area and fresh biomass, particularly for green basil.
As mentioned by Meinen et al. [
40], when exposed to increased levels of UV radiation, plants exhibit shorter stature, diminished growth, and lower biomass output, although the specific effects of UV transmittance levels vary among different species. This suggests that certain plastic covers may provide more favorable conditions for basil growth, leading to improved biomass accumulation. Regarding this, Cramer et al. [
2] reported that the UVO plastic cover film can allow the transmittance of 36% and 7% UV-B and UV-A radiation, respectively, compared to 78% and 74% for the TIV plastic cover. Results from this study contradict the findings of Semenova et al. [
20] who reported that basil plants exposure to increased UV registered a dry weight/fresh weight ratio higher by 45 and 35%, respectively, for UV-A and UV-B, and a leaf area higher by 40 and 20%. Given the trends seen in this study, it is also possible that light diffusion played a role with UVO and UVT films which also transmitted a high percentage of PAR. The KLP film also diffused light but transmitted less PAR, which could have affected biomass production as well. These results emphasize the significance of considering multiple factors when aiming to optimize basil growth conditions.
Examining fresh yield, the significant film × basil genotype effect observed on the first, second, and total fresh yield (
Table 2,
Figure 4) suggested that while green basil generally outperformed purple basil, basil yield response was influenced also by the photo-selective films tested. For the green basil plants, both the UVT and UVO plastic covers resulted in the highest fresh yield values during the second harvest as well as the overall total harvest while for the purple basil genotype, there were no significant differences observed in terms of second harvest and total fresh yield among the different plastic cover films. This implies that the choice of plastic cover film did not significantly affect the yield of the purple basil plants. Moreover, Dou et al. [
28] reported an inhibition of plant growth in green (‘Improved Genovese Compact’) and red (‘Red Rubin’) basil cultivars performing as lower plant height, width, and leaf area resulted from exposure to increasing UV-B radiation. Additionally, comprehensive investigations were conducted to assess the impact of UV-blocking covering materials on plant yield and most of them reported a positive effect across various plant species [
17].
Overall, this study emphasizes the importance of considering both the choice of plastic cover film and the specific basil genotypes when managing downy mildew in high tunnel production. The TIV and KLP plastic cover films were associated with higher severity, while the UVT film appeared to provide some level of protection. Thus, the UVT and UVO displaying, respectively, the greatest and least UV-B and UV-A radiation [
2], provided better protection against downy mildew in both basil selections. Additionally, Patel et al. [
41] argued that prioritizing increased light exposure to the lower leaf surface held greater significance compared to illuminating the upper leaf surface. This emphasis on diffusion might play a crucial role. It is conceivable that this effect could be more pronounced under a plastic material that not only allowed more light transmission but also diffused it, such as UVO and UVT. This is where the lower downy mildew ratings were observed. These results are consistent with the findings of Raviv and Antignus [
7] who documented that numerous plant responses to UV radiation are considered defense mechanisms that can potentially hinder fungal growth. Evidently, the green and purple basil genotypes showed varying susceptibility to downy mildew. This suggests that there may be a genetic difference between the green and purple basil genotypes in terms of their susceptibility to downy mildew [
42]. The potential resistance to basil downy mildew was first reported by Wyenandt et al. [
43] showing that the development of resistant cultivars may be possible. Pyne et al. [
44] suggest that downy mildew resistance in basil is under major gene control. Regarding this, the green genotype, in general, appeared to be more susceptible to downy mildew compared to the purple genotype in the current study.
This study highlighted that basil genotypes can exhibit differences in nutrient concentrations, both in the leaves and stems. The photo-selective cover films tested had a lower impact on nutrient concentration compared to the basil genotype; however, the film × basil genotype interaction effect observed on N, P, K, and S in basil stems, and only for P in basil leaves, suggest that both factors played a role in determining the nutrient concentration in basil leaves and stems. The higher concentration of N, P, and K observed in the stems of purple basil plants grown under KLP compared to those grown under UVT, and UVT and TIV, respectively, could be explained by the lower levels of visible solar radiation transmitted by KLP films (
Figure 1 and
Figure 2). These results are consistent with the findings of Walters and Currey [
45] who reported that the concentrations of N, K, Ca, Zn, Mn, Cu, Fe, and B in sweet basil tissue exhibited a decrease with higher daily light levels. However, in the present study minimal or no covering film effects were observed on the macro-mineral profile of green basil (
Table 4), which suggests that like for other factors, the effects of photo-selective films may be genotype-specific in the case of basil [
23,
46].
While minimal effects were observed on the basil mineral profile, the results of this study highlight the influence of plastic cover films on chlorophyll content and the interaction effect between plastic cover and basil genotype on total antioxidant activity. The chlorophyll content was affected by both plastic cover films and basil genotypes. Both genotypes exhibited higher chlorophyll content under the KLP cover film which allows the transmission of medium amounts of UV-B and UV-A and reduced PAR levels (
Figure 1 and
Figure 2), compared to other films tested. However, considering that a lower chlorophyll content was observed under UVO films, which significantly reduced the transmittance of UV radiation, it is likely that the increase in chlorophyll content observed under the KLP film was primarily associated with the reduced PAR levels.
These results are consistent with the findings of Stagnari et al. [
47] who observed an increase in chlorophyll content in basil plants exposed to lower radiation levels due to shading. Conversely, these results are not consistent with the findings of Semenova et al. [
20] who reported that exposure to high UV irradiation positively impacts the chlorophyll content in basil cultivation which could be due to the activation of mechanisms to protect against UV irradiation. Regarding carotenoid content, no significant difference was observed among the plastic cover films. This implies that the choice of plastic cover film did not have a noticeable impact on the accumulation of carotenoids in the basil plants.
The results showed that the carotenoid content was determined genetically in basil cultivation with the purple genotype displaying a higher carotenoid content. The carotenoid content levels observed in this study for green basil are consistent with the values reported by Naznin et al. [
48] in green lemon basil and are lower compared to those reported by Proz et al. [
49] or both green and purple basil grown indoors, in a greenhouse or in open field. Moreover, in contrast with our results, Proz et al. [
49] reported higher total carotenoid content in green basil than in purple basil.
Total phenolic content was not influenced by the photo-selective films tested and was influenced by the basil genotype only on a dry weight basis. Purple basil exhibited higher total phenolic content values only when assessed on a dry weight basis compared to the green genotype. The total phenolic content levels and tendency observed in this study corroborated Blunk et al. [
50], Cruz et al. [
51], and Prinsi et al. [
52] who reported a higher value for this parameter in purple (‘Dark Opal’) basil compared to green (‘Italian Genovese’, ‘Italiano Classico’, and ‘Mitikas’) varieties. While this study did not find any significant differences among the plastic cover films concerning total phenolic compounds, Dou et al. [
28] reported that the total phenolic compounds in basil cultivation increase significantly with an increase in daily light.
Consistent with the content of total phenolic compounds, the total antioxidant activity was not influenced by the covering films tested and was higher in purple basil compared to green basil, both on a fresh and dry weight basis. Nevertheless, the significant film × basil genotype interaction effect observed revealed once more that the effect of the photo-selective films tested was modulated by the basil genotype. The higher antioxidant activity observed in green basil grown under UVT film is consistent with the reports of previous studies and the hypothesis that higher exposure to UV radiation could serve as eustress and stimulate the plant to generate more antioxidant compounds [
29,
53] compared to plants exposed to lower UV and PAR levels as in the case of plants grown under KLP film. Instead, the effects observed on purple basil contrast with such a hypothesis considering that the highest level of antioxidant activity was observed in plants grown under the UVO film, and the lowest levels were observed in plants grown under the UVT film.



 ,
, 



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}