
Limosilactobacillus reuteri Regulating Intestinal Function: A Review


Abstract
1. Source and Diversity of L. Reuteri
2. Physiological Role of L. Reuteri
2.1. Improvement of Intestinal Physiological Function
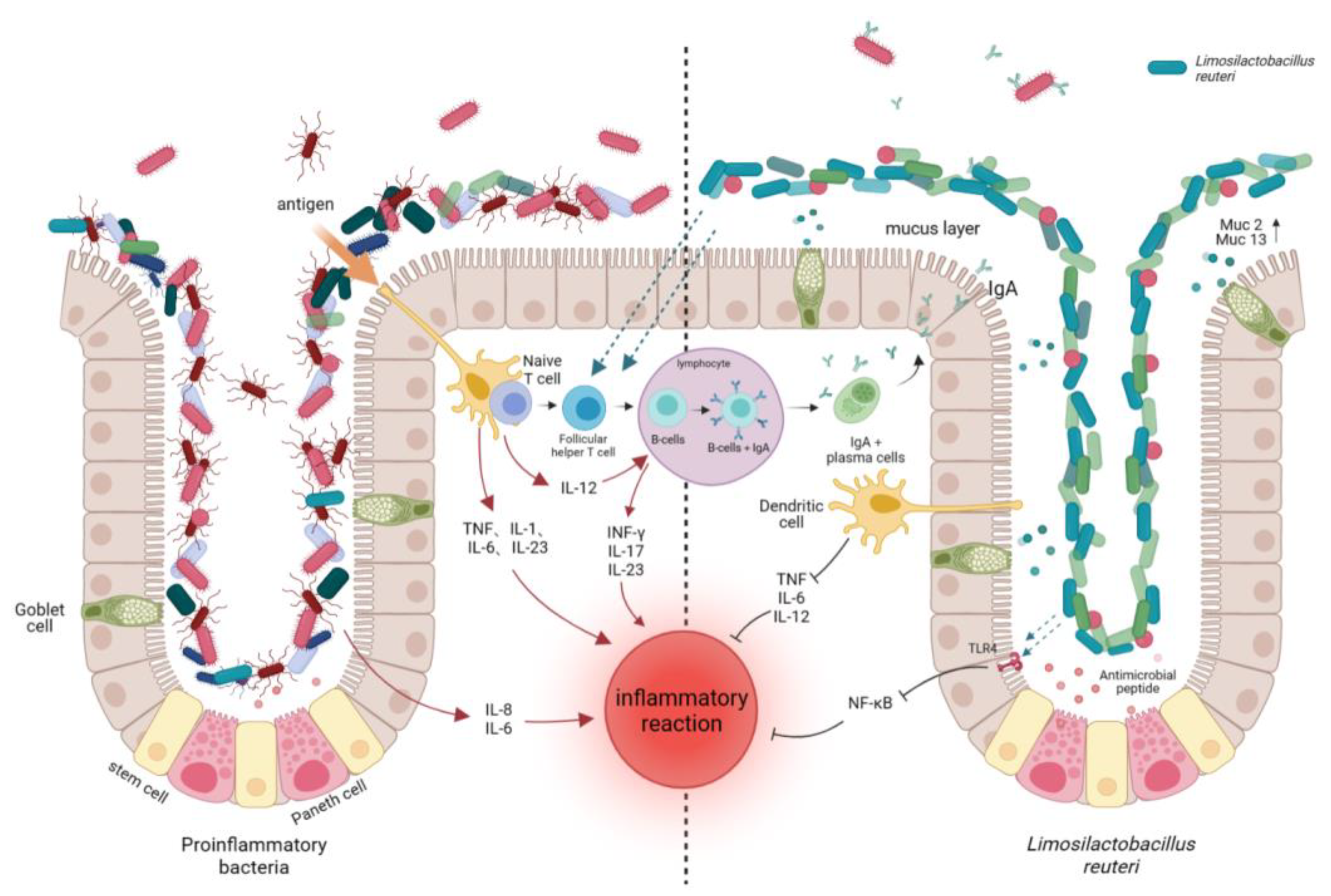
2.1.1. Modulation of Gut Flora
2.1.2. Improvement of Intestinal Structure
2.2. Inhibition of the Growth and Colonization of Intestinal Pathogenic Bacteria
2.2.1. Production of Antimicrobial Metabolites
2.2.2. Site Competition with Pathogenic Bacteria through Colonization
2.2.3. Promotion of Mucus Secretion and Reduction in the Colonization of Harmful Microorganisms
2.3. Immunomodulatory Effects
2.3.1. Influence on the Production of Immunoglobulin A
2.3.2. Regulation of TREG Cells
2.3.3. Inhibition of the Production of Pro-inflammatory Factors
2.3.4. More Studies
2.4. Amelioration of Obesity
2.5. Other Functions
3. Application of L. Reuteri
3.1. Application in the Feed Industry
3.2. Application in Functional Food
3.3. Clinical Application
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mu, Q.; Tavella, V.J.; Luo, X.M. Role of Lactobacillus reuteri in human health and diseases. Front. Microbiol. 2018, 9, 757. [Google Scholar] [CrossRef] [PubMed]
- Duar, R.M.; Frese, S.A.; Lin, X.B.; Fernando, S.C.; Burkey, T.E.; Tasseva, G.; Peterson, D.A.; Blom, J.; Wenzel, C.Q.; Szymanski, C.M.; et al. Experimental evaluation of host adaptation of Lactobacillus reuteri to different vertebrate species. Appl. Environ. Microbiol. 2017, 83, e00132-17. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Chen, L.; Chen, L.; Ren, X.; Ge, H.; Li, B.; Ma, G.; Ke, X.; Zhu, J.; Li, L.; et al. Potential probiotic characterization of Lactobacillus reuteri from traditional Chinese highland barley wine and application for room-temperature-storage drinkable yogurt. J. Dairy Sci. 2018, 101, 5780–5788. [Google Scholar] [CrossRef] [PubMed]
- İspirli, H.; Şimşek, Ö.; Skory, C.; Sağdıç, O.; Dertli, E. Characterization of a 4, 6-α-glucanotransferase from Lactobacillus reuteri E81 and production of malto-oligosaccharides with immune-modulatory roles. Int. J. Biol. Macromol. 2019, 124, 1213–1219. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Tao, Q.; Teixeira, J.S.; Su, M.S.-W.; Gänzle, M.G. Contribution of glutaminases to glutamine metabolism and acid resistance in Lactobacillus reuteri and other vertebrate host adapted lactobacilli. Food Microbiol. 2020, 86, 103343. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Siepmann, F.B.; Tovar, L.E.R.; Chen, X.; Gänzle, M.G. Effect of copy number of the spoVA2mob operon, sourdough and reutericyclin on ropy bread spoilage caused by Bacillus spp. Food Microbiol. 2020, 91, 103507. [Google Scholar] [CrossRef]
- Hu, Z.; Jia, P.; Bai, Y.; Fan, T.-P.; Zheng, X.; Cai, Y. Characterisation of five alcohol dehydrogenases from Lactobacillus reuteri DSM20016. Process Biochem. 2019, 86, 73–79. [Google Scholar] [CrossRef]
- Zabed, H.M.; Zhang, Y.; Guo, Q.; Yun, J.; Yang, M.; Zhang, G.; Qi, X. Co-biosynthesis of 3-hydroxypropionic acid and 1,3-propanediol by a newly isolated Lactobacillus reuteri strain during whole cell biotransformation of glycerol. J. Clean. Prod. 2019, 226, 432–442. [Google Scholar] [CrossRef]
- Garg, S.; Singh, T.P.; Malik, R.K. In vivo implications of potential probiotic Lactobacillus reuteri lr6 on the gut and immunological parameters as an adjuvant against protein energy malnutrition. Probiotics Antimicrob. Proteins 2020, 12, 517–534. [Google Scholar] [CrossRef]
- Turco, R.; Russo, M.; Bruzzese, D.; Staiano, A. Efficacy of a partially hydrolysed formula, with reduced lactose content and with Lactobacillus reuteri DSM 17938 in infant colic: A double blind, randomised clinical trial. Clin. Nutr. 2021, 40, 412–419. [Google Scholar] [CrossRef]
- Morita, H.; Toh, H.; Fukuda, S.; Horikawa, H.; Oshima, K.; Suzuki, T.; Murakami, M.; Hisamatsu, S.; Kato, Y.; Takizawa, T.; et al. Comparative genome analysis of Lactobacillus reuteri and Lactobacillus fermentum reveal a genomic island for reuterin and cobalamin production. DNA Res. 2008, 15, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Choi, S.H.; Kim, Y.J.; Jeong, H.J.; Ryu, J.S.; Lee, H.J.; Kim, T.W.; Im, S.-H.; Oh, J.Y.; Kim, M.K. Clinical effect of IRT-5 probiotics on immune modulation of autoimmunity or alloimmunity in the eye. Nutrients 2017, 9, 1166. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Huang, Y.; Cai, W.; Li, D.; Zheng, W.; Xiao, Y.; Zhao, H.; Pan, S. Effect of oral Lactobacillus rhamnosus GR-1 and Lactobacillus reuteri RC-14 on vaginal Group B Streptococcus colonization and vaginal microbiome in late pregnancy. Nan Fang Yi Ke Da Xue Xue Bao = J. South. Med. Univ. 2020, 40, 1753–1759. [Google Scholar] [CrossRef]
- Yamato, M.; Nakada, R.; Nakamura, Y. Release of spirosin associated with potassium phosphate-induced autolysis in Lactobacillus reuteri DSM 20016. Microbiol. Res. 1998, 153, 29–35. [Google Scholar] [CrossRef]
- Jones, M.L.; Martoni, C.J.; Prakash, S. Oral supplementation with probiotic L. reuteri NCIMB 30242 increases mean circulating 25-hydroxyvitamin D: A post hoc analysis of a randomized controlled trial. J. Clin. Endocrinol. Metab. 2013, 98, 2944–2951. [Google Scholar] [CrossRef]
- Wells, J.G.; Davis, B.; Wachsmuth, I.; Riley, L.W.; Remis, R.S.; Sokolow, R.; Morris, G.K. Laboratory investigation of hemorrhagic colitis outbreaks associated with a rare Escherichia coli serotype. J. Clin. Microbiol. 1983, 18, 512–520. [Google Scholar] [CrossRef]
- Zhang, D.; Liu, H.; Wang, S.; Zhang, W.; Wang, J.; Tian, H.; Wang, Y.; Ji, H. Fecal microbiota and its correlation with fatty acids and free amino acids metabolism in piglets after a Lactobacillus strain oral administration. Front. Microbiol. 2019, 10, 785. [Google Scholar] [CrossRef]
- Bajpai, V.K.; Chandra, V.; Kim, N.-H.; Rai, R.; Kumar, P.; Kim, K.; Aeron, A.; Kang, S.C.; Maheshwari, D.; Na, M.; et al. Ghost probiotics with a combined regimen: A novel therapeutic approach against the Zika virus, an emerging world threat. Crit. Rev. Biotechnol. 2018, 38, 438–454. [Google Scholar] [CrossRef]
- Li, X.; Yue, L.; Guan, X.; Qiao, S. The adhesion of putative probiotic lactobacilli to cultured epithelial cells and porcine intestinal mucus. J. Appl. Microbiol. 2008, 104, 1082–1091. [Google Scholar] [CrossRef]
- Wang, G.; Chen, Y.; Fei, S.; Xie, C.; Xia, Y.; Ai, L. Colonisation with endogenous Lactobacillus reuteri R28 and exogenous Lactobacillus plantarum AR17-1 and the effects on intestinal inflammation in mice. Food Funct. 2021, 12, 2481–2488. [Google Scholar] [CrossRef]
- Leonard, M.T.; Valladares, R.B.; Ardissone, A.; Gonzalez, C.F.; Lorca, G.L.; Triplett, E.W. Complete genome sequences of Lactobacillus johnsonii strain N6.2 and Lactobacillus reuteri strain TD1. Genome Announc. 2014, 2, e00397-14. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Kang, Z.; Zhou, J.; Chen, J.; Du, G. High-level expression and characterization of recombinant acid urease for enzymatic degradation of urea in rice wine. Appl. Microbiol. Biotechnol. 2015, 99, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Klein, G. Antibiotic resistance and molecular characterization of probiotic and clinical Lactobacillus strains in relation to safety aspects of probiotics. Foodborne Pathog. Dis. 2011, 8, 267–281. [Google Scholar] [CrossRef] [PubMed]
- Jatuponwiphat, T.; Namrak, T.; Supataragul, A.; Nitisinprasert, S.; Nakphaichit, M.; Vongsangnak, W. Comparative genome analysis reveals metabolic traits associated with probiotics properties in Lactobacillus reuteri KUB-AC5. Gene Rep. 2019, 17, 100536. [Google Scholar] [CrossRef]
- Kim, D.; Cho, M.-J.; Cho, S.; Lee, Y.; Byun, S.J.; Lee, S. Complete genome sequence of Lactobacillus reuteri SKKU-OGDONS-01, isolated from a chicken’s small intestine. Microbiol. Resour. Announc. 2018, 7, e01251-18. [Google Scholar] [CrossRef]
- Hu, R.; Lin, H.; Wang, M.; Zhao, Y.; Liu, H.; Min, Y.; Yang, X.; Gao, Y.; Yang, M. Lactobacillus reuteri-derived extracellular vesicles maintain intestinal immune homeostasis against lipopolysaccharide-induced inflammatory responses in broilers. J. Anim. Sci. Biotechnol. 2021, 12, 1–18. [Google Scholar] [CrossRef]
- Lv, M.; Lei, Q.; Yin, H.; Hu, T.; Wang, S.; Dong, K.; Pan, H.; Liu, Y.; Lin, Q.; Cao, Z. Effects of Prebiotics and Synbiotics on Gut Microbiota. Pol. J. Microbiol. 2021, 70, 511–520. [Google Scholar] [CrossRef]
- Chen, L.-H.; Chen, Y.-H.; Cheng, K.-C.; Chien, T.-Y.; Chan, C.-H.; Tsao, S.-P.; Huang, H.-Y. Antiobesity effect of Lactobacillus reuteri 263 associated with energy metabolism remodeling of white adipose tissue in high-energy-diet-fed rats. J. Nutr. Biochem. 2018, 54, 87–94. [Google Scholar] [CrossRef]
- Britton, R.A.; Irwin, R.; Quach, D.; Schaefer, L.; Zhang, J.; Lee, T.; Parameswaran, N.; McCabe, L.R. Probiotic L. reuteri treatment prevents bone loss in a menopausal ovariectomized mouse model. J. Cell. Physiol. 2014, 229, 1822–1830. [Google Scholar] [CrossRef]
- Ding, Y.-H.; Qian, L.-Y.; Pang, J.; Lin, J.-Y.; Xu, Q.; Wang, L.-H.; Huang, D.-S.; Zou, H. The regulation of immune cells by Lactobacilli: A potential therapeutic target for anti-atherosclerosis therapy. Oncotarget 2017, 8, 59915. [Google Scholar] [CrossRef]
- Jones, S.E.; Versalovic, J. Probiotic Lactobacillus reuteribiofilms produce antimicrobial and anti-inflammatory factors. BMC Microbiol. 2009, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Li, Y.; Xiao, H.; Shi, Y.; Le, G.W.; Sun, J. Isolation of Lactobacillus reuteri from Peyer’s patches and their effects on sIgA production and gut microbiota diversity. Mol. Nutr. Food Res. 2016, 60, 2020–2030. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Tran, D.Q.; Fatheree, N.Y.; Marc Rhoads, J. Lactobacillus reuteri DSM 17938 differentially modulates effector memory T cells and Foxp3+ regulatory T cells in a mouse model of necrotizing enterocolitis. Am. J. Physiol.-Gastrointest. Liver Physiol. 2014, 307, G177–G186. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Sun, Q.; Wang, J.; Qiu, X.; Qi, R.; Huang, J. Identification of differentially expressed miRNAs after Lactobacillus reuteri treatment in the ileum mucosa of piglets. Genes Genom. 2020, 42, 1327–1338. [Google Scholar] [CrossRef]
- Yang, F.; Wang, A.; Zeng, X.; Hou, C.; Liu, H.; Qiao, S. Lactobacillus reuteri I5007 modulates tight junction protein expression in IPEC-J2 cells with LPS stimulation and in newborn piglets under normal conditions. BMC Microbiol. 2015, 15, 1–11. [Google Scholar] [CrossRef]
- Liu, H.; Hou, C.; Wang, G.; Jia, H.; Yu, H.; Zeng, X.; Thacker, P.A.; Zhang, G.; Qiao, S. Lactobacillus reuteri I5007 modulates intestinal host defense peptide expression in the model of IPEC-J2 cells and neonatal piglets. Nutrients 2017, 9, 559. [Google Scholar] [CrossRef]
- Zheng, J.; Ruan, L.; Sun, M.; Gänzle, M. A genomic view of lactobacilli and pediococci demonstrates that phylogeny matches ecology and physiology. Appl. Environ. Microbiol. 2015, 81, 7233–7243. [Google Scholar] [CrossRef]
- El-Ziney, M.; Van Den Tempel, T.; Debevere, J.; Jakobsen, M. Application of reuterin produced by Lactobacillus reuteri 12002 for meat decontamination and preservation. J. Food Prot. 1999, 62, 257–261. [Google Scholar] [CrossRef]
- Talarico, T.; Casas, I.; Chung, T.C.; Dobrogosz, W. Production and isolation of reuterin, a growth inhibitor produced by Lactobacillus reuteri. Antimicrob. Agents Chemother. 1988, 32, 1854–1858. [Google Scholar] [CrossRef]
- Vollenweider, S.; Evers, S.; Zurbriggen, K.; Lacroix, C. Unraveling the hydroxypropionaldehyde (HPA) system: An active antimicrobial agent against human pathogens. J. Agric. Food Chem. 2010, 58, 10315–10322. [Google Scholar] [CrossRef]
- Chung, T.; Axelsson, L.; Lindgren, S.; Dobrogosz, W. In vitro studies on reuterin synthesis by Lactobacillus reuteri. Microb. Ecol. Health Dis. 1989, 2, 137–144. [Google Scholar] [CrossRef]
- Greifová, G.; Májeková, H.; Greif, G.; Body, P.; Greifová, M.; Dubničková, M. Analysis of antimicrobial and immunomodulatory substances produced by heterofermentative Lactobacillus reuteri. Folia Microbiol. 2017, 62, 515–524. [Google Scholar] [CrossRef] [PubMed]
- Ganzle, M.G.; Holtzel, A.; Walter, J.; Jung, G.N.; Hammes, W.P. Characterization of reutericyclin produced by Lactobacillus reuteri LTH2584. Appl. Environ. Microbiol. 2000, 66, 4325–4333. [Google Scholar] [CrossRef]
- Kabuki, T.; Saito, T.; Kawai, Y.; Uemura, J.; Itoh, T. Production, purification and characterization of reutericin 6, a bacteriocin with lytic activity produced by Lactobacillus reuteri LA6. Int. J. Food Microbiol. 1997, 34, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Toba, T.; Samant, S.; Yoshioka, E.; Itoh, T. Reutericin 6, a new bacteriocin produced by Lactobacillus reuteri LA 6. Lett. Appl. Microbiol. 1991, 13, 281–286. [Google Scholar] [CrossRef]
- Arakawa, K.; Kawai, Y.; Ito, Y.; Nakamura, K.; Chujo, T.; Nishimura, J.; Kitazawa, H.; Saito, T. HPLC purification and re-evaluation of chemical identity of two circular bacteriocins, gassericin A and reutericin 6. Lett. Appl. Microbiol. 2010, 50, 406–411. [Google Scholar] [CrossRef] [PubMed]
- Maqueda, M.; Sánchez-Hidalgo, M.; Fernández, M.; Montalbán-López, M.; Valdivia, E.; Martínez-Bueno, M. Genetic features of circular bacteriocins produced by Gram-positive bacteria. FEMS Microbiol. Rev. 2008, 32, 2–22. [Google Scholar] [CrossRef]
- Bøhle, L.A.; Brede, D.A.; Diep, D.B.; Holo, H.; Nes, I.F. Specific degradation of the mucus adhesion-promoting protein (MapA) of Lactobacillus reuteri to an antimicrobial peptide. Appl. Environ. Microbiol. 2010, 76, 7306–7309. [Google Scholar] [CrossRef]
- Seo, B.J.; Mun, M.R.; Kim, C.-J.; Lee, I.; Chang, Y.-H.; Park, Y.-H. Bile tolerant Lactobacillus reuteri isolated from pig feces inhibits enteric bacterial pathogens and porcine rotavirus. Vet. Res. Commun. 2010, 34, 323–333. [Google Scholar] [CrossRef]
- Moussavi, M.; Adams, M.C. An in vitro study on bacterial growth interactions and intestinal epithelial cell adhesion characteristics of probiotic combinations. Curr. Microbiol. 2010, 60, 327–335. [Google Scholar] [CrossRef]
- Saulnier, D.M.; Santos, F.; Roos, S.; Mistretta, T.-A.; Spinler, J.K.; Molenaar, D.; Teusink, B.; Versalovic, J. Exploring metabolic pathway reconstruction and genome-wide expression profiling in Lactobacillus reuteri to define functional probiotic features. PLoS ONE 2011, 6, e18783. [Google Scholar] [CrossRef] [PubMed]
- Singh, T.P.; Kaur, G.; Kapila, S.; Malik, R.K. Antagonistic activity of Lactobacillus reuteri strains on the adhesion characteristics of selected pathogens. Front. Microbiol. 2017, 8, 486. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, G.P.; Lee, S.M.; Mazmanian, S.K. Gut biogeography of the bacterial microbiota. Nat. Rev. Microbiol. 2016, 14, 20–32. [Google Scholar] [CrossRef] [PubMed]
- Son, Y.S.; Ki, S.J.; Thanavel, R.; Kim, J.J.; Lee, M.O.; Kim, J.; Jung, C.R.; Han, T.S.; Cho, H.S.; Ryu, C.M.; et al. Maturation of human intestinal organoids in vitro facilitates colonization by commensal lactobacilli by reinforcing the mucus layer. FASEB J. 2020, 34, 9899–9910. [Google Scholar] [CrossRef]
- Wang, G.; Huang, S.; Cai, S.; Yu, H.; Wang, Y.; Zeng, X.; Qiao, S. Lactobacillus reuteri ameliorates intestinal inflammation and modulates gut microbiota and metabolic disorders in dextran sulfate sodium-induced colitis in mice. Nutrients 2020, 12, 2298. [Google Scholar] [CrossRef]
- Burgueño, J.F.; Abreu, M.T. Epithelial Toll-like receptors and their role in gut homeostasis and disease. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 263–278. [Google Scholar] [CrossRef]
- Li, L.; Fang, Z.; Liu, Z.; Zhao, J.; Zhang, H.; Wang, S.; He, J.; Lu, W.; Chen, W. Lactobacillus reuteri CCFM1072 and CCFM1040 with the role of Treg cells regulation alleviate airway inflammation through modulating gut microbiota in allergic asthma mice. J. Funct. Foods 2021, 76, 104286. [Google Scholar] [CrossRef]
- Liu, Y.; Fatheree, N.Y.; Mangalat, N.; Rhoads, J.M. Lactobacillus reuteri strains reduce incidence and severity of experimental necrotizing enterocolitis via modulation of TLR4 and NF-κB signaling in the intestine. Am. J. Physiol.-Gastrointest. Liver Physiol. 2012, 302, G608–G617. [Google Scholar] [CrossRef]
- Lu, P.; Sodhi, C.P.; Hackam, D.J. Toll-like receptor regulation of intestinal development and inflammation in the pathogenesis of necrotizing enterocolitis. Pathophysiology 2014, 21, 81–93. [Google Scholar] [CrossRef]
- Zegarra-Ruiz, D.F.; El Beidaq, A.; Iñiguez, A.J.; Di Ricco, M.L.; Vieira, S.M.; Ruff, W.E.; Mubiru, D.; Fine, R.L.; Sterpka, J.; Greiling, T.M.; et al. A diet-sensitive commensal Lactobacillus strain mediates TLR7-dependent systemic autoimmunity. Cell Host Microbe 2019, 25, 113–127.e6. [Google Scholar] [CrossRef]
- Shornikova, A.-V.; Casas, I.A.; Mykkänen, H.; Salo, E.; Vesikari, T. Bacteriotherapy with Lactobacillus reuteri in rotavirus gastroenteritis. Pediatr. Infect. Dis. J. 1997, 16, 1103–1107. [Google Scholar] [CrossRef] [PubMed]
- Anukam, K.C.; Hayes, K.; Summers, K.; Reid, G. Probiotic Lactobacillus rhamnosus GR-1 and Lactobacillus reuteri RC-14 may help downregulate TNF-Alpha, IL-6, IL-8, IL-10 and IL-12 (p70) in the neurogenic bladder of spinal cord injured patient with urinary tract infections: A two-case study. Adv. Urol. 2009, 2009, 680363. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hsieh, F.-C.; Lan, C.-C.E.; Huang, T.-Y.; Chen, K.-W.; Chai, C.-Y.; Chen, W.-T.; Fang, A.-H.; Chen, Y.-H.; Wu, C.-S. Heat-killed and live Lactobacillus reuteri GMNL-263 exhibit similar effects on improving metabolic functions in high-fat diet-induced obese rats. Food Funct. 2016, 7, 2374–2388. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Fatheree, N.Y.; Mangalat, N.; Rhoads, J.M. Human-derived probiotic Lactobacillus reuteri strains differentially reduce intestinal inflammation. Am. J. Physiol.-Gastrointest. Liver Physiol. 2010, 299, G1087–G1096. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, E.; Serata, M.; Sako, T. Suppressive effect on activation of macrophages by Lactobacillus casei strain Shirota genes determining the synthesis of cell wall-associated polysaccharides. Appl. Environ. Microbiol. 2008, 74, 4746–4755. [Google Scholar] [CrossRef]
- Hajishengallis, G.; Connell, T.D. Type II heat-labile enterotoxins: Structure, function, and immunomodulatory properties. Vet. Immunol. Immunopathol. 2013, 152, 68–77. [Google Scholar] [CrossRef]
- Kšonžeková, P.; Bystrický, P.; Vlčková, S.; Pätoprstý, V.; Pulzová, L.; Mudroňová, D.; Kubašková, T.; Csank, T.; Tkáčiková, Ľ. Exopolysaccharides of Lactobacillus reuteri: Their influence on adherence of E. coli to epithelial cells and inflammatory response. Carbohydr. Polym. 2016, 141, 10–19. [Google Scholar] [CrossRef]
- Tkáčiková, Ľ.; Mochnáčová, E.; Tyagi, P.; Kiššová, Z.; Bhide, M. Comprehensive mapping of the cell response to E. coli infection in porcine intestinal epithelial cells pretreated with exopolysaccharide derived from Lactobacillus reuteri. Vet. Res. 2020, 51, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Sims, I.M.; Frese, S.A.; Walter, J.; Loach, D.; Wilson, M.; Appleyard, K.; Eason, J.; Livingston, M.; Baird, M.; Cook, G. Structure and functions of exopolysaccharide produced by gut commensal Lactobacillus reuteri 100-23. ISME J. 2011, 5, 1115–1124. [Google Scholar] [CrossRef] [PubMed]
- Round, J.L.; Mazmanian, S.K. Inducible Foxp3+ regulatory T-cell development by a commensal bacterium of the intestinal microbiota. Proc. Natl. Acad. Sci. USA 2010, 107, 12204–12209. [Google Scholar] [CrossRef]
- Hoffmann, M.; Rath, E.; Hölzlwimmer, G.; Quintanilla-Martinez, L.; Loach, D.; Tannock, G.; Haller, D. Lactobacillus reuteri 100-23 transiently activates intestinal epithelial cells of mice that have a complex microbiota during early stages of colonization. J. Nutr. 2008, 138, 1684–1691. [Google Scholar] [CrossRef] [PubMed]
- Konstantinov, S.R.; Smidt, H.; de Vos, W.M.; Bruijns, S.C.; Singh, S.K.; Valence, F.; Molle, D.; Lortal, S.; Altermann, E.; Klaenhammer, T.R.; et al. S layer protein A of Lactobacillus acidophilus NCFM regulates immature dendritic cell and T cell functions. Proc. Natl. Acad. Sci. USA 2008, 105, 19474–19479. [Google Scholar] [CrossRef] [PubMed]
- Diaz, M.; Ladero, V.; Del Rio, B.; Redruello, B.; Fernández, M.; Martin, M.C.; Alvarez, M.A. Biofilm-forming capacity in biogenic amine-producing bacteria isolated from dairy products. Front. Microbiol. 2016, 7, 591. [Google Scholar] [CrossRef] [PubMed]
- Zelante, T.; Iannitti, R.G.; Cunha, C.; De Luca, A.; Giovannini, G.; Pieraccini, G.; Zecchi, R.; D’Angelo, C.; Massi-Benedetti, C.; Fallarino, F.; et al. Tryptophan catabolites from microbiota engage aryl hydrocarbon receptor and balance mucosal reactivity via interleukin-22. Immunity 2013, 39, 372–385. [Google Scholar] [CrossRef] [PubMed]
- Koay, K.-P.; Tsai, B.C.-K.; Kuo, C.-H.; Kuo, W.-W.; Luk, H.-N.; Day, C.H.; Chen, R.-J.; Chen, M.Y.-C.; Padma, V.V.; Huang, C.-Y. Hyperglycemia-Induced Cardiac Damage Is Alleviated by Heat-Inactivated Lactobacillus reuteri GMNL-263 via Activation of the IGF1R Survival Pathway. Probiotics Antimicrob. Proteins 2021, 13, 1044–1053. [Google Scholar] [CrossRef]
- Ortiz-Rivera, Y.; Sánchez-Vega, R.; Gutiérrez-Méndez, N.; León-Félix, J.; Acosta-Muñiz, C.; Sepulveda, D. Production of reuterin in a fermented milk product by Lactobacillus reuteri: Inhibition of pathogens, spoilage microorganisms, and lactic acid bacteria. J. Dairy Sci. 2017, 100, 4258–4268. [Google Scholar] [CrossRef]
- Chang, Y.-H.; Kim, J.-K.; Kim, H.-J.; Kim, W.-Y.; Kim, Y.-B.; Park, Y.-H. Selection of a potential probiotic Lactobacillus strain and subsequent in vivo studies. Antonie Van Leeuwenhoek 2001, 80, 193–199. [Google Scholar] [CrossRef]
- Zhao, P.Y.; Kim, I.H. Effect of direct-fed microbial on growth performance, nutrient digestibility, fecal noxious gas emission, fecal microbial flora and diarrhea score in weanling pigs. Anim. Feed. Sci. Technol. 2015, 200, 86–92. [Google Scholar] [CrossRef]
- Shi, S.; Dong, J.; Cheng, X.; Hu, J.; Liu, Y.; He, G.; Zhang, J.; Yu, H.; Jia, L.; Zhou, D. Biological characteristics and whole-genome analysis of the potential probiotic, Lactobacillus reuteri S5. Lett. Appl. Microbiol. 2022, 74, 593–603. [Google Scholar] [CrossRef]
- Bao, X.; Xiang, S.; Chen, J.; Shi, Y.; Chen, Y.; Wang, H.; Zhu, X. Effect of Lactobacillus reuteri on vitamin B12 content and microbiota composition of furu fermentation. LWT 2019, 100, 138–143. [Google Scholar] [CrossRef]
- Chai, L.-J.; Qiu, T.; Lu, Z.-M.; Deng, Y.-J.; Zhang, X.-J.; Shi, J.-S.; Xu, Z.-H. Modulating microbiota metabolism via bioaugmentation with Lactobacillus casei and Acetobacter pasteurianus to enhance acetoin accumulation during cereal vinegar fermentation. Food Res. Int. 2020, 138, 109737. [Google Scholar] [CrossRef]
- Salmerón, I.; Thomas, K.; Pandiella, S.S. Effect of potentially probiotic lactic acid bacteria on the physicochemical composition and acceptance of fermented cereal beverages. J. Funct. Foods 2015, 15, 106–115. [Google Scholar] [CrossRef]
- Kakimoto, S.; Sumino, Y.; ichi Akiyama, S.; Nakao, Y. Purification and characterization of acid urease from Lactobacillus reuteri. Agric. Biol. Chem. 1989, 53, 1119–1125. [Google Scholar] [CrossRef][Green Version]
- Savino, F.; Garro, M.; Montanari, P.; Galliano, I.; Bergallo, M. Crying time and RORγ/FOXP3 expression in Lactobacillus reuteri DSM17938-treated infants with colic: A randomized trial. J. Pediatr. 2018, 192, 171–177.E1. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Guo, C.; Gong, F. Protective effect of Lactobacillus reuteri against oxidative stress in neonatal mice with necrotizing enterocolitis. Nan Fang Yi Ke Da Xue Xue Bao = J. South. Med. Univ. 2019, 39, 1221–1226. [Google Scholar] [CrossRef]
- Buckley, M.; Lacey, S.; Doolan, A.; Goodbody, E.; Seamans, K. The effect of Lactobacillus reuteri supplementation in Helicobacter pylori infection: A placebo-controlled, single-blind study. BMC Nutr. 2018, 4, 48. [Google Scholar] [CrossRef]


{kind=link}
| Source | Strain | Feature or Function | References | |
|---|---|---|---|---|
| Fermented foods | Highland barley wine | WHH1689 | No lactose fermentation, no post-acidification caused by storage at room temperature, high bacterial survival rate, good resistance, strong adhesion to HT-29 cells, can effectively inhibit Escherichia coli, Shigella flexneri, Salmonella β paratyphoid, and gold Staphylococcus aureus activity | [3] |
| Fermented dough | 81 | With 4,6-α-glucanotransferase properties, it can produce malto-oligosaccharides with immunomodulatory effects | [4] | |
| 100-23 | Contains three glutaminase enzymes which can promote the conversion of glutamine into glutamic acid and improve the flavor of fermented products | [5] | ||
| TMW1.656 | Resists proteolysis in dough and heat inactivation in baking, inhibits Streptomyces’ growth and extends shelf life, and produces the bacteriocin Reutericyclin | [6] | ||
| Humans | Stool | DSM0016 | Good dehydrogenation ability of ethanol and promotes the mutual conversion of aldehydes, alcohols, and ketones | [7] |
| FXZ014 | Produces higher amounts of 3-HP and 1,3-PD | [8] | ||
| LR6 | Has antagonistic activity against different targeted pathogens and, has cholesterol-lowering activity and anti-hypercholesterolemia effects | [9] | ||
| Breast milk | ATCC557 (Mother plant); DSM 17938 | Effectively reduces pathogen colonization, improves intestinal health, reconstructs intestinal flora, has the ability to convert glycerol into 3-HP and 1,3-pdo, and effectively reduces constipation in infants and young children | [10] | |
| JCM 1112T | Produces reuterin and vitamin B12 | [11] | ||
| IRT-5 | Immunomodulation of hyperimmune diseases and is a treatment for rheumatoid arthritis, atopic dermatitis, and inflammatory bowel disease | [12] | ||
| Vaginal | RC14 | Inhibits the adhesion of pathogenic bacteria, inhibits the gene expression of Staphylococcus superantigen protein, and reduces the inflammatory response caused by Candida albicans | [13] | |
| Gut | DSM20016 | Produces spirosin, reduces serum SUA levels in hyperuricemic rats, relieves the symptoms of hyperuricemia, and produces Roy’s protein | [7,14] | |
| Pigs | Manure | NCIMB 30242 | Improves gastrointestinal health, inhibits sterol absorption, increases average circulating 25-hydroxyvitamin D | [15] |
| DSMZ 20056 | Promotes murine secretion | [16] | ||
| ZLR003 | Positive effects on the regulation of feed intake, weight gain, gut microbiota composition, and immune performance in weanlings, growing–finishing sows, and lactating sows | [17] | ||
| Intestines | Probio-16 | Resistant to low pH and bile, inhibits enteric bacterial pathogens and rotavirus, has antibacterial and antiviral activity, and has an inhibitory effect on the Zika virus | [18] | |
| I5007 | Interacts with Caco-2 cells, has strong adhesion to porcine intestinal mucus and the host cells IPEC-J2 and IEC-6, can produce large amounts of exopolysaccharides, inhibits Salmonella Typhimurium and Escherichia coli growth, and positively regulates piglet immune function | [19] | ||
| Mice | Droppings | R28 | Good colonization effect, and can effectively reduce diarrhea and enteritis | [20] |
| TD1 | Does not produce Reutericyclin | [21] | ||
| Rats | Internal organs | DSMZ 17509 | Contains acid urease gene cluster, and can remove urea in rice wine | [22] |
| Chickens | Chicken | ATCC 55148 | Anaerobic; can be used to treat bacterial intestinal infection in poultry, and it delays and improves chronic enteritis | [23] |
| Intestines | KUB-AC5 | Effectively resists Salmonella and prevents poultry Salmonella infection, produces large amounts of exopolysaccharides, and has potential technical applications and uses for improving food properties | [24] | |
| SKKU-OGDONS-01 | Using antiviral protein as carrier as feed additive | [25] | ||
| BBC5 | Good tolerance, good adhesion to Caco-2 cells, and can significantly enhance chicken macrophages involved in immune regulation | [26] | ||
| Bees | Gut | LP4 | Good antibacterial properties, can regulate the expression of the antimicrobial peptide mRNA in the guts of adult oriental honey bees and worker bees, and increases the survival rate of bees | [27] |
| Uncertain | / | GMNL-263 | Improves renal fibrosis and insulin resistance, improve liver steatosis and hyperlipidemia, and enhances white adipose tissue browning | [28] |
| / | ATCCPTA 6475 | Reduces osteoclast activity, and improves bone health in estrogen-deficient conditions | [29] | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, J.; Li, K.; Xiao, Y.; Zhong, A.; Tang, J.; Duan, Y.; Li, Z. Limosilactobacillus reuteri Regulating Intestinal Function: A Review. Fermentation 2023, 9, 19. https://doi.org/10.3390/fermentation9010019
Jiang J, Li K, Xiao Y, Zhong A, Tang J, Duan Y, Li Z. Limosilactobacillus reuteri Regulating Intestinal Function: A Review. Fermentation. 2023; 9(1):19. https://doi.org/10.3390/fermentation9010019
Chicago/Turabian StyleJiang, Jiayan, Ke Li, Yu Xiao, Aiai Zhong, Jiaojiao Tang, Yufan Duan, and Zongjun Li. 2023. "Limosilactobacillus reuteri Regulating Intestinal Function: A Review" Fermentation 9, no. 1: 19. https://doi.org/10.3390/fermentation9010019
APA StyleJiang, J., Li, K., Xiao, Y., Zhong, A., Tang, J., Duan, Y., & Li, Z. (2023). Limosilactobacillus reuteri Regulating Intestinal Function: A Review. Fermentation, 9(1), 19. https://doi.org/10.3390/fermentation9010019