
Supplementation of 5,6-Dimethylbenzimidazole and Cobalt in High-Concentrate Diet Improves the Ruminal Vitamin B12 Synthesis and Fermentation of Sheep

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Feeding and Management
2.3. Sample Collection
2.4. Mesurement of Index
2.4.1. Measurement of Growth Performance
2.4.2. Mesurement of Vitamin B12 Content
2.4.3. Measurement of Rumen Fermentation Parameters
2.5. Statistical Analysis
3. Results
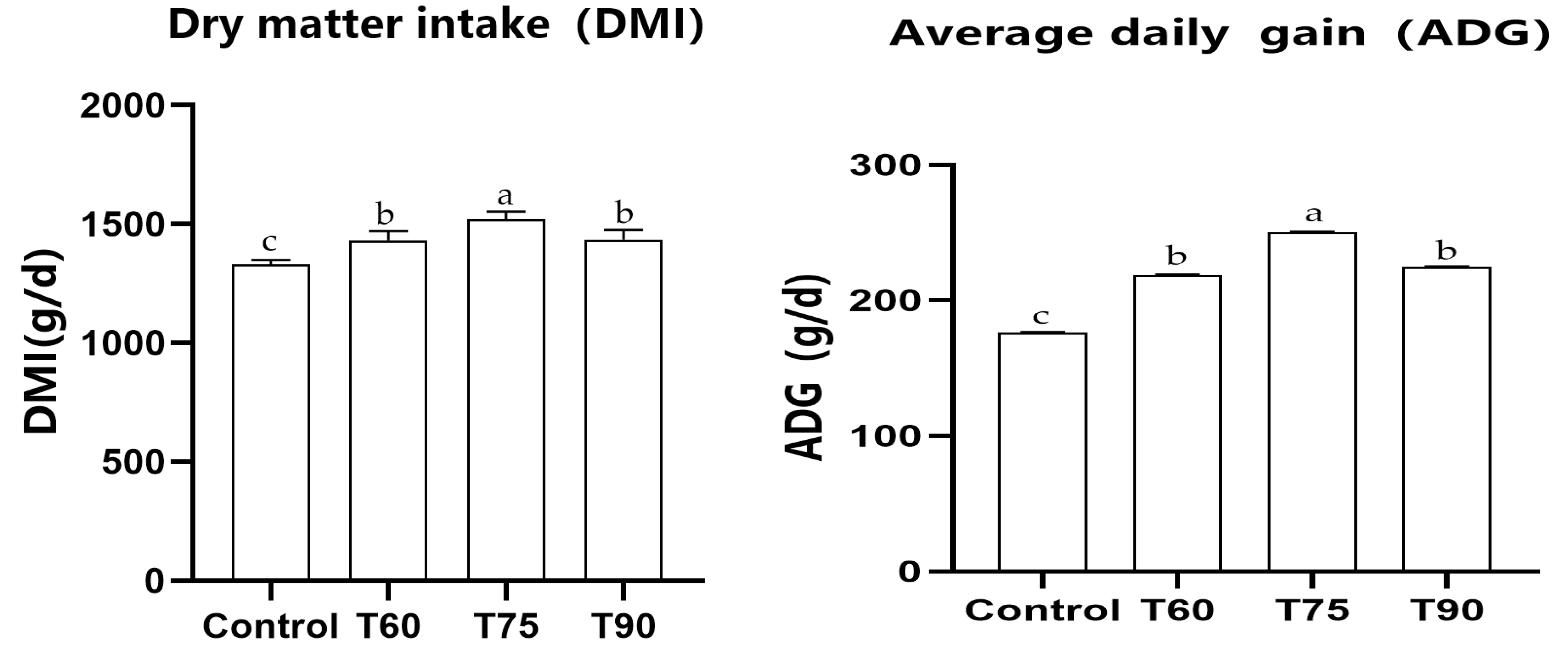
3.1. Effect of the Supplementation of 5,6-Dimethylbenzimidazole and Cobalt on Growth Performance of Sheep
3.2. Effect of the Supplementation of 5,6-Dimethylbenzimidazole and Cobalt on the Vitamin B12 Content of Sheep Rumen
3.3. Effect of the Supplementation of 5,6-Dimethylbenzimidazole and Cobalt on Ruminal pH in Sheep
3.4. Effect of the Supplementation of 5,6-Dimethylbenzimidazole and Cobalt on Ruminal NH3-N Concentration in Sheep
3.5. Effect of the Supplementation of 5,6-Dimethylbenzimidazole and Cobalt on Ruminal VFA Concentrations in Sheep
3.6. Effect of Supplementation with 5,6-Dimethylbenzimidazole and Cobalt on Ruminal L- and D-Lactate in Sheep
3.7. Effect of the Supplementation of 5,6-Dimethylbenzimidazole and Cobalt on Ruminal MCP Content in Sheep
3.8. Correlation Analysis of Growth Performance, Vitamin B12 Content and Ruminal Fermentation
4. Discussion
4.1. Effects of Supplemental 5,6-Dimethylbenzimidazole and Cobalt on Growth Performance of Sheep
4.2. Effects of Supplementation 5,6-Dimethylbenzimidazole and Cobalt on Vitamin B12 in Sheep
4.3. Effect of Supplementation 5,6-Dimethylbenzimidazole and Cobalt on Rumen Fermentation Parameters in Sheep
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fu, Y.; Li, L.G.; Wang, Y.F.; Wang, L.; Shi, W.Q. History of Natural Medicinal Chemistry: Vitamin B12. Chin. Herb. Med. 2015, 46, 1259–1264. [Google Scholar]
- Li, G.D.; Liu, M.M.; Zhan, J.S.; Zhao, G.Q. Biological functions of cobalt and its application to ruminants. China Feed 2016, 27, 5–8. [Google Scholar]
- Gonzalez Monta, J.; Escaleravalente, F.; Alonso, A.J.; Lomillos, J.M.; Robles, R.; Alonso, M.E. Relationship between Vitamin B12 and Cobalt Metabolism in Domestic Ruminant: An Update. Animals 2020, 10, 1885. [Google Scholar] [CrossRef] [PubMed]
- Clark, J.H.; Klusmeyer, T.H.; Cameron, M.R. Microbial protein synthesis and flows of nitrogen fractions to the duodenum of dairy cows. J. Dairy Sci. 1992, 75, 2304–2323. [Google Scholar] [CrossRef]
- Wang, D.M.; Zhang, B.X.; Wang, J.K.; Liu, H.Y.; Liu, J.X. Short communication: Effects of dietary 5,6-dimethylbenzimidazole supplementation on vitamin B12 supply, lactation performance, and energy balance in dairy cows during the transition period and early lactation. J. Dairy Sci. 2017, 101, 2144–2147. [Google Scholar] [CrossRef]
- Wu, C.; Yao, Z.H.; Mei, W.Q.; Feng, Y.Y.; Chen, Q.; Ni, Y.D. Effect of vitamin B complex on the composition of intestinal flora and intestinal mucosa in the hind quarters of growing goats. Acta Prataculturae Sin. 2021, 30, 170–180. [Google Scholar]
- Stemme, K.; Lebzien, P.; Flachowsky, G.; Scholz, H. The influence of an increased cobalt supply on ruminal parameters and microbial vitamin B12 synthesis in the rumen of dairy cows. Arch. Anim. Nutr. 2008, 62, 207–218. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.L.; Zhang, W.; Zhang, Y.Z.; Zhang, C.X.; Chen, J.B.; Jia, Z.H. Effects of different cobalt levels on ruminal vitamin B12 synthesis, ruminal fermentation and hematopoiesis in meat sheep. Chin. J. Anim. Nutr. 2007, 19, 534–538. [Google Scholar]
- Gagnon, D.M.; Stich, T.A.; Mehta, A.P.; Abdelwahed, S.H.; Begley, T.P.; Britt, R.D. An Aminoimidazole Radical Intermediate in the Anaerobic Biosynthesis of the 5,6-Dimethylbenzimidazole Ligand to Vitamin B12. J. Am. Chem. Soc. 2018, 140, 12798–12807. [Google Scholar] [CrossRef] [PubMed]
- Brito, A.; Chiquette, J.; Stabler, S.P.; Allen, R.H.; Girard, C.L. Supplementing lactating dairy cows with a vitamin B12 precursor, 5,6-dimethylbenzimidazole, increases the apparent ruminal synthesis of vitamin B12. Animal 2015, 9, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Ruan, C.; Xiong, S.Y.; Zang, C.J.; Cheng, Z.Q.; Jiao, Y.L.; Yu, Y.Y.; Li, X.B.; Li, F.M.; Luo, Q.J. Effects of adding 5,6-dimethylbenzimidazole and cobalt to high-concentrate rations on growth performance and digestive metabolism in sheep. Chin. J. Anim. Nutr. 2022, 34, 6576–6586. [Google Scholar]
- Feng, Z.C.; Gao, M. Improvement of the method for determining the ammonia-nitrogen content of rumen fluid by colorimetry. Inn. Mong. Anim. Husb. Sci. 2010, 40–41. [Google Scholar]
- Jun-Yu, Z.; Ling-Ling, S.U. Effects of Different Pretreatment Methods on the Determination of Volatile Fatty Acids in Rumen by Gas Chromatography. Grass-Feed. Livestock 2017, 38, 30–34. [Google Scholar]
- Gao, Y.F. Effect of Niacin on Rumen Microbiota of Cattle under High Concentrate Diet Conditions. Ph.D. Thesis, Jiangxi Agricultural University, Nanchang, China, 2017. [Google Scholar]
- Yao, Z.H.; Mei, W.Q.; Feng, Y.Y. Effects of dietary supplementation with different doses of vitamin B-complex on the growth performance and microflora composition in goats. Anim. Husb. Vet. Med. 2020, 52, 47–53. [Google Scholar]
- Li, N.; Li, M.Y.; Peng, Q.H. Research progress of vitamin B in ruminant Nutrition. Chin. J. Anim. Nutr. 2021, 33, 4909–4919. [Google Scholar]
- Reng, X.W.; Zhao, H.S. Research progress of cobalt in sheep nutrition. J. Grassl. Forage Sci. 2010, 6, 48–60. [Google Scholar]
- Singh, K.K. Effect of dietary cobalt on ruminal vitamin B12 synthesis and rumen metabolites. J. Nucl. Agric. Biol. 1995, 24, 112–116. [Google Scholar]
- Rickard, T.R.; Bigger, G.W.; Elliot, J.M. Effects of 5,6-dimethylbenzimidazole, adenine and riboflavin on ruminal vitamin B12 synthesis. J. Anim. Sci. 1975, 40, 1199–1204. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y. Effects of different cobalt levels in diets on growth performance, digestibility and meat quality of 2~3 month old New Zealand meat rabbits. Shandong J. Anim. Sci. Vet. Med. 2020, 41, 4–7. [Google Scholar]
- Bhawsar, S. Microbial Production of Vitamin B12. Appl. Microbiol. Biotechnol. 2002, 58, 275–285. [Google Scholar]
- Stangl, G.I.; Schwarz, F.J.; Jahn, B.; Kirchgessner, M. Cobalt-deficiency-induced hyperhomocysteinaemia and oxidative status of cattle. Br. J. Nutr. 2000, 83, 3–6. [Google Scholar] [CrossRef] [PubMed]
- Franco-Lopez, J.; Duplessis, M.; Bui, A.; Reymond, C.; Ronholm, J. Correlations between the Composition of the Bovine Microbiota and Vitamin B 12 Abundance. mSystems 2020, 5, e00107–e00120. [Google Scholar] [CrossRef] [PubMed]
- Tiffany, M.E.; Spears, J.W.; Xi, L.; Horton, J. Influence of dietary cobalt source and concentration on performance, vitamin B12 status, and ruminal and plasma metabolites in growing and finishing steers. J. Anim. Sci. 2003, 81, 3151–3159. [Google Scholar] [CrossRef] [PubMed]
- Sui, Y.N. Effects of Different Levels of Cobalt Carbonate on Performance, Rumen Fermentation and Serum Biochemical Indices in Lactating Dairy Cows. Ph.D. Thesis, Yangzhou University, Yangzhou, China, 2018. [Google Scholar]
- Schwab, E.C.; Schwab, C.G.; Shaver, R.D.; Girard, C.L.; Putnam, D.E.; Whitehouse, N.L. Dietary forage and nonfiber carbohydrate contents influence B-vitamin intake, duodenal flow, and apparent ruminal synthesis in lactating dairy cows. J. Dairy Sci. 2006, 89, 174–187. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.S.; Lu, Y. Effects of a micro-ecological complex on growth performance, rumen fermentation parameters, blood biochemical indexes and immunity indexes in calves. China Feed 2021, 31, 46–51. [Google Scholar]
- Anantasook, N.; Wanapat, M.; Cherdthong, A.; Gunun, P. Effect of Plants Containing Secondary Compounds with Palm Oil on Feed Intake, Digestibility, Microbial Protein Synthesis and Microbial Population in Dairy Cows. Asian-Australas. J. Anim. Sci. 2013, 26, 820–826. [Google Scholar] [CrossRef]
- Liu, W.T.; Li, X.B.; Zang, C.J.; Guo, T.J.; Li, F.M.; Zeng, F.X.; Yu, Y.Y. Effect of feeding diets with different ratios of structural and nonstructural carbohydrates on rumen fermentation parameters in sheep. Feed Ind. 2019, 40, 18–24. [Google Scholar]
- Bao, J.Y.; Han, D.; Su, T.T.; Liu, H.; Wang, C.Y.; Zhu, Y.X.; Wang, C. Effects of different ratios of carbohydrates in diets on rumen fermentation parameters and serum biochemical indices in Liaoning velvet goats. Feed Res. 2021, 44, 1–4. [Google Scholar]
- Wang, C.; Wang, Z.S.; Hu, R.; Ma, J.; Cao, G.; Yao, X.H.; Zou, H.W.; Wang, X.Y.; Xue, B.; Wang, L.Z. Effects of different types of white wine lees on growth performance, apparent digestibility of nutrients, serum biochemical indexes and rumen fermentation parameters of western hybrid cattle. Chin. J. Anim. Nutr. 2021, 33, 913–922. [Google Scholar]
- Yao, Z.H.; Mei, W.Q.; Feng, Y.Y.; Ni, D.Y. Effect of vitamin B complex on growth performance and intestinal microbiota of goats. Livest. Vet. Med. 2020, 52, 47–53. [Google Scholar]
- Zhang, R.; Liu, J.; Jiang, L.; Mao, S. Effect of high-concentrate diets on microbial composition, function, and the VFAs formation process in the rumen of dairy cows. Anim. Feed Sci. Technol. 2020, 269, 114619–114630. [Google Scholar] [CrossRef]
- Dezfoulia, A.H.; Aliarabi, H. A comparison between different concentrations and sources of cobalt in goat kid nutrition. Anim. Int. J. Anim. Biosci. 2016, 11, 600–607. [Google Scholar] [CrossRef]
- Liang, J.; Zhang, W.J.; Wang, B. Research progress on the physiological function of L-malic acid and its application in ruminant production. China Anim. Husb. Vet. Med. 2016, 43, 1916–1921. [Google Scholar]
- Feng, Y.L. Ruminant Nutrition; Science Press: Beijing, China, 2004. [Google Scholar]
- Yang, Y. Study of the Effects of Niacin on Acid Metabolism and Microflora in Rumen of Jinjiang Cattle and the Mechanism of Preventing Acidosis. Ph.D. Thesis, Jiangxi Agricultural University, Nanchang, China, 2019. [Google Scholar]
- Wang, J.Y. Biological Chemistry, Previous, 3rd ed.; Higher Education Press: Beijing, China, 2002. [Google Scholar]
- Wu, G.Y. Principles of Animal Nutrition; Science Press: Beijing, China, 2019. [Google Scholar]
- Yang, C.T. Effects of Shrub Encroachment on Grazing Behavior, Rumen Fermentation and Serum Parameters of Yaks in Alpine Meadows. Ph.D. Thesis, Lanzhou University, Lanzhou, China, 2021. [Google Scholar]
- Li, Y.X. Effect of Added Vitamin B12 and Cobalt on Rumen Fermentation In Vitro and Its Mechanism. Ph.D. Thesis, Zhejiang University, Hangzhou, China, 2012. [Google Scholar]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef] [PubMed]
- Sutton, A.L.; Elliot, J.M. Effect of ratio of roughage to concentrate and level of feed intake on ovine ruminal vitamin B12 production. J. Nutr. 1972, 102, 1341. [Google Scholar] [CrossRef]
- Zhang, T.; Mu, Y.; Zhang, R.; Xue, Y.; Guo, C.; Qi, W.; Zhang, J.; Mao, S. Responsive changes of rumen microbiome and metabolome in dairy cows with different susceptibility to subacute ruminal acidosis. Anim. Nutr. 2022, 8, 331–340. [Google Scholar] [CrossRef]
- Liu, W.; Wang, Q.; Song, J.; Xin, J.; Zhang, S.; Lei, Y.; Yang, Y.; Xie, P.; Suo, H. Comparison of Gut Microbiota of Yaks from Different Geographical Regions. Front. Microbiol. 2021, 12, 666940–666950. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Items | Content | Nutrient Levels 2 | Content |
|---|---|---|---|
| Ingredients | DM | 91.33 | |
| Corn stalk | 21.00 | CP | 17.13 |
| Alfalfa | 9.00 | EE | 1.91 |
| Corn | 35.00 | CEL | 13.59 |
| Wheat bran | 8.40 | HC | 9.71 |
| Soybean meal | 14.00 | Lignin | 3.31 |
| Cottonseed meal | 9.10 | Ca | 1.02 |
| Premix 1 | 3.50 | P | 0.55 |
| Total | 100.00 | ME (MJ/kg) | 14.12 |
| Sampling Time | Control Group | T60 Group | T75 Group | T90 Group | SEM | p-Value |
|---|---|---|---|---|---|---|
| 0 h before feeding | 40.72 | 41.32 | 40.73 | 41.23 | 0.51 | 0.967 |
| 1 h after feeding | 40.11 | 41.77 | 41.95 | 41.48 | 0.58 | 0.697 |
| 3 h after feeding | 39.68 b | 43.40 a | 44.19 a | 43.46 a | 0.50 | 0.001 |
| 5 h after feeding | 39.33 | 42.29 | 43.31 | 42.58 | 0.59 | 0.073 |
| 7 h after feeding | 39.17 | 40.79 | 42.17 | 42.07 | 0.48 | 0.087 |
| Sampling Time | Control Group | T60 Group | T75 Group | T90 Group | SEM | p-Value |
|---|---|---|---|---|---|---|
| 0 h before feeding | 6.45 | 6.43 | 6.49 | 6.44 | 0.02 | 0.852 |
| 1 h after feeding | 5.81 b | 5.98 a | 6.03 a | 6.01 a | 0.03 | 0.001 |
| 3 h after feeding | 5.55 b | 5.79 a | 5.80 a | 5.75 a | 0.03 | 0.001 |
| 5 h after feeding | 5.77 c | 5.91 b | 6.06 a | 5.92 b | 0.03 | 0.002 |
| 7 h after feeding | 5.96 c | 6.09 b | 6.20 a | 6.08 b | 0.02 | <0.001 |
| Sampling Time | Control Group | T60 Group | T75 Group | T90 Group | SEM | p-Value |
|---|---|---|---|---|---|---|
| 0 h before feeding | 18.81 | 19.94 | 19.50 | 18.93 | 0.70 | 0.943 |
| 1 h after feeding | 22.26 c | 26.14 b | 29.84 a | 26.07 b | 0.74 | <0.001 |
| 3 h after feeding | 16.59 c | 19.74 b | 23.70 a | 19.15 b | 0.59 | <0.001 |
| 5 h after feeding | 14.14 b | 16.70 ab | 17.67 a | 16.26 ab | 0.34 | 0.052 |
| 7 h after feeding | 12.56 b | 13.74 ab | 16.31 a | 13.35 ab | 0.60 | 0.135 |
| Items | Sampling Time | Control Group | T60 Group | T75 Group | T90 Group | SEM | p-Value |
|---|---|---|---|---|---|---|---|
| Acetate | 0 h before feeding | 59.14 | 59.43 | 58.55 | 57.04 | 0.85 | 0.784 |
| 1 h after feeding | 66.09 | 68.71 | 70.43 | 69.59 | 1.25 | 0.663 | |
| 3 h after feeding | 74.40 | 76.82 | 81.12 | 75.91 | 1.43 | 0.407 | |
| 5 h after feeding | 67.70 | 72.80 | 74.54 | 69.55 | 1.77 | 0.542 | |
| 7 h after feeding | 63.70 | 66.84 | 68.20 | 66.16 | 1.15 | 0.597 | |
| Propionate | 0 h before feeding | 15.89 | 16.48 | 16.50 | 16.83 | 0.17 | 0.264 |
| 1 h after feeding | 19.15 c | 21.74 b | 25.48 a | 22.16 b | 0.52 | <0.001 | |
| 3 h after feeding | 24.37 c | 27.09 b | 30.48 a | 27.56 b | 0.49 | <0.001 | |
| 5 h after feeding | 22.41 b | 23.45 ab | 24.56 a | 23.68 a | 0.24 | 0.007 | |
| 7 h after feeding | 19.77 | 19.95 | 20.59 | 20.66 | 0.27 | 0.589 | |
| Isobutyrate | 0 h before feeding | 1.39 | 1.36 | 1.45 | 1.24 | 0.08 | 0.861 |
| 1 h after feeding | 1.40 | 1.31 | 1.27 | 1.34 | 0.08 | 0.960 | |
| 3 h after feeding | 1.48 | 1.31 | 1.36 | 1.46 | 0.07 | 0.825 | |
| 5 h after feeding | 1.68 | 1.49 | 1.32 | 1.36 | 0.06 | 0.169 | |
| 7 h after feeding | 1.71 | 1.53 | 1.45 | 1.42 | 0.05 | 0.203 | |
| Butyrate | 0 h before feeding | 15.95 | 18.11 | 16.79 | 15.04 | 0.57 | 0.285 |
| 1 h after feeding | 10.89 | 11.48 | 9.99 | 10.19 | 0.37 | 0.500 | |
| 3 h after feeding | 9.13 | 8.25 | 9.06 | 9.66 | 0.29 | 0.413 | |
| 5 h after feeding | 9.31 | 9.21 | 8.25 | 8.64 | 0.33 | 0.656 | |
| 7 h after feeding | 12.43 | 11.25 | 10.59 | 10.70 | 0.27 | 0.054 | |
| Isovalerate | 0 h before feeding | 2.44 | 3.00 | 2.72 | 2.32 | 0.14 | 0.301 |
| 1 h after feeding | 1.49 | 1.42 | 1.30 | 1.40 | 0.05 | 0.684 | |
| 3 h after feeding | 1.24 | 1.24 | 1.24 | 1.42 | 0.05 | 0.419 | |
| 5 h after feeding | 1.39 | 1.39 | 1.31 | 1.28 | 0.06 | 0.859 | |
| 7 h after feeding | 1.94 | 1.51 | 1.62 | 1.48 | 0.08 | 0.177 | |
| Valerate | 0 h before feeding | 2.14 | 2.36 | 2.08 | 1.77 | 0.16 | 0.658 |
| 1 h after feeding | 1.70 | 1.78 | 1.54 | 1.66 | 0.05 | 0.442 | |
| 3 h after feeding | 1.59 | 1.67 | 1.41 | 1.59 | 0.05 | 0.400 | |
| 5 h after feeding | 1.65 | 1.52 | 1.25 | 1.35 | 0.07 | 0.119 | |
| 7 h after feeding | 1.69 | 1.53 | 1.40 | 1.40 | 0.09 | 0.469 | |
| Acetate/propionate | 0 h before feeding | 3.73 | 3.61 | 3.55 | 3.39 | 0.07 | 0.353 |
| 1 h after feeding | 3.45 a | 3.16 ab | 2.76 b | 3.16 ab | 0.08 | 0.013 | |
| 3 h after feeding | 3.05 | 2.84 | 2.66 | 2.75 | 0.06 | 0.077 | |
| 5 h after feeding | 3.02 | 3.10 | 3.04 | 2.94 | 0.07 | 0.894 | |
| 7 h after feeding | 3.23 | 3.35 | 3.31 | 3.20 | 0.06 | 0.872 | |
| T-VFA | 0 h before feeding | 96.95 | 100.74 | 98.09 | 94.24 | 1.26 | 0.347 |
| 1 h after feeding | 100.72 | 106.44 | 110.01 | 106.34 | 1.55 | 0.207 | |
| 3 h after feeding | 112.21 | 116.38 | 124.67 | 117.60 | 1.77 | 0.081 | |
| 5 h after feeding | 104.14 | 109.86 | 111.23 | 105.86 | 1.92 | 0.551 | |
| 7 h after feeding | 101.24 | 102.61 | 103.85 | 101.82 | 1.29 | 0.911 |
| Items | Sampling Time | Control Group | T60 Group | T75 Group | T90 Group | SEM | p-Value |
|---|---|---|---|---|---|---|---|
| L-lactate | 0 h before feeding | 31.11 | 32.20 | 26.36 | 28.13 | 2.08 | 0.766 |
| 1 h after feeding | 45.22 | 46.04 | 30.96 | 31.76 | 3.13 | 0.154 | |
| 3 h after feeding | 62.64 | 51.97 | 48.34 | 43.71 | 2.68 | 0.068 | |
| 5 h after feeding | 75.76 | 55.38 | 54.75 | 56.81 | 3.50 | 0.091 | |
| D-lactate | 0 h before feeding | 1.74 | 1.69 | 1.32 | 1.49 | 0.07 | 0.155 |
| 1 h after feeding | 1.54 | 1.44 | 1.03 | 1.44 | 0.07 | 0.106 | |
| 3 h after feeding | 1.25 | 1.14 | 0.96 | 1.02 | 0.06 | 0.272 | |
| 5 h after feeding | 1.09 | 0.83 | 0.95 | 0.96 | 0.05 | 0.255 |
| Sampling Time | Control Group | T60 Group | T75 Group | T90 Group | SEM | p-Value |
|---|---|---|---|---|---|---|
| 0 h before feeding | 116.85 | 119.97 | 125.92 | 124.15 | 4.51 | 0.906 |
| 1 h after feeding | 123.09 | 137.51 | 147.79 | 151.41 | 7.47 | 0.567 |
| 3 h after feeding | 126.78 b | 149.07 a | 164.54 a | 164.26 a | 4.01 | <0.001 |
| 5 h after feeding | 109.01 | 130.88 | 142.37 | 146.15 | 5.37 | 0.051 |
| 7 h after feeding | 106.03 | 120.66 | 134.13 | 128.09 | 4.23 | 0.094 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, R.; Cheng, Z.; Zang, C.; Cui, C.; Zhang, C.; Jiao, Y.; Li, F.; Li, X.; Yang, K.; Luo, Q. Supplementation of 5,6-Dimethylbenzimidazole and Cobalt in High-Concentrate Diet Improves the Ruminal Vitamin B12 Synthesis and Fermentation of Sheep. Fermentation 2023, 9, 956. https://doi.org/10.3390/fermentation9110956
Zhang R, Cheng Z, Zang C, Cui C, Zhang C, Jiao Y, Li F, Li X, Yang K, Luo Q. Supplementation of 5,6-Dimethylbenzimidazole and Cobalt in High-Concentrate Diet Improves the Ruminal Vitamin B12 Synthesis and Fermentation of Sheep. Fermentation. 2023; 9(11):956. https://doi.org/10.3390/fermentation9110956
Chicago/Turabian StyleZhang, Rui, Zhiqiang Cheng, Changjiang Zang, Changyun Cui, Changwen Zhang, Yiling Jiao, Fengming Li, Xiaobin Li, Kailun Yang, and Qiujiang Luo. 2023. "Supplementation of 5,6-Dimethylbenzimidazole and Cobalt in High-Concentrate Diet Improves the Ruminal Vitamin B12 Synthesis and Fermentation of Sheep" Fermentation 9, no. 11: 956. https://doi.org/10.3390/fermentation9110956
APA StyleZhang, R., Cheng, Z., Zang, C., Cui, C., Zhang, C., Jiao, Y., Li, F., Li, X., Yang, K., & Luo, Q. (2023). Supplementation of 5,6-Dimethylbenzimidazole and Cobalt in High-Concentrate Diet Improves the Ruminal Vitamin B12 Synthesis and Fermentation of Sheep. Fermentation, 9(11), 956. https://doi.org/10.3390/fermentation9110956