
Effect of Replacing Fish Meal Using Fermented Soybean Meal on Growth Performance, Intestine Bacterial Diversity, and Key Gene Expression of Largemouth Bass (Micropterus salmoides)


 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation Method of Fermented Soybean Meal
2.2. The Feed Preparation of Different Treatments
2.3. Experiment Process of Breeding Largemouth Bass
2.4. Determination of Nutrient Availability
2.5. Determination Method of Physicochemical Parameters
2.6. Determination Method of Blood Key Biochemical Parameters
2.7. Determination of Amino Acid Content in Perch Muscle, SBM, and FSBM
2.8. Methods of Fish Liver and Intestinal Tissue Sections
2.9. Quantitative Method of Key Gene Expression
2.10. DNA Extraction and 16S rRNA Gene Sequencing of Intestinal Microorganisms
2.11. Detection Methods of Metabolites in Intestine (LC-MS/MS Analysis)
3. Results
3.1. Changes in Soybean Meal Properties before and after Fermentation (Amino Acid Phytic Acid Content)
3.2. Effect of FSBM Instead of Fish Meal on Growth Performance of Largemouth Bass
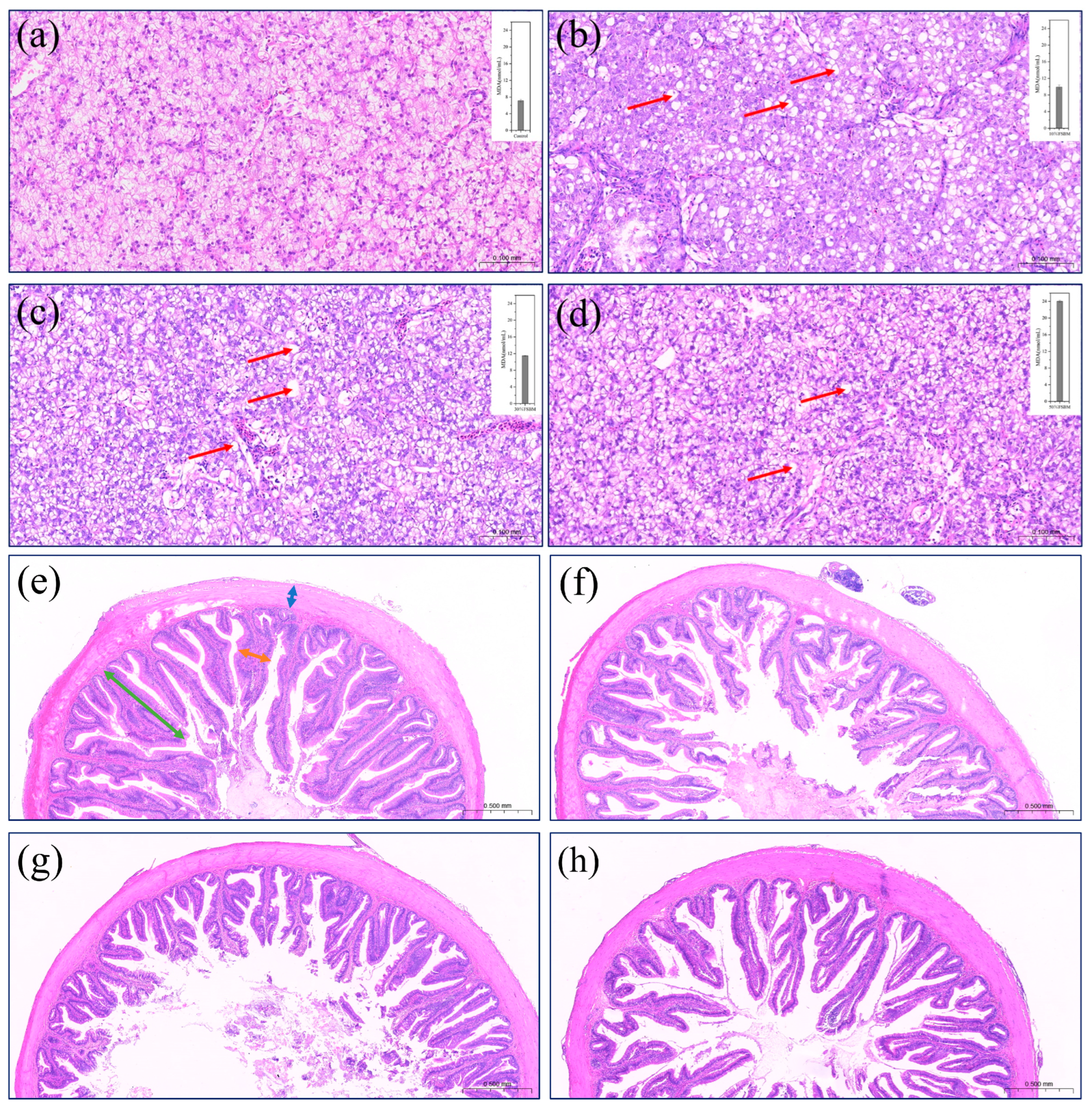
3.3. Effect of FSBM Replacing Fish Meal on the Morphology of Key Organs (Intestine and Liver)
3.4. Effect of FSBM Replacing Fish Meal on Blood Biochemical Parameters
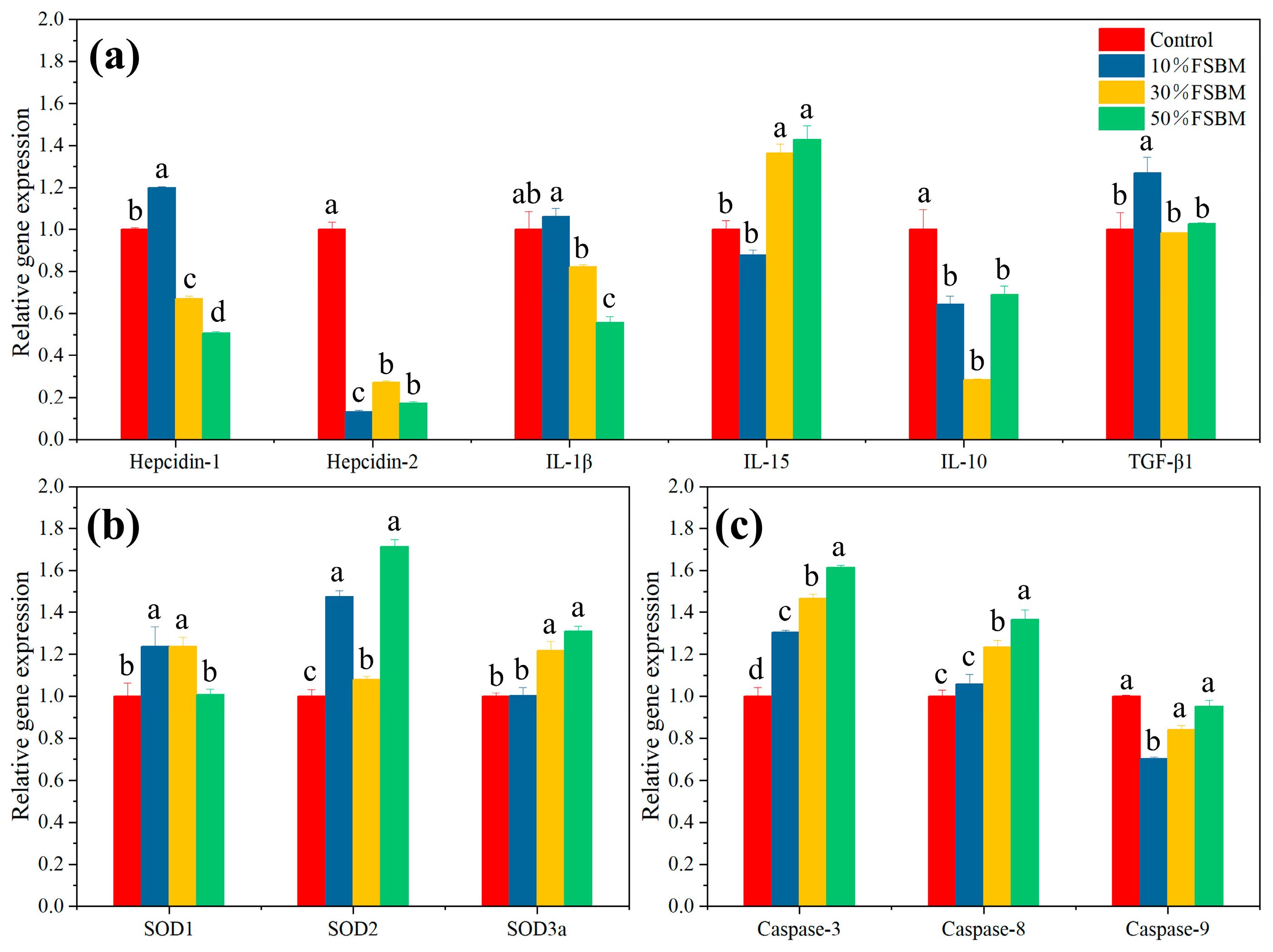
3.5. Effect of FSBM Replacing Fish Meal on Key Gene Expression
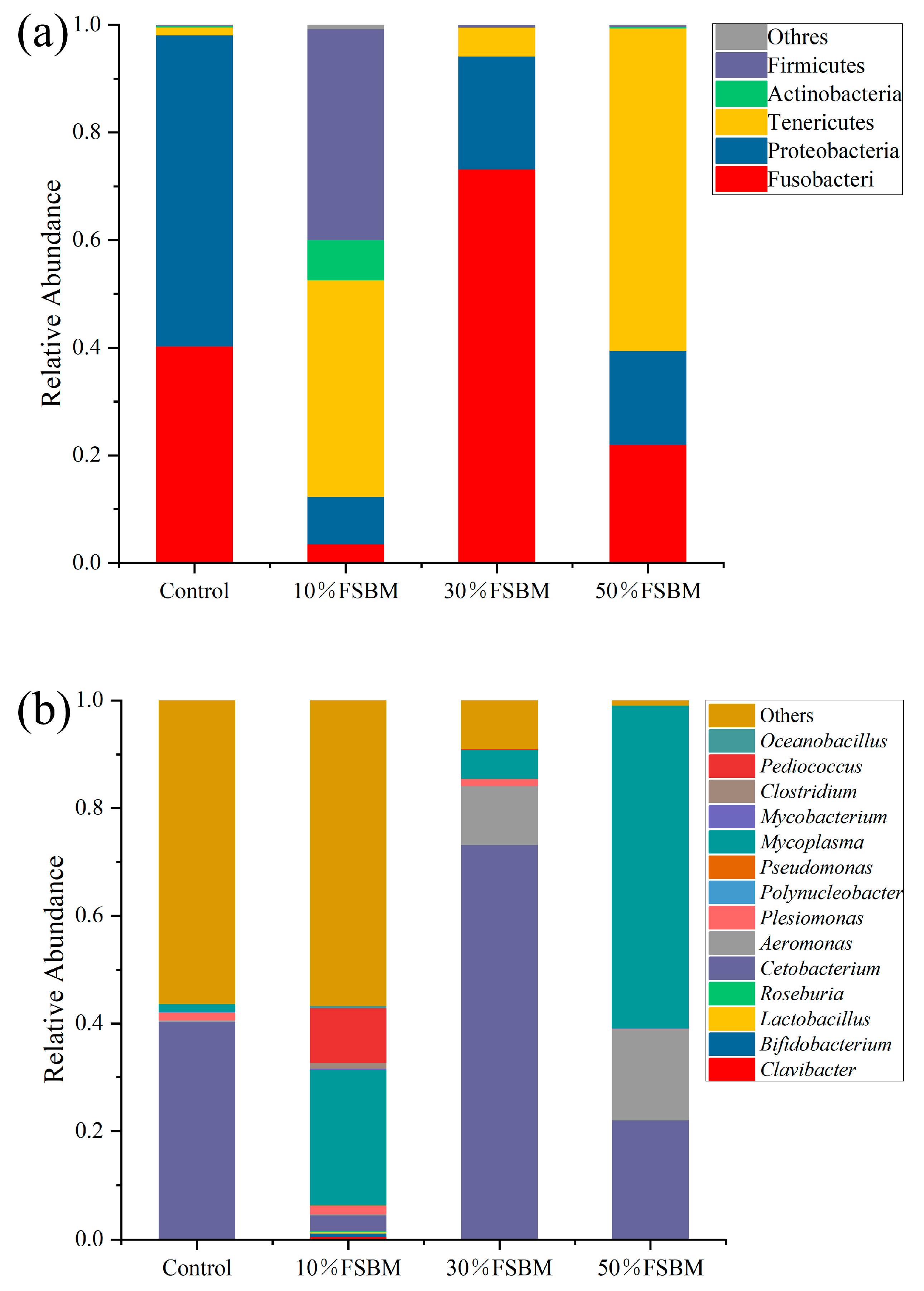
3.6. Effects of Fermented Soybean Meal Replacing Fish Meal on Intestinal Microbial Community Structure
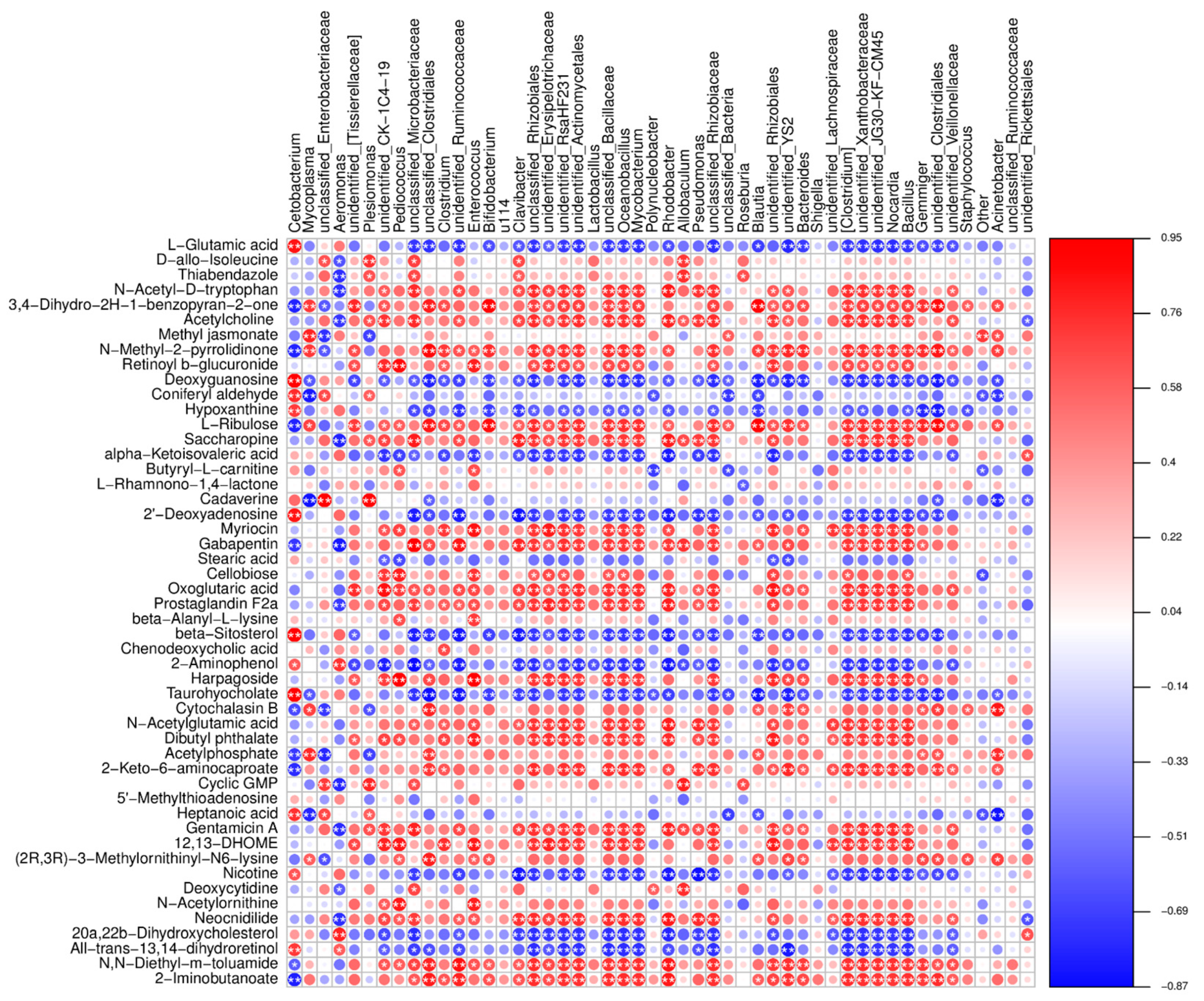
3.7. Effects of Fermented Soybean Meal Replacing Fish Meal on the Relationship between Metabolic Pathways and Metabolites
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lee, S.M.; Azarm, H.M.; Chang, K.H. Effects of dietary inclusion of fermented soybean meal on growth, body composition, antioxidant enzyme activity and disease resistance of rockfish (Sebastes schlegeli). Aquaculture 2016, 459, 110–116. [Google Scholar] [CrossRef]
- Watanabe, T. Strategies for further development of aquatic feeds. Fish. Sci. 2002, 68, 242–252. [Google Scholar] [CrossRef]
- Hassaan, M.S.; Soltanb, M.A.; Abdel-Moezc, A.M. Nutritive value of soybean meal after solid state fermentation with Saccharomyces cerevisiae for Nile tilapia, Oreochromis niloticus. Anim. Feed Sci. Technol. 2015, 201, 89–98. [Google Scholar] [CrossRef]
- Hua, K.; Cobcroft, J.M.; Cole, A.; Condon, K.; Jerry, D.R.; Mangott, A.; Praeger, C.; Vucko, M.J.; Zeng, C.; Zenger, K.; et al. The future of aquatic protein: Implications for protein sources in aquaculture diets. One Earth 2019, 1, 316–329. [Google Scholar] [CrossRef]
- Seong, M.; Lee, S.; Lee, S.; Song, Y.; Bae, J.; Chang, K.; Bai, S.C. The effects of different levels of dietary fermented plant-based protein concentrate on growth, hematology and non-specific immune responses in juvenile olive flounder, Paralichthys olivaceus. Aquaculture 2018, 483, 196–202. [Google Scholar] [CrossRef]
- Drew, M.D.; Borgeson, T.L.; Thiessen, D.L. A review of processing of feed in-gredients to enhance diet digestibility in finfish. Anim. Feed Sci. Technol. 2007, 138, 118–136. [Google Scholar] [CrossRef]
- Lin, Y.H.; Chen, Y.T. Lactobacillus spp. fermented soybean meal partially substitution to fish meal enhances innate immune responses and nutrient digestibility of white shrimp (Litopenaeus vannamei) fed diet with low fish meal. Aquaculture 2022, 548, 737634. [Google Scholar] [CrossRef]
- Refstie, S.; Sahlström, S.; Bråthen, E.; Baeverfjord, G.; Krogdahl, P. Lactic acid fermentation eliminates indigestible carbohydrates and antinutritional factors in soybean meal for Atlantic salmon (Salmo salar). Aquaculture 2005, 246, 331–345. [Google Scholar] [CrossRef]
- Hong, Y.C.; Chu, J.H.; Kirby, R.; Sheen, S.S.; Chien, A. The effects of replacing fish meal protein with a mixture of poultry by-product meal and fermented soybean meal on the growth performance and tissue nutritional composition of Asian seabass (Lates calcarifer). Aquac. Res. 2021, 52, 4105–4115. [Google Scholar] [CrossRef]
- Daniel, N. A review on replacing fish meal in aqua feeds using plant protein sources. Int. J. Fish. Aquat. Stud. 2018, 6, 164–179. [Google Scholar]
- He, M.; Yu, Y.F.; Li, X.Q.; Poolsawat, L.; Yang, P.X.; Bian, Y.H.; Leng, X.J. An evaluation of replacing fish meal with fermented soybean meal in the diets of largemouth bass (Micropterus salmoides): Growth, nutrition utilization and intestinal histology. Aquac. Res. 2020, 51, 4302–4314. [Google Scholar] [CrossRef]
- AOAC-Association of Official Analytical Chemists. Official Methods of Analysis, Agricultural Chemicals, 19th ed.; Gaithersburg: Maryland, VA, USA, 2007; pp. 46–48. [Google Scholar]
- Haug, W.; Lantzsch, H.J. Sensitive method for the rapid determination of phytate in cereals and cereal products. J. Sci. Food Agric. 1983, 34, 1423–1426. [Google Scholar] [CrossRef]
- Sun, J.-L.; Zhao, L.-L.; Liao, L.; Tang, X.-H.; Cui, C.; Liu, Q.; He, K.; Ma, J.-D.; Jin, L.; Yan, T.; et al. Interactive effect of thermal and hypoxia on largemouth bass (Micropterus salmoides) gill and liver: Aggravation of oxidative stress, inhibition of immunity and promotion of cell apoptosis. Fish Shellfish Immunol. 2020, 98, 923–936. [Google Scholar] [CrossRef]
- Hong, K.J.; Lee, C.H.; Kim, S.W. Aspergillus oryzae GB-107 fermentation improves nutritional quality of food soybeans and feed soybean meals. J. Med. Food. 2004, 7, 430–435. [Google Scholar] [CrossRef] [PubMed]
- Olukomaiya, O.O.; Adiamo, O.Q.; Fernando, W.C.; Mereddy, R.; Li, X.; Sultanbawa, Y. Effect of solid-state fermentation on proximate composition, anti-nutritional factor, microbiological and functional properties of lupin flour. Food Chem. 2020, 315, 126238. [Google Scholar] [CrossRef] [PubMed]
- Zamudio, M.; Gonzalez, A.; Medina, J.A. Lactobacillus plantarum phytase activity is due to non-specific acid phosphatase. Lett. Appl. Microbiol. 2001, 32, 181–184. [Google Scholar] [CrossRef]
- Lee, S.M. Apparent digestibility coefficients of various feed ingredients for juvenile and grower rockfish (Sebastes schlegeli). Aquaculture 2002, 207, 79–95. [Google Scholar] [CrossRef]
- Kari, Z.A.; Kabir, M.A.; Mat, K.; Rusli, N.D.; Razab, M.K.A.A.; Ariff, N.S.N.A.; Wei, L.S. The possibility of replacing fishmeal with fermented soy pulp on the growth performance, blood biochemistry, liver, and intestinal morphology of African catfish (Clarias gariepinus). Aquac. Rep. 2021, 21, 100815. [Google Scholar] [CrossRef]
- Lim, C.; Lückstädt, C.; Webster, C.D.; Kesius, P. Organic acids and their salts. In Dietary Nutrients, Additives, and Fish Health; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015; pp. 305–319. [Google Scholar]
- Ribeiro, L.; Moura, J.; Santos, M.; Colen, R.; Rodrigues, V.; Bandarra, N.; Soares, F.; Ramalho, P.; Barata, M.; Moura, P.; et al. Effect of vegetable based diets on growth, intestinal morphology, activity of intestinal enzymes and haematological stress indicators in meagre (Argyrosomus regius). Aquaculture 2015, 447, 116–128. [Google Scholar] [CrossRef]
- Huang, B.; Zhang, S.; Dong, X.; Chi, S.; Yang, Q.; Liu, H.; Tan, B.; Xie, S. Effects of fishmeal replacement by black soldier fly on growth performance, digestive enzyme activity, intestine morphology, intestinal flora and immune response of pearl gentian grouper (Epinephelus fuscoguttatus ♀ × Epinephelus lanceolatus ♂). Fish Shellfish Immunol. 2022, 120, 497–506. [Google Scholar] [CrossRef]
- Soltan, M.; Hanafy, M.; Wafa, M. Effect of replacing fish meal by a mixture of different plant protein sources in Nile tilapia (Oreochromis niloticus L.) diets. Glob. Vet. 2008, 2, 157–164. [Google Scholar]
- Xu, Q.Y.; Wang, C.A.; Zhao, Z.G.; Luo, L. Effects of replacement of fish meal by soy protein isolate on the growth, digestive enzyme activity and serum biochemical parameters for juvenile Amur sturgeon (Acipenser schrenckii). Asian-Australas. J. Anim. Sci. 2012, 25, 1588–1594. [Google Scholar] [CrossRef] [PubMed]
- Magnadottir, B.; Lange, S.; Gudmundsdottir, S.; Bogwald, J.; Dalmo, R.A. Ontogeny of humoral immune parameters in fish. Fish Shellfish Immunol. 2005, 19, 429–439. [Google Scholar] [CrossRef]
- Akhter, N.; Wu, B.; Memon, A.M.; Mohsin, M. Probiotics and prebiotics associated with aquaculture: A review. Fish Shellfish Immunol. 2015, 45, 733–741. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Kim, N.; Roh, H.; Park, J.; Kim, M.; Lee, J.; Kim, D.H. Hepcidin-1 in olive flounder (Paralichthys olivaceus): Gene expression, antimicrobial and therapeutic effects of synthetic peptides against bacterial and viral infections. Aquaculture 2020, 560, 738480. [Google Scholar] [CrossRef]
- Hu, Y.; Kurobe, T.; Liu, X.; Zhang, Y.A.; Su, J.; Yuan, G. Hamp type-1 promotes antimicrobial defense via direct microbial killing and regulating iron metabolism in grass carp (Ctenopharyngodon idella). Biomolecules 2020, 10, 825. [Google Scholar] [CrossRef]
- Fridovich, I. Superoxide radical and superoxide dismutases. Annu. Rev. Biochem. 1995, 64, 97–112. [Google Scholar] [CrossRef]
- Verni, M.; Verardo, V.; Rizzello, C.G. How fermentation affects the antioxidant properties of cereals and legumes. Foods 2019, 8, 362. [Google Scholar] [CrossRef]
- McIlwain, D.R.; Berger, T.; Mak, T.W. Caspase functions in cell death and disease. Cold Spring Harb. Perspect. Biol. 2013, 5, a008656. [Google Scholar] [CrossRef]
- Yu, L.L.; Yu, H.H.; Liang, X.F.; Li, N.; Wang, X.; Li, F.H.; Wu, X.F.; Zheng, Y.H.; Xue, M.; Liang, X.F. Dietary butylated hydroxytoluene improves lipid metabolism, antioxidant and anti-apoptotic response of largemouth bass (Micropterus salmoides). Fish Shellfish Immunol. 2018, 72, 220–229. [Google Scholar] [CrossRef]
- Chen, G.F.; Yin, B.; Liu, H.Y.; Tan, B.P.; Dong, X.H.; Yang, Q.H.; Zhang, S. Effects of fish meal replacement with cottonseed protein concentrate on growth, digestive proteinase, intestinal morphology and microflora in pearl gentian grouper (♀ Epinephelus fuscoguttatus × ♂ Epinephelus lanceolatu). Aquac. Res. 2020, 51, 2870–2884. [Google Scholar] [CrossRef]
- Wei, H.; Xing, S.; Chen, P.; Wu, X.; Gu, X.; Luo, L.; Xue, M. Plant protein diet-induced hypoimmunity by affecting the spiral valve intestinal microbiota and bile acid enterohepatic circulation in Amur sturgeon (Acipenser schrenckii). Fish Shellfish Immunol. 2020, 106, 421–430. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Cai, H.; Li, H.; You, F.; Jiang, A.; Hu, W.; Li, K.; Zhang, X.; Zhang, Y.; Chang, X.; et al. Clostridium butyricum-fermented Chinese herbal medicine enhances the immunity by modulating the intestinal microflora of largemouth bass (Micropterus salmoides). Aquaculture 2023, 562, 738768. [Google Scholar] [CrossRef]
- Rimoldi, S.; Gini, E.; Iannini, F.; Gasco, L.; Terova, G. The effects of dietary insect meal from Hermetia illucens prepupae on autochthonous gut microbiota of rainbow trout (Oncorhynchus mykiss). Animals 2019, 9, 143. [Google Scholar] [CrossRef] [PubMed]
- Porto, M.C.W.; Kuniyoshi, T.M.; Azevedo, P.O.S.; Vitolo, M.; Oliveira, R.P.S. Pediococcus spp.: An important genus of lactic acid bacteria and pediocin producers. Biotechnol. Adv. 2017, 35, 361–374. [Google Scholar] [CrossRef] [PubMed]
- Ghatak, S.; Blom, J.; Das, S.; Sanjukta, R.; Puro, K.; Mawlong, M.; Shakuntala, I.; Sen, A.; Goesmann, A.; Kumar, A.; et al. Pan-genome analysis of Aeromonas hydrophila, Aeromonas veronii and Aeromonas caviae indicates phylogenomic diversity and greater pathogenic potential for Aeromonas hydrophila. Antonie Leeuwenhoek 2016, 109, 945–956. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Kim, S.; Kim, A.; Kang, S.; Park, M.; Sung, M. Dietary fat intake and age modulate the composition of the gut microbiota and colonic inflammation in C57BL/6J mice. BMC Microbiol. 2019, 19, 193. [Google Scholar] [CrossRef]
- Do, M.; Lee, E.; Oh, M.; Kim, Y.; Park, H. High-glucose or -fructose diet cause changes of the gut microbiota and metabolic disorders in mice without body weight change. Nutrients 2018, 10, 761. [Google Scholar] [CrossRef]
- Xia, Y.; Yu, E.M.; Lu, M.; Xie, J.J.A. Effects of probiotic supplementation on gut microbiota as well as metabolite profiles within Nile tilapia, Oreochromis niloticus. Aquaculture 2020, 527, 735428. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | Control | 10% FSBM | 30% FSBM | 50% FSBM |
|---|---|---|---|---|
| Fish meal | 350 | 315 | 245 | 175 |
| Fermented soybean meal | 0 | 47.7 | 143.1 | 238.5 |
| Corn gluten meal | 180 | 180 | 180 | 180 |
| Wheat flour | 145.8 | 132.7 | 106.3 | 80 |
| Wheat gluten meal | 80 | 80 | 80 | 80 |
| Fish oil | 20 | 20 | 20 | 20 |
| Corn oil | 20 | 21.3 | 23.2 | 25.7 |
| Soybean phospholipid | 30 | 30 | 30 | 30 |
| Lysine | 4 | 4 | 4 | 4 |
| Arginine | 6 | 6 | 6 | 6 |
| Methionine | 8 | 8 | 8 | 8 |
| Monocalcium phosphate | 20 | 20 | 20 | 20 |
| Mineral premix | 5 | 5 | 5 | 5 |
| Vitamin premix | 5 | 5 | 5 | 5 |
| Carboxymethyl cellulose | 40 | 40 | 40 | 40 |
| Y2O3 | 1 | 1 | 1 | 1 |
| Microcrystalline cellulose | 75.2 | 74.3 | 73.4 | 71.8 |
| Alpha starch | 10 | 10 | 10 | 10 |
| Proximate composition (% dry-matter basis) | ||||
| Moisture | 7.27 ± 0.031 | 7.4 ± 0.055 | 7.72 ± 0.046 | 7.84 ± 0.07 |
| Crude Protein | 45.97 ± 0.16 | 46.42 ± 0.22 | 45.98 ± 0.44 | 45.6 ± 0.1 |
| Crude fiber | 8.93 ± 0.13 | 9.45 ± 0.19 | 9.53 ± 0.59 | 11.32 ± 0.32 |
| Ash | 8.65 ± 0.07 | 8.74 ± 0.09 | 9.33 ± 0.24 | 9.34 ± 0.184 |
| Crude lipid | 10.86 ± 0.099 | 11.61 ± 0.19 | 11.55 ± 0.31 | 11.92 ± 0.2 |
| NFE | 25.59 | 23.59 | 23.61 | 21.82 |
| GE (kJ/g) | 18.67 | 18.51 | 18.60 | 18.20 |
| The Type of Amino Acid | SBM (mg g−1 Dry Matter Basis) | FSBM (mg g−1 Dry Matter Basis) | Change (%) |
|---|---|---|---|
| Essential amino acid | |||
| Valine | 19.343 | 20.263 | +4.76 |
| Methionine | 10.535 | 11.175 | +6.07 |
| Isoleucine | 26.410 | 27.559 | +4.35 |
| Leucine | 33.681 | 36.324 | +7.85 |
| Threonine | 17.776 | 18.168 | +2.21 |
| Phenylalanine | 22.223 | 24.508 | +10.28 |
| Histidine | 14.507 | 13.782 | +5.00 |
| Lysine | 26.435 | 26.589 | +0.58 |
| Arginine | 26.740 | 27.548 | +3.02 |
| Non-essential amino acid | |||
| Aspartic acid | 45.331 | 45.359 | +0.06 |
| Proline | 39.149 | 39.581 | +1.10 |
| Serine | 22.291 | 22.423 | +0.59 |
| Glutamic acid | 71.521 | 73.187 | +2.33 |
| Glycine | 17.248 | 17.950 | +4.07 |
| Alanine | 17.636 | 18.015 | +2.15 |
| Cysteine | 2.633 | 3.274 | +24.34 |
| Tyrosine | 17.741 | 18.198 | +2.58 |
| Total amino acid | 431.2 | 443.903 | +2.95 |
| Crude protein | 460.3 | 494.6 | +7.45 |
| Phytic acid | 29.8 ± 0.2 | 15.3 ± 0.16 | −48.66 |
| Parameters | Control | 10% FSBM | 30% FSBM | 50% FSBM |
|---|---|---|---|---|
| IBW | 60.45 ± 0.69 a | 60.39 ± 0.42 a | 60.72 ± 0.54 a | 60.28 ± 0.92 a |
| FBW | 172.99 ± 6.65 a | 162.65 ± 5.73 a | 169.04 ± 6.26 a | 130.32 ± 5.20 b |
| SR | 100 | 100 | 100 | 100 |
| WG | 186.18 ± 10.99 a | 169.33 ± 9.49 a | 178.39 ± 10.30 a | 116.18 ± 8.63 b |
| SGR | 1.88 ± 0.069 a | 1.77 ± 0.063 b | 1.83 ± 0.066 ab | 1.38 ± 0.071 c |
| FCR | 1.11 ± 0.067 b | 1.21 ± 0.066 b | 1.22 ± 0.070 b | 1.79 ± 0.13 a |
| FE | 90.12 ± 5.32 a | 82.73 ± 4.64 a | 82.24 ± 4.75 a | 55.93 ± 4.16 b |
| VSI | 8.37 ± 0.40 a | 7.84 ± 0.48 ab | 7.39 ± 0.16 b | 7.66 ± 0.12 ab |
| CF | 2.34 ± 0.038 a | 2.32 ± 0.048 ab | 2.30 ± 0.032 b | 2.29 ± 0.040 b |
| HIS 1 | 2.54 ± 0.068 a | 2.32 ± 0.18 ab | 1.79 ± 0.093 c | 1.95 ± 0.069 bc |
| Control | 10% FSBM | 30% FSBM | 50% FSBM | |
|---|---|---|---|---|
| ADDM | 72.09 ± 0.64 | 73.53 ± 0.06 | 68.02 ± 0.27 | 69.16 ± 0.04 |
| ADCP | 74.17 ± 0.10 | 75.64 ± 0.29 | 68.43 ± 0.30 | 69.85 ± 0.85 |
| ADCL | 80.21 ± 0.44 | 83.39 ± 0.55 | 85.57 ± 0.25 | 84.22 ± 0.15 |
| Control | 10% FSBM | 30% FSBM | 50% FSBM | |
|---|---|---|---|---|
| The height of intestinal villi (mm) | 0.9571 ± 0.0073 a | 0.9437 ± 0.0758 a | 0.6888 ± 0.0468 b | 0.7704 ± 0.0518 b |
| The width of intestinal villi (mm) | 0.1606 ± 0.0134 a | 0.1345 ± 0.1366 b | 0.1332 ± 0.0130 b | 0.1315 ± 0.0026 b |
| Muscle layer thickness (mm) | 0.1867 ± 0.2575 a | 0.1608 ± 0.0343 a | 0.1549 ± 0.0113 a | 0.1668 ± 0.0060 a |
| Control | 10% FSBM | 30% FSBM | 50% FSBM | |
|---|---|---|---|---|
| ALT (U/L) | 1.2 | 2.2 | 1.3 | 1.7 |
| AST (U/L) | 13.2 | 12.4 | 11.6 | 12.7 |
| TP (g/L) | 20 | 14.4 | 18.5 | 13.3 |
| ALB (g/L) | 9.6 | 6.3 | 8.4 | 6 |
| GLB (g/L) | 10.4 | 8.1 | 10.1 | 7.3 |
| GLU (mmol/L) | 8.7 | 3.7 | 4.35 | 3.61 |
| ALP (U/L) | 64 | 95 | 102 | 70 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weng, L.; Wang, Z.; Zhuang, W.; Yang, T.; Xu, X.; Liu, J.; Liu, J.; Xu, Z.; Chen, R.; Wang, Q.; et al. Effect of Replacing Fish Meal Using Fermented Soybean Meal on Growth Performance, Intestine Bacterial Diversity, and Key Gene Expression of Largemouth Bass (Micropterus salmoides). Fermentation 2023, 9, 520. https://doi.org/10.3390/fermentation9060520
Weng L, Wang Z, Zhuang W, Yang T, Xu X, Liu J, Liu J, Xu Z, Chen R, Wang Q, et al. Effect of Replacing Fish Meal Using Fermented Soybean Meal on Growth Performance, Intestine Bacterial Diversity, and Key Gene Expression of Largemouth Bass (Micropterus salmoides). Fermentation. 2023; 9(6):520. https://doi.org/10.3390/fermentation9060520
Chicago/Turabian StyleWeng, Longfei, Zhi Wang, Wei Zhuang, Tiezhu Yang, Xinxin Xu, Jinle Liu, Jixiang Liu, Zhengzhong Xu, Ruitao Chen, Qi Wang, and et al. 2023. "Effect of Replacing Fish Meal Using Fermented Soybean Meal on Growth Performance, Intestine Bacterial Diversity, and Key Gene Expression of Largemouth Bass (Micropterus salmoides)" Fermentation 9, no. 6: 520. https://doi.org/10.3390/fermentation9060520
APA StyleWeng, L., Wang, Z., Zhuang, W., Yang, T., Xu, X., Liu, J., Liu, J., Xu, Z., Chen, R., Wang, Q., Wang, S., Cai, Y., & Ying, H. (2023). Effect of Replacing Fish Meal Using Fermented Soybean Meal on Growth Performance, Intestine Bacterial Diversity, and Key Gene Expression of Largemouth Bass (Micropterus salmoides). Fermentation, 9(6), 520. https://doi.org/10.3390/fermentation9060520