
Nipa Sap Can Be Both Carbon and Nutrient Source for Acetic Acid Production by Moorella thermoacetica (f. Clostridium thermoaceticum) and Reduced Minimal Media Supplements


Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Hydrolysis of Nipa Sap
2.3. Batch Fermentation of Nipa Sap Hydrolysate
2.4. Analyses
3. Results and Discussion
3.1. Chemical Composition and Hydrolysis of Nipa Sap
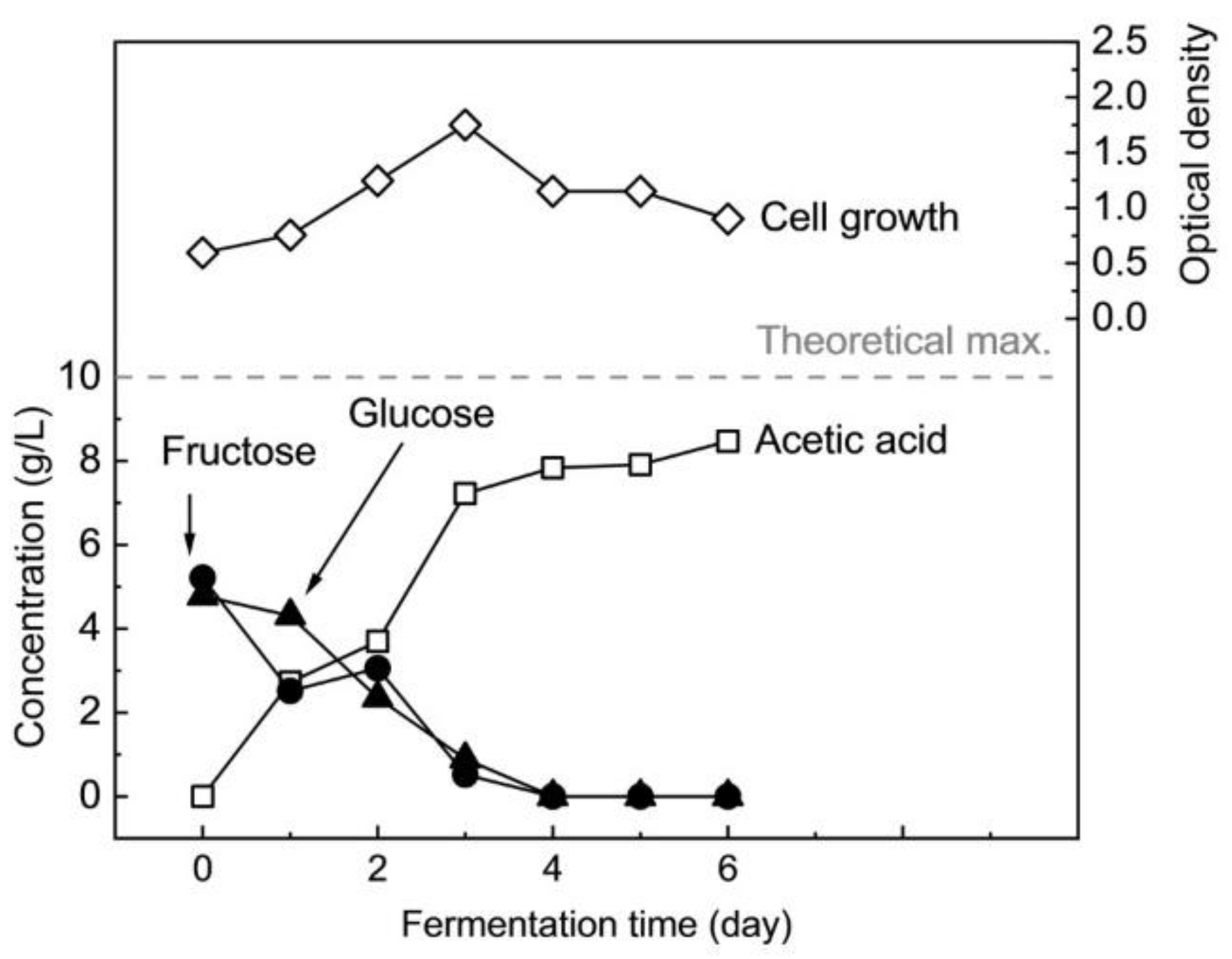
3.2. Batch Fermentation of Nipa Sap Hydrolysate
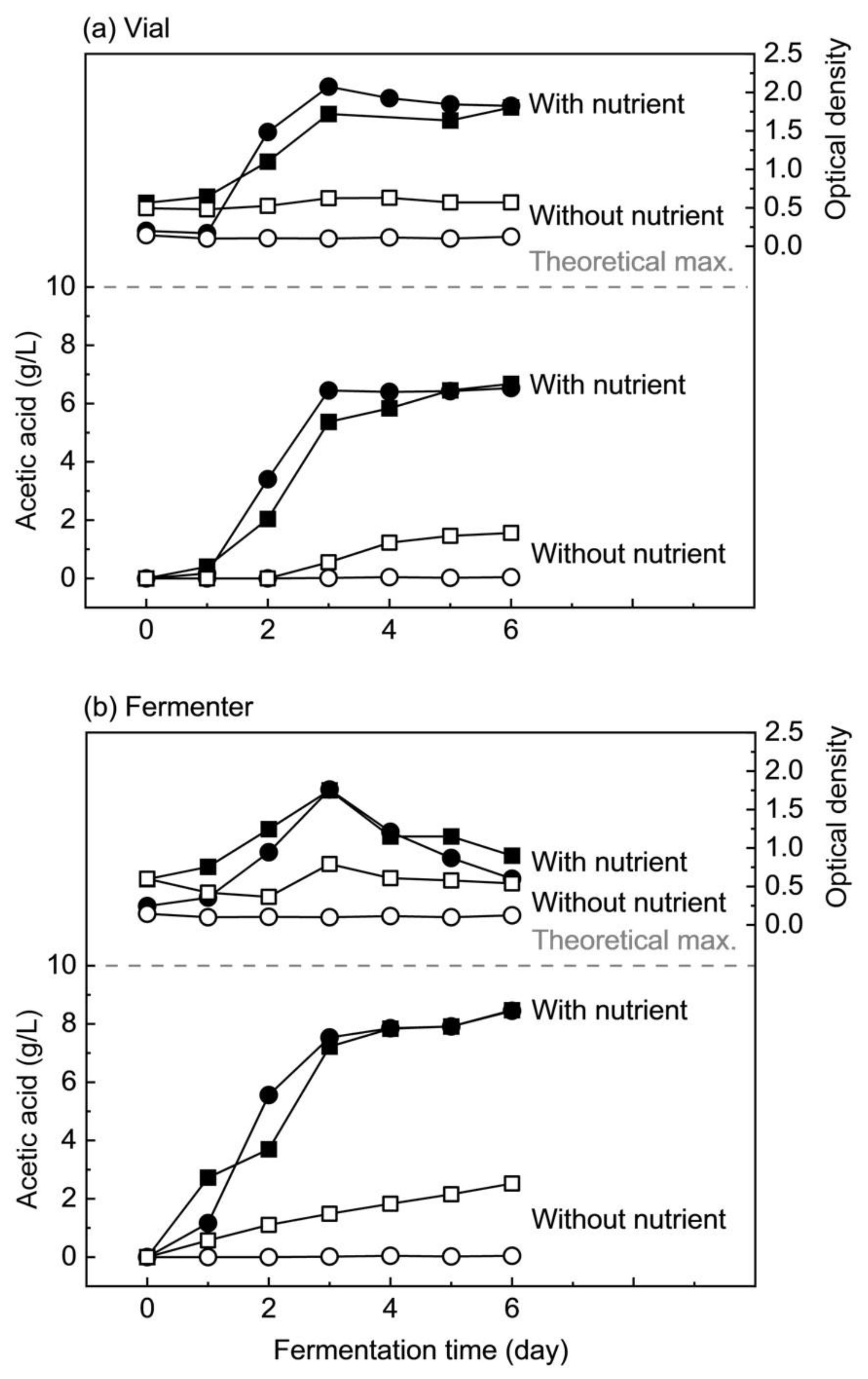
3.2.1. Fermentation of Hydrolyzed Nipa Sap with and without Nutrient Supplements in Comparison with Standard Sugars
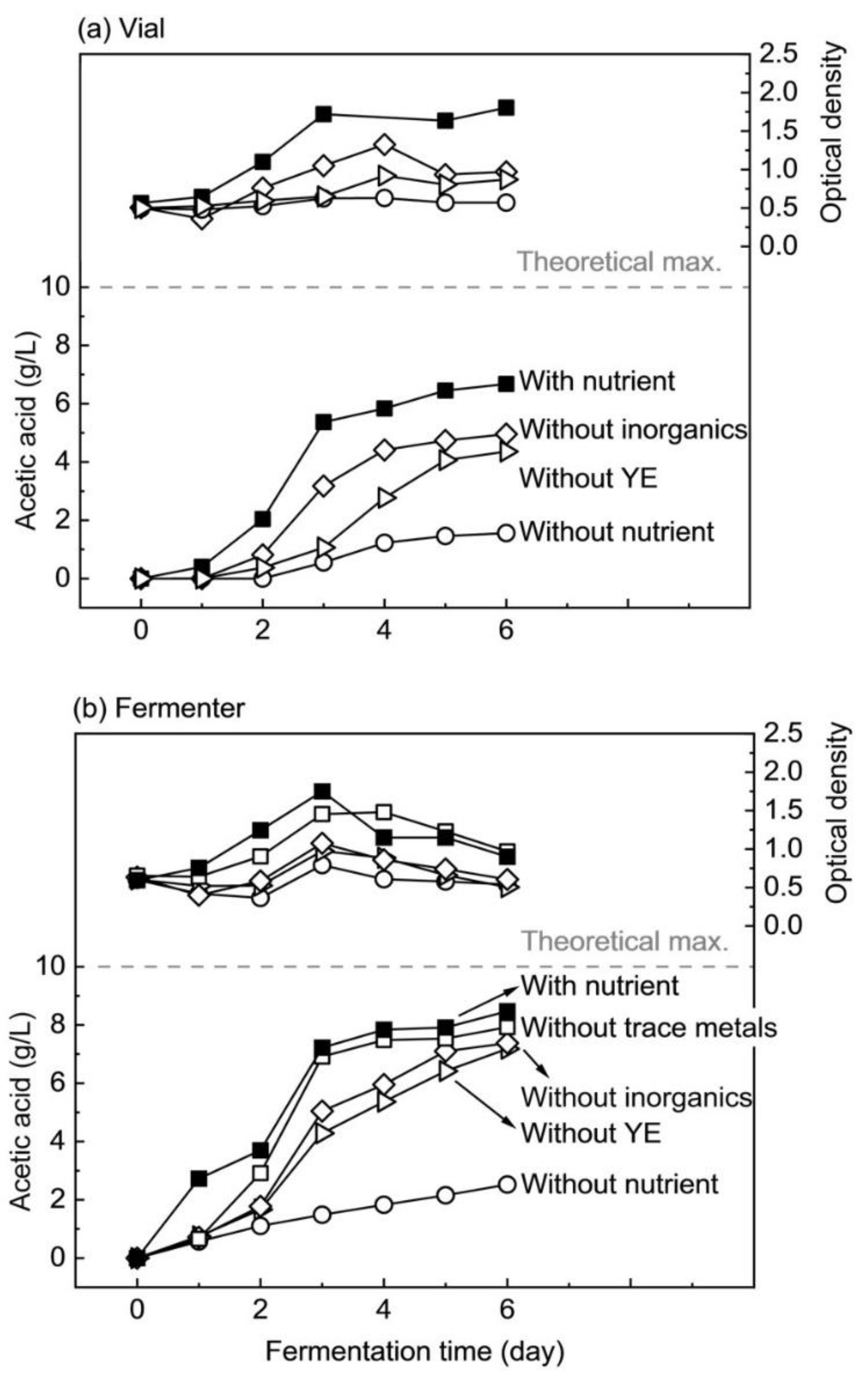
3.2.2. Fermentation of Hydrolyzed Nipa Sap without Trace Metals
3.2.3. Fermentation of Hydrolyzed Nipa Sap without Inorganics
3.2.4. Fermentation of Hydrolyzed Nipa Sap without Yeast Extract
3.2.5. Nipa Sap as Nutrient Source for Minimal Medium Supplementation
3.3. Comparison with Vinegar Process

4. Conclusions
- ➢
- Natural nipa sap contains crucial inorganic elements and vitamins in addition to its high sugar content.
- ➢
- Using only one feedstock, namely nipa sap, covered all inorganic needs of M. thermoacetica. Nipa sap was demonstrated to be a good source of nutrients for M. thermoacetica in small as well as in higher fermentation scales and can reduce the number of media supplements from 8–27 chemicals to as few as 2. The most economical nutrient condition was without yeast extract. It could reduce the price of a liter of the medium by 83%, and the price of supplementary nutrients by 80% per ton of acetic acid produced.
- ➢
- Sugar conversion efficiencies to acetic acid in fermenters without additional inorganics or yeast extract are more than threefold those obtained from the vinegar process and in a much shorter fermentation time. Thus, anaerobic fermentation using M. thermoacetica and nipa sap can be economically viable to afford efficient commercial acetic acid production from renewable resources.
- ➢
- The results also open opportunities for inexpensive co-digestion or co-fermentation of nipa sap with other biomass resources where nipa sap will serve both as a carbon and nutrient source.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ijmker, H.M.; Gramblička, M.; Kersten, S.R.A.; van der Ham, A.G.J.; Schuur, B. Acetic acid extraction from aqueous solutions using fatty acids. Sep. Purif. Technol. 2014, 125, 256–263. [Google Scholar] [CrossRef]
- Merli, G.; Becci, A.; Amato, A.; Beolchini, F. Acetic acid bioproduction: The technological innovation change. Sci. Total Environ. 2021, 798, 149292. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.V.; Sethapokin, P.; Rabemanolontsoa, H.; Minami, E.; Kawamoto, H.; Saka, S. Efficient production of acetic acid from nipa (Nypa fruticans) sap by Moorella thermoacetica (f. Clostridium thermoaceticum). Int. J. Green Technol. 2016, 2, 1–12. [Google Scholar]
- Rabemanolontsoa, H.; Kuninori, Y.; Saka, S. High conversion efficiency of Japanese cedar hydrolyzates into acetic acid by co-culture of Clostridium thermoaceticum and Clostridium thermocellum. J. Chem. Technol. Biotechnol. 2016, 91, 1040–1047. [Google Scholar] [CrossRef]
- Rabemanolontsoa, H.; Triwahyuni, E.; Takada, M. Consolidated bioprocessing of paper sludge to acetic acid by clostridial co-culture. Bioresour. Technol. Rep. 2021, 16, 100842. [Google Scholar] [CrossRef]
- Lim-Castillo, P. Traditional Philippine vinegars and their role in shaping the culinary culture. In Authenticity in the Kitchen: Proceedings of the Oxford Symposium on Food and Cookery; Hosking, R., Ed.; Oxford Symposium: Devon, UK, 2006; pp. 295–306. [Google Scholar]
- Shah, M.M.; Cheryan, M. Acetate production by Clostridium thermoaceticum in corn steep liquor media. J. Ind. Microbiol. 1995, 15, 424–428. [Google Scholar] [CrossRef]
- Lundie, L.L.; Drake, H.L. Development of a minimally defined medium for the acetogen Clostridium thermoaceticum. J. Bacteriol. 1984, 159, 700–703. [Google Scholar] [CrossRef]
- Witjitra, K.; Shah, M.M.; Cheryan, M. Effect of nutrient sources on growth and acetate production by Clostridium thermoaceticum. Enzym. Microb. Technol. 1996, 19, 322–327. [Google Scholar] [CrossRef]
- Peralta, A.S. Nipa based products and agro-processing. In Proceedings of the 14th Agriculture and Fisheries Technology Forum and Product Exhibition Seminar Series, SM Megamall, Mandaluyong City, Philippines, 31 August 2018. [Google Scholar]
- Hamilton, L.S.; Murphy, D.H. Use and management of nipa palm (Nypa fruticans, Arecaceae): A review. Econ. Bot. 1988, 42, 206–213. [Google Scholar] [CrossRef]
- Päivöke, A.E.A. Tapping practices and sap yields of the nipa palm (Nypa fruticans) in Papua New Guinea. Agric. Ecosyst. Environ. 1985, 13, 59–72. [Google Scholar] [CrossRef]
- Nakamura, Y.; Miyafuji, H.; Kawamoto, H.; Saka, S. Acetic acid fermentability with Clostridium thermoaceticum and Clostridium thermocellum of standard compounds found in beech wood as produced in hot-compressed water. J. Wood Sci. 2011, 57, 331–337. [Google Scholar] [CrossRef][Green Version]
- Sigma Aldrich. Prices of Nutrients for Microbial Growth Medium. Available online: https://www.sigmaaldrich.com/US/en (accessed on 11 October 2022).
- Van Die, J.; Tammes, P.M.L. Phloem exudation from monocotyledonous axes. In Transport in Plants I: Phloem Transport; Zimmermann, M.H., Milburn, J.A., Eds.; Springer: Berlin, Germany, 1975; pp. 196–222. [Google Scholar]
- Santiago-Urbina, J.; Ruíz-Terán, F. Microbiology and biochemistry of traditional palm wine produced around the world. Int. Food Res. J. 2014, 21, 1261–1269. [Google Scholar]
- Marquez, L.D.S.; Cabral, B.V.; Freitas, F.F.; Cardoso, V.L.; Ribeiro, E.J. Optimization of invertase immobilization by adsorption in ionic exchange resin for sucrose hydrolysis. J. Mol. Catal. B Enzym. 2008, 51, 86–92. [Google Scholar] [CrossRef]
- Kulshrestha, S.; Tyagi, P.; Sindhi, V.; Yadavilli, K.S. Invertase and its applications—A brief review. J. Pharm. Res. 2013, 7, 792–797. [Google Scholar] [CrossRef]
- Tamunaidu, P.; Saka, S. Comparative study of nutrient supplements and natural inorganic components in ethanolic fermentation of nipa sap. J. Jpn. Inst. Energy 2013, 92, 181–186. [Google Scholar] [CrossRef]
- Saengkrajang, W.; Chaijan, M.; Panpipat, W. Physicochemical properties and nutritional compositions of nipa palm (Nypa fruticans Wurmb) syrup. NFS J. 2021, 23, 58–65. [Google Scholar] [CrossRef]
- Phetrit, R.; Chaijan, M.; Sorapukdee, S.; Panpipat, W. Characterization of nipa palm’s (Nypa fruticans Wurmb.) sap and syrup as functional food ingredients. Sugar Tech. 2020, 22, 191–201. [Google Scholar] [CrossRef]
- Thorasin, T. A comparative study of the physicochemical, nutritional characteristics and microbiological contamination of fresh nipa palm (Nypa fruticans) sap. BUSCIJ 2018, 23, 1301–1316. [Google Scholar]
- Nguyen, V.D.; Rabemanolontsoa, H.; Saka, S. Sap from various palms as a renewable energy source for bioethanol production. Chem. Ind. Chem. Eng. 2016, 22, 355–373. [Google Scholar] [CrossRef]
- Mokhtar, S.I.; Zakaria, F.; Ayob, M.A.; Siow, W.S.; Nurshahida, A.S.; Latiff, Z.A.A. Study on the nutritional values and customer acceptance of Lansium domesticum & Nephelium lappaceum newly fermented natural fruit vinegars in Malaysia. Asian Pac. J. Adv. Bus. Soc. Stud. 2016, 2, 402–413. [Google Scholar]
- Reardon, K.F.; Bailey, J.E. Effects of pH and added metabolites on bioconversions by immobilized non-growing Clostridium acetobutylicum. Biotechnol. Bioeng. 1989, 34, 825–837. [Google Scholar] [CrossRef] [PubMed]
- Seifritz, C.; Daniel, S.L.; Gössner, A.; Drake, H.L. Nitrate as a preferred electron sink for the acetogen Clostridium thermoaceticum. J. Bacteriol. 1993, 175, 8008–8013. [Google Scholar] [CrossRef] [PubMed]
- Ehsanipour, M.; Suko, A.V.; Bura, R. Fermentation of lignocellulosic sugars to acetic acid by Moorella thermoacetica. J. Ind. Microbiol. Biotechnol. 2016, 43, 807–816. [Google Scholar] [CrossRef] [PubMed]
- Koesnandar; Ago, S.; Nishio, N.; Nagai, S. Production of extracellular 5-aminolevulinic acid by Clostridium thermoaceticum grown in minimal medium. Biotechnol. Lett. 1989, 11, 567–572. [Google Scholar] [CrossRef]
- Chongkhong, S.; Puangpee, S. Alternative energy under the royal initiative of his majesty the king: Ethanol from nipa sap using isolated yeast. Songklanakarin J. Sci. Technol. 2018, 40, 11. [Google Scholar]
- Saithong, P.; Nitipan, S.; Permpool, J. Optimization of vinegar production from nipa (Nypa fruticans Wurmb.) sap using surface culture fermentation process. Appl. Food Biotechnol. 2019, 6, 193–200. [Google Scholar]
- Paivoke, A.; Adams, M.R.; Twiddy, D.R. Nipa palm vinegar in Papua New Guinea. Process Biochem. 1984, 19, 84–87. [Google Scholar]
- Cheryan, M. Acetic acid production. In Encyclopedia of Microbiology; Schaechter, M., Ed.; Academic Press: Oxford, UK, 2009; pp. 144–149. [Google Scholar]
- Fontaine, F.E.; Peterson, W.H.; McCoy, E.; Johnson, M.J.; Ritter, G.J. A new type of glucose fermentation by Clostridium thermoaceticum. J. Bacteriol. 1942, 43, 701–715. [Google Scholar] [CrossRef]
- Koesnandar, A.; Nishio, N.; Nagai, S. Effects of trace metal ions on the growth, homoacetogenesis and corrinoid production by Clostridium thermoaceticum. J. Ferment. Bioeng. 1991, 71, 181–185. [Google Scholar] [CrossRef]
- Coelho, E.; Genisheva, Z.; Oliveira, J.M.; Teixeira, J.A.; Domingues, L. Vinegar production from fruit concentrates: Effect on volatile composition and antioxidant activity. J. Food Sci. Technol. 2017, 54, 4112–4122. [Google Scholar] [CrossRef]
- Coelho, E.; Vilanova, M.; Genisheva, Z.; Oliveira, J.M.; Teixeira, J.A.; Domingues, L. Systematic approach for the development of fruit wines from industrially processed fruit concentrates, including optimization of fermentation parameters, chemical characterization and sensory evaluation. LWT Food Sci. Technol. 2015, 62, 1043–1052. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Component | Concentration (g/L) | Price * ($/kg) | ||||
|---|---|---|---|---|---|---|
| With full Nutrients | Without Trace Metals | Without Inorganics | Without Yeast Extract | Without Nutrient | ||
| Inorganics | ||||||
| Trace metals | ||||||
| MgSO4·7H2O | 0.25 | - | - | 0.25 | - | 148 |
| Fe(NH4)2(SO4)2·6H2O | 0.04 | - | - | 0.04 | - | 388 |
| NiCl2·6H2O | 0.24 × 10−3 | - | - | 0.24 × 10−3 | - | 292 |
| ZnSO4·7H2O | 0.29 × 10−3 | - | - | 0.29 × 10−3 | - | 218 |
| Na2SeO3 | 0.017 × 10−3 | - | - | 0.017 × 10−3 | - | 1830 |
| Ammonium sulfate | 1.00 | 1.00 | - | 1.00 | - | 139 |
| Vitamin and organic nitrogen source | ||||||
| Yeast extract | 5.00 | 5.00 | 5.00 | - | - | 281 |
| Reducing agent | ||||||
| Cysteine | 0.10 | 0.10 | 0.10 | 0.10 | 0.10 | 906 |
| Price per liter of broth ($/L) | 1.7 | 1.6 | 1.5 | 0.3 | - | - |
| Chemical Composition | Concentration (g/L) | |
|---|---|---|
| Before Hydrolysis | After Hydrolysis | |
| Organic compounds | ||
| 82.7 ± 0.9 | 0.9 ± 0.0 |
| 29.9 ± 0.1 | 71.4 ± 1.1 |
| 38.5 ± 0.2 | 83.1 ± 2.6 |
| 1.6 ± 0.1 | 1.6 ± 0.1 |
| Total organics | 152.7 ± 1.3 | 157.0 ± 3.8 |
| Inorganic elements | ||
| 3.0 ± 0.3 | 3.0 ± 0.3 |
| 5.0 ± 2.8 | 5.0 ± 2.8 |
| 1.0 ± 0.3 | 1.0 ± 0.3 |
| (2.6 ± 3.5) × 10−2 | (2.6 ± 3.5) × 10−2 |
| (5.7 ± 0.3) × 10−2 | (5.7 ± 0.3) × 10−2 |
| (4.4 ± 0.9) × 10−2 | (4.4 ± 0.9) × 10−2 |
| (3.6 ± 3.8) × 10−3 | (3.6 ± 3.8) × 10−3 |
| (2.9 ± 2.2) × 10−4 | (2.9 ± 2.2) × 10−4 |
| (2.4 ± 2.2) × 10−4 | (2.4 ± 2.2) × 10−4 |
| Total inorganics | 9.1 ± 3.5 | 9.1 ± 3.5 |
| Total chemical composition | 161.8 ± 4.8 | 162.7 ± 7.3 |
| Inorganics Elements | Required Concentration for M. thermoacetica (g/L) | Concentration in Nipa Sap (g/L) | Reference |
|---|---|---|---|
| Trace elements | |||
| Magnesium (Mg) | 2.5 × 10−2 | (4.4 ± 0.9) × 10−2 | This work |
| Sulfur (S) | 0.28 | (5.7 ± 0.3) × 10−2 | This work |
| Iron (Fe) | 5.7 × 10−3 | 5.9 × 10−3 | Saengkrajang et al. [20] |
| Nickel (Ni) | 5.9 × 10−5 | Unknown | - |
| Chlorine (Cl) | Less than 0.24 × 10−3 | 5.0 ± 2.8 | This work |
| Zinc (Zn) | 0.066 × 10−3 | 0.5 × 10−3 | Saengkrajang et al. [20] |
| Sodium (Na) | Less than 1.0 × 10−3 | 1.0 ± 0.3 | This work |
| Selenium (Se) | 7.8 × 10−6 | Unknown | - |
| Nitrogen source | |||
| Nitrogen (N) | 0.2 from ammonium sulfate 0.5 from yeast extract | 0.6 ± 0.1 | Tamunaidu and Saka [19] |
| Experiment | Substrate Concentration (g/L) | Substrate Consumption (%) | Acetic Acid | Max. Cell Density at 660 nm | |||||
|---|---|---|---|---|---|---|---|---|---|
| Glc* | Frc* | Glc* | Frc* | Overall | Concentration (g/L) | Conversion Efficiency (%) | Production (g/L/h) | ||
| Nipa sap | |||||||||
| With nutrient | |||||||||
| Vial | 4.69 | 5.31 | 63 | 100 | 80 | 6.7 | 67 | 0.046 | 1.72 |
| Fermenter | 4.78 | 5.22 | 100 | 100 | 100 | 8.5 | 85 | 0.059 | 1.75 |
| Without trace metals | |||||||||
| Vial | - | - | - | - | - | - | - | - | - |
| Fermenter | 4.75 | 5.25 | 98 | 100 | 99 | 7.9 | 79 | 0.055 | 1.48 |
| Without inorganics | |||||||||
| Vial | 4.64 | 5.36 | 37 | 100 | 71 | 5.0 | 50 | 0.034 | 1.33 |
| Fermenter | 4.77 | 5.23 | 98 | 91 | 94 | 7.4 | 74 | 0.051 | 1.08 |
| Without yeast extract | |||||||||
| Vial | 4.68 | 5.32 | 29 | 100 | 67 | 4.4 | 44 | 0.030 | 0.92 |
| Fermenter | 4.77 | 5.23 | 98 | 92 | 95 | 7.2 | 72 | 0.050 | 0.98 |
| Without nutrient | |||||||||
| Vial | 4.69 | 5.31 | 10 | 31 | 21 | 1.6 | 16 | 0.011 | 0.63 |
| Fermenter | 4.77 | 5.23 | 67 | 52 | 59 | 2.5 | 25 | 0.018 | 0.79 |
| Standard sugars | |||||||||
| With nutrient | |||||||||
| Vial | 5.12 | 4.88 | 74 | 85 | 80 | 6.4 | 64 | 0.044 | 2.08 |
| Fermenter | 5.11 | 4.89 | 100 | 100 | 100 | 8.4 | 84 | 0.059 | 1.76 |
| Without nutrient | |||||||||
| Vial | 5.10 | 4.90 | 20 | 18 | 19 | 0.0 | 4 | 0.000 | 0.13 |
| Fermenter | 5.10 | 4.90 | 1 | 2 | 1 | 0.0 | 0 | 0.000 | 0.19 |
| References | Carbon Source | Nitrogen Source | Vitamin | Reducing Agent | Number of Other Mineral Salts | Chelator | Total Number of Supplements* | Conversion Efficiency (%) | |
|---|---|---|---|---|---|---|---|---|---|
| Lundie et al. [8] | Glucose | Ammonium sulfate | Nicotinic acid | Cysteine | 19 | Potassium sulfate | 23 | Not reported | |
| Seifritz et al. [26] | Vanillin, glucose | Yeast extract and various amino-acids | Nicotinic acid | - | 15 | - | 27 | Not reported | |
| Ehsanipour et al. [27] | Xylose, glucose | Yeast extract | Cysteine | 11 | - | 13 | 71 | ||
| This work | With full nutrient | Nipa sap | Yeast extract, ammonium sulfate | Cysteine | 5 | - | 8 | 85 | |
| Without trace metals | Nipa sap | Yeast extract, ammonium sulfate | Cysteine | - | - | 3 | 79 | ||
| Without inorganics | Nipa sap | Yeast extract | Cysteine | - | - | 2 | 74 | ||
| Without yeast extract | Nipa sap | Ammonium sulfate | - | Cysteine | 5 | - | 7 | 72 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen, D.V.; Rabemanolontsoa, H. Nipa Sap Can Be Both Carbon and Nutrient Source for Acetic Acid Production by Moorella thermoacetica (f. Clostridium thermoaceticum) and Reduced Minimal Media Supplements. Fermentation 2022, 8, 663. https://doi.org/10.3390/fermentation8110663
Nguyen DV, Rabemanolontsoa H. Nipa Sap Can Be Both Carbon and Nutrient Source for Acetic Acid Production by Moorella thermoacetica (f. Clostridium thermoaceticum) and Reduced Minimal Media Supplements. Fermentation. 2022; 8(11):663. https://doi.org/10.3390/fermentation8110663
Chicago/Turabian StyleNguyen, Dung Van, and Harifara Rabemanolontsoa. 2022. "Nipa Sap Can Be Both Carbon and Nutrient Source for Acetic Acid Production by Moorella thermoacetica (f. Clostridium thermoaceticum) and Reduced Minimal Media Supplements" Fermentation 8, no. 11: 663. https://doi.org/10.3390/fermentation8110663
APA StyleNguyen, D. V., & Rabemanolontsoa, H. (2022). Nipa Sap Can Be Both Carbon and Nutrient Source for Acetic Acid Production by Moorella thermoacetica (f. Clostridium thermoaceticum) and Reduced Minimal Media Supplements. Fermentation, 8(11), 663. https://doi.org/10.3390/fermentation8110663