Abstract
The effects of additives on perennial ryegrass (Lolium perenne L., LP) silage’s metabolites, microbial diversity and microbial metabolic pathways have been less studied in karst areas. This experiment sought to ascertain the impact of formic acid (F) and lactic acid bacteria (LAB) preparations introduced in karst locations on the fermentation products of LP silage, the makeup of bacterial population and the projected functional characteristics. The experiment proceeded as follows: (1) for the CK treatment, 5 mL kg−1 fresh weight (FW) of distilled water was added; (2) for the F treatment (88%), 5 mL kg−1 FW of formic acid was added; (3) for the L treatment, Lactobacillus plantarum was mixed with Lactobacillus brucei at 2 × 107 cfu/g FW. For 7, 15 and 45 days, the silage samples were kept at room temperature (20–25 °C). On day 45, the amount of lactic acid (LA) in the silage samples was substantially higher (p < 0.05) in the F (6.56% DM) and L (6.94% DM) treatments than in the CK treatment (4.47% DM), and the F treatment also had significantly lower pH and NH3-N contents than the CK and L treatments. The concentration of lactic acid (LA) was significantly higher (p < 0.05) in the F (6.56% DM) and L (6.94% DM) treatments than in the CK treatment (4.47% DM). On day 45, the dominant genera for the F and L treatments were Lactiplantibacillus (28.78% and 20.34%), Lentilactobacillus (18.85% and 12.67%) and Secundilacillus (5.01% and 13.25%), while Hafnia-Obesumbacterium (16.94%) had a higher abundance in the CK treatment. The F and L treatments promoted microbial metabolic pathways such as “metabolism”, “genetic information processing” and “organismal systems”. They reduced other microbial metabolic pathways such as “membrane transport”, “signal transduction” and “ABC transport”. In summary, F and L can enhance the quality of LP silage in karst areas by improving the structure and function of microbial communities.
1. Introduction
Karst areas have large areas of exposed bedrock, fragile ecological environments, serious soil erosion and rocky desertification [1]. Rocky desertification (mild or worse) has occurred in approximately 31% of karst areas in southern China [2]. In addition, in karst areas of southern China, green hay is not easy to make and preserve due to humid air, rain and heat at the same time. Moreover, the rugged terrain of karst areas, the small number of people and the small area of available grassland directly affect the development of grass-fed livestock farming [3]. By creating high-quality, long-lasting silage, farmers attempt to boost animal production and maintain ruminant performance when feed is in limited supply [4]. The foundation of silage, according to Keshri et al.’s [5] research report, is that lactic acid bacteria (LAB) use soluble carbohydrates (WSC) in the raw materials for silage as a fermentation substrate to produce lactic acid (LA), which creates an acidic environment, inhibits microbial growth and preserves nutrients in feed for a long time. To address the issue of an imbalanced seasonal fodder supply and a lack of winter and spring feed for livestock development in karst areas, high-quality silage research and development are important [6].
Perennial ryegrass is a perennial herb of the genus of ryegrass. It has the advantages of high yield, strong tillering ability, fast growth rate and multiple cuttings [7,8,9]. Wang et al. [10] showed that undesired fermentation may result when the level of epiphytic LAB in silage feedstocks is below 105 cfu/g. Some studies have shown that the number of attached LAB of LP is small, which is lower than the number of attached LAB required to produce high-quality forage (105 cfu/g). Additives are commonly employed to improve silage performance and preserve nutrients; formic acid (F) and lactic acid (L) are two common additives that can enhance silage fermentation qualities and extend silage storage time in different ways. Muck et al. [11] showed that F enhances silage quality by direct acidification and inhibition of undesirable microbial activities, while L enhances silage quality by increasing the amount of LAB in the feed to accelerate fermentation. Homotypic and heterotypic fermenting lactic acid bacteria regulate the fermentation of silage in different ways. Lactobacillus plantarum belongs to homotypic lactic acid bacteria, which rapidly reduces environmental pH and inhibits the growth and reproduction of microorganisms, such as yeasts, molds and Clostridium perfringens, in the pre-fermentation stage of silage, thus improving the fermentation quality of silage [12]. Lactobacillus buchneri is the dominant strain in specialized heterotypic silage additives, which improves the aerobic stability of silage by converting lactic acid to acetic acid in the silage fermentation system and inhibiting the activity of yeasts, such as Z. bailii and C. tropicalis, and the reproduction of microorganisms, such as Pantoea and molds [13]. The combination of Lactobacillus plantarum and Lactobacillus buchneri can effectively promote the whole silage fermentation process [14]. In our conclusion, we predicted that the addition of L and F might alter the microbial composition of LP and enhance its silage qualities.
We are aware that previous studies on the addition of F to LP silage have mainly focused on the effects of F on silage fermentation quality, in vitro rumen digestion and protease activity [15,16,17]. The main subjects of studies on adding L to LP silage are the effects of L on the bacterial colony composition and fermentation characteristics of LP silage [18]. Silage undergoes an anaerobic fermentation process driven by microorganisms. There are few studies on the effects of additives on microbial community structure and metabolic pathways of LP silage in karst areas; thus, this study focuses on the effects of additives on microbial community structure and metabolic pathways. We analyzed the effects of various additives on the fermentation qualities, bacterial community dynamics and functional properties of LP silage, and we provided some insights for production of high-quality LP silage in karst areas through high-throughput sequencing and other research methods.
2. Materials and Methods
2.1. Silage Preparation
LP was taken from Guanling County, Guiyang City, Guizhou Province, on 11 April 2021. LP was in the heading stage and the land was not fertilized. The experiment proceeded as follows: (1) for CK, 5 mL kg−1 fresh weight (FW) of distilled water was added; (2) for F (88%), 5 mL kg−1 FW of formic acid was added; (3) for L, Lactobacillus plantarum was mixed with Lactobacillus brucei (Lactobacillus plantarum and Lactobacillus buchneri were produced by Xi‘an Polysource Biotechnology Co., Ltd., (Xi’an, China) and preserved in the China General Microbiological Culture Collection Center under the accession numbers HH-LP56 (Lactobacillus plantarum), JY-16LE8 (Lactobacillus buchneri)), at 2 × 107 cfu/g FW. Each part was divided into 9 parts and placed in bags (each bag was approximately 300 g), and then a vacuum packaging machine was used to vacuum seal the bags. A total of 27 packs (3 treatments × 3 silage periods × 3 repetitions) were kept at room temperature (20–25 °C). The bags were opened on the 7th, 15th and 45th days of silage, with three vacuum bags opened for each treatment.
2.2. Analysis of Silage Microbial Populations, Chemical Composition and Fermentation Quality
To determine the microbial populations in the fresh fodder, a plate count approach was adopted [19]. The silage bags were handled after putting on sterile gloves, and then each sample was homogeneously mixed in a clean plastic container. For one hour, ultraviolet light was shone on the plastic containers. Ninety milliliters of sterilized brine (8.5 g/L NaCl) was added to 10 g of samples that were obtained from each treatment. On deMan, Rogosa–Sharpe, mauve bile, and Rose Bengal agars, LAB, coliforms, yeasts and molds were counted.
We calculated the dry matter (DM) content by drying 100 g of each sample at 65 °C. The amount of CP was measured using the Kjeldahl nitrogen determination method [20]. The contents of acid detergent fiber (ADF) and neutral detergent fiber (NDF) were determined as described by Van Soest et al. [21]. The concentration of soluble sugar (WSC) was measured using anthone colorimetry [22].
Ninety milliliters of distilled water was mixed with 10 g of each sample, and the mixture was kept at 4 degrees Celsius for 24 h before being filtered through four layers of gauze. The pH value was determined using a pH meter. According to Jia et al. [23], high-performance liquid chromatography (HPLC) was used to measure the amounts of LA, acetic acid (AA), propionic acid (PA) and butyric acid (BA). The amount of ammonia nitrogen (NH3-N) was measured using the phenol-hypochlorite reaction [24].
2.3. Microbial Community Analysis
The silage samples were processed to extract microbial DNA using the Wang et al. [19] technique. As soon as possible, Beijing Biomarker Technology Co., Ltd. (Beijing, China) received all samples of microbial DNA for PCR amplification and bioinformatics analysis. Using a forward primer (50-ACTCCTACGGGAGGCAGCA-30) and a reverse primer (50-GGACTACHVGGGTWTCTAAT-30), the V3–V4 region of 16S rDNA was amplified. The operational taxonomic units (OTUs) were grouped together using the QIIME (v1.8.0) software with a 97% similarity rate. Utilizing the MOTHUR software(v.1.41.1), alpha diversity was calculated and included the OTU, Shannon, Chao1, Sobs, Simpson, Ace, coverage and Shannon indices. Community phylogenetic surveys reconstructed using Tax4Fun (version 0.3.1) were used to forecast bacterial functions in the Kyoto Encyclopedia of Genes and Genomes (KEGG) database, as described by Asshauer et al. [25].
2.4. Statistical Analysis
The differences between the treatments were examined using the Statistical Package for Social Sciences (SPSS, version 26.0, SPSS Inc., Chicago, IL, USA). To analyze the fermentation byproducts, chemical composition and microbial populations, a two-factor ANOVA was conducted. The relative abundance of each KEGG pathway was analyzed using a one-way ANOVA. The Duncan multiple range method was used to compare significance. p < 0.05 was considered as indicating significant difference. All graphs were created using GraphPad Prism (version 8, IBM, Armonk, NY, USA).
3. Results and Discussion
3.1. Chemical Composition and Microbial Populations of Fresh Perennial Ryegrass
Table 1 displays the chemical make-up and microbiological populations of the fresh material. Fresh LP had a DM content of 30.60% of FM in the current study. The crude protein content of fresh LP (8.22% DM) was lower than that reported in a previous study (10.46% DM) [26]. In the current study, fresh LP had lower NDF (47.43% DM) and ADF (24.22% DM) contents than those in previous studies [26]. This needs to be confirmed further; however, the results might be related to the moisture level of the raw material during harvest, as well as the climate and soil texture of the growing area. Fresh LP met the criteria for an ideal WSC concentration over 5% DM for raw materials with a WSC content of 9.59% DM, which was comparable to fresh ryegrass samples [27,28]. Silage undergoes an anaerobic fermentation process that is primarily driven by LAB, and high-quality silage raw materials should contain more epiphytic LAB than 105 cfu/g FM. The fresh LP in this investigation had a greater yeast concentration (103.31 cfu/g FM) and less L attachment (104.97 cfu/g FM). Consequently, silage additives are required to raise the standard of LP fermentation.

Table 1.
Chemical compositions and microbial counts of perennial ryegrass prior to ensiling.
3.2. Effect of Additives on Chemical Composition
Table 2 displays the LP chemical composition. Throughout the silage period, the DM content of each treatment gradually decreased. According to Oladosu et al. [29], microbial substrate metabolism might be responsible for this decline. In line with the findings of this study, He et al. [30] demonstrated that a drop in DM occurred primarily during the pre-fermentation period, with a considerable accumulation of LA in the late fermentation period and a fall in pH, which inhibited microbial activity. During silage, the crude protein (CP) content gradually decreased in the CK treatment, and the CP content decreased and then increased in the L and F treatments. At 45 d of silage, the CP content in the L and F treatments was significantly higher than that in the CK treatment (p < 0.05). This is inconsistent with the results of previous studies [5], which may be attributed to the reduction in CP content due to aerobic metabolism of microbial substrates during the pre-fermentation period, the addition of L and F treatments to inhibit microbial reproductive activity, and the formation of bacterial proteins after microbial death to increase CP content in silage. The WSC content of each treatment gradually dropped during the ensiling period. The F treatment had a greater WSC level than the L and CK treatments (p < 0.05), and the WSC concentration of all treatments was significantly lower at 45 days of silage than it was at 7 days. This might be because F prevented harmful microbes, such as clostridia, from consuming WSC and because the F group had a lower pH (Table 3) and fewer harmful organisms (Figure 1B), which is in line with previous findings [31].
Table 2.
Effects of additives on the dynamics of chemical compositions in perennial ryegrass silage during ensiling.
Table 3.
Effects of additives on the dynamics of fermentation quality in perennial ryegrass during ensiling.
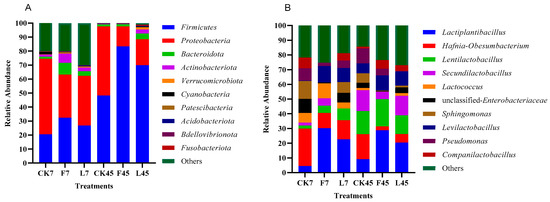
Figure 1.
The relative abundance of bacteria at the phylum (A) and genus (B) levels in perennial ryegrass silage treated with different additives. CK, without additives; F, formic acid; L, Lactiplantibacillus plantarum combined with Lactiplantibacillus buchneri; 7, 7 days of ensiling; 45, 45 days of ensiling.
3.3. Additives’ Impact on Fermentation Properties
Table 3 displays the results of the fermentation of the LP silage samples. The pH of silage is considered an important indicator of the interaction between soluble substrate effectiveness, metabolism and undesirable microbial protein hydrolysis activity, and is one of the most important indicators to evaluate the long-term preservation of silage. A silage’s pH should be less than 4.2 to effectively suppress the growth of dangerous bacteria and enhance the fermentation process [30,32]. At 15 days of silage fermentation, all treatments’ LA contents were significantly higher (p < 0.05); the L treatment’s LA content was significantly higher than the CK treatment’s and the F treatment’s (p < 0.05) contents, and the L treatment’ and the F treatment’s LA contents were significantly higher than the CK treatment at 45 days (p < 0.05). Each treatment’s pH value gradually declined throughout the ensiling process, and the pH value of the F treatment was below 4.0, which was substantially lower than the pH values of the other treatments (p < 0.05). This agrees with Wang et al.’s [33] conclusion. In comparison to the L and CK treatments, the F treatment had a greater proportion of lactic acid bacteria at 45 days of silage, which resulted in a higher lactic acid concentration and a lower pH. Small quantities of AA (<3.00% DM) and PA (<0.30% DM) were discovered in all treatments, but BA was not discovered. According to previous research, the presence of PA and BA is undesirable because their creation uses up metabolic energy [5] and because AA concentrations below 3% DM inhibit aerobic degeneration and enhance energy recovery [34]. Throughout the ensiling process, the NH3-N content gradually increased in each treatment, but the F and L treatments had significantly lower NH3-N contents than the CK treatment. Thus, it is evident that the addition of F and L can effectively reduce the pH and protein loss of LP silage.
3.4. Effect of Additives on Microbial Communities
As shown in Table 4, after seven days of silage fermentation, the number of lactic acid bacteria in each treatment increased compared to the original sample, while the numbers of coliforms and yeasts were lower than 102 cfu/g. The number of lactic acid bacteria in the F treatment was significantly higher than that in the CK and L treatments, while the number in the CK treatment was significantly lower than in the L treatment (p < 0.05). This might be because the addition of formic acid inhibited the reproduction of undesirable microorganisms and reduced the competition between lactic acid bacteria and undesirable microorganisms so that lactic acid bacteria obtained more fermentation substrate, and then proliferated. In addition, the number of lactic acid bacteria in each treatment increased significantly at 45 days of silage (p < 0.05). This is consistent with previous research results. With the formation of an acidic anaerobic environment, lactic acid bacteria can continue to multiply while most undesirable microorganisms are inhibited [35].
Table 4.
Microbial populations after silage of perennial ryegrass.
Table 5 displays the alpha diversity of the silage samples. The kind and quantity of microorganisms present during the fermentation of silage has a direct impact on fermentation quality [36]. The coverage values of the perennial ryegrass silage samples ranged from 0.997~0.999, indicating that the microbial populations in the silage samples had been reliably identified and could adequately reflect the diversity of bacterial communities in the samples [37]. The fact that the internal environment of the silage samples did not reach a strictly anaerobic state at the beginning of fermentation, but rather an acidic anaerobic environment was essentially developed after 45 days of silage, might account for the decrease in diversity among the treatments. The Simpson’s and Shannon’s indices were lower in the F treatment after 45 days of silage than in the L and CK treatments. The results at the microbial genus level suggest that this might be because the F treatment inhibited the growth of unfavorable microorganisms [11], leading to a bacterial community that was rather simple and consistent with the results of Du et al. [38]. This demonstrates how L and F can change the microbial community structure and composition of LP silage.
Table 5.
Effects of additives on the diversity and richness of bacterial microbiota in the mixed silage of alfalfa and perennial ryegrass during ensiling.
Using high-throughput sequencing based on the 16S ribosomal DNA gene, the dynamics of the bacterial community during fermentation were evaluated. The relative density of the bacterial populations during the fermentation of LP silage is shown in Figure 1. The two primary bacterial phyla found in the silage are Firmicutes and Proteobacteria [39]. Proteobacteria speed up the nitrogen and carbon cycles and breakdown organic materials [40]. Firmicutes can produce a wide range of enzymes, including proteases, cellulases and other extracellular enzymes, and their acid hydrolysis function is vital in a variety of anaerobic settings [41]. In terms of phyla (Figure 1A), the phyla Firmicutes and Proteobacteria (20.46% and 50.01%), F (32.34% and 30.85%) and L (26.80% and 35.52%) were found in the CK treatment, accounting for more than 60.0% of the total bacterial flora. This is in line with a study published by Wang et al. [42]. The relative abundance of Firmicutes rose to varying degrees at 45 days during silage fermentation. The relative abundance of Firmicutes was higher in the F treatment (83.39%) and L treatment (69.93%) than in the CK treatment (48.19%). Both the F treatment (14.29%) and the L treatment (18.51%) showed a decline in the relative abundance of Proteobacteria. This could be because F and L can produce anaerobic conditions and an acidic environment more quickly, which helps Firmicutes grow and reproduce during anaerobic fermentation [43].
According to the research by Duniere et al. [6], LAB have the ability to exploit soluble carbohydrates in a fermentation substrate to convert glucose to lactic acid and produce significant volumes of LA, which can be used to reduce the pH of silage and suppress unwanted microbes such as Clostridium. After seven days of silage, at the genus level (Figure 2B), Hafnia-Obesumbacterium was detected in the CK treatment (25.50%), and Lactiplantibacterium was found in the F (30.17%) and L (22.57%) treatments as the dominant bacteria. This might be a result of F and L inhibiting the action of harmful microbes. According to previous reports, adding F and L to maize and sorghum can effectively reduce the number of undesirable microorganisms in their silage and raise the quality of silage [44]. Wang et al. [38] found the presence of Hafnia-Obesumbacterium in Enterobacterium in Italian ryegrass silage, and the presence of Hafnia-Obesumbacterium was found to accelerate the hydrolytic activity of proteins during silage. In all treatments, the relative abundance of lactic acid bacteria grew significantly after 45 days of silage, with Hafnia-Obesumbacterium in the CK treatment (16.94%) having a much greater relative abundance than in the F (2.60%) and L (5.92%) treatments. The dominant bacterial genera of the F and L treatments were Lactobacillus (28.78% and 20.34%) and Subbacillus (18.85% and 13.25%). F and L could efficiently decrease the relative abundance of undesirable microorganisms such as Hafnia-Obesumbacterium in LP silage, increase LA content and lower pH, thus improving LP silage quality. These effects were observed when combined with a thorough analysis of LP silage chemical composition (Table 2) and fermentation index (Table 3).
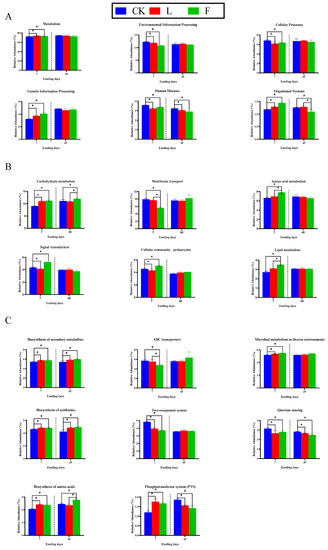
Figure 2.
* means that different treatments differ significantly in the same silage period (p < 0.05); Bar graphs showing 16S rRNA gene-predicted functional profiles on level 1 metabolic pathways (A), level 2 metabolic pathways (B) and level 3 metabolic pathways (C) of silage of perennial ryegrass treated with different additives for 7 and 45 days obtained using Tax4Fun. CK, without additives; F, formic acid; L, Lactiplantibacillus plantarum combined with Lactiplantibacillus buchneri; 7, 7 days of ensiling; 45, 45 days of ensiling.
3.5. The Effect of Additives on the Pathways
A method for assessing the impact of flora structure changes on silage quality is provided by using the KEGG to investigate changes in flora structure at a higher and whole-genome level. This study examined the effects of various additive amounts on the silage of LP using the KEGG technology based on Tax4Fun.
Figure 2A depicts the metabolic route at the first pathway level. At seven days of silage, the L and F treatments had significantly higher gene abundances than the CK treatment for “organism system”, “genetic information processing” and “metabolism” (p < 0.05). In comparison to the CK treatment, genes involved in “environmental information processing”, “cellular processes,” and “human diseases” were significantly less common (p < 0.05). A stabilized inner environment might have developed at a later stage of fermentation because there was no discernible difference in “metabolism” between the treatments at 45 days of silage (p > 0.05). The aforementioned findings demonstrated that whereas L might improve the fermentation quality of LP mostly by boosting the population of advantageous bacteria such as LAB, F might do so by limiting the “membrane transport” and “signal transduction” of unwanted bacteria.
Figure 2B depicts the second horizontal metabolic pathway. At days 7 and 45 of silage, the relative abundance of “carbohydrate metabolism” was noticeably higher in the L and F treatments than in CK treatment (p < 0.05). This is in line with Table 3′s greater LAB and LA concentrations, which show that L and F improved the microbial metabolism of WSC. Compared to the L and CK treatments, the relative abundance of genes involved in “membrane transport” was significantly lower in the F treatment at seven days of silage (p < 0.05), and the relative abundance of genes involved in “amino acid metabolism” was significantly higher in the F treatment (p < 0.05). When silage was seven days old, the F treatment had a larger relative abundance of “signal transduction” than the CK treatment, and the L treatment had a lower relative abundance than the CK treatment. The fact that F suppressed the metabolic activity of harmful microbes in silage might have something to do with this. This is in line with previous studies where it was found that transit proteins were more abundant in untreated silage, which might be related to the symbiotic connection of bacterial communities in silage [43].
Figure 2C displays the modifications to metabolic pathways at the third pathway level. After seven days of ensiling, the abundances of genes involved in the “biosynthesis of secondary metabolites”, “microbial metabolism in diverse environments”, “biosynthesis of amino acids”, “biosynthesis of antibiotics” and “phosphotransferase system (PTS)” were significantly higher in the L and F treatments than in the CK treatment (p < 0.05), while the abundances of genes involved in the “two-component system” and “quorum sensing” were significantly lower in the CK treatment (p < 0.05). At seven days of silage, the F treatment had a significantly lower relative abundance of “ABC transport” than the L and CK treatments (p < 0.05). The relative abundances of genes involved in the “biosynthesis of secondary metabolites” and “biosynthesis of antibiotics” were substantially greater in the L and F treatments than in the CK treatment at 45 days after ensiling (p < 0.05). In comparison to the CK treatment, the relative abundances of the “phosphotransferase system (PTS)” and “quorum sensing” genes were significantly lower in the L and F treatments (p < 0.05). In comparison to the L and CK treatments, the relative abundance of genes involved in the “biosynthesis of amino acids” was significantly higher in the F treatment (p < 0.05). These findings suggested that pathogenic bacterial movement via the membrane system is constrained by the acidic environment created by L and F, which impacts the quality of LP silage.
4. Conclusions
In this study, it was discovered that F and L could enhance the structure of microbial community and the metabolic pathway, leading to an improvement in perennial ryegrass silage quality in a karst region. F and L can promote the growth of good bacteria (Lactiplantibacillus, Lentilactobacillus and Secundilacillus), which increase the LA content of perennial ryegrass silage and lower the pH and ammonia nitrogen content while inhibiting the growth of harmful bacteria (Hafnia-Obesumbacterium). According to the results of the functional prediction, F and L encourage metabolic pathways in microorganisms, such as “metabolism”, “genetic information processing” and “organismal systems”, while inhibiting transduction pathways such as “membrane transport”, “signal transduction” and “ABC transports”. This demonstrates that the L treatment increased the fermentation quality of perennial ryegrass by accelerating the growth rate and metabolic level of helpful bacteria such as LAB strains. The F treatment largely reduced the “membrane transport” and “signaling transduction” of unwanted bacteria.
Author Contributions
Q.C. and C.C. designed the experiments and revised the manuscript. M.L., X.F. and Y.L. performed the experiments. Q.C., Y.L. and Y.C. wrote the manuscript. M.L. and Q.C. performed the data analysis. P.L., H.S., Y.X., Y.Z., C.W. and R.D. contributed to the grammar and language revision of the paper and contributed to this experiment. All authors have read and agreed to the published version of the manuscript.
Funding
This project was supported by the Guizhou Provincial Science and Technology Project (Qiankehe Chengguo [2022]005), the National Key Research and Development Subject (2021YFD1300302), the Guizhou Provincial Science and Technology Project (Qiankehe Jichu-ZK [2023]120), and the Guizhou Provincial Science and Technology Project (QKHPTRC-CXTD [2022]011).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Our raw sequence data are available under the NCBI project PRJNA778048 with the accession number SRP344966.
Acknowledgments
We thank the Biomarker Technologies Company (Beijing, China) for providing technical support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chang, J.J.; Zhu, J.X.; Xu, L.; Su, H.X.; Gao, Y.; Cai, X.L.; Peng, T.; Wen, X.F.; Zhang, J.J.; He, N.P. Rational land-use types in the karst regions of China: Insights from soil organic matter composition and stability. Catena 2018, 160, 345–353. [Google Scholar] [CrossRef]
- He, K.; Jia, Y.; Wang, F.; Lu, Y. Overview of karst geo-environments and karst water resources in north and south China. Environ. Earth Sci. 2011, 64, 1865–1873. [Google Scholar]
- Song, S.; Xiong, K.; Chi, Y.; Shen, X.; Guo, T.; Lu, N. Research Progress and Prospect of Grassland Establishment and Ecological Animal Husbandry in the Karst Rocky Desertification Area. Fresenius Environ. Bull. 2018, 27, 7017–7030. [Google Scholar]
- Wilkinson, J.M.; Lee, M.R.F.; Rivero, M.J.; Chamberlain, A.T. Some challenges and opportunities for grazing dairy cows on temperate pastures. Grass Forage Sci. 2020, 75, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Keshri, J.; Chen, Y.; Pinto, R.; Kroupitski, Y.; Weinberg, Z.G.; Sela Saldinger, S. Bacterial dynamics of wheat silage. Front. Microbiol. 2019, 10, 1532. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.; Sun, L.; Chen, C.; Lin, J.; Yang, F.; Cai, Y. Exploring microbial community structure and metabolic gene clusters during silage fermentation of paper mulberry, a high-protein woody plant. Anim. Feed Sci. Technol. 2021, 275, 114766. [Google Scholar] [CrossRef]
- Yu, X.; Pijut, P.M.; Byrne, S.; Asp, T.; Bai, G.; Jiang, Y. Candidate gene association mapping for winter survival and spring regrowth in perennial ryegrass. Plant Sci. 2013, 235, 37–45. [Google Scholar] [CrossRef]
- Fu, Z.; Song, J.; Zhao, J.; Jameson, P.E. Identification and expression of genes associated with the abscission layer controlling seed shattering in Lolium perenne. AoB Plants 2019, 11, ply076. [Google Scholar] [CrossRef]
- Xie, L.; Teng, K.; Tan, P.; Chao, Y.; Li, Y.; Guo, W.; Han, L. PacBio single-molecule long-read sequencing shed new light on the transcripts and splice isoforms of the perennial ryegrass. Mol. Genet. Genomics. 2020, 295, 475–489. [Google Scholar] [CrossRef]
- Wang, S.; Yuan, X.; Dong, Z.; Li, J.; Guo, G.; Bai, Y.; Zhang, J.; Shao, T. Characteristics of isolated lactic acid bacteria and their effects on the silage quality. Asian Australas. J. Anim. Sci. 2017, 30, 819–827. [Google Scholar] [CrossRef] [PubMed]
- Muck, R.E.; Nadeau, E.M.G.; McAllister, T.A.; Contreras-Govea, F.E.; Santos, M.C.; Kung, L., Jr. Silage review: Recent advances and future uses of silage additives. J. Dairy Sci. 2018, 101, 3980–4000. [Google Scholar] [CrossRef]
- Yang, F.; Wang, Y.; Zhao, S.; Wang, Y. Lactobacillus plantarum inoculants delay spoilage of high moisture alfalfa silages by regulating bacterial community composition. Front. Microbiol. 2020, 11, 1989. [Google Scholar] [CrossRef] [PubMed]
- Wambacq, E.; Latré, J.P.; Haesaert, G. The effect of Lactobacillus buchneri inoculation on the aerobic stability and fermentation characteristics of alfalfa-ryegrass, red clover and maize silage. Agr. Food Sci. 2013, 22, 127–136. [Google Scholar] [CrossRef]
- Li, M.; Zhou, S.; Tang, X.; Liao, C.; Li, P.; Xie, Y.X.; Cheng, Q.M.; Chen, C. Effect of co-inoculation of Lactobacillus plantarum and Lentilactobacillus buchneri on aerobic stability and microbial community composition of perennial sorghum silage. Biomass Bioenergy 2023, 173, 106801. [Google Scholar] [CrossRef]
- Yuan, X.; Wen, A.; Desta, S.T.; Dong, Z.; Shao, T. Effects of four short-chain fatty acids or salts on the dynamics of nitrogen transformations and intrinsic protease activity of alfalfa silage. J. Sci. Food Agric. 2017, 97, 2759–2766. [Google Scholar] [CrossRef] [PubMed]
- Xie, H.; Xie, F.; Guo, Y.; Liang, X.; Peng, L.; Li, M.; Tang, Z.H.; Peng, K.P.; Yang, C.J. Fermentation quality, nutritive value and in vitro ruminal digestion of Napier grass, sugarcane top and their mixed silages prepared using lactic acid bacteria and formic acid. Grassl. Sci. 2023, 69, 23–32. [Google Scholar] [CrossRef]
- Lv, J.; Fang, X.; Feng, G.; Zhang, G.; Zhao, C.; Zhang, Y.; Li, Y. Effects of Sodium Formate and Calcium Propionate Additives on the Fermentation Quality and Microbial Community of Wet Brewers Grains after Short-Term Storage. Animals 2020, 10, 1608. [Google Scholar] [CrossRef]
- Wang, S.; Yuan, X.; Dong, Z.; Li, J.; Shao, T. Effect of ensiling corn stover with legume herbages in different proportions on fermentation characteristics, nutritive quality and in vitro digestibility on the Tibetan Plateau. Grassl. Sci. 2017, 63, 236–244. [Google Scholar] [CrossRef]
- Wang, S.; Zhao, J.; Dong, Z.; Li, J.; Kaka, N.A.; Shao, T. Sequencing and microbiota transplantation to determine the role of microbiota on the fermentation type of oat silage. Bioresour. Technol. 2020, 309, 123371. [Google Scholar] [CrossRef]
- Krishnamoorthy, U.; Muscato, T.V.; Sniffen, C.J.; Van Soest, P.J. Nitrogen Fractions in Selected Feedstuffs. J. Dairy Sci. 1982, 65, 217–225. [Google Scholar] [CrossRef]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Thomas, T. An automated procedure for the determination of soluble carbohydrates in herbage. J. Sci. Food Agric. 1977, 28, 639–642. [Google Scholar] [CrossRef]
- Jia, T.; Sun, Z.; Gao, R.; Yu, Z. Lactic acid bacterial inoculant effects on the vitamin content of alfalfa and Chinese leymus silage. Asian Australas. J. Anim. Sci. 2019, 32, 1873–1881. [Google Scholar] [CrossRef] [PubMed]
- Broderick, G.A.; Kang, J.H. Automated simultaneous determination of ammonia and total amino acids in ruminal fluid and in vitro media. J. Dairy Sci. 1980, 63, 64–75. [Google Scholar] [CrossRef] [PubMed]
- Asshauer, K.P.; Wemheuer, B.; Daniel, R.; Meinicke, P. Tax4Fun: Predicting functional profiles from metagenomic 16S rRNA data. Bioinformatics 2015, 31, 2882–2884. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.; Zhang, H.; Gao, Y.; Diao, Q. Dynamic profiles of fermentation characteristics and bacterial community composition of Broussonetia papyrifera ensiled with perennial ryegrass. Bioresour. Technol. 2020, 310, 123396. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Zhang, Y.; Gou, W.; Cheng, Q.; Bai, S.; Cai, Y. Silage fermentation and bacterial community of bur clover, annual ryegrass and their mixtures prepared with microbial inoculant and chemical additive. Anim. Feed Sci. Technol. 2019, 247, 285–293. [Google Scholar] [CrossRef]
- Conaghan, P.; O’Kiely, P.; O’Mara, F.P. Conservation characteristics of wilted perennial ryegrass silage made using biological or chemical additives. J. Dairy Sci. 2010, 93, 628–643. [Google Scholar] [CrossRef]
- Oladosu, Y.; Rafii, M.Y.; Abdullah, N.; Magaji, U.; Hussin, G.; Ramli, A.; Miah, G. Fermentation Quality and Additives: A Case of Rice Straw Silage. Biomed. Res. Int. 2016, 2016, 7985167. [Google Scholar] [CrossRef]
- He, L.; Lv, H.; Xing, Y.; Wang, C.; You, X.; Chen, X.; Chen, X.; Zhang, Q. The nutrients in Moringa oleifera leaf contribute to the improvement of stylo and alfalfa silage: Fermentation, nutrition and bacterial community. Bioresour. Technol. 2020, 301, 122733. [Google Scholar] [CrossRef]
- Ran, Q.; Guan, H.; Li, H.; He, W.; Zhu, R.; Zhang, L.; Huang, Y.; Xu, Y.; Fan, Y. Effect of Formic Acid and Inoculants on Microbial Community and Fermentation Profile of Wilted or Un-Wilted Italian Ryegrass Silages during Ensiling and Aerobic Exposure. Fermentation 2022, 8, 755. [Google Scholar] [CrossRef]
- Yang, L.; Yuan, X.; Li, J.; Dong, Z.; Shao, T. Dynamics of microbial community and fermentation quality during ensiling of sterile and nonsterile alfalfa with or without Lactobacillus plantarum inoculant. Bioresour. Technol. 2019, 275, 280–287. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; He, L.; Xing, Y.; Zhou, W.; Yang, F.; Chen, X.; Zhang, Q. Fermentation quality and microbial community of alfalfa and stylo silage mixed with Moringa oleifera leaves. Bioresour. Technol. 2019, 284, 240–247. [Google Scholar] [CrossRef] [PubMed]
- Ni, K.; Wang, F.; Zhu, B.; Yang, J.; Zhou, G.; Pan, Y.; Tao, Y.; Zhong, J. Effects of lactic acid bacteria and molasses additives on the microbial community and fermentation quality of soybean silage. Bioresour. Technol. 2017, 238, 706–715. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Yin, X.J.; Wang, S.R.; Li, J.F.; Shao, T. Separating the effects of chemical and microbial factors on fermentation quality and bacterial community of Napier grass silage by using gamma-ray irradiation and epiphytic microbiota transplantation. Anim. Feed Sci. Technol. 2021, 280, 115082. [Google Scholar] [CrossRef]
- Guan, H.; Yan, Y.; Li, X.; Li, X.; Shuai, Y.; Feng, G.; Ran, Q.; Cai, Y.; Li, Y.; Zhang, X. Microbial communities and natural fermentation of corn silages prepared with farm bunker-silo in Southwest China. Bioresour. Technol. 2018, 265, 282–290. [Google Scholar] [CrossRef]
- He, L.; Wang, C.; Xing, Y.; Zhou, W.; Pian, R.; Chen, X.; Zhang, Q. Ensiling characteristics, proteolysis and bacterial community of high-moisture corn stalk and stylo silage prepared with Bauhinia variegate flower. Bioresour. Technol. 2020, 296. [Google Scholar] [CrossRef]
- Du, Z.; Lin, Y.; Sun, L.; Yang, F.; Cai, Y. Microbial community structure, co-occurrence network and fermentation characteristics of woody plant silage. J. Sci. Food Agric. 2022, 102, 1193–1204. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, X.Y.; Wang, C.; He, L.W.; Zhou, W.; Yang, F.Y.; Zhang, Q. The bacterial community and fermentation quality of mulberry (Morus alba) leaf silage with or without Lactobacillus casei and sucrose. Bioresour. Technol. 2019, 293, 122059. [Google Scholar] [CrossRef]
- Ma, S.S.; Fang, C.; Sun, X.X.; Han, L.J.; He, X.Q.; Huang, G.Q. Bacterial community succession during pig manure and wheat straw aerobic composting covered with a semi-permeable membrane under slight positive pressure. Bioresour. Technol. 2018, 259, 221–227. [Google Scholar] [CrossRef]
- Wang, S.; Sun, Y.; Zhao, J.; Dong, Z.; Li, J.; Nazar, M.; Shao, T. Assessment of inoculating various epiphytic microbiota on fermentative profile and microbial community dynamics in sterile Italian ryegrass. J. Appl. Microbiol. 2020, 129, 509–520. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; He, L.; Xing, Y.; Zhou, W.; Pian, R.; Yang, F.; Chen, X.; Zhang, Q. Bacterial diversity and fermentation quality of Moringa oleifera leaves silage prepared with lactic acid bacteria inoculants and stored at different temperatures. Bioresour. Technol. 2019, 284, 349–358. [Google Scholar] [CrossRef] [PubMed]
- Keshri, J.; Chen, Y.; Pinto, R.; Kroupitski, Y.; Weinberg, Z.G.; Sela, S. Microbiome dynamics during ensiling of corn with and without Lactobacillus plantarum inoculant. Appl. Microbiol. Biotechnol. 2018, 102, 4025–4037. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.N.; Li, Y.F.; Jeong, E.C.; Kim, H.J.; Kim, J.G. Effects of formic acid and lactic acid bacteria inoculant on main summer crop silages in Korea. J. Anim. Sci. Technol. 2021, 63, 91–103. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).