
Textural and Functional Properties of Skimmed and Whole Milk Fermented by Novel Lactiplantibacillus plantarum AG10 Strain Isolated from Silage

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Milk Fermentation
2.2. Preparing of Protein-Free Extract (PFE)
2.3. Quantitative Chemical Analysis of Fermented Skimmed and Whole Milk
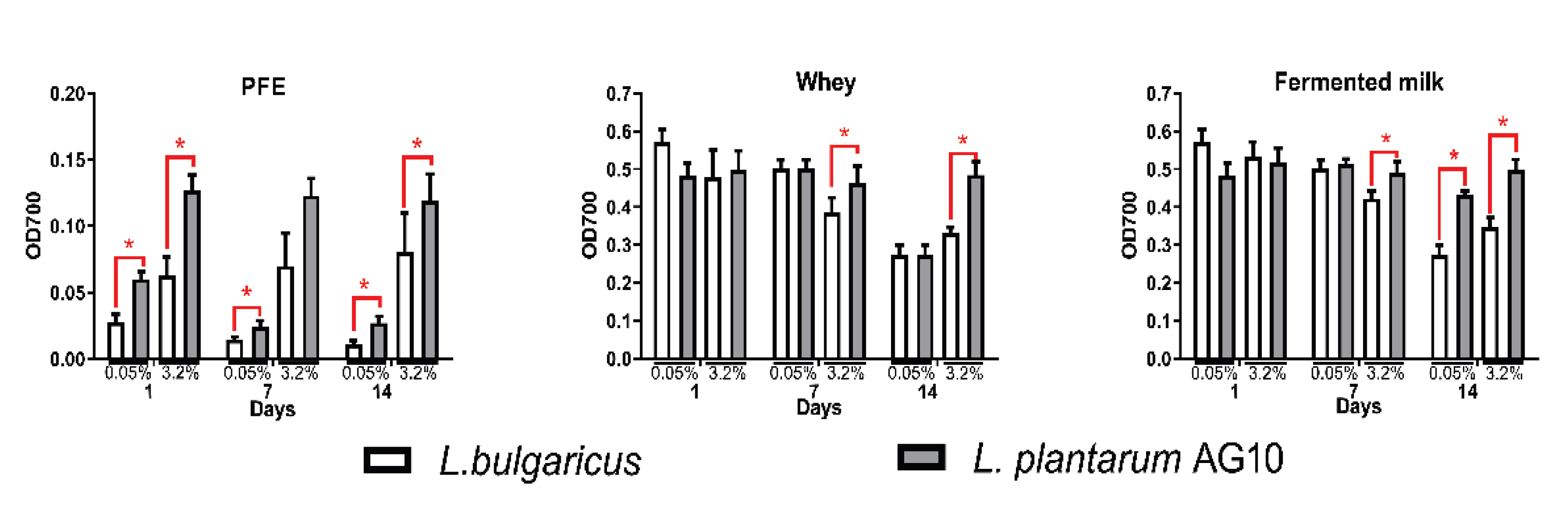
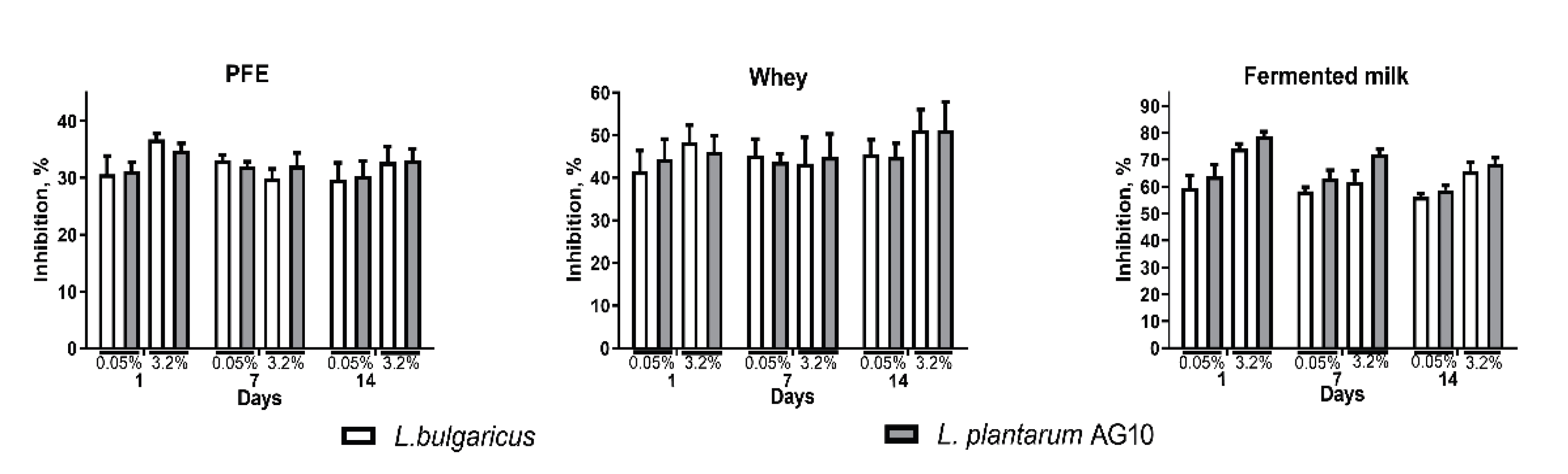
2.4. Antibacterial Activity
2.5. Determination of the Total Amount of Exopolysaccharides (EPS)
2.6. Apparent Viscosity
2.7. Textural Studies
2.8. Scanning Electron Microscopy
2.9. Ferric Reducing Antioxidant Power Assay (FRAP)
2.10. Evaluation of Radical-Scavenging Ability (RSA) by 2,2-Di-phenyl-1-picrylhydrazyl (DPPH) Assay
2.11. Evaluation of Peroxide Value of Fat
2.12. Evaluation of Thiobarbituric Acid-Reactive Substance (TBARS)
2.13. Evaluation of Free Fatty Acid Value
2.14. CFUs Count
2.15. Statistical Analysis
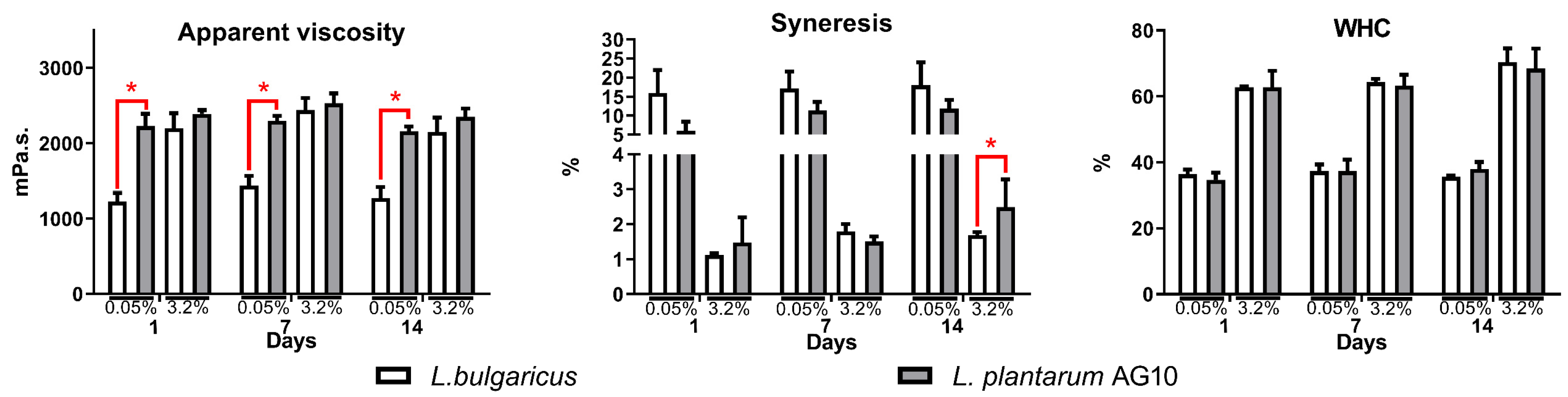
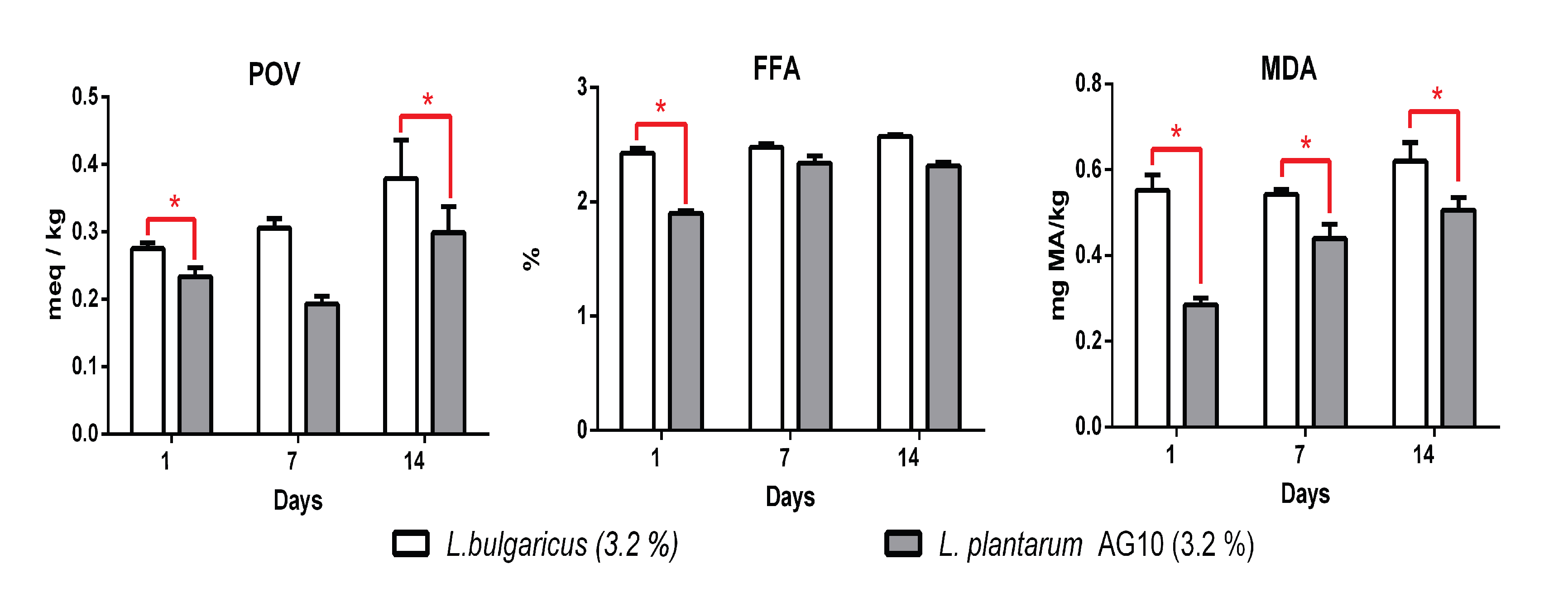
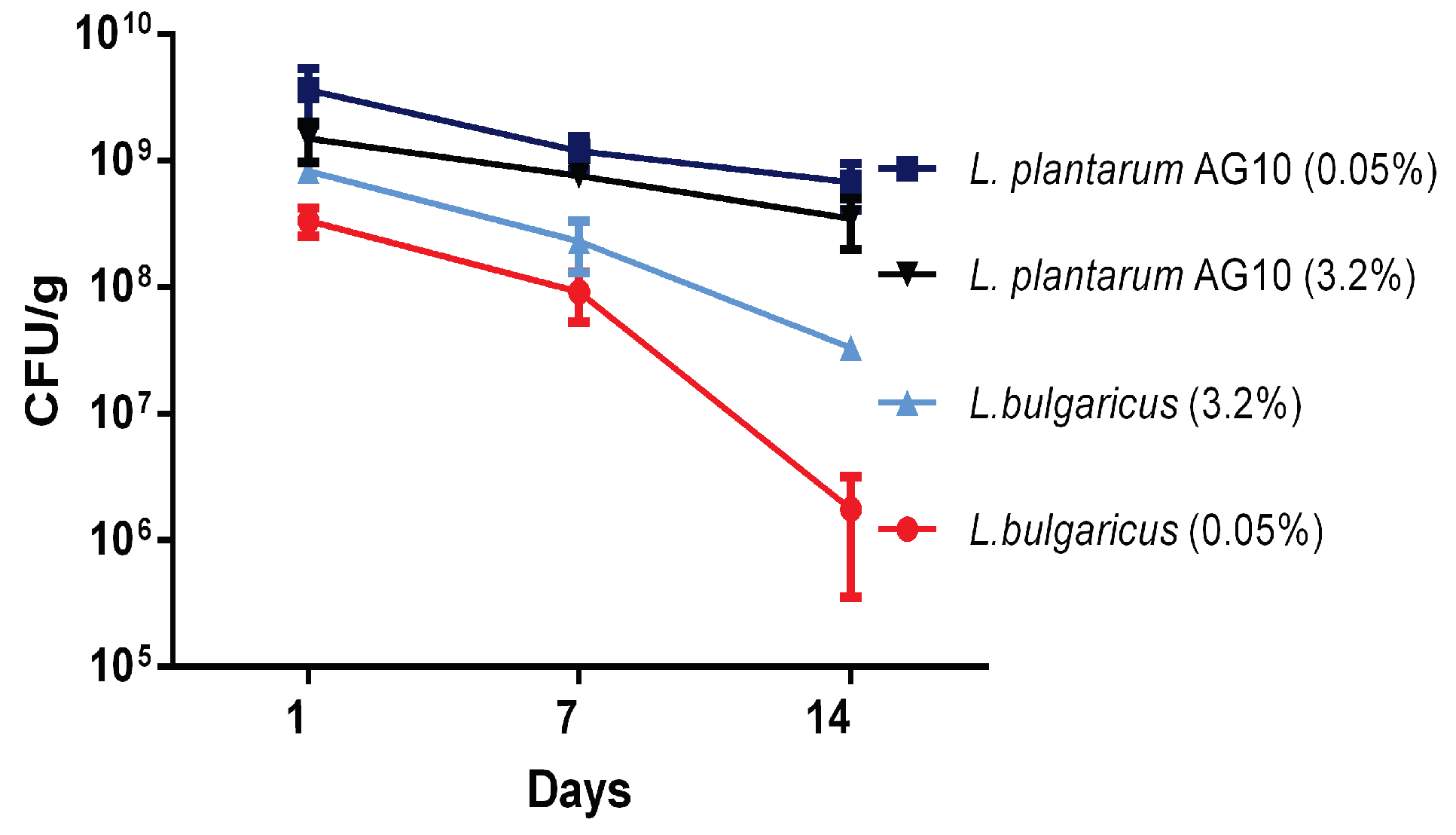
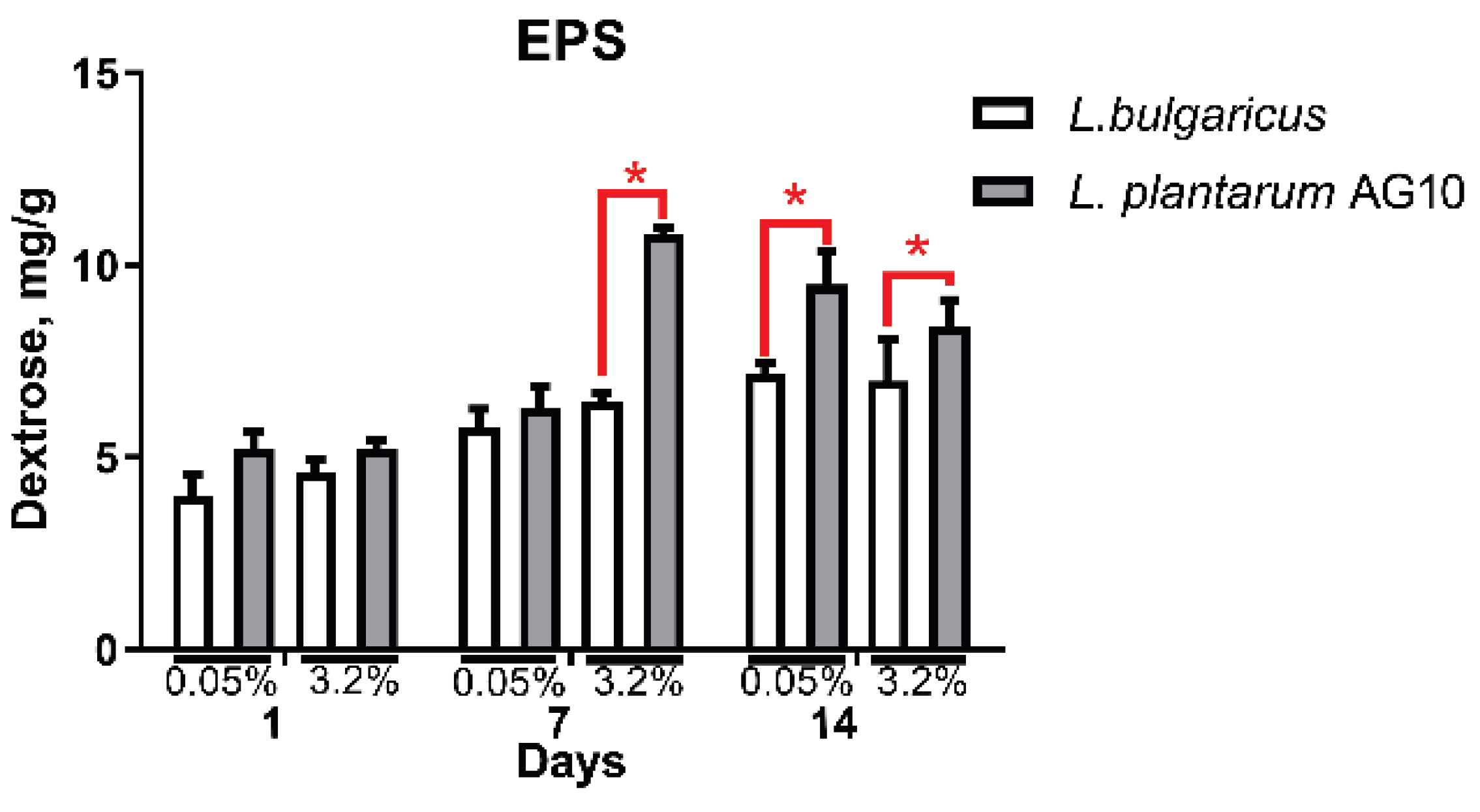
3. Results
3.1. Characteristics of Fermented Milk
3.2. The Exopolysaccharides Production
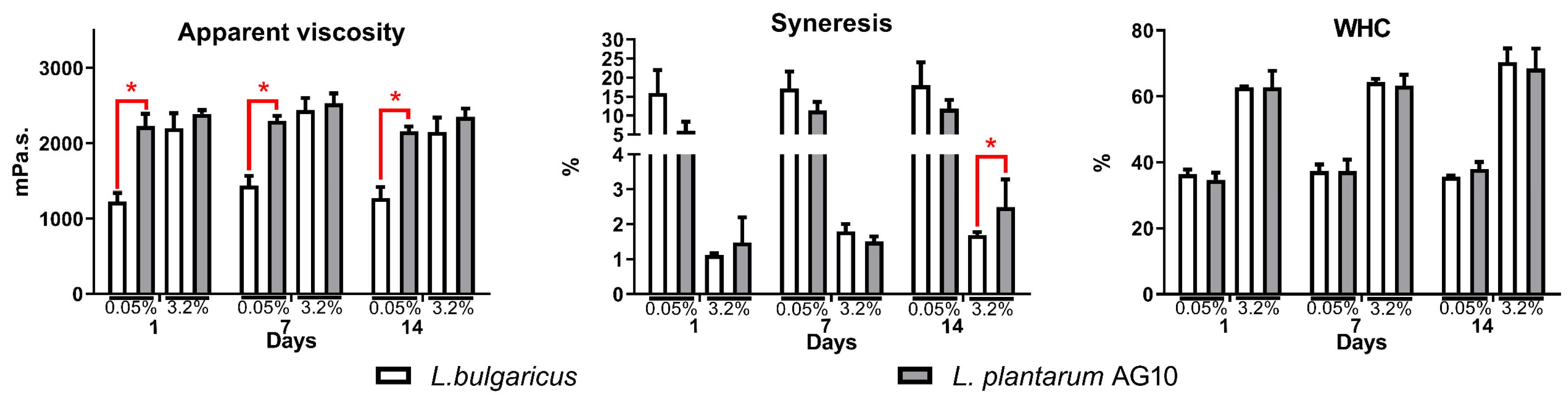
3.3. Textural Properties of Fermented Milk
3.4. Scanning Electron Microscopy
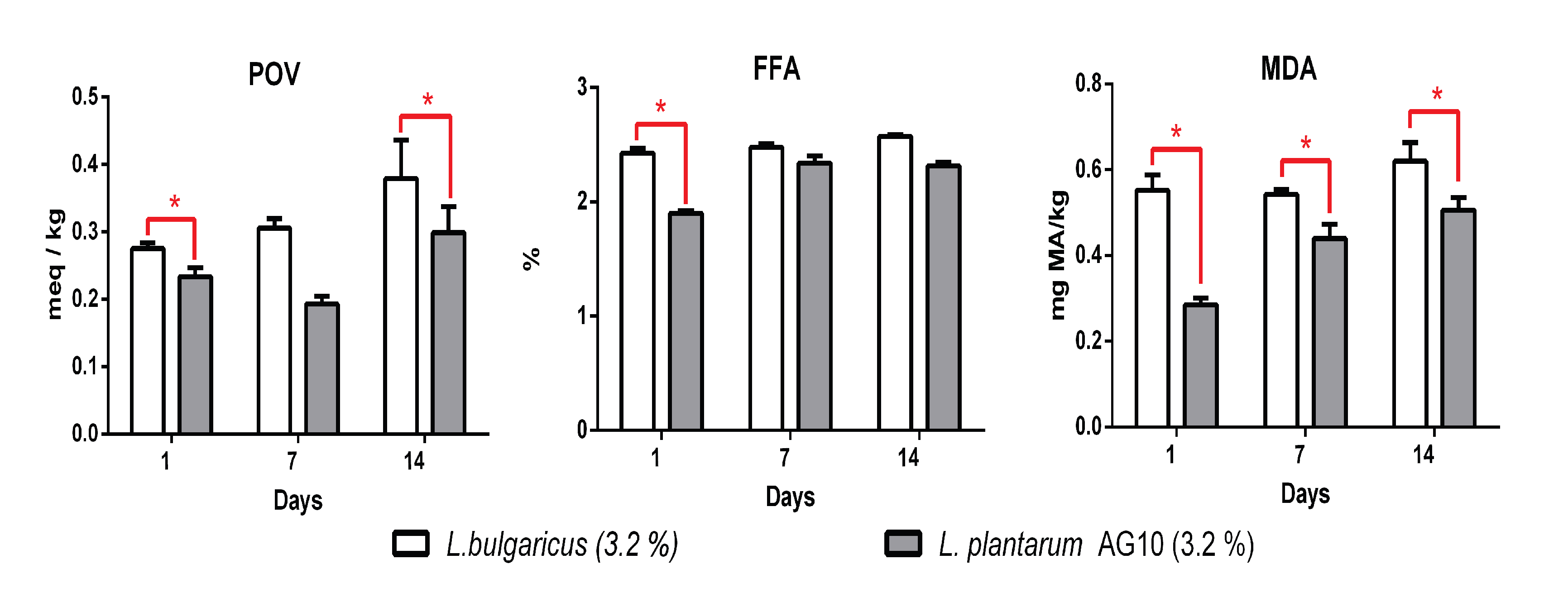
3.5. Antioxidant Properties
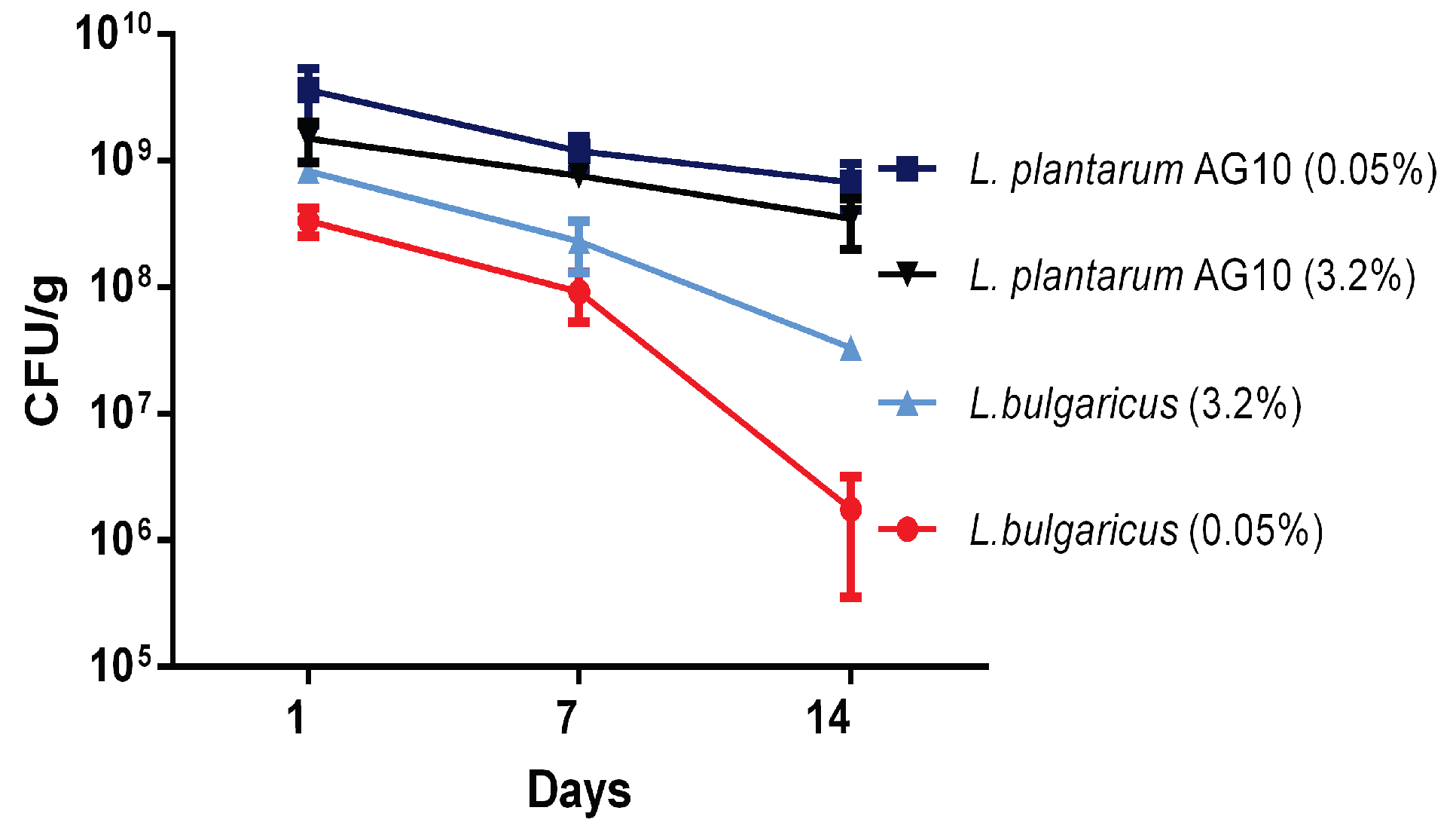
3.6. Viability of L. plantarum AG10 and L. bulgaricus during Storage
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| LAB | Lactic acid bacteria |
| NSLAB | Non-specialized lactic acid bacteria |
| TTA | Total titratable acidity |
| FRAP | Ferric reducing antioxidant power |
| RSA | Radical-scavenging ability |
| WHC | Water-holding capacity |
| DPPH | 2,2-di-phenyl-1-picrylhydrazyl |
| PFE | Protein-free extract |
| TBARS | Thiobarbituric acid reactive substance |
| TBA | 2-thiobarbituric acid |
| MA | Malonaldehyde |
| FFA | Free fatty acid |
| POV | Peroxide value |
References
- Bhat, Z.F.; Bhat, H. Milk and Dairy Products as Functional Foods: A Review. Int. J. Dairy Sci. 2011, 6, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Giraffa, G.; Chanishvili, N.; Widyastuti, Y. Importance of lactobacilli in food and feed biotechnology. Res. Microbiol. 2010, 161, 480–487. [Google Scholar] [CrossRef] [PubMed]
- Wen, R.; Sun, F.; Wang, Y.; Chen, Q.; Kong, B. Evaluation the potential of lactic acid bacteria isolates from traditional beef jerky as starter cultures and their effects on flavor formation during fermentation. LWT 2021, 142, 110982. [Google Scholar] [CrossRef]
- Weerathilake, W.; Rasika, D.; Ruwanmali, J.; Munasinghe, M. The evolution, processing, varieties and health benefits of yogurt. Int. J. Sci. Res. Publ. 2014, 4, 1–10. [Google Scholar]
- Bintsis, T. Lactic acid bacteria: Their applications in foods. J. Bacteriol. Mycol. Open Access 2018, 6, 89–94. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Salam, A.M.; Ammar, N.; Abdel-Hamid, A.Z. Effectiveness of Probiotic Labneh Supplemented with Garlic or Onion Oil Against Schistosoma mansoni in Infected Mice. Int. J. Dairy Sci. 2008, 3, 97–104. [Google Scholar] [CrossRef]
- Admassie, M. A review on food fermentation and the biotechnology of lactic acid bacteria. World J. Food Sci. Technol. 2018, 2, 19–24. [Google Scholar] [CrossRef] [Green Version]
- Zapaśnik, A.; Sokołowska, B.; Bryła, M.J.F. Role of Lactic Acid Bacteria in Food Preservation and Safety. Foods 2022, 11, 1283. [Google Scholar] [CrossRef]
- Schnürer, J.; Magnusson, J. Antifungal lactic acid bacteria as biopreservatives. Trends Food Sci. Technol. 2005, 16, 70–78. [Google Scholar] [CrossRef]
- Favaro, L.; Penna, A.L.B.; Todorov, S.D. Bacteriocinogenic LAB from cheeses–Application in biopreservation? Trends Food Sci. Technol. 2015, 41, 37–48. [Google Scholar] [CrossRef]
- Gemechu, T. Review on lactic acid bacteria function in milk fermentation and preservation. Afr. J. Food Sci. 2015, 9, 170–175. [Google Scholar] [CrossRef] [Green Version]
- Leyva Salas, M.; Mounier, J.; Valence, F.; Coton, M.; Thierry, A.; Coton, E. Antifungal microbial agents for food biopreservation—A review. Microorganisms 2017, 5, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shehata, M.G.; Badr, A.N.; El Sohaimy, S.A.; Asker, D.; Awad, T.S. Characterization of antifungal metabolites produced by novel lactic acid bacterium and their potential application as food biopreservatives. Ann. Agric. Sci. 2019, 64, 71–78. [Google Scholar] [CrossRef]
- Vieco-Saiz, N.; Belguesmia, Y.; Raspoet, R.; Auclair, E.; Gancel, F.; Kempf, I.; Drider, D. Benefits and inputs from lactic acid bacteria and their bacteriocins as alternatives to antibiotic growth promoters during food-animal production. Front. Microbiol. 2019, 10, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferencik, M.; Ebringer, L. Probiotics, allergy and asthma. Alergie 2003, 5, 224–230. [Google Scholar]
- Gill, H.S.; Guarner, F. Probiotics and human health: A clinical perspective. Postgrad. Med. J. 2004, 80, 516–526. [Google Scholar] [CrossRef] [Green Version]
- Gursoy, O.; Kinik, O. Probiotics: A New Popular Option for Cancer Inhibition. Int. J. Dairy Sci. 2006, 1, 100–103. [Google Scholar] [CrossRef] [Green Version]
- Lolou, V.; Mihalis, I.P. Functional Role of Probiotics and Prebiotics on Skin Health and Disease. Fermentation 2019, 5, 41. [Google Scholar] [CrossRef] [Green Version]
- Bajaj, B.K.; Claes, I.J.; Lebeer, S. Functional mechanisms of probiotics. J. Microbiol. Biotechnol. Food Sci. 2021, 2021, 321–327. [Google Scholar] [CrossRef] [Green Version]
- Bata, A.; Lasztity, R. Detoxification of mycotoxin-contaminated food and feed by microorganisms. Trends Food Sci. Technol. 1999, 10, 223–228. [Google Scholar] [CrossRef]
- Panesar, P.S. Fermented Dairy Products: Starter Cultures and Potential Nutritional Benefits. Food Nutr. Sci. 2011, 2, 47–51. [Google Scholar] [CrossRef] [Green Version]
- Jeong, J.-J.; Park, H.J.; Cha, M.G.; Park, E.; Won, S.-M.; Ganesan, R.; Gupta, H.; Gebru, Y.A.; Sharma, S.P.; Lee, S.B.J.M. The Lactobacillus as a Probiotic: Focusing on Liver Diseases. Microorganisms 2022, 10, 288. [Google Scholar] [CrossRef] [PubMed]
- Meisel, H.; Bockelmann, W. Bioactive peptides encrypted in milk proteins: Proteolytic activation and thropho-functional properties. Antonie Van Leeuwenhoek 1999, 76, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Hagi, T.; Kobayashi, M.; Nomura, M. Metabolome analysis of milk fermented by γ-aminobutyric acid–producing Lactococcus lactis. J. Dairy Sci. 2016, 99, 994–1001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohanty, D.; Mohapatra, S.; Misra, S.; Sahu, P. Milk derived bioactive peptides and their impact on human health–A review. Saudi J. Biol. Sci. 2016, 23, 577–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pessione, E.; Cirrincione, S.J.F.i.m. Bioactive molecules released in food by lactic acid bacteria: Encrypted peptides and biogenic amines. Front. Microbiol. 2016, 7, 876. [Google Scholar] [CrossRef] [Green Version]
- Kliche, T.; Li, B.; Bockelmann, W.; Habermann, D.; Klempt, M.; De Vrese, M.; Wutkowski, A.; Clawin-Raedecker, I.; Heller, K. Screening for proteolytically active lactic acid bacteria and bioactivity of peptide hydrolysates obtained with selected strains. Appl. Microbiol. Biotechnol. 2017, 101, 7621–7633. [Google Scholar] [CrossRef]
- Aguilar-Toalá, J.E.; Santiago-López, L.; Peres, C.M.; Peres, C.; Garcia, H.S.; Vallejo-Cordoba, B.; González-Córdova, A.F.; Hernández-Mendoza, A. Assessment of multifunctional activity of bioactive peptides derived from fermented milk by specific Lactobacillus plantarum strains. J. Dairy Sci. 2017, 100, 65–75. [Google Scholar] [CrossRef]
- Granier, A.; Goulet, O.; Hoarau, C. Fermentation products: Immunological effects on human and animal models. Pediatr. Res. 2013, 74, 238–244. [Google Scholar] [CrossRef] [Green Version]
- Sasikumar, K.; Vaikkath, D.K.; Devendra, L.; Nampoothiri, K.M. An exopolysaccharide (EPS) from a Lactobacillus plantarum BR2 with potential benefits for making functional foods. Bioresour. Technol. 2017, 241, 1152–1156. [Google Scholar] [CrossRef]
- Zhu, Y.; Wang, X.; Pan, W.; Shen, X.; He, Y.; Yin, H.; Zhou, K.; Zou, L.; Chen, S.; Liu, S. Exopolysaccharides produced by yogurt-texture improving Lactobacillus plantarum RS20D and the immunoregulatory activity. Int. J. Biol. Macromol. 2019, 121, 342–349. [Google Scholar] [CrossRef] [PubMed]
- Ayyash, M.; Abu-Jdayil, B.; Itsaranuwat, P.; Galiwango, E.; Tamiello-Rosa, C.; Abdullah, H.; Esposito, G.; Hunashal, Y.; Obaid, R.S.; Hamed, F. Characterization, bioactivities, and rheological properties of exopolysaccharide produced by novel probiotic Lactobacillus plantarum C70 isolated from camel milk. Int. J. Biol. Macromol. 2020, 144, 938–946. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.; Wang, Z.; Zhang, Y.; Zhang, J.; Wang, L.; Dong, X.; Su, F.; Yao, G.; Wang, S.; Zhang, H. Effect of Lactobacillus plantarum P-8 on lipid metabolism in hyperlipidemic rat model. Eur. J. Lipid Sci. Technol. 2012, 114, 1230–1236. [Google Scholar] [CrossRef]
- Song, M.; Park, S.; Lee, H.; Min, B.; Jung, S.; Park, S.; Kim, E.; Oh, S. Effect of Lactobacillus acidophilus NS1 on plasma cholesterol levels in diet-induced obese mice. J. Dairy Sci. 2015, 98, 1492–1501. [Google Scholar] [CrossRef] [Green Version]
- Bintsis, T. Lactic acid bacteria as starter cultures: An update in their metabolism and genetics. AIMS Microbiol. 2018, 4, 665–684. [Google Scholar] [CrossRef]
- Waters, D.M.; Mauch, A.; Coffey, A.; Arendt, E.K.; Zannini, E. Lactic acid bacteria as a cell factory for the delivery of functional biomolecules and ingredients in cereal-based beverages: A review. Crit. Rev. Food Sci. Nutr. 2015, 55, 503–520. [Google Scholar] [CrossRef]
- Liu, J.; Chan, S.H.J.; Chen, J.; Solem, C.; Jensen, P.R. Systems biology–A guide for understanding and developing improved strains of lactic acid bacteria. Front. Microbiol. 2019, 10, 876. [Google Scholar] [CrossRef]
- Morandi, S.; Silvetti, T.; Tamburini, A.; Brasca, M. Changes in oxidation-reduction potential during milk fermentation by wild lactic acid bacteria. J. Dairy Res. 2016, 83, 387–394. [Google Scholar] [CrossRef]
- Kim, S.; Lee, J.Y.; Jeong, Y.; Kang, C.-H.J.F. Antioxidant Activity and Probiotic Properties of Lactic Acid Bacteria. Fermentation 2022, 8, 29. [Google Scholar] [CrossRef]
- Yang, J.; Cao, Y.; Cai, Y.; Terada, F. Natural populations of lactic acid bacteria isolated from vegetable residues and silage fermentation. J. Dairy Sci. 2010, 93, 3136–3145. [Google Scholar] [CrossRef]
- Muelas, R.; Olives, A.M.d.; Romero, G.; Díaz, J.R.; Sayas-Barberá, M.E.; Sendra, E. Evaluation of individual lactic acid bacteria for the fermentation of goat milk: Quality parameters. LWT 2018, 98, 506–514. [Google Scholar] [CrossRef]
- Argyri, A.A.; Zoumpopoulou, G.; Karatzas, K.A.; Tsakalidou, E.; Nychas, G.J.; Panagou, E.Z.; Tassou, C.C. Selection of potential probiotic lactic acid bacteria from fermented olives by in vitro tests. Food Microbiol. 2013, 33, 282–291. [Google Scholar] [CrossRef] [PubMed]
- Gavrilova, E.; Anisimova, E.; Gabdelkhadieva, A.; Nikitina, E.; Vafina, A.; Yarullina, D.; Bogachev, M.; Kayumov, A. Newly isolated lactic acid bacteria from silage targeting biofilms of foodborne pathogens during milk fermentation. BMC Microbiol. 2019, 19, 248. [Google Scholar] [CrossRef] [PubMed]
- Zheng, G.; Slavik, M.F. Isolation, partial purification and characterization of a bacteriocin produced by a newly isolated Bacillus subtilis strain. Lett. Appl. Microbiol. 1999, 28, 363–367. [Google Scholar] [CrossRef]
- Klose, V.; Bayer, K.; Bruckbeck, R.; Schatzmayr, G.; Loibner, A.P. In vitro antagonistic activities of animal intestinal strains against swine-associated pathogens. Vet. Microbiol. 2010, 144, 515–521. [Google Scholar] [CrossRef]
- Feldmane, J.; Ciprovica, I.; Semjonovs, P.; Linde, R. The influence of fermentation temperature on the development of exopolysaccharide in yoghurt production. In Proceedings of the 9th Baltic Conference on Food Science and Technology “Food for Consumer Well-Being”, Jelgava, Latvia, 8–9 May 2014; 2014; p. 266. [Google Scholar]
- Iranmanesh, M.; Ezzatpanah, H.; Mojgani, N. Antibacterial activity and cholesterol assimilation of lactic acid bacteria isolated from traditional Iranian dairy products. LWT-Food Sci. Technol. 2014, 58, 355–359. [Google Scholar] [CrossRef]
- Najgebauer-Lejko, D.; Witek, M.; Żmudziński, D.; Ptaszek, A. Changes in the viscosity, textural properties, and water status in yogurt gel upon supplementation with green and Pu-erh teas. J. Dairy Sci. 2020, 103, 11039–11049. [Google Scholar] [CrossRef]
- Nikitina, E.V.; Yurtaeva, T.A.; Tsyganov, M.S.; Ezhkova, G.O. Physico-Chemical and Antioxidant Properties of Skimmed Varenets (Slavic Baked Milk Yogurt) Mixed with Enzyme-Modified Potato Starches. Curr. Res. Nutr. Food Sci. 2021, 9, 88–99. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT-Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Sallam, K.I.; Ishioroshi, M.; Samejima, K. Antioxidant and antimicrobial effects of garlic in chicken sausage. Lebensm Wiss Technol. 2004, 37, 849–855. [Google Scholar] [CrossRef] [Green Version]
- Schmedes, A.; Hølmer, G. A new thiobarbituric acid (TBA) method for determining free malondialdehyde (MDA) and hydroperoxides selectively as a measure of lipid peroxidation. J. Am. Oil Chem. Soc. 1989, 66, 813–817. [Google Scholar] [CrossRef]
- Rukunudin, I.; White, P.; Bern, C.; Bailey, T. A modified method for determining free fatty acids from small soybean sample sizes. J. Am. Oil Chem. Soc. 1998, 75, 563–568. [Google Scholar] [CrossRef]
- Herigstad, B.; Hamilton, M.; Heersink, J. How to optimize the drop plate method for enumerating bacteria. J. Microbiol. Methods 2001, 44, 121–129. [Google Scholar] [CrossRef]
- Sharafutdinov, I.S.; Trizna, E.Y.; Baidamshina, D.R.; Ryzhikova, M.N.; Sibgatullina, R.R.; Khabibrakhmanova, A.M.; Latypova, L.Z.; Kurbangalieva, A.R.; Rozhina, E.V.; Klinger-Strobel, M.; et al. Antimicrobial Effects of Sulfonyl Derivative of 2(5H)-Furanone against Planktonic and Biofilm Associated Methicillin-Resistant and -Susceptible Staphylococcus aureus. Front. Microbiol. 2017, 8, 2246. [Google Scholar] [CrossRef]
- Ma, C.; Cheng, G.; Liu, Z.; Gong, G.; Chen, Z. Determination of the essential nutrients required for milk fermentation by Lactobacillus plantarum. LWT-Food Sci. Technol. 2016, 65, 884–889. [Google Scholar] [CrossRef]
- Zha, M.; Li, K.; Zhang, W.; Sun, Z.; Kwok, L.-Y.; Menghe, B.; Chen, Y. Untargeted mass spectrometry-based metabolomics approach unveils molecular changes in milk fermented by Lactobacillus plantarum P9. LWT 2021, 140, 110759. [Google Scholar] [CrossRef]
- Zaunmüller, T.; Eichert, M.; Richter, H.; Unden, G. Variations in the energy metabolism of biotechnologically relevant heterofermentative lactic acid bacteria during growth on sugars and organic acids. Appl. Microbiol. Biotechnol. 2006, 72, 421–429. [Google Scholar] [CrossRef]
- Sandoval-Castilla, O.; Lobato-Calleros, C.; Aguirre-Mandujano, E.; Vernon-Carter, E.J. Microstructure and texture of yogurt as influenced by fat replacers. Int. Dairy J. 2004, 14, 151–159. [Google Scholar] [CrossRef]
- Li, C.; Song, J.; Kwok, L.-y.; Wang, J.; Dong, Y.; Yu, H.; Hou, Q.; Zhang, H.; Chen, Y. Influence of Lactobacillus plantarum on yogurt fermentation properties and subsequent changes during postfermentation storage. J. Dairy Sci. 2017, 100, 2512–2525. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Hui, Y.; Gao, T.; Shu, G.; Chen, H. Function and characterization of novel antioxidant peptides by fermentation with a wild Lactobacillus plantarum 60. LWT 2021, 135, 110162. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | L. bulgaricus | L. plantarum AG10 | |||||
|---|---|---|---|---|---|---|---|
| Days | 1 | 7 | 14 | 1 | 7 | 14 | |
| Skimmed Milk (fat = 0.05%) | |||||||
| pH | 4.5 ± 0.3 | 4.5 ± 0.1 | 4.4 ± 0.2 | 5 ± 0.1 * | 5.1 ± 0.2 * | 5.1 ± 0.1 * | |
| Total titratable acidity, T° | 122 ± 8 | 130 ± 6 | 136 ± 4 | 87 ± 4 * | 91 ± 2 * | 93 ± 9 * | |
| Total proteins, % | 3.8 ± 0.6 | 3.9 ± 0.4 | 3.7 ± 0.5 | 3.8 ± 0.1 | 3.8 ± 0.1 | 3.8 ± 0.1 | |
| Total carbohydrates, % | 4.6 ± 0.1 | 4.5 ± 0.1 | 4.5 ± 0.2 | 4.2 ± 0.1 * | 4.2 ± 0.1 * | 4.0 ± 0.3 * | |
| Dextrose, mmol/l of whey | 5.3 ± 0.1 | 5.4 ± 0.3 | 5.2 ± 0.6 | 5.2 ± 0.1 | 5.6 ± 0.3 | 4.5 ± 0.6 * | |
| Salts, % | 0.73 ± 0,03 | 0.73 ± 0.02 | 0.74 ± 0.03 | 0.63 ± 0.04 | 0.68 ± 0.01 | 0.64 ± 0.21 | |
| Density, kg/m3 | 1037.2 ± 0.6 | 1037.6 ± 1.4 | 1037.0 ± 1.3 | 1035.4 ± 1.5 | 1035.8 ± 0.2 | 1038.9 ± 1.1 | |
| Dry matter, % | 9.54 ± 0.50 | 9.59 ± 0.29 | 9.42 ± 0.35 | 8.95 ± 0.22 | 9.07 ± 0.01 | 8.94 ± 0.42 | |
| Antimicrobial activity * | E. coli | ++ | ++ | ++ | ++ | ++ | + |
| S. aureus | ++ | ++ | ++ | + | + | + | |
| Whole Milk (fat = 3.2 %) | |||||||
| pH | 4.2 ± 0.1 | 4.1 ± 0.1 | 4.1 ± 0.1 | 4.8 ± 0.1 * | 4.8 ± 0.1 * | 4.9 ± 0.1 * | |
| Total titratable acidity, T° | 143 ± 67 | 152 ± 5 | 155 ± 6 | 92 ± 7 * | 95 ± 6 * | 99 ± 3.1 * | |
| Total proteins, % | 3.7 ± 0.1 | 3.6 ± 0.0 | 3.5 ± 0.1 | 3.5 ± 0.1 | 3.5 ± 0.1 | 3.4 ± 0.1 | |
| Total carbohydrates, % | 4.0 ± 0.1 | 4.1 ± 0.1 | 4.1 ± 0.0 | 3.9 ± 0.1 | 3.9 ± 0.1 | 3.9 ± 0.0 | |
| Dextrose, mmol/l of whey | 2.3 ± 0.3 | 2.2 ± 0.4 | 2.1 ± 0.2 | 1.8 ± 0.1 * | 1.7 ± 0.2 * | 1.6 ± 0.1 * | |
| Fat, % | 3.2 ± 0.3 | 3 ± 0.3 | 2.6 ± 0.3 | 2.7 ± 0.1 * | 2.2 ± 0.2 * | 2 ± 0.1 * | |
| Salts, % | 0.68 ± 0.04 | 0.63 ± 0.04 | 0.63 ± 0.01 | 0.68 ± 0.02 | 0.65 ± 0.01 | 0.64 ± 0.01 | |
| Density, kg/m3 | 1031 ± 0.1 | 1032 ± 0.6 | 1032 ± 1.1 | 1031 ± 1.0 | 1031 ± 0.5 | 1031 ± 0.4 | |
| Dry matter, % | 11.69 ± 0.22 | 11.35 ± 0.13 | 11.05 ± 0.30 | 10.86 ± 0.60 | 10.04 ± 0.09 | 10.09 ± 0.12 | |
| Antimicrobial activity 1 | E. coli | ++ | ++ | ++ | ++ | ++ | + |
| S. aureus | ++ | ++ | ++ | ++ | ++ | + | |
| L. bulgaricus, (fat = 0.05%) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Storage | Firmness, g | Fracturability | Elasticity | Adhesion Force, g | Adhesiveness | Cohesiveness | Gumminess | Springiness, s | Chewiness |
| 1 | 32.6 ± 1.0 | 6.35 ± 0.02 | 0.50 ± 0.01 | 6.80 ± 0.04 | 3.13 ± 1.20 | 0.27 ± 0.00 | 8.8 ± 0.3 | 20.8 ± 1.1 | 164.2 ± 6.5 |
| 7 | 32.5 ± 1.1 | 5.86 ± 0.03 | 0.57 ± 0.01 | 6.30 ± 0.10 | 2.98 ± 0.91 | 0.25 ± 0.02 | 9.2 ± 1.1 | 19.8 ± 0.9 | 185.1 ± 4.5 |
| 14 | 32.5 ± 1.0 | 5.96 ± 0.03 | 0.57 ± 0.02 | 6.10 ± 0.21 | 4.32 ± 1.03 | 0.32 ± 0.01 | 10.5 ± 0.9 | 19.7 ± 0.2 | 208.7 ± 35.2 |
| L. plantarum AG10, (fat = 0.05%) | |||||||||
| 1 | 30.2 ± 1.1 | 4.46 ± 0.02 | 0.69 ± 0.03 | 5.80 ± 0.22 | 4.32 ± 1.31 | 0.30 ± 0.003 | 17.7 ± 0.1 * | 9 ± 0.3 * | 160.4 ± 6.3 |
| 7 | 31.7 ± 1.1 | 5.84 ± 0.02 | 0.75 ± 0.01 | 5.90 ± 0.1 | 4.46 ± 0.64 | 0.33 ± 0.01 * | 18.5 ± 0.5 * | 10.3 ± 0.6 * | 191.2 ± 2.5* |
| 14 | 32.3 ± 1.0 | 5.72 ± 0.05 | 0.62 ± 0.01 | 6.10 ± 0.02 | 4.87 ± 0.52 | 0.38 ± 0.03 * | 19.3 ± 1.2 * | 12.2 ± 1.2 * | 2365.8 ± 8.5* |
| L. bulgaricus, (fat = 3.2%) | |||||||||
| 1 | 39.2 ± 1.2 | 11.92 ± 0.01 | 0.22 ± 0.02 | 10.00 ± 0.01 | 3.70 ± 2.01 | 0.25 ± 0.01 | 20 ± 2.1 | 9.6 ± 0.6 | 194.1 ± 2.3 |
| 7 | 51 ± 1.4 | 16.29 ± 0.01 | 0.14 ± 0.02 | 13.80 ± 0.10 | 4.77 ± 0.51 | 0.20 ± 0.01 | 27.5 ± 1.2 | 10.2 ± 0.2 | 281.4 ± 6.5 |
| 14 | 54.7 ± 1.4 | 19.69 ± 0.03 | 0.12 ± 0.04 | 14.40 ± 0.05 | 4.82 ± 0.86 | 0.20 ± 0.01 | 27.8 ± 0.9 | 11.1 ± 0.9 | 309.9 ± 10.5 |
| L. plantarum AG10, (fat = 3.2%) | |||||||||
| 1 | 38.3 ± 1.0 | 10.28 ± 0.02 | 0.30 ± 0.03 | 8.70 ± 0.03 | 4.14 ± 1.26 | 0.28 ± 0.01 | 18.9 ± 1.7 | 10.8 ± 1.1 | 205.1 ± 6.8 * |
| 7 | 52.4 ± 1.0 | 17.18 ± 0.02 | 0.13 ± 0.01 | 13.30 ± 0.02 | 5.35 ± 0.68 | 0.23 ± 0.01 | 26.6 ± 0.9 | 12.2 ± 1.0 | 324.5 ± 2.5 * |
| 14 | 59 ± 1.1 | 22,07 ± 0.04 | 0.10 ± 0.01 | 15.80 ± 0.02 | 6.13 ± 1.11 | 0.22 ± 0.01 | 27.8 ± 1.1 | 13.2 ± 0.3 | 368.8 ± 10.4 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nikitina, E.; Petrova, T.; Vafina, A.; Ezhkova, A.; Nait Yahia, M.; Kayumov, A. Textural and Functional Properties of Skimmed and Whole Milk Fermented by Novel Lactiplantibacillus plantarum AG10 Strain Isolated from Silage. Fermentation 2022, 8, 290. https://doi.org/10.3390/fermentation8060290
Nikitina E, Petrova T, Vafina A, Ezhkova A, Nait Yahia M, Kayumov A. Textural and Functional Properties of Skimmed and Whole Milk Fermented by Novel Lactiplantibacillus plantarum AG10 Strain Isolated from Silage. Fermentation. 2022; 8(6):290. https://doi.org/10.3390/fermentation8060290
Chicago/Turabian StyleNikitina, Elena, Tatyana Petrova, Adel Vafina, Asya Ezhkova, Monyr Nait Yahia, and Airat Kayumov. 2022. "Textural and Functional Properties of Skimmed and Whole Milk Fermented by Novel Lactiplantibacillus plantarum AG10 Strain Isolated from Silage" Fermentation 8, no. 6: 290. https://doi.org/10.3390/fermentation8060290
APA StyleNikitina, E., Petrova, T., Vafina, A., Ezhkova, A., Nait Yahia, M., & Kayumov, A. (2022). Textural and Functional Properties of Skimmed and Whole Milk Fermented by Novel Lactiplantibacillus plantarum AG10 Strain Isolated from Silage. Fermentation, 8(6), 290. https://doi.org/10.3390/fermentation8060290