
Highly Efficient Biosynthesis of γ-Bisabolene with a New Sesquiterpene Synthase AcTPS5 by Dual Cytoplasmic-Peroxisomal Engineering in Saccharomyces cerevisiae
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Bioinformatic Analysis of Bisabolene Synthases
2.2. Medium, Culture Conditions, and Chemicals
2.3. Construction of Plasmids and Strains
2.4. Shake-Flask Fermentation
2.5. Fed-Batch Fermentation for γ-Bisabolene Biosynthesis
2.6. Analytical Methods
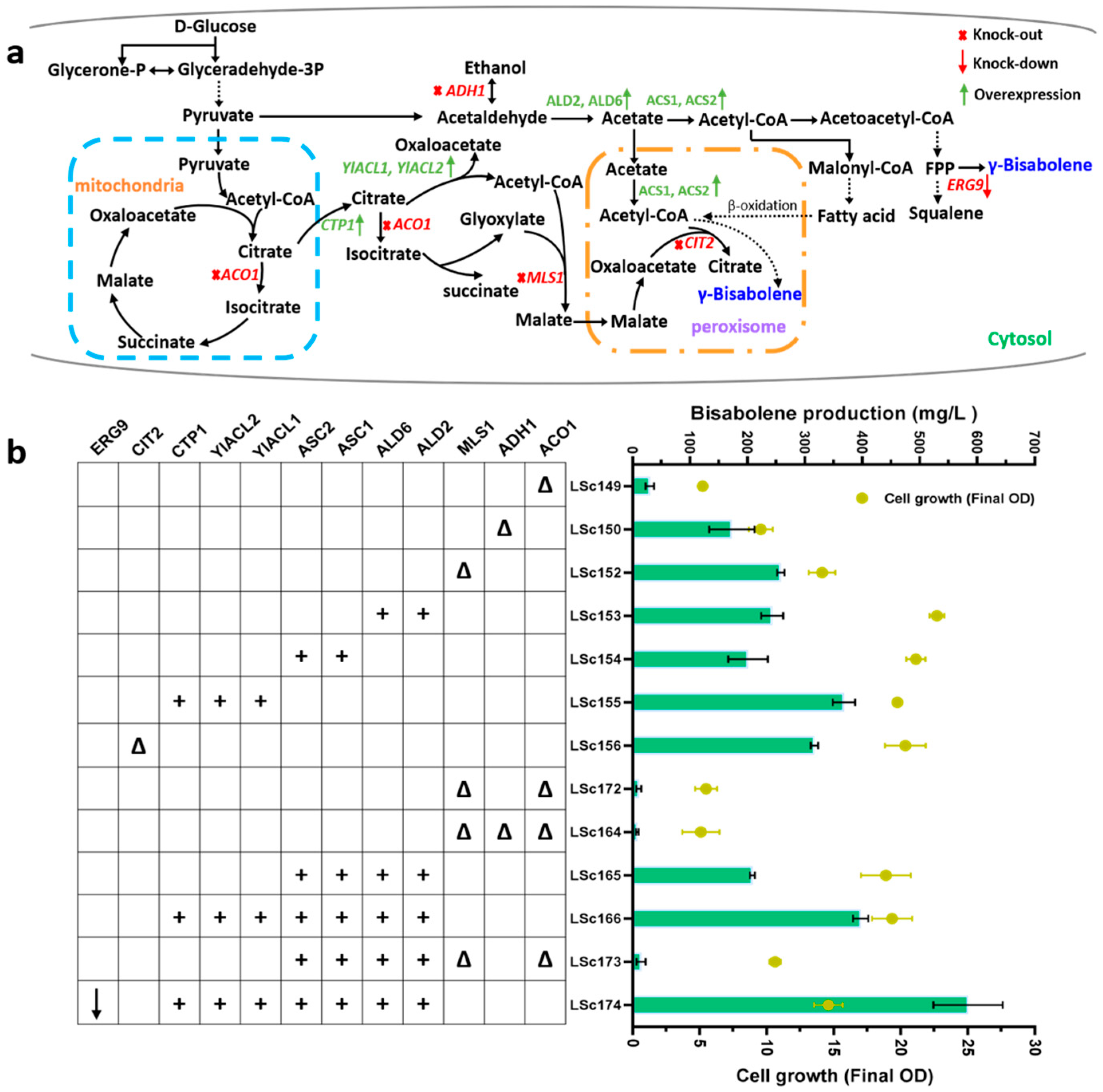
3. Results and Discussion
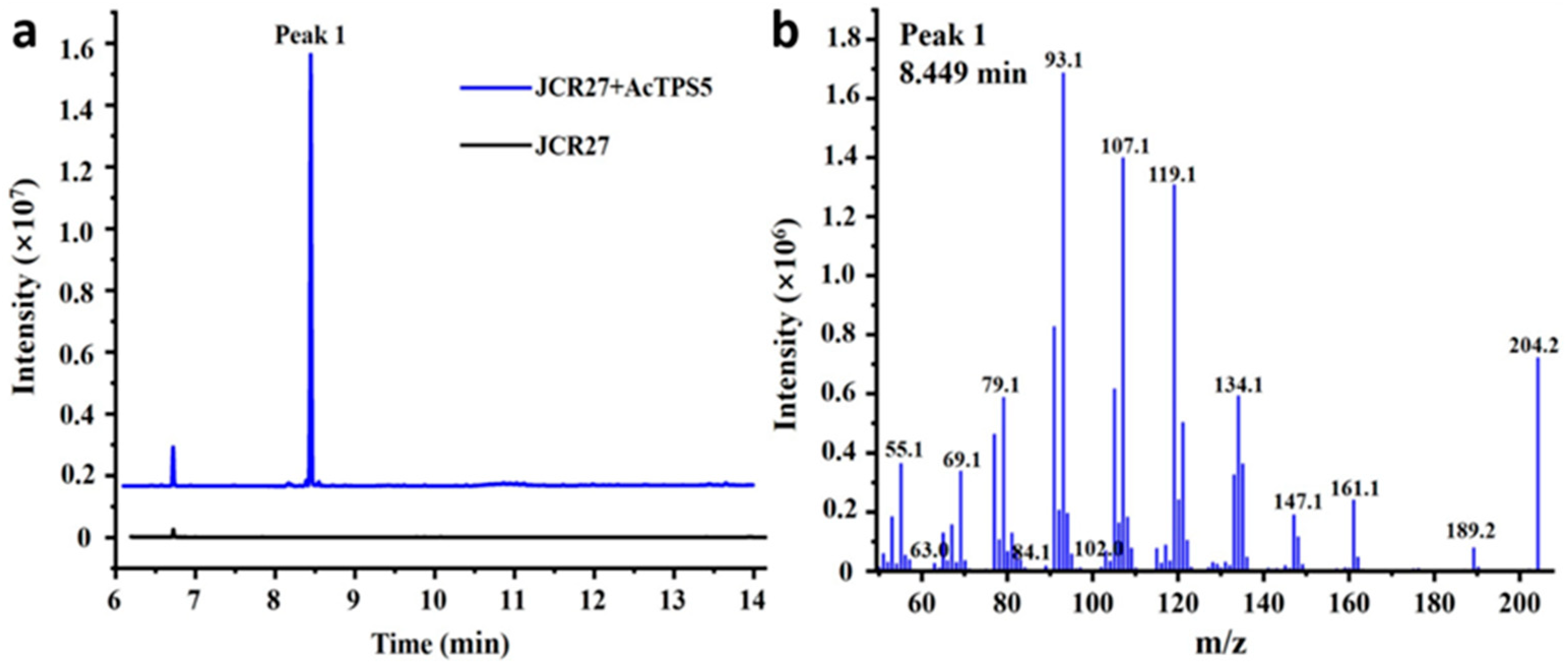
3.1. Functional Characterization and Sequence Analysis of AcTPS5
3.2. Producing γ-Bisabolene in S. cerevisiae Peroxisomes
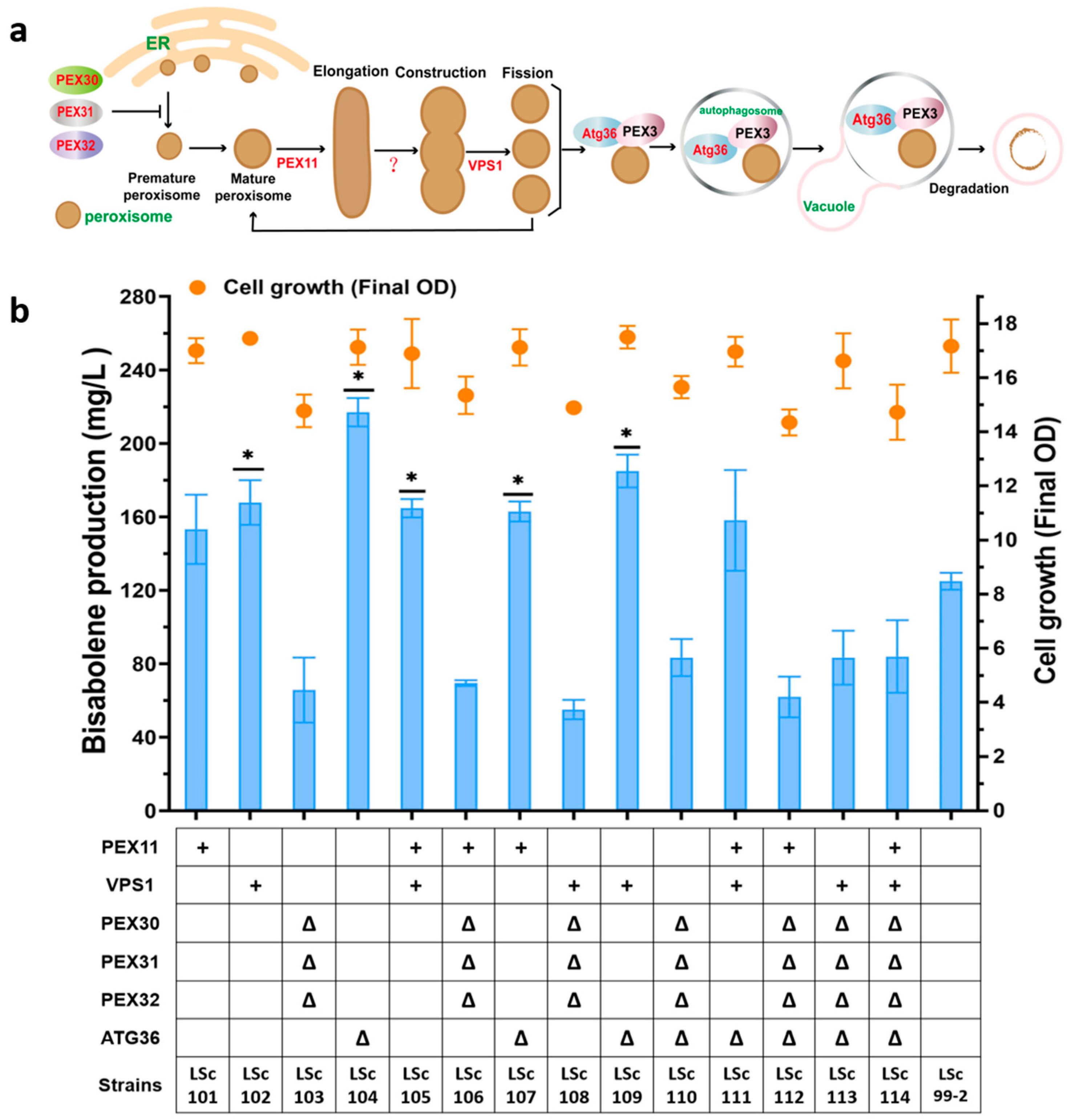
3.3. Harnessing Peroxisomes to Enhance γ-Bisabolene Production
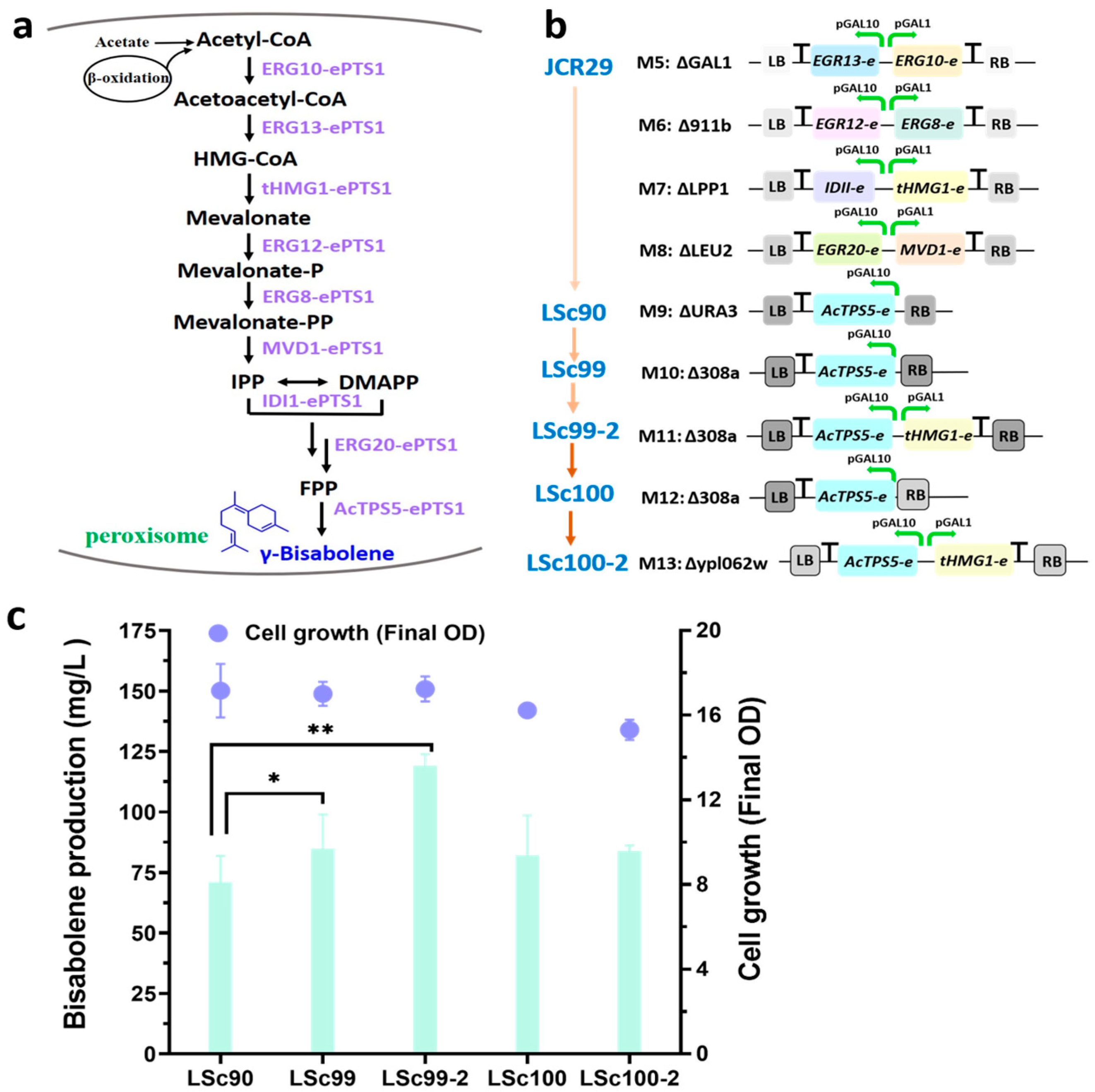
3.4. Dual Cytoplasmic-Peroxisomal Engineering to Optimize γ-Bisabolene Production
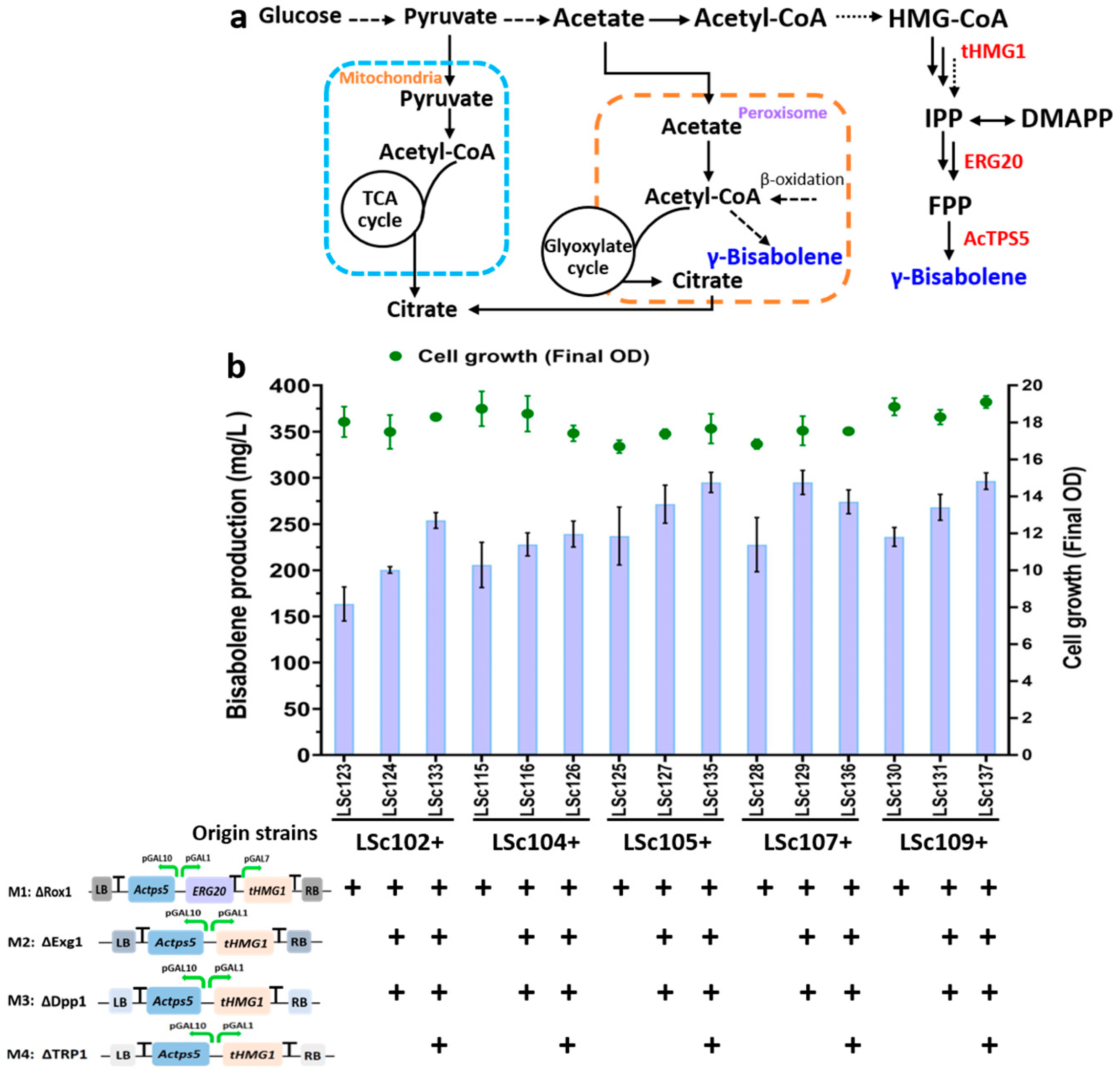
3.5. Engineering Acetyl-CoA Supply to Overproduce Bisabolene
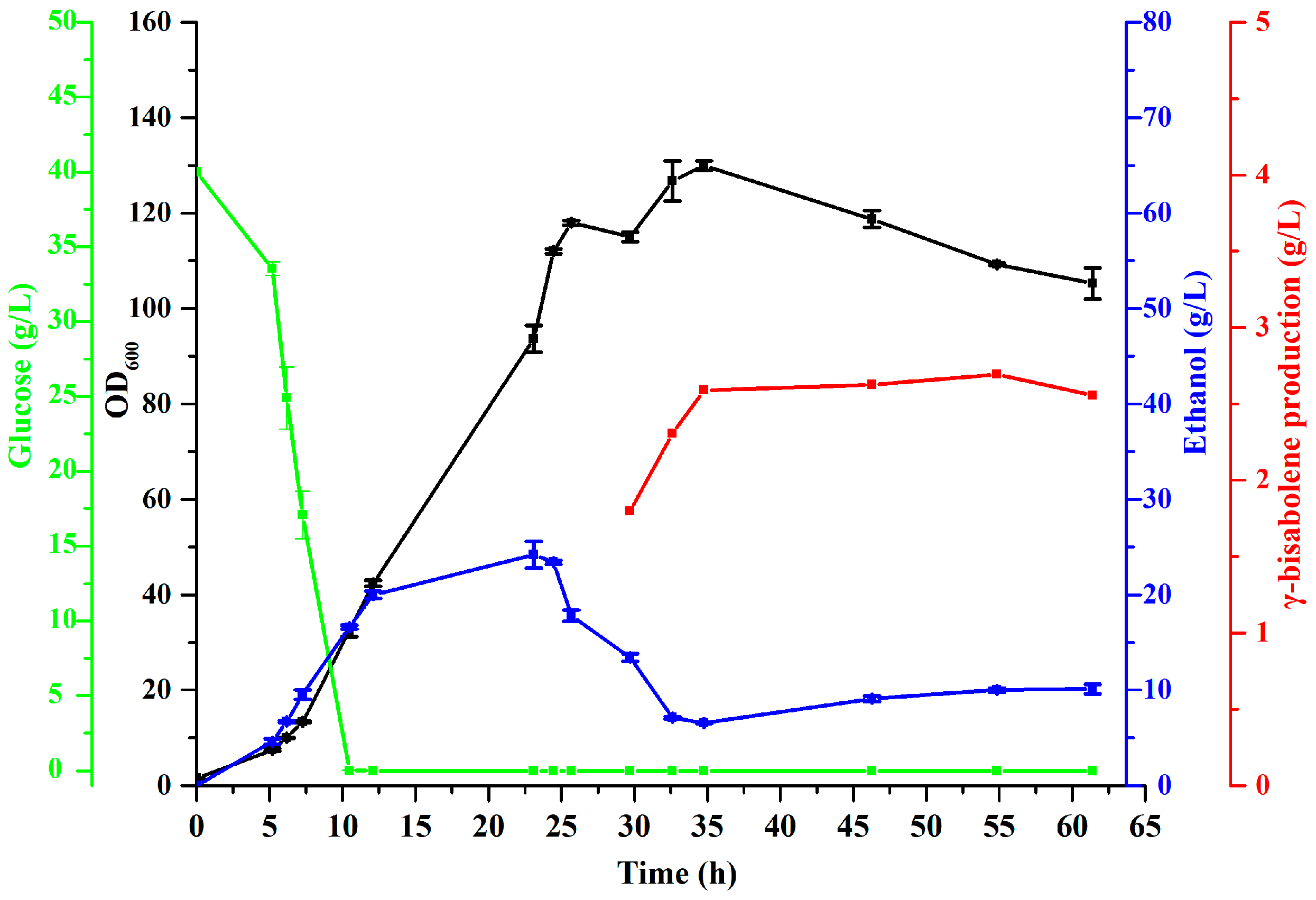
3.6. High-Density Fermentation for γ-Bisabolene Production
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, C.L.; Xue, K.; Yang, Y.K.; Liu, X.X.; Li, Y.; Lee, T.; Bai, Z.; Tan, T. Metabolic engineering strategies for sesquiterpene production in microorganism. Crit. Rev. Biotechnol. 2022, 42, 3–92. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Wang, X. Biosynthesis of monoterpenoid and sesquiterpenoid as natural flavors and fragrances. Biotechnol. Adv. 2023, 65, 108151. [Google Scholar] [CrossRef] [PubMed]
- Hewage, R.T.; Tseng, C.C.; Liang, S.; Lai, C.Y.; Lin, H.C. Genome mining of cryptic bisabolenes that were biosynthesized by intramembrane terpene synthases from Antrodia cinnamomea. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2023, 378, 20220033. [Google Scholar] [CrossRef] [PubMed]
- Peralta-Yahya, P.P.; Ouellet, M.; Chan, R.; Mukhopadhyay, A.; Keasling, J.D.; Lee, T.S. Identification and microbial production of a terpene-based advanced biofuel. Nat. Commun. 2011, 2, 483. [Google Scholar] [CrossRef]
- Kamatou, G.P.P.; Viljoen, A.M. A review of the application and pharmacological properties of alpha-bisabolol and alpha-bisabolol-rich oils. J. Am. Oil Chem. Soc. 2010, 87, 1–7. [Google Scholar] [CrossRef]
- Yeo, S.K.; Ali, A.Y.; Hayward, O.A.; Turnham, D.; Jackson, T.; Bowen, I.D.; Clarkson, R. β-Bisabolene, a sesquiterpene from the essential oil extract of Opoponax (Commiphora guidottii), exhibits cytotoxicity in breast cancer cell lines. Phytother. Res. 2016, 30, 418–425. [Google Scholar] [CrossRef]
- Nascimento, A.M.; Brandao, M.G.; Oliveira, G.B.; Fortes, I.C.; Chartone-Souza, E. Synergistic bactericidal activity of Eremanthus erythropappus oil or β-bisabolene with ampicillin against Staphylococcus aureus. Antonie Leeuwenhoek. 2007, 92, 95–100. [Google Scholar] [CrossRef]
- Shu, H.Z.; Peng, C.; Bu, L.; Guo, L.; Liu, F.; Xiong, L. Bisabolane-type sesquiterpenoids: Structural diversity and biological activity. Phytochemistry 2021, 192, 112927. [Google Scholar] [CrossRef]
- Jou, Y.J.; Hua, C.H.; Lin, C.S.; Wang, C.Y.; Wan, L.; Lin, Y.J.; Huang, S.H.; Lin, C.W. Anticancer activity of γ-bisabolene in human neuroblastoma cells via induction of p53-mediated mitochondrial apoptosis. Molecules 2016, 21, 601. [Google Scholar] [CrossRef]
- Govindarajan, M.; Vaseeharan, B.; Alharbi, N.S.; Kadaikunnan, S.; Khaled, J.M.; Al-Anbr, M.N.; Alyahya, S.A.; Maggi, F.; Benelli, G. High efficacy of (Z)-γ-bisabolene from the essential oil of Galinsoga parviflora (Asteraceae) as larvicide and oviposition deterrent against six mosquito vectors. Environ. Sci. Pollut. Res. Int. 2018, 25, 10555–10566. [Google Scholar] [CrossRef]
- Lim, D.S.; Shi, L.L.; Guo, K.; Luo, S.; Liu, Y.; Chen, Y.; Liu, Y.; Li, S.H. A new sesquiterpene synthase catalyzing the formation of (R)-β-bisabolene from medicinal plant Colquhounia coccinea var. mollis and its anti-adipogenic and antibacterial activities. Phytochemistry 2023, 211, 113681. [Google Scholar]
- Chemat, F.; Vian, M.A.; Cravotto, G. Green extraction of natural products: Concept and principles. Int. J. Mol. Sci. 2012, 13, 8615–8627. [Google Scholar] [CrossRef] [PubMed]
- Walls, L.E.; Otoupal, P.; Ledesma-Amaro, R.; Velasquez-Orta, S.B.; Gladden, J.M.; Rios-Solis, L. Bioconversion of cellulose into bisabolene using Ruminococcus flavefaciens and Rhodosporidium toruloides. Bioresour. Technol. 2023, 368, 128216. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhu, K.; Li, J.; Zhao, Y.; Li, S.L.; Zhang, C.; Xiao, D.; Yu, A.Q. High-efficiency production of bisabolene from waste cooking oil by metabolically engineered Yarrowia lipolytica. Microb. Biotechnol. 2021, 14, 2497–2513. [Google Scholar] [CrossRef] [PubMed]
- Kulagina, N.N.; Besseau, S.; Papon, N.; Courdavault, V. Peroxisomes: A New Hub for Metabolic Engineering in Yeast. Front. Bioeng. Biotechnol. 2021, 9, 659431. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.B.; Li, M.; Zhao, G.; Lu, W.Y. Harnessing Yeast Peroxisomes and cytosol acetyl-CoA for sesquiterpene α-Humulene production. J. Agric. Food Chem. 2020, 68, 1382–1389. [Google Scholar] [CrossRef]
- Gerke, J.; Frauendorf, H.; Schneider, D.; Wintergoller, M.; Hofmeister, T.; Poehlein, A.; Zebec, Z.; Takano, E.; Scrutton, N.S.; Braus, G.H. Production of the fragrance geraniol in peroxisomes of a product-tolerant baker’s yeast. Front. Bioeng. Biotechnol. 2020, 8, 582052. [Google Scholar] [CrossRef]
- Liu, G.S.; Li, T.; Zhou, W.; Jiang, M.; Tao, X.Y.; Liu, M.; Zhao, M.; Ren, Y.; Gao, B.; Wang, F.Q.; et al. The yeast peroxisome: A dynamic storage depot and subcellular factory for squalene overproduction. Metab. Eng. 2020, 57, 151–161. [Google Scholar] [CrossRef]
- Hammer, S.K.; Avalos, J.L. Harnessing yeast organelles for metabolic engineering. Nat. Chem. Biol. 2017, 13, 823–832. [Google Scholar] [CrossRef]
- Liu, J.J.; Chen, C.; Wan, X.K.; Yao, G.; Bao, S.H.; Wang, F.L.; Wang, K.; Song, T.Y.; Han, P.G.; Jiang, H. Identification of the sesquiterpene synthase AcTPS1 and high production of (–)-germacrene D in metabolically engineered Saccharomyces cerevisiae. Microb. Cell Fact. 2022, 21, 89. [Google Scholar] [CrossRef]
- Rinkel, J.; Dickschat, J.S. Stereochemical investigations on the biosynthesis of achiral (Z)-γ-bisabolene in Cryptosporangium arvum. Beilstein J. Org. Chem. 2019, 15, 789–794. [Google Scholar] [CrossRef] [PubMed]
- Ichinose, H.; Ukeba, S.; Kitaoka, T. Latent potentials of the white-rot basidiomycete Phanerochaete chrysosporium responsible for sesquiterpene metabolism: CYP5158A1 and CYP5144C8 decorate (E)-α-bisabolene. Enzym. Microb. Technol. 2022, 158, 110037. [Google Scholar] [CrossRef] [PubMed]
- Siemon, T.; Wang, Z.Q.; Bian, G.K.; Seitz, T.; Ye, Z.L.; Lu, Y.; Cheng, S.; Ding, Y.K.; Huang, Y.L.; Deng, Z.X.; et al. Semisynthesis of plant-derived englerin A enabled by microbe engineering of guaia-6,10(14)-diene as building block. J. Am. Chem. Soc. 2020, 142, 2760–2765. [Google Scholar] [CrossRef]
- Wang, F.; Lv, X.; Xie, W.; Zhou, P.; Zhu, Y.; Yao, Z.; Yang, C.; Yang, X.; Ye, L.; Yu, H. Combining Gal4p-mediated expression enhancement and directed evolution of isoprene synthase to improve isoprene production in Saccharomyces cerevisiae. Metab. Eng. 2017, 39, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Jiang, G.Z.; Yao, M.D.; Wang, Y.; Zhou, L.; Song, T.; Liu, H.; Xiao, W.H.; Yuan, Y.J. Manipulation of GES and ERG20 for geraniol overproduction in Saccharomyces cerevisiae. Metab. Eng. 2017, 41, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Apel, A.R.; d’Espaux, L.; Wehrs, M.; Sachs, D.; Li, R.A.; Tong, G.J.; Garber, M.; Nnadi, O.; Zhuang, W.; Hillson, N.; et al. A Cas9-based toolkit to program gene expression in Saccharomyces cerevisiae. Nucleic Acids Res. 2017, 45, 496–508. [Google Scholar]
- Chen, Y.; Wang, Y.; Liu, M.; Qu, J.Z.; Yao, M.D.; Li, B.; Ding, M.Z.; Liu, H.; Xiao, W.H.; Yuan, Y.J. Primary and secondary metabolic effects of a key gene deletion (Δypl062w) in metabolically engineered terpenoid-producing Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2019, 85, e01990-18. [Google Scholar] [CrossRef] [PubMed]
- Akşit, A.; Klei, I.J. Yeast peroxisomes: How are they formed and how do they grow? Int. J. Biochem. Cell Biol. 2018, 105, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Rottensteiner, H.; Stein, K.; Sonnenhol, E.; Erdmann, R. Conserved Function of Pex11p and the Novel Pex25p and Pex27p in Peroxisome Biogenesis. Mol. Biol. Cell 2003, 14, 4316–4328. [Google Scholar] [CrossRef][Green Version]
- Fagarasanu, A.; Fagarasanu, M.; Rachubinski, R.A. Maintaining peroxisome populations: A story of division and inheritance. Annu. Rev. Cell Dev. Biol. 2007, 23, 321–344. [Google Scholar] [CrossRef]
- Motley, A.M.; Nuttall, J.M.; Hettema, E.H. Pex3-anchored Atg36 tags peroxisomes for degradation in Saccharomyces cerevisiae. EMBO J. 2012, 31, 2852–2868. [Google Scholar] [CrossRef]
- Donald, K.A.G.; Hampton, R.Y.; Fritz, I.B. Effects of overproduction of the catalytic domain of 3-hydroxy-3-methylglutaryl coenzyme A reductase on squalene synthesis in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 1997, 63, 3341–3344. [Google Scholar] [CrossRef]
- Lv, X.M.; Wang, F.; Zhou, P.P.; Ye, L.D.; Xie, W.P.; Xu, H.M.; Yu, H.W. Dual regulation of cytoplasmic and mitochondrial acetyl-CoA utilization for improved isoprene production in Saccharomyces cerevisiae. Nat. Commun. 2016, 7, 12851. [Google Scholar] [CrossRef]
- Jiang, Y.K.; Xia, L.; Gao, S.; Li, N.; Yu, S.Q.; Zhou, J.W. Engineering Saccharomyces cerevisiae for enhanced (–)-α-bisabolol production. Synth. Syst. Biotechnol. 2023, 8, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.M.; Shi, B.; Ye, Z.L.; Huang, M.; Chen, R.; Cai, Y.S.; Kuang, Z.L.; Sun, X.; Bian, G.K.; Deng, Z.X.; et al. Systematic identification of Ocimum sanctum sesquiterpenoid synthases and (–)-eremophilene overproduction in engineered yeast. Metab. Eng. 2022, 69, 122–133. [Google Scholar] [CrossRef] [PubMed]
- Henry, K.W.; Nickels, J.T.; Edlind, T.D. ROX1 and ERG regulation in Saccharomyces cerevisiae: Implications for antifungal susceptibility. Eukaryot. Cell 2002, 1, 1041–1044. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Lv, X.; Ye, L.; Zhou, P.; Yu, H. Construction of lycopene-overproducing Saccharomyces cerevisiae by combining directed evolution and metabolic engineering. Metab. Eng. 2015, 30, 69–78. [Google Scholar] [CrossRef]
- Faulkner, A.; Chen, X.; Rush, J.; Horazdovsky, B.; Waechter, C.J.; Carman, G.M.; Sternweis, P.C. The LPP1 and DPP1 gene products account for most of the isoprenoid phosphate phosphatase activities in Saccharomyces cerevisiae. J. Biol. Chem. 1999, 274, 14831–14837. [Google Scholar] [CrossRef]
- Wang, H.M.; Yang, Y.; Lin, L.; Zhou, W.L.; Liu, M.Z.; Cheng, K.D.; Wang, W. Engineering Saccharomyces cerevisiae with the deletion of endogenous glucosidases for the production of flavonoid glucosides. Microb. Cell Factories 2016, 15, 134. [Google Scholar] [CrossRef]
- Paramasivan, K.; Mutturi, S. Progress in terpene synthesis strategies through engineering of Saccharomyces cerevisiae. Crit. Rev. Biotechnol. 2017, 37, 974–989. [Google Scholar] [CrossRef]
- Lian, J.Z.; Si, T.; Nair, N.U.; Zhao, H.M. Design and construction of acetyl-CoA overproducing Saccharomyces cerevisiae strains. Metab. Eng. 2014, 24, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Krivoruchko, A.; Amatriain, C.S.; Chen, Y.; Siewers, V.; Nielsen, J. Improving biobutanol production in engineered Saccharomyces cerevisiae by manipulation of acetyl-CoA metabolism. J. Ind. Microbiol. Biotechnol. 2013, 40, 1051–1056. [Google Scholar] [CrossRef] [PubMed]
- Scalcinati, G.; Knuf, C.; Partow, S.; Chen, Y.; Maury, J.; Schalk, M.; Daviet, L.; Nielsen, J.; Siewers, V. Dynamic control of gene expression in Saccharomyces cerevisiae engineered for the production of plant sesquitepene α-santalene in a fed-batch mode. Metab. Eng. 2012, 14, 91–103. [Google Scholar] [CrossRef] [PubMed]
- Idnurm, A.; Giles, S.S.; Perfect, J.R. Peroxisome function regulates growth on glucose in the basidiomycete fungus Cryptococcus neoformans. Eukaryot. Cell 2007, 6, 60–72. [Google Scholar] [CrossRef]
- Vitolo, M.; Duranti, M.A.; Pellegrim, M.B. Effect of pH, aeration and sucrose feeding on the invertase activity of intact S. cerevisiae cells grown in sugarcane blackstrap molasses. J. Ind. Microbiol. 1995, 15, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Bohlmann, J.; Crock, J.; Jetter, R.; Croteau, R. Terpenoid-based defenses in conifers: cDNA cloning, characterization, and functional expression of wound-inducible (E)-α-bisabolene synthase from grand fir (Abies grandis). Proc. Natl. Acad. Sci. USA 1998, 95, 6756–6761. [Google Scholar] [CrossRef]
- Huber, D.W.; Philippe, R.N.; Godard, K.A.; Sturrock, R.N.; Bohlmann, J. Characterization of four terpene synthase cDNAs from methyl jasmonate-induced Douglas-fir, Pseudotsuga menziesii. Phytochemistry 2005, 66, 1427–1439. [Google Scholar] [CrossRef]
- Fujisawa, M.; Harada, H.; Kenmoku, H.; Mizutani, S.; Misawa, N. Cloning and characterization of a novel gene that encodes (S)-beta-bisabolene synthase from ginger, Zingiber officinale. Planta 2010, 232, 121–130. [Google Scholar] [CrossRef]
- Parveen, I.; Wang, M.; Zhao, J.; Chittiboyina, A.G.; Tabanca, N.; Ali, A.; Baerson, S.R.; Techen, N.; Chappell, J.; Khan, I.A.; et al. Investigating sesquiterpene biosynthesis in Ginkgo biloba: Molecular cloning and functional characterization of (E,E)-farnesol and alpha-bisabolene synthases. Plant Mol. Biol. 2015, 89, 451–462. [Google Scholar] [CrossRef]
- Srivastava, P.L.; Daramwar, P.P.; Krithika, R.; Pandreka, A.; Shankar, S.S.; Thulasiram, H.V. functional characterization of novel sesquiterpene synthases from indian sandalwood, Santalum album. Sci. Rep. 2015, 5, 10095. [Google Scholar] [CrossRef]
- Ro, D.K.; Ehlting, J.; Keeling, C.I.; Lin, R.; Mattheus, N.; Bohlmann, J. Microarray expression profiling and functional characterization of AtTPS genes_ duplicated Arabidopsis thaliana sesquiterpene synthase genes. Arch. Biochem. Biophys. 2006, 448, 104–116. [Google Scholar] [CrossRef]
- Aschenbrenner, A.K.; Kwon, M.; Conrad, J.; Ro, D.; Spring, O. Identification and characterization of two bisabolene synthases from linear glandular trichomes of sunflower (Helianthus annuus L., Asteraceae). Phytochemistry 2016, 124, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Lancaster, J.; Lehner, B.; Khrimian, A.; Muchlinski, A.; Luck, K.; Köllner, T.G.; Weber, D.C.; Gundersen-Rindal, D.; Tholl, D. An IDS-Type sesquiterpene synthase produces the pheromone precursor (Z)-α-Bisabolene in Nezara viridula. J. Chem. Ecol. 2019, 45, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Niu, M.; Silva, J.T.; Zhang, Y.Y.; Yuan, Y.F.; Jia, Y.; Xiao, Y.; Li, Y.; Fang, L.; Zeng, S.; et al. Identification and functional characterization of three new terpene synthase genes involved in chemical defense and abiotic stresses in Santalum album. BMC Plant Biol. 2019, 19, 115. [Google Scholar] [CrossRef] [PubMed]
- Muchlinski, A.; Chen, X.L.; Lovell, J.T.; Köllner, T.G.; Pelot, K.A.; Zerbe, P.; Ruggiero, M.; Callaway, L.; Laliberte, S.; Chen, F.; et al. Biosynthesis and emission of stress-induced volatile terpenes in roots and leaves of switchgrass (Panicum virgatum L.). Front Plant Sci. 2019, 10, 1144. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.P.; Wang, J.; Wang, Z.B.; Zhang, Y.M.; Shi, S.B.; Nielsen, J.; Liu, Z.H. A gRNA-tRNA array for CRISPR-Cas9 based rapid multiplexed genome editing in Saccharomyces cerevisiae. Nat. Commun. 2019, 5, 1053. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Yao, G.; Wan, X.; Wang, F.; Han, P.; Bao, S.; Wang, K.; Song, T.; Jiang, H. Highly Efficient Biosynthesis of γ-Bisabolene with a New Sesquiterpene Synthase AcTPS5 by Dual Cytoplasmic-Peroxisomal Engineering in Saccharomyces cerevisiae. Fermentation 2023, 9, 779. https://doi.org/10.3390/fermentation9090779
Liu J, Yao G, Wan X, Wang F, Han P, Bao S, Wang K, Song T, Jiang H. Highly Efficient Biosynthesis of γ-Bisabolene with a New Sesquiterpene Synthase AcTPS5 by Dual Cytoplasmic-Peroxisomal Engineering in Saccharomyces cerevisiae. Fermentation. 2023; 9(9):779. https://doi.org/10.3390/fermentation9090779
Chicago/Turabian StyleLiu, Jiajia, Ge Yao, Xiukun Wan, Fuli Wang, Penggang Han, Shaoheng Bao, Kang Wang, Tianyu Song, and Hui Jiang. 2023. "Highly Efficient Biosynthesis of γ-Bisabolene with a New Sesquiterpene Synthase AcTPS5 by Dual Cytoplasmic-Peroxisomal Engineering in Saccharomyces cerevisiae" Fermentation 9, no. 9: 779. https://doi.org/10.3390/fermentation9090779
APA StyleLiu, J., Yao, G., Wan, X., Wang, F., Han, P., Bao, S., Wang, K., Song, T., & Jiang, H. (2023). Highly Efficient Biosynthesis of γ-Bisabolene with a New Sesquiterpene Synthase AcTPS5 by Dual Cytoplasmic-Peroxisomal Engineering in Saccharomyces cerevisiae. Fermentation, 9(9), 779. https://doi.org/10.3390/fermentation9090779