
Characterization of Green Manure Sunn Hemp Crop Silage Prepared with Additives: Aerobic Instability, Nitrogen Value, and In Vitro Rumen Methane Production

 ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. GMSH Preparation
2.2. Design, Silage Making, and Measurement of Ensiling Loss, and Aerobic Stability
2.3. Analytical Procedures
2.4. Statistical Analysis
3. Results
3.1. GMSH Crop Material
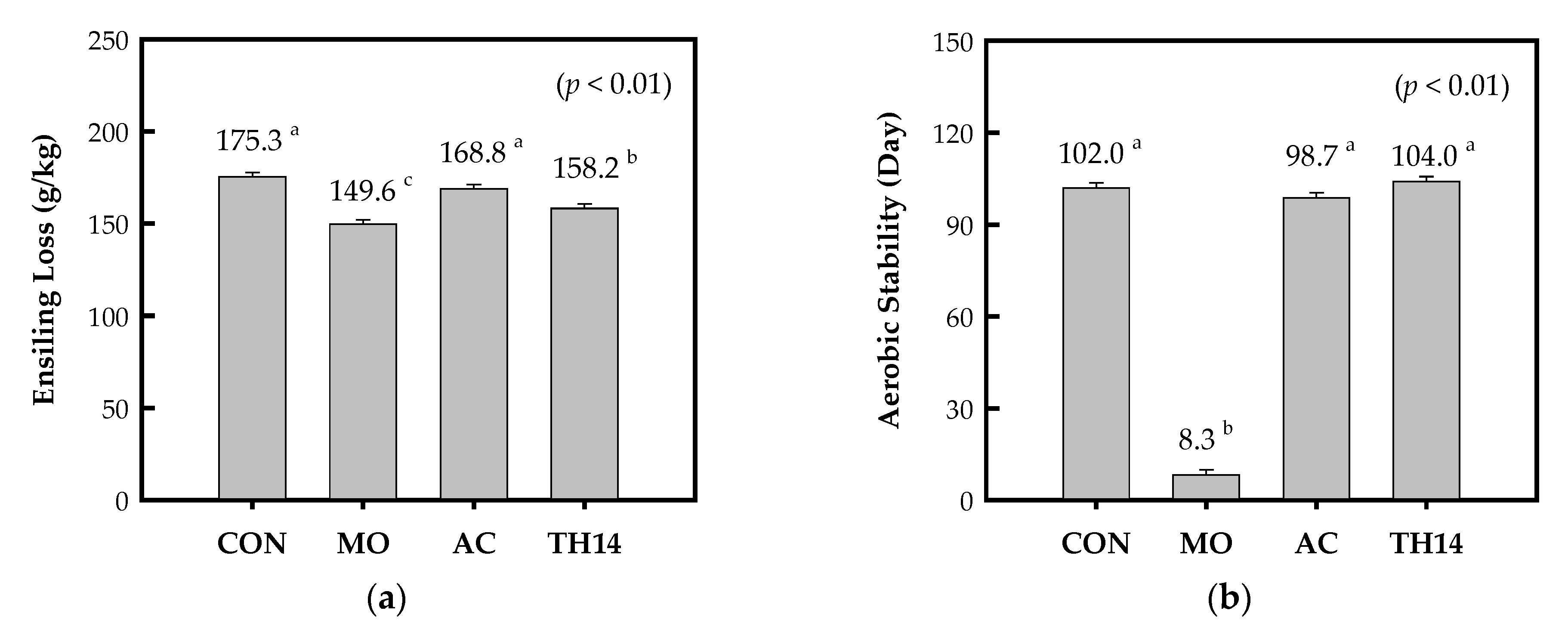
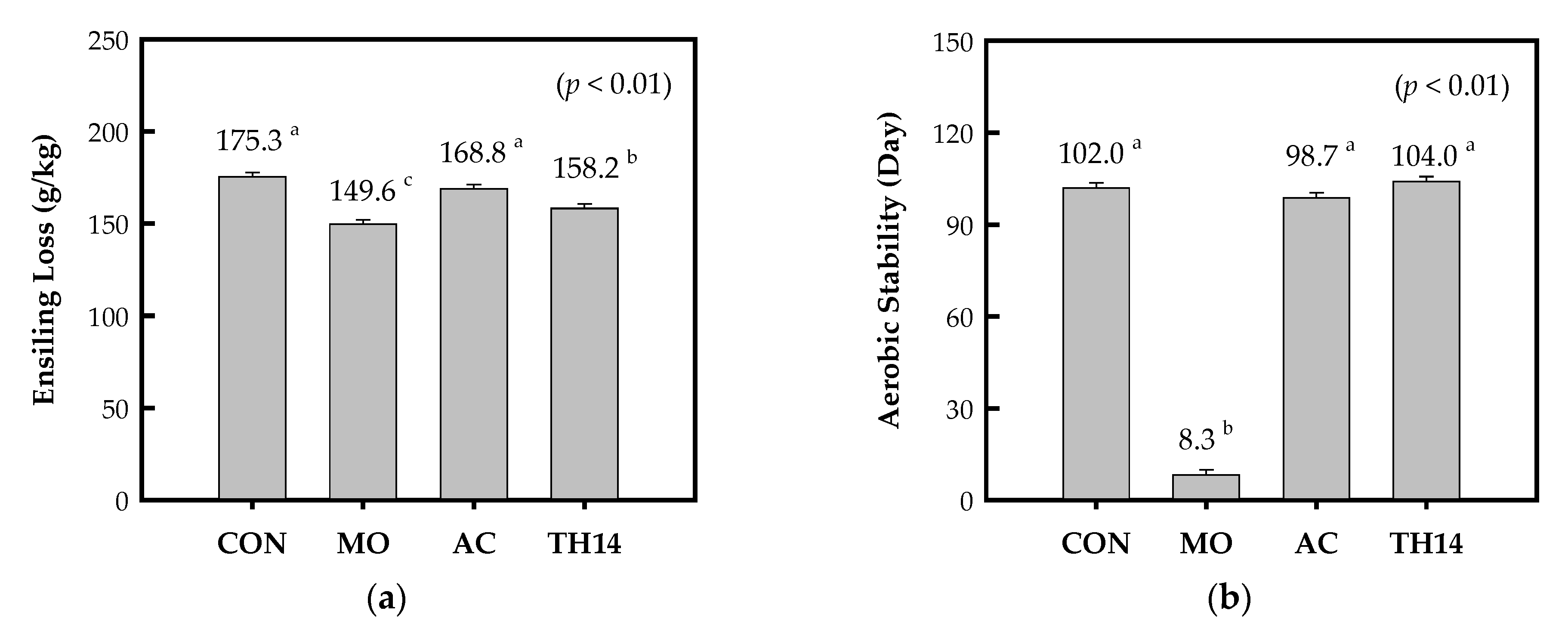
3.2. Ensiling Loss and Aerobic Stability
3.3. Silage Quality of GMSH Crop
3.4. Microbial Populations of GMSH Crop Silage
3.5. Chemical Compositions of GMSH Crop Silage
3.6. N Fractions of GMSH Crop Silage
3.7. In Vitro Digestibility, and Productions of Gas and Methane of GMSH Crop Silage
4. Discussion
4.1. Characteristics of GMSH Crop as Ensiling Material and Potent Forage
4.2. Characteristics of Ensiling Loss, Aerobic Stability, and Silage Quality in GMSH Crop
4.3. Characteristics of Microbial Populations in GMSH Crop Silage
4.4. Characteristics of Chemical Compositions in GMSH Crop Silage
4.5. Characteristics of N Fractions in GMSH Crop Silage
4.6. Characteristics of In Vitro Rumen Test in GMSH Crop Silage
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Garzon, J.; Vendramini, J.M.B.; Silveira, M.L.; Moriel, P.; da Silva, H.M.S.; Dubeux, J.C.B.; Kaneko, M.; Carnelos, C.C.; Mamede, P.A. Harvest management and genotype effects on Sunn hemp forage characteristics. Agron. J. 2021, 113, 298–307. [Google Scholar] [CrossRef]
- Ashworth, A.J.; West, C.P.; Allen, F.L.; Keyser, P.D.; Weiss, S.A.; Tyler, D.D.; Taylor, A.M.; Warwick, K.L.; Beamer, K.P. Biologically fixed nitrogen in legume intercropped systems: Comparison of nitrogen-difference and nitrogen-15 enrichment techniques. Agron. J. 2015, 107, 2419–2430. [Google Scholar] [CrossRef] [Green Version]
- Wanapat, M.; Totakul, P.; Viennasay, B.; Matra, M. Sunnhemp (Crotalaria juncea, L.) silage can enrich rumen fermentation process, microbial protein synthesis, and nitrogen utilization efficiency in beef cattle crossbreds. Trop. Anim. Health Prod. 2021, 53, 187. [Google Scholar] [CrossRef] [PubMed]
- McDonald, P.; Henderson, A.; Heron, S. The Biochemistry of Silage; Chalcombe Publications: Marlow, UK, 1991. [Google Scholar]
- Driehuis, F.; Wilkinson, J.M.; Jiang, Y.; Ogunade, I.; Adesogan, A.T. Silage review: Animal and human health risks from silage. J. Dairy Sci. 2018, 101, 4093–4110. [Google Scholar] [CrossRef]
- Sun, J.; Wang, T.; Huang, F.; Liu, Y.; Shi, W.; Ma, C.; Zhong, J. Silage fermentation: A potential microbial approach for the forage utilization of Cyperus esculentus L. by-product. Fermentation 2021, 7, 273. [Google Scholar] [CrossRef]
- Munekata, P.E.S.; Domínguez, R.; Pateiro, M.; Nawaz, A.; Hano, C.; Walayat, N.; Lorenzo, J.M. Strategies to increase the value of pomaces with fermentation. Fermentation 2021, 7, 299. [Google Scholar] [CrossRef]
- Khota, W.; Pholsen, S.; Higgs, D.; Cai, Y. Natural lactic acid bacteria population of tropical grasses and their fermentation factor analysis of silage prepared with cellulase and inoculant. J. Dairy Sci. 2016, 99, 9768–9781. [Google Scholar] [CrossRef] [Green Version]
- Ma, Z.-Y.; Ungerfeld, E.; Ouyang, Z.; Zhou, X.-L.; Han, X.-F.; Zeng, Y.-Q.; Tan, Z.-L. Effect of Lactobacillus plantarum inoculation on chemical composition, fermentation, and bacterial community composition of ensiled sweet corn whole plant or stover. Fermentation 2022, 8, 24. [Google Scholar] [CrossRef]
- Kaewpila, C.; Khota, W.; Gunun, P.; Kesorn, P.; Cherdthong, A. Strategic addition of different additives to improve silage fermentation, aerobic stability and in vitro digestibility of Napier grasses at late maturity stage. Agriculture 2020, 10, 262. [Google Scholar] [CrossRef]
- Wilson, R.F.; Wilkins, R.J. Formic acid as a silage additive for wet crops of cocksfoot and lucerne. J. Agric. Sci. 1973, 80, 225–231. [Google Scholar] [CrossRef]
- Panyawoot, N.; So, S.; Cherdthong, A.; Chanjula, P. Effect of feeding discarded durian peel ensiled with Lactobacillus casei TH14 and additives in total mixed rations on digestibility, ruminal fermentation, methane mitigation, and nitrogen balance of Thai Native-Anglo-Nubian goats. Fermentation 2022, 8, 43. [Google Scholar] [CrossRef]
- Kaewpila, C.; Gunun, P.; Kesorn, P.; Subepang, S.; Thip-Uten, S.; Cai, Y.; Pholsen, S.; Cherdthong, A.; Khota, W. Improving ensiling characteristics by adding lactic acid bacteria modifies in vitro digestibility and methane production of forage-sorghum mixture silage. Sci. Rep. 2021, 11, 1968. [Google Scholar] [CrossRef]
- Cao, Y.; Cai, Y.; Takahashi, T.; Yoshida, N.; Tohno, M.; Uegaki, R.; Nonaka, K.; Terada, F. Effect of lactic acid bacteria inoculant and beet pulp addition on fermentation characteristics and in vitro ruminal digestion of vegetable residue silage. J. Dairy Sci. 2011, 94, 3902–3912. [Google Scholar] [CrossRef] [PubMed]
- Ellis, J.L.; Bannink, A.; Hindrichsen, I.K.; Kinley, R.D.; Pellikaan, W.F.; Milora, N.; Dijkstra, J. The effect of lactic acid bacteria included as a probiotic or silage inoculant on in vitro rumen digestibility, total gas and methane production. Anim. Feed Sci. Technol. 2016, 211, 61–74. [Google Scholar] [CrossRef]
- Kaewpila, C.; Sommart, K. Development of methane conversion factor models for Zebu beef cattle fed low-quality crop residues and by-products in tropical regions. Ecol. Evol. 2016, 6, 7422–7432. [Google Scholar] [CrossRef]
- Kaewpila, C.; Sommart, K.; Mitsumori, M. Dietary fat sources affect feed intake, digestibility, rumen microbial populations, energy partition and methane emissions in different beef cattle genotypes. Animal 2018, 12, 2529–2538. [Google Scholar] [CrossRef] [PubMed]
- Khota, W.; Pholsen, S.; Higgs, D.; Cai, Y. Comparative analysis of silage fermentation and in vitro digestibility of tropical grass prepared with Acremonium and Tricoderma species producing cellulases. Asian-Australas. J. Anim. Sci. 2018, 31, 1913–1922. [Google Scholar] [CrossRef] [Green Version]
- Pholsen, S.; Khota, W.; Pang, H.; Higgs, D.; Cai, Y. Characterization and application of lactic acid bacteria for tropical silage preparation. Anim. Sci. J. 2016, 87, 1202–1211. [Google Scholar] [CrossRef]
- Weiß, K.; Kroschewski, B.; Auerbach, H.U. The influence of delayed sealing and repeated air ingress during the storage of maize silage on fermentation patterns, yeast development and aerobic stability. Fermentation 2022, 8, 48. [Google Scholar] [CrossRef]
- Moselhy, M.A.; Borba, J.P.; Borba, A.E.S. Improving the nutritive value, in vitro digestibility and aerobic stability of Hedychium gardnerianum silage through application of additives at ensiling time. Anim. Feed Sci. Technol. 2015, 206, 8–18. [Google Scholar] [CrossRef]
- Kozaki, M.; Uchimura, T.; Okada, S. Experimental Manual for Lactic Acid Bacteria; Asakurasyoten: Tokyo, Japan, 1992. [Google Scholar]
- AOAC. Official Methods of Analysis, 15th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 1990. [Google Scholar]
- Faichney, G.; White, G. Methods for the Analysis of Feeds Eaten by Ruminants; Division of Animal Production, Ian Clunies Ross Animal Research Laboratory, Commonwealth Scientific and Industrial Research Organization: Melbourne, Australia, 1983. [Google Scholar]
- Mertens, D.R. Creating a system for meeting the fiber requirements of dairy cows. J. Dairy Sci. 1997, 80, 1463–1481. [Google Scholar] [CrossRef]
- Krishnamoorthy, U.; Muscato, T.V.; Sniffen, C.J.; Van Soest, P.J. Nitrogen fractions in selected feedstuffs. J. Dairy Sci. 1982, 65, 217–225. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Cai, Y.; Benno, Y.; Ogawa, M.; Kumai, S. Effect of applying lactic acid bacteria isolated from forage crops on fermentation characteristics and aerobic deterioration of silage. J. Dairy Sci. 1999, 82, 520–526. [Google Scholar] [CrossRef]
- Darwin; Charles, W.; Cord-Ruwisch, R. Concurrent lactic and volatile fatty acid analysis of microbial fermentation samples by gas chromatography with heat pre-treatment. J. Chromatogr. Sci. 2018, 56, 1–5. [Google Scholar] [CrossRef]
- Fawcett, J.K.; Scott, J.E. A rapid and precise method for the determination of urea. J. Clin. Pathol. 1960, 13, 156–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makkar, H.P.; Blümmel, M.; Becker, K. Formation of complexes between polyvinyl pyrrolidones or polyethylene glycols and tannins, and their implication in gas production and true digestibility in in vitro techniques. Br. J. Nutr. 1995, 73, 897–913. [Google Scholar] [CrossRef] [Green Version]
- Muizelaar, W.; Bani, P.; Kuhla, B.; Larsen, M.; Tapio, I.; Yáñez-Ruiz, D. Rumen fluid sampling via oral stomach tubing method. In Methods in Cattle Physiology and Behaviour Research—Recommendations from the SmartCow Consortium; Mesgaran, S.D., Baumont, R., Munksgaard, L., Humphries, D., Kennedy, E., Dijkstra, J., Dewhurst, R., Ferguson, H., Terré, M., Kuhla, B., Eds.; PUBLISSO: Cologne, Germany, 2020; p. 6. [Google Scholar]
- Steel, R.G.D.; Torrie, J.H. Principles and Procedures of Statistics: A Biometrical Approach, 2nd ed.; McGraw–Hill Book Co. Inc.: New York, NY, USA, 1980. [Google Scholar]
- Allen, M.S. Effects of diet on short-term regulation of feed intake by lactating dairy cattle. J. Dairy Sci. 2000, 83, 1598–1624. [Google Scholar] [CrossRef]
- Kearl, L.C. Nutrient Requirements of Ruminants in Developing Countries; International Feedstuffs Institute: Logan, UT, USA, 1982. [Google Scholar]
- Lepcha, I.; Naumann, H.D. Partitioning of forage mass and nutritive value in Sunn hemp leaf and stem components. Int. J. Agron. 2021, 2021, 5547120. [Google Scholar] [CrossRef]
- Muck, R.E. Silage microbiology and its control through additives. Rev. Bras. Zootec. 2010, 39, 183–191. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Xu, W.; Yang, J.; Zhao, H.; Pan, C.; Ding, X. Effects of applying lactic acid bacteria to the fermentation on a mixture of corn steep liquor and air-dried rice straw. Anim. Nutr. 2016, 2, 229–233. [Google Scholar] [CrossRef]
- Xue, Z.; Liu, N.; Wang, Y.; Yang, H.; Wei, Y.; Moriel, P.; Palmer, E.; Zhang, Y. Combining orchardgrass and alfalfa: Effects of forage ratios on in vitro rumen degradation and fermentation characteristics of silage compared with hay. Animals 2020, 10, 59. [Google Scholar] [CrossRef] [Green Version]
- Heinritz, S.N.; Martens, S.D.; Avila, P.; Hoedtke, S. The effect of inoculant and sucrose addition on the silage quality of tropical forage legumes with varying ensilability. Anim. Feed Sci. Technol. 2012, 174, 201–210. [Google Scholar] [CrossRef]
- Kaewpila, C.; Thip-Uten, S.; Cherdthong, A.; Khota, W. Impact of cellulase and lactic acid bacteria inoculant to modify ensiling characteristics and in vitro digestibility of sweet corn stover and cassava pulp silage. Agriculture 2021, 11, 66. [Google Scholar] [CrossRef]
- Gao, J.L.; Wang, P.; Zhou, C.H.; Li, P.; Tang, H.Y.; Zhang, J.B.; Cai, Y. Chemical composition and in vitro digestibility of corn stover during field exposure and the fermentation characteristics of silage prepared with microbial additives. Asian-Australas. J. Anim. Sci. 2019, 32, 1854–1863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinne, M.; Winquist, E.; Pihlajaniemi, V.; Niemi, P.; Seppälä, A.; Siika-Aho, M. Fibrolytic enzyme treatment prior to ensiling increased press-juice and crude protein yield from grass silage. Bioresour. Technol. 2020, 299, 122572. [Google Scholar] [CrossRef]
- Van Soest, P.J. Nutritional Ecology of the Ruminant, 2nd ed.; Cornell University Press: Ithaca, NY, USA, 1994. [Google Scholar]
- Musco, N.; Koura, I.B.; Tudisco, R.; Awadjihè, G.; Adjolohoun, S.; Cutrignelli, M.I.; Mollica, M.P.; Houinato, M.; Infascelli, F.; Calabrò, S. Nutritional characteristics of forage grown in south of Benin. Asian-Australas. J. Anim. Sci. 2016, 29, 51–61. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
| Item * | GMSH Crop | |
|---|---|---|
| Microbial counts (cfu/g FM) | LAB | 6.4 × 103 |
| Coliform bacteria | 1.8 × 105 | |
| Aerobic bacteria | 2.7 × 109 | |
| Yeasts | 2.9 × 105 | |
| Molds | 1.3 × 109 | |
| pH value | 6.37 | |
| LBC (mEq/kg DM) | 2343 | |
| Chemical composition (g/kg DM) | DM (g/kg) | 256.1 |
| OM | 915.2 | |
| CP | 160.2 | |
| EE | 19.5 | |
| NDF | 623.8 | |
| ADF | 434.2 | |
| ADL | 94.7 | |
| NFC | 111.7 | |
| N fractions (g/kg total N) | Insoluble N | 564.9 |
| Soluble TPN | 178.5 | |
| Soluble NPN | 257.5 | |
| In vitro parameters | IVDMD (g/kg) | 557.6 |
| IVNDFD (g/kg) | 422.1 | |
| IVADFD (g/kg) | 293.2 | |
| Gas production (L/kg DM) | 135.9 | |
| Methane production (L/kg DM) | 17.24 | |
| Methane production (L/kg IVDMD) | 30.92 |
| Item * | pH | Lactic Acid | Acetic Acid | Propionic Acid | Butyric Acid | NH3N | |
|---|---|---|---|---|---|---|---|
| (g/kg DM) | (g/kg Total N) | ||||||
| AE | CON | 5.74 a | 7.22 b | 32.63 a | 8.57 a | 19.41 b | 112.7 b |
| MO | 4.33 d | 56.35 a | 17.38 b | 0.54 b | 0.78 c | 64.4 c | |
| AC | 4.76 c | 46.06 a | 26.80 a | 2.38 b | 5.84 c | 107.4 b | |
| TH14 | 5.63 b | 8.43 b | 26.55 a | 8.68 a | 26.96 a | 130.2 a | |
| SEM | 0.027 | 6.776 | 2.828 | 1.235 | 1.797 | 3.002 | |
| p | <0.01 | <0.01 | 0.018 | <0.01 | <0.01 | <0.01 | |
| AE + AIS | CON | 5.34 b | 4.39 b | 25.22 a | 5.71 a | 11.26 b | 5.14 b |
| MO | 4.75 c | 16.35 a | 10.14 b | 1.19 b | 1.94 c | 1.80 c | |
| AC | 8.76 a | 3.99 b | 3.27 c | 0.73 b | 1.69 c | 12.03 a | |
| TH14 | 5.40 b | 2.64 b | 21.11 a | 6.62 a | 16.92 a | 6.37 b | |
| SEM | 0.120 | 1.237 | 1.651 | 0.641 | 0.617 | 0.601 | |
| p | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | |
| Additive (A) | CON | 5.54 b | 5.80 c | 28.93 a | 7.14 a | 15.34 b | 58.94 b |
| MO | 4.54 c | 36.35 a | 13.76 b | 0.86 b | 1.36 c | 33.08 c | |
| AC | 6.76 a | 25.03 b | 15.03 b | 1.56 b | 3.76 c | 59.72 b | |
| TH14 | 5.52 b | 5.54 c | 23.83 a | 7.65 a | 21.94 a | 68.27 a | |
| Condition (C) | AE | 5.12 b | 29.52 a | 25.84 a | 5.04 a | 13.25 a | 103.67 a |
| AE + AIS | 6.07 a | 6.84 b | 14.93 b | 3.56 b | 7.95 b | 6.34 b | |
| SEM | 0.084 | 5.183 | 2.191 | 0.693 | 1.384 | 2.145 | |
| p | A | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
| C | <0.01 | <0.01 | <0.01 | 0.011 | <0.01 | <0.01 | |
| A × C | <0.01 | <0.01 | <0.01 | 0.123 | <0.01 | <0.01 | |
| Item * | LAB | Coliform Bacteria | Aerobic Bacteria | Yeasts | Molds | |
|---|---|---|---|---|---|---|
| (Log10 cfu/g FM) | ||||||
| AE | CON | 7.79 a | ND | 9.50 a | 6.76 a | 1.22 |
| MO | 5.49 c | ND | 7.67 b | 5.94 c | ND | |
| AC | 6.28 b | ND | 7.47 c | 6.55 b | ND | |
| TH14 | 7.42 a | ND | 7.40 c | 6.60 b | ND | |
| SEM | 0.147 | − | 0.030 | 0.044 | − | |
| p | <0.01 | − | <0.01 | <0.01 | − | |
| AE + AIS | CON | 4.40 b | ND | 5.54 b | 5.09 b | ND c |
| MO | 8.09 a | 1.51 | 8.76 a | 8.29 a | 1.22 b | |
| AC | 1.87 c | 1.49 | 8.83 a | ND c | 6.94 a | |
| TH14 | 4.92 b | ND | 5.65 b | 5.32 b | ND c | |
| SEM | 0.667 | 0.749 | 0.285 | 0.104 | 0.509 | |
| p | <0.01 | 0.309 | <0.01 | <0.01 | <0.01 | |
| Additive (A) | CON | 6.10 a | ND | 7.52 b | 5.92 b | ND c |
| MO | 6.79 a | 0.75 | 8.21 a | 7.11 a | 0.61 b | |
| AC | 4.07 b | 0.74 | 8.15 a | 3.28 c | 3.47 a | |
| TH14 | 6.17 a | ND | 6.52 c | 5.96 b | ND c | |
| Condition (C) | AE | 6.74 a | ND | 8.01 a | 6.46 a | ND b |
| AE + AIS | 4.82 b | 0.75 | 7.19 b | 4.67 b | 2.04 a | |
| SEM | 0.435 | 0.530 | 0.201 | 0.087 | 0.360 | |
| p | A | <0.01 | 0.309 | <0.01 | <0.01 | <0.01 |
| C | <0.01 | 0.069 | <0.01 | <0.01 | <0.01 | |
| A × C | <0.01 | 0.309 | <0.01 | <0.01 | <0.01 | |
| Item * | DM | OM | CP | EE | NDF | ADF | ADL | |
|---|---|---|---|---|---|---|---|---|
| (g/kg) | (g/kg DM) | |||||||
| AE | CON | 308.4 b | 931.8 | 154.0 b | 32.7 a | 574.8 a | 477.1 a | 104.9 a |
| MO | 347.0 a | 933.9 | 164.2 a | 31.7 a | 480.6 c | 411.1 c | 86.8 b | |
| AC | 307.5 b | 934.1 | 159.3 ab | 34.7 a | 517.6 b | 444.6 b | 87.6 b | |
| TH14 | 300.6 b | 935.0 | 144.1 c | 28.3 b | 579.4 a | 489.7 a | 92.8 b | |
| SEM | 2.574 | 1.349 | 2.472 | 1.070 | 4.958 | 6.951 | 2.313 | |
| p | <0.01 | 0.425 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | |
| AE + AIS | CON | 339.5 a | 930.0 a | 147.5 b | 48.5 a | 672.6 a | 551.1 a | 117.3 b |
| MO | 338.5 a | 931.5 a | 176.9 a | 48.8 a | 543.0 b | 438.4 b | 94.1 c | |
| AC | 284.6 b | 909.3 b | 143.2 b | 35.9 ab | 681.2 a | 554.3 a | 132.6 a | |
| TH14 | 347.1 a | 930.7 a | 146.2 b | 24.0 b | 658.7 a | 530.7 a | 115.0 b | |
| SEM | 3.435 | 5.427 | 3.643 | 4.513 | 10.072 | 8.523 | 3.896 | |
| p | <0.01 | 0.037 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | |
| Additive (A) | CON | 324.0 b | 930.9 a | 150.8 b | 40.60 a | 623.7 a | 514.1 a | 111.1 a |
| MO | 342.8 a | 932.7 a | 170.6 a | 40.25 a | 511.8 c | 424.7 b | 90.4 b | |
| AC | 296.1 c | 921.7 b | 151.3 b | 35.32 a | 599.4 b | 499.4 a | 110.1 a | |
| TH14 | 323.9 b | 932.9 a | 145.2 b | 26.13 b | 619.1 a | 510.2 a | 103.9 a | |
| Condition (C) | AE | 315.9 b | 933.7 a | 155.4 | 31.8 b | 538.1 b | 455.6 b | 93.0 b |
| AE + AIS | 327.4 a | 925.4 b | 153.5 | 39.3 a | 638.9 a | 518.6 a | 114.8 a | |
| SEM | 1.754 | 4.138 | 2.653 | 3.272 | 7.910 | 7.291 | 3.083 | |
| p | A | <0.01 | 0.059 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
| C | <0.01 | 0.015 | 0.327 | <0.01 | <0.01 | <0.01 | <0.01 | |
| A × C | <0.01 | 0.047 | <0.01 | 0.015 | <0.01 | <0.01 | <0.01 | |
| Item * | Insoluble N | Soluble TPN | Soluble NPN | |
|---|---|---|---|---|
| (g/kg Total N) | ||||
| AE | CON | 354.7 | 189.4 a | 456.0 |
| MO | 285.7 | 189.8 a | 524.5 | |
| AC | 357.1 | 162.9 b | 479.9 | |
| TH14 | 302.5 | 190.7 a | 506.8 | |
| SEM | 26.061 | 5.263 | 26.035 | |
| p | 0.176 | <0.01 | 0.308 | |
| AE + AIS | CON | 336.9 b | 157.1 b | 506.0 a |
| MO | 245.5 c | 157.4 b | 597.1 a | |
| AC | 405.0 a | 295.2 a | 299.9 b | |
| TH14 | 357.1 ab | 163.9 b | 479.0 a | |
| SEM | 16.291 | 28.238 | 42.106 | |
| p | <0.01 | 0.011 | <0.01 | |
| Additive (A) | CON | 345.8 b | 173.2 b | 481.0 b |
| MO | 265.6 c | 173.6 b | 560.8 a | |
| AC | 381.0 a | 229.1 a | 389.9 c | |
| TH14 | 329.8 b | 177.3 b | 492.9 b | |
| Condition (C) | AE | 325.0 | 183.2 | 491.8 |
| AE + AIS | 336.1 | 193.4 | 470.5 | |
| SEM | 28.527 | 21.484 | 46.572 | |
| p | A | 0.011 | 0.062 | 0.024 |
| C | 0.592 | 0.515 | 0.530 | |
| A × C | 0.297 | <0.01 | 0.072 | |
| Item * | IVDMD | IVNDFD | IVADFD | Gas Production | Methane Production | ||
|---|---|---|---|---|---|---|---|
| (g/kg) | (g/kg) | (g/kg) | (L/kg DM) | (L/kg DM) | (L/kg IVDMD) | ||
| AE | CON | 458.8 c | 229.6 c | 224.1 c | 106.3 c | 13.83 c | 30.18 b |
| MO | 613.7 a | 333.8 a | 318.6 a | 148.4 a | 18.67 a | 30.41 b | |
| AC | 482.4 bc | 202.1 c | 182.1 d | 124.7 b | 16.31 b | 33.96 a | |
| TH14 | 511.2 b | 282.3 b | 266.1 b | 126.7 b | 17.94 a | 35.30 a | |
| SEM | 12.396 | 12.368 | 12.120 | 2.618 | 0.299 | 0.883 | |
| p | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | |
| AE + AIS | CON | 460.7 b | 330.1 ab | 287.8 a | 120.0 ab | 14.97 b | 32.47 a |
| MO | 575.3 a | 352.5 a | 310.6 a | 137.4 a | 18.51 a | 32.23 a | |
| AC | 402.0 c | 268.1 c | 226.2 c | 74.3 c | 9.08 c | 22.46 c | |
| TH14 | 456.8 b | 305.6 b | 259.4 b | 108.1 b | 13.07 b | 28.61 b | |
| SEM | 12.372 | 8.112 | 8.171 | 6.056 | 0.746 | 1.126 | |
| p | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | |
| Additive (A) | CON | 459.8 bc | 279.9 b | 256.0 b | 113.2 b | 14.41 b | 31.34 a |
| MO | 594.5 a | 343.2 a | 314.6 a | 142.9 a | 18.60 a | 31.32 a | |
| AC | 442.2 c | 235.1 c | 204.2 c | 99.5 c | 12.72 c | 28.19 b | |
| TH14 | 484.0 b | 294.0 b | 262.7 b | 117.4 b | 15.53 b | 31.95 a | |
| Condition (C) | AE | 516.5 a | 262.0 b | 247.8 b | 126.5 a | 16.70 a | 32.46 a |
| AE + AIS | 473.7 b | 314.1 a | 271.0 a | 110.0 b | 13.93 b | 28.94 b | |
| SEM | 9.806 | 9.768 | 11.077 | 4.379 | 0.477 | 0.904 | |
| p | A | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
| C | <0.01 | <0.01 | 0.012 | <0.01 | <0.01 | <0.01 | |
| A × C | <0.01 | <0.01 | 0.014 | <0.01 | <0.01 | <0.01 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaewpila, C.; Khota, W.; Gunun, P.; Kesorn, P.; Kimprasit, T.; Sarnklong, C.; Cherdthong, A. Characterization of Green Manure Sunn Hemp Crop Silage Prepared with Additives: Aerobic Instability, Nitrogen Value, and In Vitro Rumen Methane Production. Fermentation 2022, 8, 104. https://doi.org/10.3390/fermentation8030104
Kaewpila C, Khota W, Gunun P, Kesorn P, Kimprasit T, Sarnklong C, Cherdthong A. Characterization of Green Manure Sunn Hemp Crop Silage Prepared with Additives: Aerobic Instability, Nitrogen Value, and In Vitro Rumen Methane Production. Fermentation. 2022; 8(3):104. https://doi.org/10.3390/fermentation8030104
Chicago/Turabian StyleKaewpila, Chatchai, Waroon Khota, Pongsatorn Gunun, Piyawit Kesorn, Thachawech Kimprasit, Chaweng Sarnklong, and Anusorn Cherdthong. 2022. "Characterization of Green Manure Sunn Hemp Crop Silage Prepared with Additives: Aerobic Instability, Nitrogen Value, and In Vitro Rumen Methane Production" Fermentation 8, no. 3: 104. https://doi.org/10.3390/fermentation8030104
APA StyleKaewpila, C., Khota, W., Gunun, P., Kesorn, P., Kimprasit, T., Sarnklong, C., & Cherdthong, A. (2022). Characterization of Green Manure Sunn Hemp Crop Silage Prepared with Additives: Aerobic Instability, Nitrogen Value, and In Vitro Rumen Methane Production. Fermentation, 8(3), 104. https://doi.org/10.3390/fermentation8030104