Abstract
The aim of this study was the evaluation of the effect of the sequential inoculation of Metschnikowia pulcherrima and Saccharomyces cerevisiae for the production of Pecorino wine on the winery scale. Sequential inoculation allowed for wines with interesting oenological characteristics to be obtained. In particular, these wines showed a lower ethanol content than those obtained with S. cerevisiae alone, and the presence of M. pulcherrima induced a reduction in the wine titratable and volatile acidity of about 30%. The effect of M. pulcherrima was relevant also in terms of the volatile profile of the wines. In fact, an increase in the esters content of 60% and a reduction of 53% of the acetic acid was observed. Moreover, the sequential inoculation resulted in an increase in the terpenes content of about 80%. The wines also showed a different sensory profile; in fact, the wines obtained with the sequential inoculation showed higher scores than the other wines for the following descriptors: floral, tropical fruit, citrusy, balsamic, and freshness.
1. Introduction
The process of wine fermentation encompasses a multifaceted assemblage of microorganisms; indeed, there exists a wide range of genera, species, and strains, which are interactive and thus produce a fluctuating array of metabolites according to changes in dominance [1]. These microorganisms include yeasts, bacteria, and filamentous fungi [2]. The process of grape transformation into wine is heavily influenced by the dynamic evolution of the microbial community and metabolic interactions. This transformation primarily involves the conversion of sugars into ethanol and other metabolites [3].
The first steps of fermentation are characterized by the dominance of apiculate yeasts and other non-Saccharomyces, e.g., the Metschnikowia, Pichia, Candida, Lachancea, and Torulaspora genera, followed by the prevalence of Saccharomyces cerevisiae [4]. Due to the challenges associated with managing spontaneous fermentation and the inherent unpredictability and difficulty in reproducing the oenological characteristics of the resulting wines, the prevalent approach in the global wine industry has shifted towards the practice of inoculated fermentation using specific strains of S. cerevisiae [5]. This approach facilitates the production of wines that exhibit the intended oenological characteristics. Nevertheless, it has been widely acknowledged that this particular method leads to the production of wines that lack the unique attributes and intricacy typically associated with the presence of non-Saccharomyces strains during the fermentation process [6,7]. The use of sequential inoculation or co-inoculum with non-Saccharomyces has been described as a suitable microbiological alternative to mimic spontaneous fermentation [8]. Non-Saccharomyces yeast can potentially enhance wine flavor and aroma, counteract spoilage microbes, and reduce ethanol content. Moreover, they were discovered to affect glycerol and mannoprotein amounts, volatile acidity, and/or color stability [9].
Among the different non-Saccharomyces wine yeasts used in sequential inoculation with S. cerevisiae in winemaking, an increasing interest is gaining in Metschnikowia pulcherrima. M. pulcherrima (anamorph form called Candida pulcherrima) is a ubiquitous yeast that has been found in grapes, fruits (fresh and spoiled), flowers, nectars, and tree sap fluxes. Several studies revealed that it has the ability to regulate the production of secondary metabolites in order to enhance the sensory characteristics of wine and serve as a biocontrol agent [10,11,12,13]. An additional noteworthy characteristic of this non-Saccharomyces yeast is its extensive range of enzymatic activities found among the various strains. These activities include pectinase, protease, glucanase, lichenase, β-glucosidase, cellulase, xylanase, amylase, sulfite reductase, lipase, and β-lyase activity [10,14]. In the context of sequential inoculation, it is observed that S. cerevisiae and non-Saccharomyces yeast do not simply coexist but rather exhibit dynamic interactions that have the potential to impact the analytical characteristics of the resulting wine [15]. The proteolytic activity of M. pulcherrima is a significant characteristic during the fermentation processes, as it facilitates the release of amino acids that serve as nutrients for S. cerevisiae. Additionally, this activity acts as a means of controlling the formation of protein haze in wines, functioning as a biological fining agent [16]. According to multiple studies, the utilization of M. pulcherrima has been observed to exhibit a consistent decrease in volatile acidity, with estimated variations ranging from 10% to 75% [17]. Metschnikowia pulcherrima has the potential to serve as a biocontrol agent, as it produces pulcherrimin, a red pigment that exhibits antifungal properties [18].
On the basis of its characteristics, M. pulcherrima seems to be a good candidate to be used in winemaking. Therefore, in this study, the impact of S. cerevisiae and M. pulcherrima on the main oenological parameters, the copper content, and the aroma and sensory profiles of Pecorino wines were evaluated at the industrial level. Pecorino grapes are cultivated in central Italy, mainly in the Abruzzo and Marche regions. Its cultivation is documented starting from the second half of the 19th century, and it is considered an ancient variety originating from the Sibillini mountains, imported into Italy during the Greek migrations. The attribution of this name to the vine is supported by some hypotheses. It seems that the grapes, being on the route of the ancient sheep transhumance (seasonal movement of livestock) carried out from the Abruzzo mountains to the Apulian lands, were particularly appreciated by the sheep that found them well-ripe in the late summer period. A second hypothesis of the etymology is that of the strong resemblance between the winged bunch of Pecorino and the head of the sheep. For several years, this variety was completely abandoned since wineries preferred high-yield grapes. It was accidentally rediscovered in an old vineyard and has been reclaimed as a signature white grape of the Abruzzo region. After its introduction to the national register of vines, the typical geographical indication “Colline Pescaresi” for Pecorino wine was subsequently approved [19].
2. Materials and Methods
2.1. Origin of the Strains
The strain Metschnikowia pulcherrima (GS80) and the strain S. cerevisiae (SRS1) were obtained from organic grapes and musts and were previously identified based on the sequence analysis of 26S rRNA gene [5]. The strains were stored at the Culture Collection of the Microbial Biotechnology Laboratory (Department of BioScience and Technology for Food, Agriculture, and Environment, University of Teramo in Italy). Yeast peptone dextrose (YPD) medium, consisting of 1% (w/v) yeast extract, 2% (w/v) peptone, and 2% (w/v) glucose, was employed for the cultivation of yeast cells. The cells were subjected to incubation at a temperature of 28 °C for 48 h. The strains were maintained at a temperature of −80 °C in YPD broth, which was supplemented with glycerol (20% v/v, Sigma-Aldrich, Milan, Italy).
2.2. Cellar Vinification
Vinifications were carried out in a cellar located in Tollo in Abruzzo region (Italy) during the harvest of 2022. Pecorino must (213.83 g/L fermentable sugars, 6.1 g/ L tartaric acid titratable acidity (TTA), and pH 3.45) was collected in tanks of 100 hL after destemming and crushing and treated following the standard winemaking procedure. Each yeast strain (M. pulcherrima and S. cerevisiae) was inoculated at 106 cells/mL from 24 h pre-cultures grown in the same pasteurized must. S. cerevisiae was inoculated after 48 h following the scheme reported in Table 1. All fermentations were carried out in triplicate at room temperature (maximum temperature variation from 9 to 19 °C). Sulfur dioxide (SO2) was not added. After the completion of fermentation, the yeast sediment was allowed to settle for 7 days. Subsequently, the wines were transferred to a different vessel, a process known as racking, and stored for 14 days. The wines were subsequently transferred into glass bottles with a capacity of 750 mL. These bottles were sealed with crown caps and kept in storage at a temperature range of 15–20 °C for a maximum duration of 30 days. Following this storage period, the wines were subjected to sensory analyses.

Table 1.
Fermentation trials carried out in this study.
2.3. Determination of Oenological Parameters
The FOSS WineScan™ FT120 rapid scanning Fourier Transform Infrared Spectroscopy was employed to monitor ethanol, residual sugars, titratable acidity, and volatile acidity in accordance with the instructions provided by the manufacturer. The calibration was conducted by utilizing wine samples that were subjected to analysis using the established OIV methods [20]. The pH measurement was conducted utilizing an InoLab 730 pH meter (WTW, Weilheim, Germany). The content of copper was determined with EnzytecColor COPPER (R-Biopharm, Darnstadt, Germany) on a Gallery Discrete Analyzer (ThermoFisher, Waltham, MA, USA) as previously described [21].
2.4. Viable Yeast Count
The samples underwent serial dilution using a sterile peptone water solution (0.1% w/v). Subsequently, aliquots of the diluted samples were plated to determine the number of colony-forming units per milliliter (CFU/mL). Lysine agar medium (Oxoid, Hampshire, UK) was utilized for non-Saccharomyces strains, while WL nutrient agar medium (Oxoid, Hampshire, UK) was employed for the total yeast count. Plates were incubated at 28 °C for 5 days. In particular, the analyses were performed at the following sampling points: T0 (after the inoculation of S. cerevisiae in F1 trial, and after the inoculation of M. pulcherrima in F2 trial), after 2 days (after the inoculation of S. cerevisiae in F2 trial), and after 5, 7, and 14 days of fermentation. All analyses were performed in triplicate.
2.5. Determination of Volatile Organic Compounds
The measurement of volatile organic compounds (VOCs) was conducted using solid-phase microextraction coupled with gas-chromatography (SPME-GC-MS) with the utilization of a Clarus SQ8S chromatography/mass spectrometer (GC-MS) (Perkin Elmer, Boston, MA, USA) [22]. The column employed had the following dimensions: 30 m × 0.25 mm i.d. 0.25 μm film thickness, and was coated with polyethyleneglycol, serving as the stationary phase, with a film thickness of 1.2 μm. The carboxen–polydimethylsiloxane-coated fiber (85 μm) (Sigma-Aldrich) was introduced into the desorption chamber and allowed to remain for a duration of 15 min. Subsequently, a program consisting of specific temperature settings was implemented. Initially, the temperature was set at 50 °C for 2 min. Following this, the temperature was gradually increased at a rate of 1 °C per minute until reaching 65 °C. Subsequently, a second ramp was initiated, elevating the temperature at a rate of 10 °C per minute until reaching 150 °C where it was maintained for 10 min. Finally, a third ramp was executed, increasing the temperature at a rate of 10 °C per minute until reaching 200 °C where it was held for 1 min. The pure standards (Sigma-Aldrich) utilized for the identification of aroma compounds included ethyl acetate, ethyl butanoate, ethyl propanoate, ethyl isobutanoate, ethyl isovalerate, ethyl hexanoate, ethyl lactate, ethyl octanoate, ethyl decanoate, iso-amyl acetate, ethyl 2-phenyl acetate, 1-propanol, 1-hexanol, 2-phenylethanol, hexanoic acid, octanoic acid, decanoic acid, and dodecanoic acid. The internal standard used was 2-methyl-hexanol at a concentration of 0.1 M. The identification was additionally validated through the comparison of mass spectra and MS fragmentation patterns with those found in the National Institute for Standards and Technology database (NIST version 2005). Compounds lacking pure reference standards were provisionally identified solely through the comparison of mass spectra. All measurements were conducted in triplicate.
2.6. Sensorial Analysis
A quantitative descriptive analysis (QDA) was conducted on the wines by a group of 15 trained sensory panelists, consisting of 7 women and 8 men, following the methodology outlined by Perpetuini et al. [23]. The assessors were provided with information regarding the analysis in accordance with the ISO 8586-2012 regulation [24]. The judges underwent specialized training in order to assess the following descriptors: floral, tropical fruit, citrusy, herbal, balsamic, persistence, structure, mineral, freshness, and balance. Furthermore, the judges underwent training in order to familiarize themselves with the intensity scale employed, which ranged from 0 to 10. The individual denoted “zero (0)” if the descriptor was not perceived, whereas a ten (10) represented the utmost level of perception. Sensory evaluations were conducted on the samples under controlled conditions at a temperature of 12 °C. Then, 30-milliliter samples were transferred into glasses with a capacity of 150 milliliters. The analysis was conducted in a controlled environment known as a sensory room where participants were placed in separate booths illuminated by white lights [25].
2.7. Statistical Analysis
Prism 7.0 program (GraphPad Software Inc., La Jolla, CA, USA) was used to analyze data and prepare graphs. Results were expressed as mean value ± standard deviation. t-test was used to determine significance (p < 0.05) concerning the oenological parameters and volatile compounds.
3. Results
3.1. Yeast Dynamics
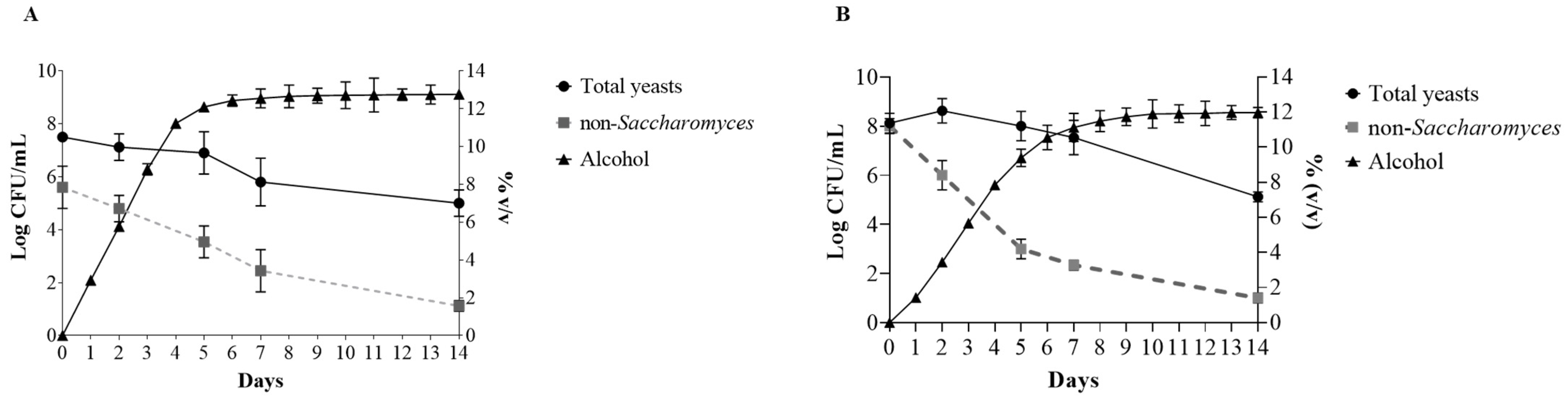
The yeast growth dynamics are shown in Figure 1. In the F1 trial, the number of total yeasts remained stable for the first days and then started to decrease, and a value of 5 log CFU/mL was detected at the end of the fermentation. The non-Saccharomyces yeasts were 5.6 log CFU/mL at the beginning, and a decrease of about a log was observed after 2 days, and a cell count of 3.54 log CFU/mL was detected after 5 days. At the end of the alcoholic fermentation, they almost disappeared, probably due to nutrient depletion or/and the presence of significant levels of alcohol. In the F2 trial, the total yeast count was 8.12 Log CFU/mL at the beginning, and reached a value of 8.62 Log CFU/mL after 2 days, when S. cerevisiae was inoculated. The total yeasts started to decrease after 5 days, and a value of 5.12 Log CFU/mL was detected at the end of fermentation.
Figure 1.
Yeast growth dynamics (log CFU/mL) and alcohol content (% v/v) in F1 (A) and F2 (B) trials.
The number of non-Saccharomyces yeast was higher in F2 than in F1 because of the inoculum of M. pulcherrima. However, a faster decrease in their concentration was observed. This phenomenon could be related to M. pulcherrima biocontrol activity. Multiple studies have demonstrated that this particular yeast exhibits the ability to control the growth of various genera, including Hanseniaspora, Pichia, Torulaspora, Zygosaccharomyces, Saccharomycodes, Candida, Issatchenkia, Brettanomyces, and Schizosaccharomyces [18,26]. The biocontrol mechanism employed for M. pulcherrima involves the depletion of iron from the medium by binding it to pulcherrimic acid [18]. Based on the aforementioned attitudes, it can be inferred that the utilization of M. pulcherrima species may serve as a viable approach in mitigating the proliferation of spoilage microorganisms and minimizing the reliance on sulfur dioxide (SO2), especially within the context of organic and biodynamic wine production. A recent survey examining consumer perceptions, preferences, and willingness to pay for wine, revealed that wines devoid of added sulfites were linked to health advantages and were perceived distinctively from other categories of sustainable wines [27]. This finding suggests that the use of a multi-starter culture M. pulcherrima + S. cerevisiae could represent a valuable approach to producing more sustainable Pecorino wines.
Concerning the fermentation kinetic, the F1 trial showed a faster kinetic with an alcohol content of 5.77% after 2 days; in F2, the content was 3.66% at the same time, underlining the effect of S. cerevisiae inoculation. This difference remained until the end of fermentation; in fact, after 14 days, the F1 wines had about 1% more alcohol. These results highlighted that both fermentations were driven by S. cerevisiae. Indeed, the longer the time when M. pulcherrima acted in the absence of S. cerevisiae, the lower the fermentation kinetics. This evidence was supported by the lower fermentation kinetics of the trials inoculated with S. cerevisiae after 48 h (F2 trial) compared to the S. cerevisiae inoculation at 24 h and by the highest fermentation kinetics when S. cerevisiae was in pure culture (F1 trial).
3.2. Oenological Parameters
The assessment of the influence on the chemical composition of wine resulting from the interaction between S. cerevisiae and M. pulcherrima was conducted quantifying the main analytical components of the wines at the end of fermentation (Table 2). Both wines exhibited a minimal residual sugar concentration (less than 0.5 g/L), thereby confirming the successful completion of the yeast-driven alcoholic fermentation process within a span of 15 days. The sequential inoculation resulted in a decrease in ethanol levels, in agreement with previous studies which also observed a decrease in the ethanol content during fermentations carried out using a sequential inoculation approach involving M. pulcherrima and S. cerevisiae strains [28,29,30]. This phenomenon has been linked to the respiratory metabolism of M. pulcherrima strains. When provided with adequate aeration conditions, these strains can respire 40 to 100% of the sugar they consume, resulting in a decrease in the final ethanol yields [30]. The search for new tools to reduce ethanol concentration represents one of the main challenges of the wine industry posed by climate change, which is causing an increase in grape ripeness levels, resulting in more sugar and alcohol in wines [31].
Table 2.
Main oenological parameters. Different letters in the same line indicate significant differences (p < 0.05).
No significant differences were observed in the final pH of the wines. However, the sequential inoculation induced a reduction in the wine titratable and volatile acidity. Some authors have also documented that M. pulcherrima has the ability to reduce the titratable acidity of the resulting wines [32,33]. Moreover, it has been observed that the utilization of M. pulcherrima results in a range of a 10% to 75% reduction in volatile acidity [17,34,35]. This suggests that the reduction in volatile acidity in such conditions seems to be a trend. However, other studies have reported equal quantities of acetic acid when sequential fermentation was employed [13,36], or even an increase of around 20% [37].
3.3. Effect of M. pulcherrima on Copper Reduction
Copper is an essential metallic element for biological growth and metabolism, as well as a cofactor for intracellular enzyme metabolism, particularly within a low concentration range [38]. However, when exceeding the optimal range, it can have an inhibitory impact on cells, potentially leading to toxicity [39]. The presence of elevated levels of copper in the process of wine production has been observed to have an impact on both the fermentation process and the overall quality of the resulting wine [40]. Therefore, the selection of resistant strains able to absorb copper is crucial, as there is a risk of high levels of copper residues in grape must [40]. In both samples, the content of copper was below the legal limit. The wines obtained with a pure culture of S. cerevisiae showed a content of 0.44 mg/L, while the sequential inoculation induced a reduction of 29.55% (Table 2). Few studies have been performed on the mechanisms of copper uptake in non-Saccharomyces yeasts, and their effect in co-inoculation or sequential inoculation with S. cerevisiae. The considerable variation in copper tolerance among various strains of wine yeast is widely recognized. Both non-living and living wine yeasts have demonstrated the ability to uptake copper, although the specific mechanism of this action remains a topic of ongoing discussion. A recent study has shown an inverse correlation between the ability to tolerate copper and the capacity to adsorb copper, in various strains. The strain that exhibited the highest removal ratio and the most efficient adsorption was found to be the most sensitive [40]. In addition, it has been observed that the primary method by which copper is eliminated by S. cerevisiae during the process of alcoholic fermentation is through the process of cell surface adsorption [40].
3.4. Aroma Profile
A total of 56 aroma compounds belonging to different chemical classes (organic acids, esters, higher alcohols, aldehydes, terpenes, and lactones) were detected. The differences detected were more quantitative than qualitative. Significant differences were obtained for almost all chemical classes, with the only exceptions of lactones and aldehydes that were present in similar amounts in both wines. The total concentration of organic acids was higher in wines obtained with S. cerevisiae (18.12 mg/L) alone than in the others (16.08 mg/L) (Table 3). In particular, the F2 wines showed a reduction of 53% in acetic acid. Different studies reported low acetic acid production for certain non-Saccharomyces yeasts (M. pulcherrima, Torulaspora delbrueckii, Starmerella bacillaris), and their capacity, in culture with S. cerevisiae, to produce lower acetic acid concentrations in comparison to a S. cerevisiae monoculture [41,42]. According to Sadoudi et al. [43], it seems that the acetic acid metabolic pathway of S. cerevisiae can be affected by the presence of M. pulcherrima, leading to a decrease in the amount of acetic acid.
Table 3.
Main volatile compounds detected in wines at the end of fermentation, expressed as mg/L. Different letters in the same line indicate significant differences.
The presence of M. pulcherrima induced an increase in the esters content of 60%. In particular, the sequential inoculation induced an increase in ethyl decanoate (fruity, juicy, pineapple), ethyl octanoate (fruity, winey, sweet, apricot, banana, brandy, pear), isoamyl acetate (sweet, fruity, banana), and phenethyl acetate (floral, rose, sweet, honey, fruity, tropical). These results are in agreement with previous studies that demonstrated that the use of M. pulcherrima as a co-starter with S. cerevisiae resulted in an increase in the esters content [11,12,44]. Zhang et al. [12] revealed that sequential inoculation (M. pulcherrima + S. cerevisiae) induced an increase in acetate esters. Similarly, Canonico et al. [44] highlighted that this combination induced an increase in the ethyl butyrate, ethyl hexanoate, phenyl ethyl acetate, and isoamyl acetate content. Varela et al. [11] showed that wines inoculated at T0 with M. pulcherrima showed significantly higher concentrations of ethyl acetate, ethyl propanoate, 2-methypropyl acetate, ethyl butanoate, and ethyl hexanoate and lower concentrations of 2-methylbutyl acetate, 2-methylbutyl acetate, hexyl acetate, and ethyl decanoate. These data could be related to the extracellular enzymatic activity of this non-Saccharomyces yeast. It is well-known that M. pulcherrima shows several enzymatic activities, such as pectinase, protease, glucanase, lichenase, β-glucosidase, cellulase, xylanase, amylase, sulfite reductase, lipase, and β-lyase activity [10,14].
The content of higher alcohol was higher in F2 wines (27.54 mg/L) than in F1 (26.65 mg/L). In particular, F2 wines were characterized by a reduced content of isoamyl alcohol (fusel, alcoholic, whiskey, fruity, banana) and an increased content of phenethyl alcohol (sweet, floral, fresh, bready, rose, honey). An increase in 2-phenylethanol is generally attributed to sequential inoculation with non-Saccharomyces [9,15,33]. Several studies highlighted the increase in higher alcohols in M. pulcherrima co-fermentations [17,45,46,47,48]. The mechanisms underlying this phenomenon are not completely understood. A possible explanation could be linked to the differential usage of amino acids between S. cerevisiae and mixed cultures (sequential inoculation and co-inoculum). However, it should be noted that the relationship between amino acid composition and the volatile profile of the wine is not necessarily straightforward in sequential inoculation [48]. Moreover, higher alcohols can also be synthesized from glucose via pyruvate [49]. Partial sugar diversion towards higher alcohols instead of ethanol remains a possibility in sequential inoculations.
Interesting results were obtained also for terpenes. In fact, the sequential inoculation resulted in an increase in their content of about 80%. Several studies reported an increase in the total terpene concentration at the end of sequential fermentations involving M. pulcherrima compared to a single fermentation of S. cerevisiae up to 100% [9,15,37,50,51,52,53]. Terpenes predominantly originating from grapes are released and modulated by microbial activity, although de novo synthesis has been reported for both S. cerevisiae and non-Saccharomyces yeasts [9,50]. In particular, the α-arabinofuranosidase and β-glucosidase activities associated with M. pulcherrima strains [53] could account for the increase in monoterpenoids seen in the sequentially inoculated wines. The main effect was observed for linalool whose concentration was 0.32 mg/L in the F1 wines and 1.44 mg/L in the F2 wines and for β-damascenone (0.01 mg/L in F1 wines and 0.02 mg/L in F2 wines). These results are in agreement with previous studies that demonstrated an increase in these compounds in the presence of M. pulcherrima [15,37,51,52].
3.5. Sensory Profile of Wines
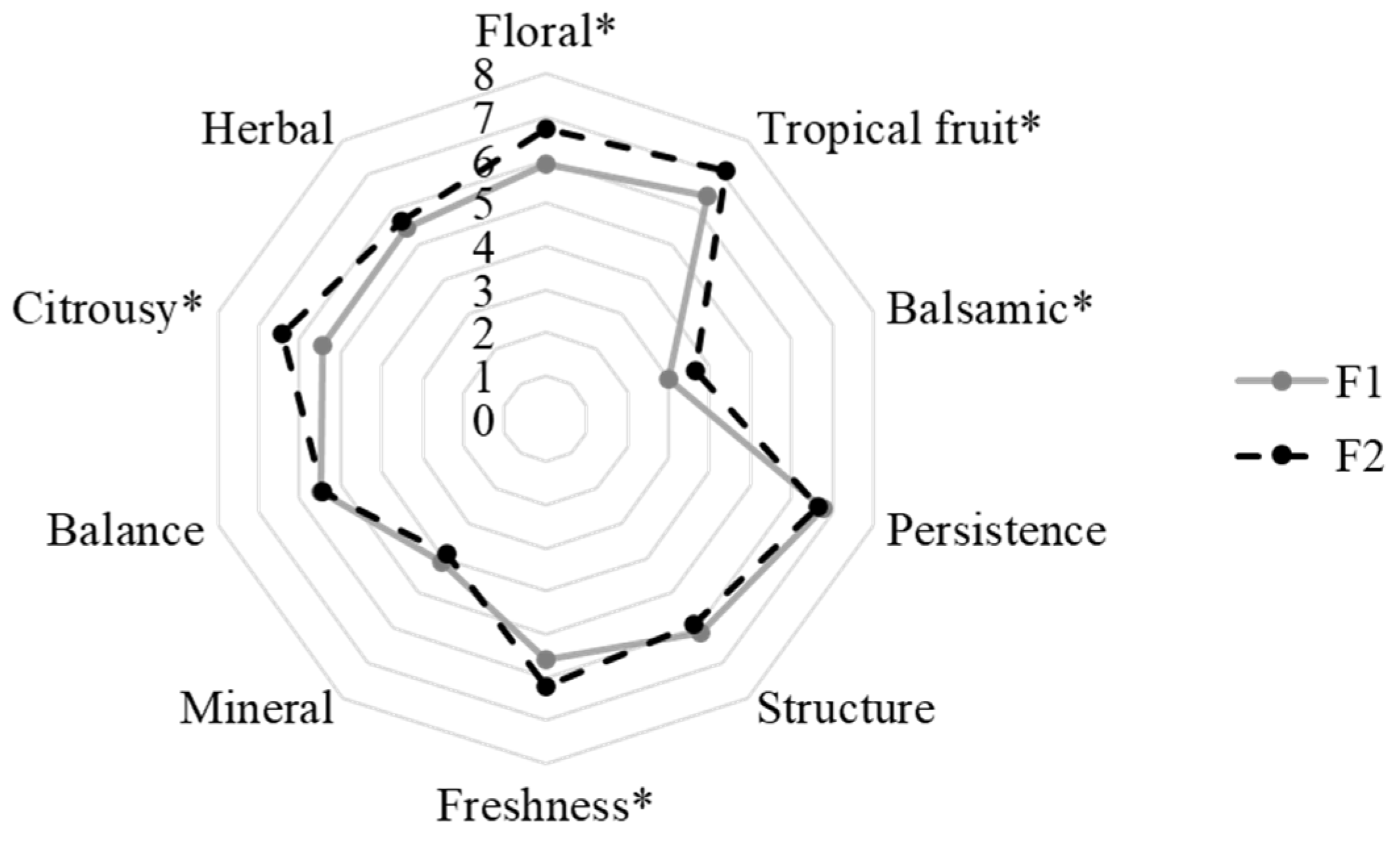
Sensory analysis was performed to better investigate the role of M. pulcherrima in aroma complexity. The results are reported in Figure 2. The obtained results highlighted a positive role of M. pulcherrima in the definition of the wine sensory profile. In particular, the F2 wines showed higher scores than the F1 wines for the following descriptors: floral, tropical fruit, citrusy, balsamic, and freshness. The floral, citrusy, and fruity notes could be related to the presence of some esters, e.g., phenethyl acetate, higher alcohols, e.g., phenethyl alcohol, and terpenes which were detected in higher concentrations in these wines than in those obtained with S. cerevisiae as a single culture. The wine's freshness could be defined by the combination of a fruity aroma reminiscent of the grape variety, moderate ethanol content, and high acidity [54]. Therefore, the perception of this descriptor could be connected with the production of esters and the lower ethanol content detected in the F2 wines.
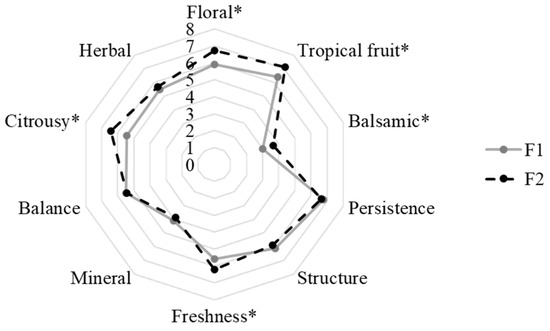
Figure 2.
Sensory analysis of obtained wines. * indicated the significant differences (p < 0.05).
4. Conclusions
In this study, we have demonstrated the collaboration between S. cerevisiae and M. pulcherrima as a biotechnological strategy to produce Pecorino wines with a reduced ethanol content and an increased freshness. Moreover, the use of sequential inoculation allowed for the improvement of the Pecorino wine's complexity, increasing the esters and terpenes content and reducing the copper content in the wines. The results indicated that the sequential inoculation of M. pulcherrima GS80 and S. cerevisiae SRS1 represents a good strategy to enhance the quality, improve the complexity, and modify some of the undesired parameters of the final wines facing the new challenges of the wine industry posed by climate change. Although M. pulcherrima GS80 and S. cerevisiae SRS1 positively cooperated to shape the Pecorino characteristics, further studies are necessary to test other Metschnikowia species, such as M. fruticola and M. viticola, in winemaking in combination with S. cerevisiae to differentiate Pecorino wines.
Author Contributions
Conceptualization, R.T.; methodology, A.P.R. and G.P.; formal analysis, G.P. and A.P.R.; investigation, A.P.R.; data curation, G.P. and A.P.R.; writing—original draft preparation, R.T., G.P. and G.S.; writing—review and editing, R.T., G.P., G.A., L.Q., A.P. and G.S.; funding acquisition, R.T. All authors have read and agreed to the published version of the manuscript.
Funding
The authors are thankful to European Union—Next Generation EU. Project Code: ECS00000041; Project CUP: C43C22000380007; Project Title: Innovation, digitalization, and sustainability for the diffused economy in Central Italy—VITALITY.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available within this article.
Acknowledgments
The authors acknowledge Luca Di Campli for technical support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Liu, P.T.; Lu, L.; Duan, C.Q.; Yan, G.L. The contribution of indigenous non-Saccharomyces wine yeast to improved aromatic quality of Cabernet Sauvignon wines by spontaneous fermentation. LWT-Food Sci. Technol. 2016, 71, 356–363. [Google Scholar] [CrossRef]
- Wei, R.T.; Chen, N.; Ding, Y.T.; Wang, L.; Liu, Y.H.; Gao, F.F.; Zhang, L.; Li, H.; Wang, H. Correlations between microbiota with physicochemical properties and volatile compounds during the spontaneous fermentation of Cabernet Sauvignon (Vitis vinifera L.) wine. LWT-Food. Sci. Technol. 2022, 163, 113529. [Google Scholar] [CrossRef]
- Liu, D.; Howell, K. Community succession of the grapevine fungal microbiome in the annual growth cycle. Environ. Microbiol. 2020, 23, 1842–1857. [Google Scholar] [CrossRef]
- Escribano-Viana, R.; Gonzalez-Arenzana, L.; Garijo, P.; Lopez, R.; Santamaría, P.; Gutierrez, A.R. Selection process of a mixed inoculum of non-Saccharomyces yeasts isolated in the D.O.Ca. Rioja. Fermentation 2021, 7, 148. [Google Scholar] [CrossRef]
- Suzzi, G.; Arfelli, G.; Schirone, M.; Corsetti, A.; Perpetuini, G.; Tofalo, R. Effect of grape indigenous Saccharomyces cerevisiae strains on Montepulciano d’Abruzzo red wine quality. Food Res. Int. 2012, 46, 22–29. [Google Scholar] [CrossRef]
- Suzzi, G.; Schirone, M.; Sergi, M.; Marianella, R.M.; Fasoli, G.; Aguzzi, I.; Tofalo, R. Multistarter from organic viticulture for red wine Montepulciano d’Abruzzo production. Front. Microbiol. 2012, 3, 135. [Google Scholar]
- Tofalo, R.; Patrignani, F.; Lanciotti, R.; Perpetuini, G.; Schirone, M.; Di Gianvito, P.; Pizzoni, D.; Arfelli, G.; Suzzi, G. Aroma profile of Montepulciano d’Abruzzo wine fermented by single and co-culture starters of autochthonous Saccharomyces and non-Saccharomyces yeasts. Front. Microbiol. 2016, 7, 610. [Google Scholar] [CrossRef] [PubMed]
- Padilla, B.; Zulian, L.; Ferreres, À.; Pastor, R.; Esteve-Zarzoso, B.; Beltran, G.; Mas, A. Sequential inoculation of native non-Saccharomyces and Saccharomyces cerevisiae strains for wine making. Front. Microbiol. 2017, 8, 1293. [Google Scholar] [CrossRef]
- Padilla, B.; Gil, J.V.; Manzanares, P. Past and future of non-Saccharomyces yeasts: From spoilage microorganisms to biotechnological tools for improving wine aroma complexity. Front. Microbiol. 2016, 7, 411. [Google Scholar] [CrossRef]
- Barbosa, C.; Lage, P.; Esteves, M.; Chambel, L.; Mendes-Faia, A.; Mendes-Ferreira, A. Molecular and phenotypic characterization of Metschnikowia pulcherrima strains from Douro wine region. Fermentation 2018, 4, 8. [Google Scholar] [CrossRef]
- Varela, C.; Sengler, F.; Solomon, M.; Curtin, C. Volatile flavour profile of reduced alcohol wines fermented with the non-conventional yeast species Metschnikowia pulcherrima and Saccharomyces uvarum. Food Chem. 2016, 209, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.-Q.; Shen, J.-Y.; Duan, C.-Q.; Yan, G.-L. Use of indigenous Hanseniaspora vineae and Metschnikowia pulcherrima co-fermentation with Saccharomyces cerevisiae to improve the aroma diversity of Vidal Blanc icewine. Front. Microbiol. 2018, 9, 2303. [Google Scholar] [CrossRef]
- Ruiz, J.; Belda, I.; Beisert, B.; Navascues, E.; Marquina, D.; Calderon, F.; Rauhut, D.; Santos, A.; Benito, S. Analytical impact of Metschnikowia pulcherrima in the volatile profile of Verdejo white wines. Appl. Microbiol. Biotechnol. 2018, 102, 8501–8509. [Google Scholar] [CrossRef] [PubMed]
- Vicente, J.; Ruiz, J.; Belda, I.; Benito-Vázquez, I.; Marquina, D.; Calderón, F.; Santos, A.; Benito, S. The genus Metschnikowia in enology. Microorganisms 2020, 8, 1038. [Google Scholar] [CrossRef]
- Sadoudi, M.; Tourdot-Marechal, R.; Rousseaux, S.; Steyer, D.; Gallardo-Chacon, J.J.; Ballester, J.; Vichi, S.; Guerin-Schneider, R.; Caixach, J.; Alexandre, H. Yeast-yeast interactions revealed by aromatic profile analysis of Sauvignon Blanc wine fermented by single or co-culture of non-Saccharomyces and Saccharomyces yeasts. Food Microbiol. 2012, 32, 243–253. [Google Scholar] [CrossRef]
- Marangon, M.; Van Sluyter, S.C.; Robinson, E.M.; Muhlack, R.A.; Holt, H.E.; Haynes, P.A.; Godden, P.W.; Smith, P.A.; Waters, E.J. Degradation of white wine haze proteins by Aspergillopepsin I and II during juice flash pasteurization. Food Chem. 2012, 135, 1157–1165. [Google Scholar] [CrossRef] [PubMed]
- Hranilovic, A.; Gambetta, J.M.; Jeffery, D.W.; Grbin, P.R.; Jiranek, V. Lower-alcohol wines produced by Metschnikowia pulcherrima and Saccharomyces cerevisiae co-fermentations: The effect of sequential inoculation timing. Int. J. Food Microbiol. 2020, 329, 108651. [Google Scholar] [CrossRef] [PubMed]
- Oro, L.; Ciani, M.; Comitini, F. Antimicrobial activity of Metschnikowia pulcherrima on wine yeasts. J. Appl. Microbiol. 2014, 116, 1209–1217. [Google Scholar] [CrossRef]
- Available online: https://biodiversita.umbria.parco3a.org/storymap/content/Scheda_Iscrizione_Registro_Regionale_Vitigno_Pecorino.pdf (accessed on 3 July 2023).
- Organisation International de la Vigne et du Vin (OIV). Compendium of International Methods of Wine and Must Analysis. Vol. 1 and Paris, France. Available online: http://188.165.107.123/en/technical-standards-and-documents/methods-of-analysis/compendium-of-international-methods-of-analysis-of-wines-and-musts-2-vol (accessed on 19 July 2023).
- Dournes, G.; Dufourcq, T.; Suc, L.; Roland, A.; Mouret, J.R. Unravelling copper effect on the production of varietal thiols during Colombard and Gros Manseng grape juices fermentation by Saccharomyces cerevisiae. Front. Microbiol. 2023, 14, 1101110. [Google Scholar] [CrossRef]
- Rossetti, A.P.; Perpetuini, G.; Battistelli, N.; Zulli, C.; Arfelli, G.; Suzzi, G.; Cichelli, A.; Tofalo, R. Capturing the fungal community associated with conventional and organic Trebbiano Abruzzese grapes and its influence on wine characteristics. Food Biosci. 2023, 52, 102382. [Google Scholar] [CrossRef]
- Perpetuini, G.; Rossetti, A.P.; Battistelli, N.; Zulli, C.; Piva, A.; Arfelli, G.; Corsetti, A.; Tofalo, R. Contribution of Starmerella bacillaris and oak chips to Trebbiano d’Abruzzo wine volatile and sensory diversity. Foods 2023, 12, 1102. [Google Scholar] [CrossRef] [PubMed]
- ISO 8586:2012; Sensory Analysis—General Guidelines for the Selection, Training and Monitoring of Selected Assessors and Expert Sensory Assessors. International Organization for Standardization: Geneva, Switzerland, 2012.
- ISO 8589:2007; Sensory Analysis—General Guidance for the Design of Test Rooms. International Organization for Standardization: Geneva, Switzerland, 2007.
- Kuchen, B.; Maturano, Y.P.; Mestre, M.V.; Combina, M.; Toro, M.E.; Vazquez, F. Selection of native non-Saccharomyces yeasts with biocontrol activity against spoilage yeasts in order to produce healthy regional wines. Fermentation 2019, 5, 60. [Google Scholar] [CrossRef]
- Checchia, I.; Binati, R.L.; Troiano, E.; Ugliano, M.; Felis, G.E.; Torriani, S. Unravelling the impact of grape washing, SO2, and multi-starter inoculation in lab-scale vinification trials of withered black grapes. Fermentation 2021, 7, 43. [Google Scholar] [CrossRef]
- Quiros, M.; Rojas, V.; Gonzalez, R.; Morales, P. Selection of non-Saccharomyces yeast strains for reducing alcohol levels in wine by sugar respiration. Int. J. Food Microbiol. 2014, 181, 85–91. [Google Scholar] [CrossRef]
- Contreras, A.; Hidalgo, C.; Henschke, P.A.; Chambers, P.J.; Curtin, C.; Varela, C. Evaluation of non-Saccharomyces yeasts for the reduction of alcohol content in wine. Appl. Environ. Microbiol. 2014, 80, 1670–1678. [Google Scholar] [CrossRef] [PubMed]
- Morales, P.; Rojas, V.; Quiros, M.; Gonzalez, R. The impact of oxygen on the final alcohol content of wine fermented by a mixed starter culture. Appl. Microbiol. Biotechnol. 2015, 99, 3993–4003. [Google Scholar] [CrossRef] [PubMed]
- Droulia, F.; Charalampopoulos, I. A Review on the Observed Climate Change in Europe and Its Impacts on Viticulture. Atmosphere 2022, 13, 837. [Google Scholar] [CrossRef]
- Sadineni Naresh, V.; Kondapalli, N.; Obulam, V. Effect of co-fermentation with Saccharomyces cerevisiae and Torulaspora delbrueckii or Metschnikowia pulcherrima on the aroma and sensory properties of mango wine. Ann. Microbiol. 2012, 62, 1353–1360. [Google Scholar] [CrossRef]
- Comitini, F.; Gobbi, M.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Selected non-Saccharomyces wine yeasts in controlled multistarter fermentations with Saccharomyces cerevisiae. Food Microbiol. 2011, 28, 873–882. [Google Scholar] [CrossRef]
- Jolly, N.; Augustyn, O.; Pretorius, I. The effect of non-Saccharomyces yeasts on fermentation and wine quality. J. South Afr. J. Enol. 2003, 24, 55–62. [Google Scholar] [CrossRef]
- Roca-Mesa, H.; Sendra, S.; Mas, A.; Beltran, G.; Torija, M.J. Nitrogen preferences during alcoholic fermentation of different non-Saccharomyces yeasts of oenological interest. Microorganisms 2020, 8, 157. [Google Scholar] [CrossRef] [PubMed]
- Benito, S.; Hofmann, T.; Laier, M.; Lochbühler, B.; Schüttler, A.; Ebert, K.; Fritsch, S.; Röcker, J.; Rauhut, D. Effect on quality and composition of Riesling wines fermented by sequential inoculation with non-Saccharomyces and Saccharomyces cerevisiae. Eur. Food Res. Technol. 2015, 241, 707–717. [Google Scholar] [CrossRef]
- Dutraive, O.; Benito, S.; Fritsch, S.; Beisert, B.; Patz, C.-D.; Rauhut, D. Effect of sequential inoculation with non-Saccharomyces and Saccharomyces yeasts on Riesling wine chemical composition. Fermentation 2019, 5, 79. [Google Scholar] [CrossRef]
- Ferreira, J.; Toit, M.; Toit, W. The effects of copper and high sugar concentrations on growth, fermentation efficiency and volatile acidity production of different commercial wine yeast strains. Aust. J. Grape Wine Res. 2006, 12, 50–56. [Google Scholar] [CrossRef]
- Robinson, N.; Winge, D. Copper Metallochaperones. Annu. Rev. Biochem. 2010, 79, 537–562. [Google Scholar] [CrossRef]
- Sun, X.Y.; Zhao, Y.; Liu, L.L.; Jia, B.; Zhao, F.; Huang, W.D.; Zhan, J.C. Copper Tolerance and biosorption of Saccharomyces cerevisiae during alcoholic fermentation. PLoS ONE 2015, 10, e0128611. [Google Scholar] [CrossRef]
- Bely, M.; Stoeckle, P.; Masneuf-Pomarède, I.; Dubourdieu, D. Impact of mixed Torulaspora delbrueckii–Saccharomyces cerevisiae culture on high-sugar fermentation. Int. J. Food Microbiol. 2008, 122, 312–320. [Google Scholar] [CrossRef]
- Milanovic, V.; Ciani, M.; Oro, L.; Comitini, F. Starmerella bombicola influences the metabolism of Saccharomyces cerevisiae at pyruvate decarboxylase and alcohol dehydrogenase level during mixed wine fermentation. Microb. Cell Fact. 2012, 11, 18. [Google Scholar] [CrossRef]
- Sadoudi, M.; Rousseaux, S.; David, V.; Alexandre, H.; Tourdot-Maréchal, R. Metschnikowia pulcherrima influences the expression of genes involved in PDH bypass and glyceropyruvic fermentation in Saccharomyces cerevisiae. Front. Microbiol. 2017, 8, 1137. [Google Scholar] [CrossRef]
- Canonico, L.; Agarbati, A.; Galli, E.; Comitini, F.; Ciani, M. Metschnikowia pulcherrima as biocontrol agent and wine aroma enhancer in combination with a native Saccharomyces cerevisiae. LWT-Food Sci. Technol. 2023, 181, 114758. [Google Scholar] [CrossRef]
- Canonico, L.; Solomon, M.; Comitini, F.; Ciani, M.; Varela, C. Volatile profile of reduced alcohol wines fermented with selected non-Saccharomyces yeasts under different aeration conditions. Food Microbiol. 2019, 84, 103247. [Google Scholar] [CrossRef] [PubMed]
- Shekhawat, K.; Bauer, F.F.; Setati, M.E. Impact of oxygenation on the performance of three non-Saccharomyces yeasts in co-fermentation with Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2017, 101, 2479–2491. [Google Scholar] [CrossRef]
- Tronchoni, J.; Curiel, J.A.; Saenz-Navajas, M.P.; Morales, P.; de-la-Fuente-Blanco, A.; Fernandez-Zurbano, P.; Ferreira, V.; Gonzalez, R. Aroma profiling of an aerated fermentation of natural grape must with selected yeast strains at pilot scale. Food Microbiol. 2018, 70, 214–223. [Google Scholar] [CrossRef] [PubMed]
- Gobert, A.; Tourdot-Marechal, R.; Morge, C.; Sparrow, C.; Liu, Y.; Quintanilla-Casas, B.; Vichi, S.; Alexandre, H. Non-Saccharomyces yeasts nitrogen source preferences: Impact on sequential fermentation and wine volatile compounds profile. Front. Microbiol. 2017, 8, 2175. [Google Scholar] [CrossRef]
- Prior, K.J.; Bauer, F.F.; Divol, B. The utilisation of nitrogenous compounds by commercial non-Saccharomyces yeasts associated with wine. Food Microbiol. 2019, 79, 75–84. [Google Scholar] [CrossRef]
- Cordente, A.G.; Curtin, C.D.; Varela, C.; Pretorius, I.S. Flavour-active wine yeasts. Appl. Microbiol. Biotechnol. 2012, 96, 601–618. [Google Scholar] [CrossRef] [PubMed]
- Rossouw, D.; Bauer, F.F. Exploring the phenotypic space of non-Saccharomyces wine yeast biodiversity. Food Microbiol. 2016, 55, 32–46. [Google Scholar] [CrossRef]
- García, M.; Esteve-Zarzoso, B.; Cabellos, J.M.; Arroyo, T. Sequential non-Saccharomyces and Saccharomyces cerevisiae fermentations to reduce the alcohol content in wine. Fermentation 2020, 6, 60. [Google Scholar] [CrossRef]
- Rodriguez, M.E.; Lopes, C.A.; Barbagelata, R.J.; Barda, N.B.; Caballero, A.C. Influence of Candida pulcherrima Patagonian strain on alcoholic fermentation behaviour and wine aroma. Int. J. Food Microbiol. 2010, 138, 19–25. [Google Scholar] [CrossRef]
- Morata, A.; Escott, C.; Bañuelos, M.A.; Loira, I.; Fresno, J.M.D.; González, C.; Suárez-Lepe, J.A. Contribution of non-Saccharomyces yeasts to wine freshness. A Review. Biomolecules 2019, 10, 34. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).