
Ex Vivo Fermentation of Hay and Corn by Rumen Bacteria from Cattle and Sheep


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Media Preparation
2.2. Animals and Mixed Rumen Microorganism Cell Suspensions
2.3. Ex Vivo Experiment
2.4. Isolation of Amylolytic Bacteria
2.5. Fermentation End-Product Analyses
2.6. Chemical Composition of Feed Substrates
2.7. Statistical Analyses
3. Results
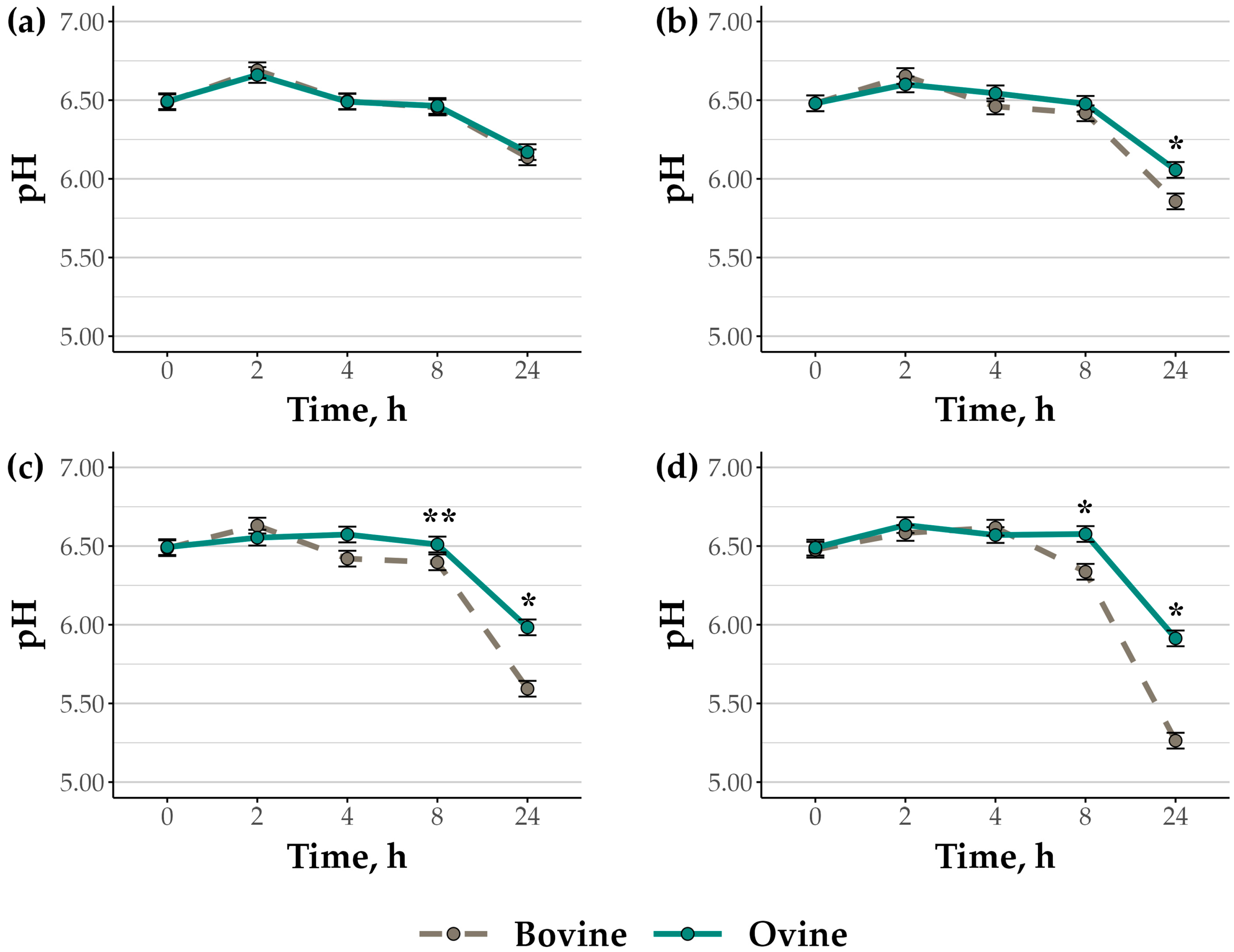
3.1. Rumen Cell Suspension pH and Fermentation End-Products
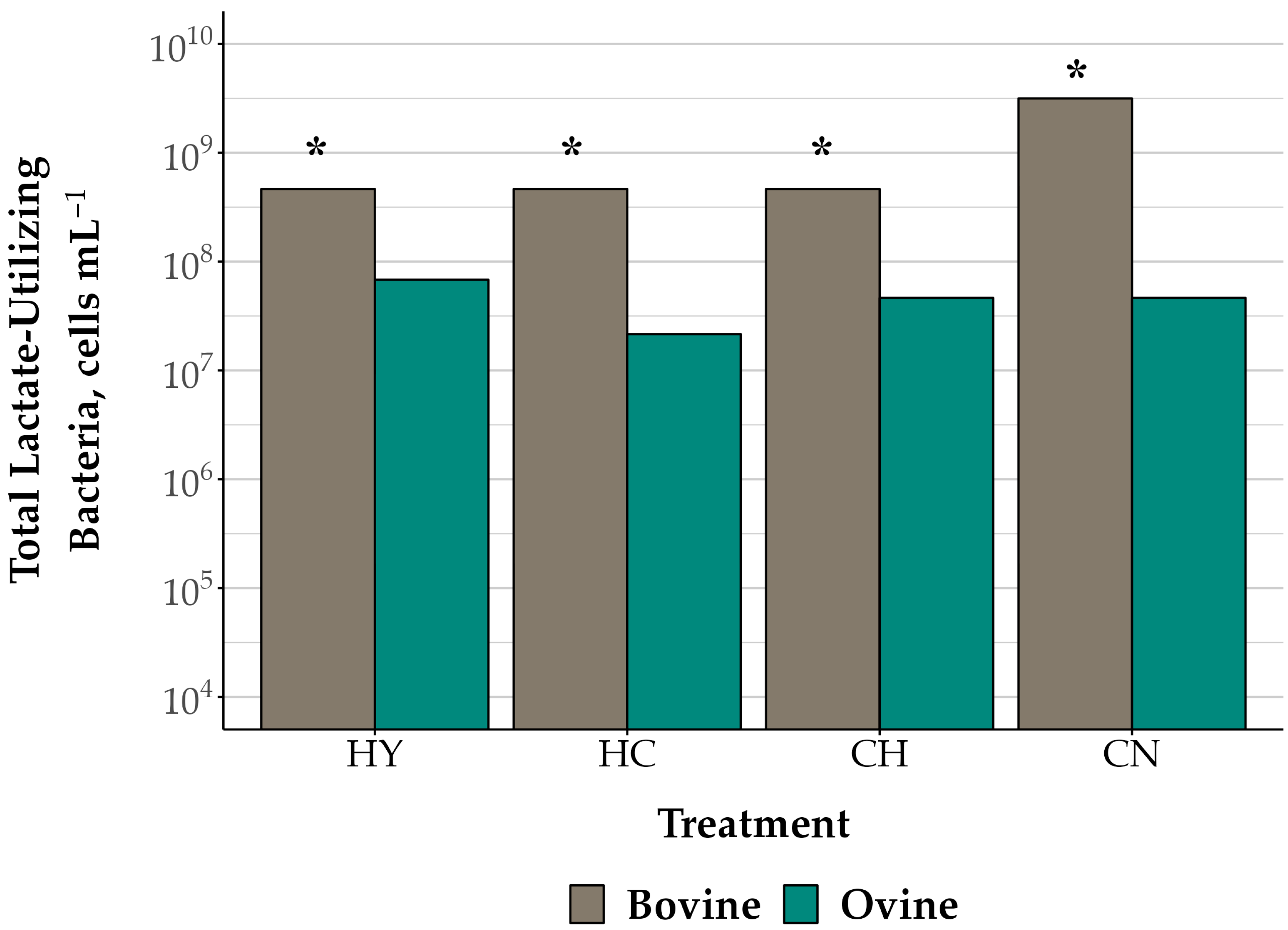
3.2. Rumen Microbiota in Mixed Microorganism Cell Suspensions
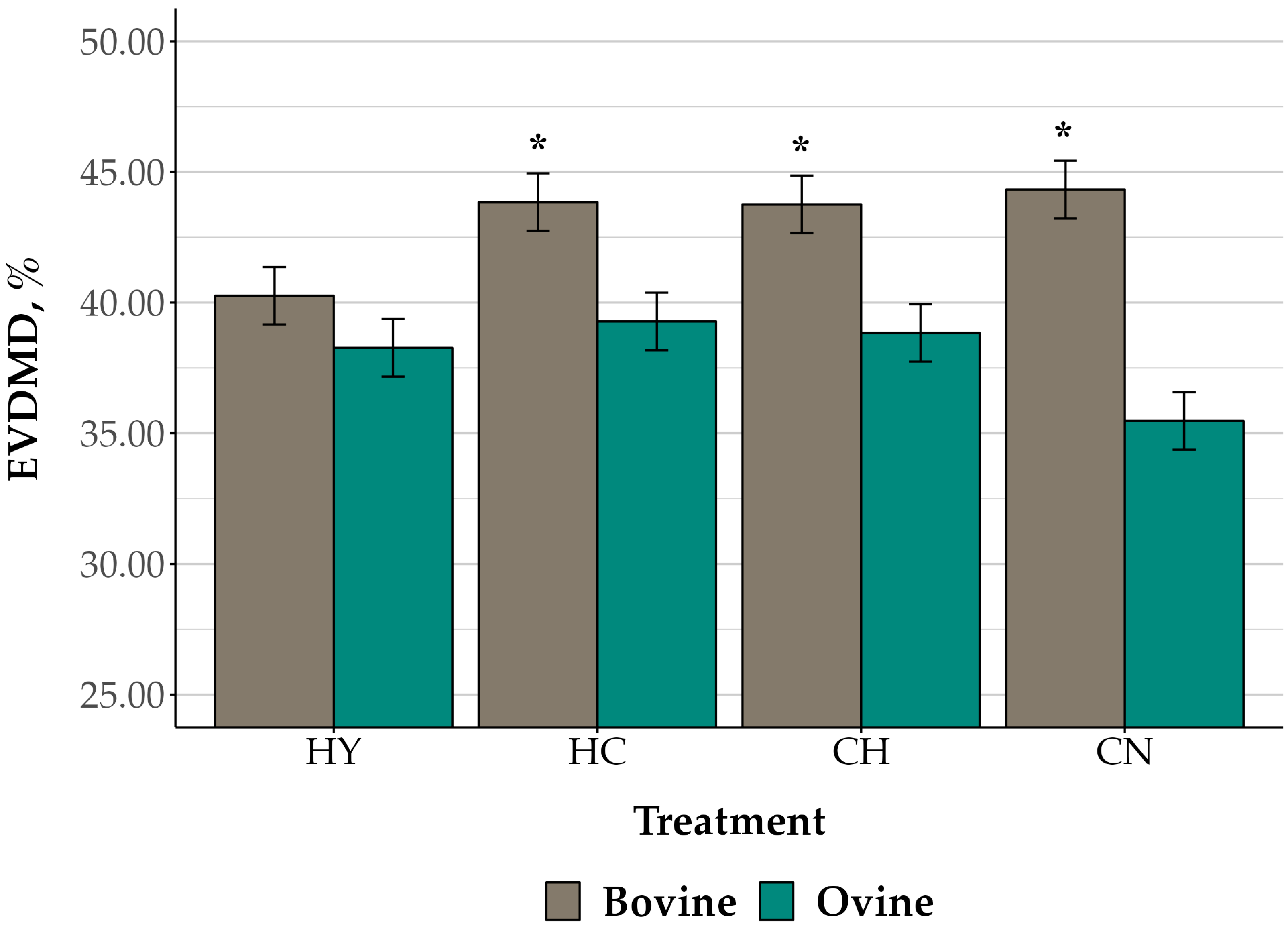
3.3. Ex Vivo Digestibility of Hay and Corn Substrates
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chishti, G.A.; Carvalho, P.H.; Pinto, A.C.; Silva, F.A.; Felix, T.L. Efficacy of sheep as a digestibility model for cattle when fed concentrate-based or forage-based diets. Transl. Anim. Sci. 2019, 3, 1106–1111. [Google Scholar] [CrossRef] [PubMed]
- van Gastelen, S.; Dijkstra, J.; Bannink, A. Are dietary strategies to mitigate enteric methane emission equally effective across dairy cattle, beef cattle, and sheep? J. Dairy Sci. 2019, 102, 6109–6130. [Google Scholar] [CrossRef] [PubMed]
- Prigge, E.C.; Baker, M.J.; Varga, G.A. Comparative digestion, rumen fermentation and kinetics of forage diets by steers and wethers. J. Anim. Sci. 1984, 59, 237–245. [Google Scholar] [CrossRef]
- Reid, R.L.; Jung, G.A.; Cox-Ganser, J.M.; Rybeck, B.F.; Townsend, E.C. Comparative utilization of warm-and cool-season forages by cattle, sheep and goats. J. Anim. Sci. 1990, 60, 2986–2994. [Google Scholar] [CrossRef] [PubMed]
- Pearson, R.A.; Archibald, R.F.; Muirhead, R.H. A comparison of the effect of forage type and level of feeding on the digestibility and gastrointestinal mean retention time of dry forages given to cattle, sheep, ponies and donkeys. Br. J. Nutr. 2006, 95, 88–98. [Google Scholar] [CrossRef] [PubMed]
- O’Mara, F.P.; Coyle, J.E.; Drennan, M.J.; Young, P.; Caffrey, P.J. A comparison of digestibility of some concentrate feed ingredients in cattle and sheep. Anim. Feed Sci. Technol. 1990, 81, 167–174. [Google Scholar] [CrossRef]
- Woods, V.B.; Moloney, A.P.; Mulligan, F.J.; Kenny, M.J.; O’Mara, F.P. The effect of animal species (cattle or sheep) and level of intake by cattle on in vivo digestibility of concentrate ingredients. Anim. Feed Sci. Technol. 1990, 80, 135–150. [Google Scholar] [CrossRef]
- Siddons, R.C.; Paradine, J. Protein degradation in the rumen of cattle and sheep. J. Sci. Food Agric. 1983, 34, 701–708. [Google Scholar] [CrossRef]
- Soto-Navarro, S.A.; Lopez, R.; Sankey, C.; Capitan, B.M.; Holland, B.P.; Balstad, L.A.; Krehbiel, C.R. Comparative digestibility by cattle versus sheep: Effect of forage quality. J. Anim. Sci. 2014, 92, 1621–1629. [Google Scholar] [CrossRef]
- Li, J.; Yan, H.; Chen, J.; Duan, C.; Guo, Y.; Liu, Y.; Zhang, Y.; Ji, S. Correlation of Ruminal Fermentation Parameters and Rumen Bacterial Community by Comparing Those of the Goat, Sheep, and Cow In vitro. Fermentation 2022, 8, 427. [Google Scholar] [CrossRef]
- Owens, F.; Secrist, D.; Hill, W.; Gill, D. Acidosis in cattle: A review. J. Anim. Sci. 1998, 76, 275–286. [Google Scholar] [CrossRef] [PubMed]
- Krause, K.M.; Oetzel, G.R. Understanding and preventing subacute ruminal acidosis in dairy herds: A review. Anim. Feed Sci. Technol. 2006, 126, 215–236. [Google Scholar] [CrossRef]
- Nagaraja, T.; Titgemeyer, E. Ruminal acidosis in beef cattle: The current microbiological and nutritional outlook 1, 2. J. Dairy Sci. 2007, 90, E17–E38. [Google Scholar] [CrossRef] [PubMed]
- Therion, J.J.; Kistner, A.; Kornelius, J.H. Effect of pH on growth rates of rumen amylolytic and lactilytic bacteria. Appl. Environ. Microbiol. 1982, 44, 428–434. [Google Scholar] [CrossRef]
- Commun, L.; Mialon, M.M.; Martin, C.; Baumont, R.; Veissier, I. Risk of subacute ruminal acidosis in sheep with separate access to forage and concentrate. J. Anim. Sci. 2009, 87, 3372–3379. [Google Scholar] [CrossRef] [PubMed]
- Gill, H.S.; Shu, Q.; Leng, R.A. Immunization with Streptococcus bovis protects against lactic acidosis in sheep. Vaccine 2000, 18, 2541–2548. [Google Scholar] [CrossRef]
- Minuti, A.; Ahmed, S.; Trevisi, E.; Piccioli-Cappelli, F.; Bertoni, G.; Jahan, N.; Bani, P. Experimental acute rumen acidosis in sheep: Consequences on clinical, rumen, and gastrointestinal permeability conditions and blood chemistry. J. Anim. Sci. 2014, 92, 3966–3977. [Google Scholar] [CrossRef]
- Stack, R.J.; Hungate, R.E.; Opsahl, W.P. Phenylacetic acid stimulation of cellulose digestion by Ruminococcus albus 8. Appl. Environ. Microbiol. 1983, 46, 539–544. [Google Scholar] [CrossRef]
- Harlow, B.E.; Flythe, M.D.; Aiken, G.E. Effect of biochanin A on corn grain (Zea mays) fermentation by bovine rumen amylolytic bacteria. J. Appl. Microbiol. 2017, 122, 870–880. [Google Scholar] [CrossRef]
- Russell, J.B.; Strobel, H.J.; Chen, G.J. Enrichment and isolation of a ruminal bacterium with a very high specific activity of ammonia production. Appl. Environ. Microbiol. 1988, 54, 872–877. [Google Scholar] [CrossRef]
- Chen, G.; Russell, J.B. More monensin-sensitive, ammonia-producing bacteria from the rumen. Appl. Environ. Microbiol. 1989, 55, 1052–1057. [Google Scholar] [CrossRef] [PubMed]
- Mackie, R.; Heath, S. Enumeration and isolation of lactate-utilizing bacteria from the rumen of sheep. Appl. Environ. Microbiol. 1979, 38, 416–421. [Google Scholar] [CrossRef] [PubMed]
- FASS. Guide for the Care and Use of Agricultural Animals in Research and Teaching; FASS Inc.: Champaign, IL, USA, 2020. [Google Scholar]
- Flythe, M.D. The antimicrobial effects of hops (Humulus lupulus L.) on ruminal hyper ammonia-producing bacteria. Lett. Appl. Microbiol. 2009, 48, 712–717. [Google Scholar] [CrossRef] [PubMed]
- Harlow, B.E.; Flythe, M.D.; Aiken, G.E. Biochanin A improves fibre fermentation by cellulolytic bacteria. J. Appl. Microbiol. 2018, 124, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Harlow, B.E.; Flythe, M.D.; Klotz, J.L.; Harmon, D.L.; Aiken, G.E. Effect of biochanin A on the rumen microbial community of Holstein steers consuming a high fiber diet and subjected to a subacute acidosis challenge. PLoS ONE 2021, 16, e0253754. [Google Scholar] [CrossRef] [PubMed]
- ANKOM Technology. Method 6: Neutral Detergent Fiber in Feeds—Filter Bag Technique (for A200 and A2001); ANKOM Technology: Macedon, NY, USA, 2006; Available online: https://www.ankom.com/analytical-methods-support/fiber-analyzer-a200?f%5B0%5D=field_faq_group%3A89#:~:text=View%20PDF-,Neutral,-Detergent%20Fiber%20Method (accessed on 26 July 2022).
- Weinert-Nelson, J.R.; Ely, D.G.; Flythe, M.D.; Hamilton, T.A.; May, J.B.; Ferrell, J.L.; Hamilton, M.C.; LeeAnn Jacks, W.; Davis, B.E. Red clover supplementation modifies rumen fermentation and promotes feed efficiency in ram lambs. J. Anim. Sci. 2023, 101, skad036. [Google Scholar] [CrossRef] [PubMed]
- Janda, J.M.; Abbott, S.L. 16S rRNA gene sequencing for bacterial identification in the diagnostic laboratory: Pluses, perils, and pitfalls. J. Clin. Microbiol. 2007, 45, 2761–2764. [Google Scholar] [CrossRef] [PubMed]
- Drummond, A.J.; Krearse, M.; Heled, J.; Moir, R.; Thierer, T.; Ashton, B.; Wilson, A.; Stone-Havas, S. Geneious v2.5. 2006. Available online: http://www.geneious.com (accessed on 19 October 2023).
- Benson, D.A.; Cavanaugh, M.; Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2013, 41, D36–D42. [Google Scholar] [CrossRef]
- Chaney, A.L.; Marbach, E.P. Modified reagents for determination of urea and ammonia. Clin. Chem. 1962, 8, 130–132. [Google Scholar] [CrossRef]
- ANKOM Technology. Method 6: Acid Detergent Fiber in Feeds—Filter Bag Technique (for A200 and A2001); ANKOM Technology: Macedon, NY, USA, 2005; Available online: https://www.ankom.com/analytical-methods-support/fiber-analyzer-a200?f%5B0%5D=field_faq_group%3A89#:~:text=Acid-,Detergent,-Fiber%20Method%20(A200) (accessed on 26 July 2022).
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- National Research Council. Nutrient Requirements of Dairy Cattle; National Academy Press: Washington, DC, USA, 2001.
- Weiss, W.P.; Conrad, H.R.; Pierre, N.S. A theoretically-based model for predicting total digestible nutrient values of forages and concentrates. Anim. Feed Sci. Technol. 1992, 39, 95–110. [Google Scholar] [CrossRef]
- Henderson, G.; Cox, F.; Ganesh, S.; Jonker, A.; Young, W.; Janssen, P.H. Rumen microbial community composition varies with diet and host, but a core microbiome is found across a wide geographical range. Sci. Rep. 2015, 5, 14567. [Google Scholar] [CrossRef]
- Huntington, J.A.; Givens, D.I. Studies on in situ degradation of feeds in the rumen: 1. Effect of species, bag mobility and incubation sequence on dry matter disappearance. Anim. Feed Sci. Technol. 1997, 64, 227–241. [Google Scholar] [CrossRef]
- Norton, B.W.; Pieris, H.; Elliott, R. Fermentation patterns and diet utilization by cattle, sheep and goats given grain or molasses based diets. In Proceedings-Australian Society of Animal Production; Australian Society of Animal Production: Coolangatta, Australia, 1994; Volume 20, p. 182. [Google Scholar]
- Colucci, P.E.; Macleod, G.K.; Grovum, W.L.; Cahill, L.W.; McMillan, I. Comparative digestion in cattle and sheep fed different forage to concentrate ratios at high and low intakes. J. Dairy Sci. 1989, 72, 1774–1785. [Google Scholar] [CrossRef] [PubMed]
- Russell, J.B.; Dombrowski, D. Effect of pH on the efficiency of growth by pure cultures of rumen bacteria in continuous culture. Appl. Environ. Microbiol. 1980, 39, 604–610. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Weimer, P. Response surface analysis of the effects of pH and dilution rate on Ruminococcus flavefaciens FD-1 in cellulose-fed continuous culture. Appl. Environ. Microbiol. 1992, 58, 2583–2591. [Google Scholar] [CrossRef] [PubMed]
- Weimer, P.J. Effects of dilution rate and pH on the ruminal cellulolytic bacterium Fibrobacter succinogenes S85 in cellulose-fed continuous culture. Arch. Microbiol. 1993, 160, 288. [Google Scholar] [CrossRef]
- Chatterton, N.J.; Harrison, P.A.; Bennett, J.H.; Asay, K.H. Carbohydrate partitioning in 185 accessions of Gramineae grown under warm and cool temperatures. J. Plant Physiol. 1989, 134, 169–179. [Google Scholar] [CrossRef]
- Kagan, I.A.; Kirch, B.H.; Thatcher, C.D.; Strickland, J.R.; Teutsch, C.D.; Elvinger, F.; Pleasant, R.S. Seasonal and diurnal variation in simple sugar and fructan composition of orchardgrass pasture and hay in the Piedmont region of the United States. J. Equine Vet. Sci. 2001, 31, 488–497. [Google Scholar] [CrossRef]
- Kagan, I.A.; Lawrence, L.M.; Seman, D.H.; Prince, K.J.; Fowler, A.L.; Smith, S.R. Effects of sampling time, cultivar, and methodology on water-and ethanol-soluble carbohydrate profiles of three cool-season grasses in Central Kentucky. J. Equine Vet. Sci. 2018, 61, 99–107. [Google Scholar] [CrossRef]
- Britton, R.; Stock, R. Acidosis: A continual problem in cattle fed high grain diets. In Proceedings of Cornell Nutrition Conference for Feed Manufactures; Cornell University: Ithaca, NY, USA, 1989; pp. 8–15. [Google Scholar]
- Atlas, R.M. Handbook of Microbiological Media, 3rd ed.; CRC Press: Boca Raton, FL, USA, 2004. [Google Scholar]
- Braun, U.; Rihs, T.; Schefer, U. Ruminal lactic acidosis in sheep and goats. Vet. Rec. 1992, 130, 343–349. [Google Scholar] [CrossRef] [PubMed]
- Castillo-Lopez, E.; Wiese, B.I.; Hendrick, S.; McKinnon, J.J.; McAllister, T.A.; Beauchemin, K.A.; Penner, G.B. Incidence, prevalence, severity, and risk factors for ruminal acidosis in feedlot steers during backgrounding, diet transition, and finishing. J. Anim. Sci. 2014, 92, 3053–3063. [Google Scholar] [CrossRef] [PubMed]
- Morgante, M.; Stelletta, C.; Berzaghi, P.; Gianesella, M.; Andrighetto, I. Subacute rumen acidosis in lactating cows: An investigation in intensive Italian dairy herds. J. Anim. Physiol. Anim. Nutr. 2007, 91, 226–234. [Google Scholar] [CrossRef]
- Bramley, E.; Lean, I.J.; Fulkerson, W.J.; Stevenson, M.A.; Rabiee, A.R.; Costa, N.D. The definition of acidosis in dairy herds predominantly fed on pasture and concentrates. J. Dairy Sci. 2008, 91, 308–321. [Google Scholar] [CrossRef] [PubMed]
- O’Grady, L.; Doherty, M.L.; Mulligan, F.J. Subacute ruminal acidosis (SARA) in grazing Irish dairy cows. Vet. J. 2008, 176, 44–49. [Google Scholar] [CrossRef]
- Kleen, J.L.; Hooijer, G.A.; Rehage, J.; Noordhuizen, J.P.T.M. Subacute ruminal acidosis in Dutch dairy herds. Vet. Rec. 2009, 164, 681–684. [Google Scholar] [CrossRef]
- Kleen, J.L.; Upgang, L.; Rehage, J. Prevalence and consequences of subacute ruminal acidosis in German dairy herds. Acta Vet. Scand. 2013, 55, 48. [Google Scholar] [CrossRef]
- Atkinson, O. Prevalence of subacute ruminal acidosis (SARA) on UK dairy farms. Cattle Pract. 2014, 22, 1–9. [Google Scholar]
- Stefańska, B.; Nowak, W.; Komisarek, J.; Taciak, M.; Barszcz, M.; Skomiał, J. Prevalence and consequence of subacute ruminal acidosis in Polish dairy herds. J. Anim. Physiol. Anim. Nutr. 2017, 101, 694–702. [Google Scholar] [CrossRef]
- Zhang, C.; Zhao, L. Strain-level dissection of the contribution of the gut microbiome to human metabolic disease. Genome Med. 2016, 8, 41. [Google Scholar] [CrossRef]
- Counotte, G.H.M.; Prins, R.A.; Janssen, R.H.A.M.; Debie, M.J.A. Role of Megasphaera elsdenii in the fermentation of DL-[2-13C] lactate in the rumen of dairy cattle. Appl. Environ. Microbiol. 1981, 42, 649–655. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Xia, G.; Jin, Y.; Wang, H. Ambient pH regulates lactate catabolism pathway of the ruminal Megasphaera elsdenni BE2-2083 and Selenomonas ruminantium HD4. J. Appl. Microbiol. 2021, 132, 2661–2672. [Google Scholar] [CrossRef] [PubMed]
- Harlow, B.E.; Flythe, M.D.; Kagan, I.A.; Goodman, J.P.; Klotz, J.L.; Aiken, G.E. Isoflavone supplementation, via red clover hay, alters the rumen microbial community and promotes weight gain of steers grazing mixed grass pastures. PLoS ONE 2020, 15, e0229200. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| VFA, mmol L−1 | Time, h | p-Values | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2 | 4 | 8 | 24 | SEM | Species | TRT | Time | Species × TRT × Time | |||||
| Bovine | Ovine | Bovine | Ovine | Bovine | Ovine | Bovine | Ovine | ||||||
| Acetate | 0.66 | <0.01 | <0.01 | 0.74 | |||||||||
| HY | 0.63 | 0.33 | 2.96 | 3.81 | 11.70 | 13.16 | 29.63 | 28.82 | 1.21 | ||||
| HC | 1.92 | 0.00 | 5.00 | 2.35 | 11.77 | 11.19 | 30.66 | 27.75 | 1.21 | ||||
| CH | 0.86 | 0.00 | 0.40 | 2.98 | 8.29 | 9.19 | 30.74 | 28.73 | 1.21 | ||||
| CN | 0.33 | 0.00 | 0.87 | 1.38 | 6.76 | 6.03 | 22.75 | 24.70 | 1.21 | ||||
| Propionate | 0.08 | <0.01 | <0.01 | <0.01 | |||||||||
| HY | 0.00 γ | 0.76 γ | 1.98 β | 4.22 β,γ | 10.17 β | 11.14 β | 27.55 α | 25.42 α | 1.17 | ||||
| HC | 0.33 γ | 0.88 γ | 2.53 β,γ | 3.16 β,γ | 10.19 β | 9.70 β | 37.74 a,α | 27.71 b,α | 1.17 | ||||
| CH | 0.00 γ | 1.04 γ | 1.76 β,γ | 3.76 β,γ | 8.95 β | 8.85 β | 42.87 a,α | 29.74 b,α | 1.17 | ||||
| CN | 0.00 γ | 0.33 γ | 1.34 β,γ | 2.45 β,γ | 8.48 β | 6.81 β | 33.17 a,α | 28.12 b,α | 1.17 | ||||
| Butyrate | 0.21 | 0.41 | <0.01 | <0.01 | |||||||||
| HY | 0.00 β | 0.00 β | 0.67 β | 1.00 β | 1.20 β | 1.46 β | 4.79 α | 5.19 α | 0.48 | ||||
| HC | 0.33 β | 0.00 β | 0.33 β | 0.53 β | 1.24 β | 1.37 β | 5.23 y,α | 6.52 x,α | 0.48 | ||||
| CH | 0.00 γ | 0.00 β | 0.00 γ | 1.00 β | 1.19 β | 1.28 β | 5.43 b,α | 8.17 a,α | 0.48 | ||||
| CN | 0.00 γ | 0.00 β | 0.67 β,γ | 0.67 β | 1.30 β | 1.02 β | 5.06 b,α | 7.46 a,α | 0.48 | ||||
| Total VFA | 0.07 | <0.01 | <0.01 | <0.01 | |||||||||
| HY | 1.63 δ | 1.76 δ | 8.54 γ | 10.36 γ | 23.74 β | 25.76 β | 63.53 α | 60.42 α | 1.99 | ||||
| HC | 3.92 δ | 1.88 γ | 10.46 γ | 7.26 γ | 24.35 β | 22.62 β | 74.69 a,α | 63.39 b,α | 1.99 | ||||
| CH | 1.53 γ | 2.04 γ | 3.47 γ | 8.75 γ | 19.70 β | 20.07 β | 80.09 a,α | 67.65 b,α | 1.99 | ||||
| CN | 0.33 γ | 0.67 γ | 4.21 γ | 4.49 γ | 17.96 β | 14.87 β | 61.98 α | 61.29 α | 1.99 | ||||
| Treatment | Species | p-Values | |||
|---|---|---|---|---|---|
| Bovine | Ovine | Species | TRT | Species × TRT | |
| HY | 10.37 ± 0.78 a | 8.11 ± 0.78 b | 0.01 | <0.01 | <0.01 |
| HC | 3.25 ± 1.04 b | 6.94 ± 0.78 a | |||
| CH | 2.31 ± 0.78 b | 6.65 ± 0.84 a | |||
| CN | 2.72 ± 0.84 | 3.76 ± 0.78 | |||
| Functional Guilds, Cells mL−1 | Species | Statistics | ||
|---|---|---|---|---|
| Bovine | Ovine | SEM | p-Value | |
| Amylolytic | 2.15 × 107 | 1.00 × 107 | 0.47 | 0.64 |
| GPC † | 5.03 × 104 x | 9.90 × 102 y | 0.51 | 0.08 |
| Lactate-Utilizing | 1.00 × 107 | 2.15 × 107 | 0.38 | 0.37 |
| Cellulolytic | 1.00 × 105 b | 4.64 × 106 a | 0.24 | <0.01 |
| HAB ‡ | 4.64 × 107 x | 4.64 × 106 y | 0.33 | 0.10 |
| Functional Guilds, Cells mL−1 | Time, h | p-Values | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 8 | 24 | SEM (Log Transformed) | Species | TRT | Time | Species × TRT × Time | |||
| Bovine | Ovine | Bovine | Ovine | ||||||
| Amylolytic | <0.01 | 0.05 | 0.20 | 0.10 | |||||
| HY | 2.15 × 1010 a | 2.15 × 108 b | 4.64 × 109 a | 1.00 × 108 b | 0.25 | ||||
| HC | 1.00 × 109 | 1.00 × 109 | 2.15 × 109 a | 2.15 × 108 b | 0.25 | ||||
| CH | 1.00 × 109 α,a | 1.00 × 108 b | 1.00 × 1010 β,a | 1.00 × 108 b | 0.25 | ||||
| CN | 4.64 × 108 a | 4.64 × 107 β,b | 2.15 × 109 x | 4.64 × 108 α,y | 0.25 | ||||
| GPC † | <0.01 | 0.11 | <0.01 | <0.01 | |||||
| HY | 4.45 × 106 a | 1.95 × 103 β,b | 7.79 × 106 | 1.81 × 105 α | 0.61 | ||||
| HC | 6.01 × 105 a | 1.21 × 103 β,b | 2.40 × 106 | 1.80 × 105 α | 0.61 | ||||
| CH | 1.08 × 108 a | 8.64 × 102 β,b | 1.38 × 107 | 1.29 × 106 α | 0.61 | ||||
| CN | 3.13 × 106 a | 8.26 × 102 β,b | 8.30 × 106 | 4.42 × 105 α | 0.61 | ||||
| Lactate-Utilizing | <0.01 | 0.04 | 0.08 | 0.12 | |||||
| HY | 2.15 × 108 | 1.00 × 108 | 2.15 × 109 | 4.64 × 107 | 0.28 | ||||
| HC | 4.64 × 108 | 4.64 × 107 | 4.64 × 108 | 2.15 × 107 | 0.28 | ||||
| CH | 2.15 × 108 | 4.64 × 107 | 1.00 × 109 | 4.64 × 107 | 0.28 | ||||
| CN | 1.00 × 109 | 4.64 × 107 | 1.00 × 1010 | 4.64 × 107 | 0.28 | ||||
| Cellulolytic | 0.08 | <0.01 | <0.01 | 0.09 | |||||
| HY | 4.64 × 105 | 2.15 × 105 | 4.64 × 105 | 1.00 × 106 | 0.29 | ||||
| HC | 4.64 × 105 α | 4.64 × 105 | 4.64 × 104 β,b | 4.64 × 105 a | 0.29 | ||||
| CH | 4.64 × 105 α | 4.64 × 106 | 2.15 × 104 β,b | 4.64 × 105 a | 0.29 | ||||
| CN | 4.64 × 104 α | 1.00 × 105 α | 2.15 × 103 β | 1.00 × 104 β | 0.29 | ||||
| HAB ‡ | <0.01 | 0.24 | 0.08 | 0.02 | |||||
| HY | 1.00 × 1010 a | 4.64 × 107 b | 2.15 × 109 a | 2.15 × 107 b | 0.36 | ||||
| HC | 1.00 × 107 β,a | 2.15 × 108 b | 2.15 × 109 α,a | 1.00 × 108 b | 0.36 | ||||
| CH | 2.15 × 109 a | 1.00 × 108 b | 2.15 × 109 a | 1.00 × 108 b | 0.36 | ||||
| CN | 1.00 × 109 a | 2.15 × 107 b | 4.64 × 109 a | 1.00 × 108 b | 0.36 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weinert-Nelson, J.R.; Ely, D.G.; Flythe, M.D.; Hamilton, T.A.; Ferrell, J.L.; Davis, B.E. Ex Vivo Fermentation of Hay and Corn by Rumen Bacteria from Cattle and Sheep. Fermentation 2023, 9, 929. https://doi.org/10.3390/fermentation9110929
Weinert-Nelson JR, Ely DG, Flythe MD, Hamilton TA, Ferrell JL, Davis BE. Ex Vivo Fermentation of Hay and Corn by Rumen Bacteria from Cattle and Sheep. Fermentation. 2023; 9(11):929. https://doi.org/10.3390/fermentation9110929
Chicago/Turabian StyleWeinert-Nelson, Jennifer R., Donald G. Ely, Michael D. Flythe, Tracy A. Hamilton, Jessica L. Ferrell, and Brittany E. Davis. 2023. "Ex Vivo Fermentation of Hay and Corn by Rumen Bacteria from Cattle and Sheep" Fermentation 9, no. 11: 929. https://doi.org/10.3390/fermentation9110929
APA StyleWeinert-Nelson, J. R., Ely, D. G., Flythe, M. D., Hamilton, T. A., Ferrell, J. L., & Davis, B. E. (2023). Ex Vivo Fermentation of Hay and Corn by Rumen Bacteria from Cattle and Sheep. Fermentation, 9(11), 929. https://doi.org/10.3390/fermentation9110929