
Effect of Metabolic Regulators and Aeration on Isocitric Acid Synthesis by Yarrowia lipolytica Grown on Ester-Aldehyde Fraction


Abstract
:1. Introduction
2. Materials and Methods
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Barth, G.; Gaillardin, C. Yarrowia lipolytica. In Nonconventional Yeasts in Biotechnology: A Handbook; Wolf, K., Ed.; Springer: Berlin/Heidelberg, Germany, 1996; pp. 313–388. [Google Scholar] [CrossRef]
- Barth, G.; Gaillardin, C. Physiology and genetics of the dimorphic fungus Yarrowia lipolytica. FEMS Microbiol. Rev. 1997, 19, 219–237. [Google Scholar] [CrossRef]
- Zinjarde, S.S. Food-related applications of Yarrowia lipolytica. Food Chem. 2014, 152, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Groenewald, M.; Boekhout, T.; Neuvéglise, C.; Gaillardin, C.; van Dijck, P.V.; Wyss, M. Yarrowia lipolytica: Safety assessment of an oleaginous yeast with a great industrial potential. Crit. Rev. Microbiol. 2014, 40, 187–206. [Google Scholar] [CrossRef] [PubMed]
- Makri, A.; Fakas, S.; Aggelis, G. Metabolic activities of biotechnological interest in Yarrowia lipolytica grown on glycerol in repeated batch cultures. Bioresour. Technol. 2010, 101, 2351–2358. [Google Scholar] [CrossRef] [PubMed]
- Blazeck, J.; Hill, A.; Liu, L.; Knight, R.; Miller, J.; Pan, A.; Otoupal, P.; Alper, H.S. Harnessing Yarrowia lipolytica lipogenesis to create a platform for lipid and biofuel production. Nat. Commun. 2014, 5, 3131. [Google Scholar] [CrossRef] [Green Version]
- Rakicka, M.; Rywińska, A.; Cybulski, K.; Rymowicz, W. Enhanced production of erythritol and mannitol by Yarrowia lipolytica in media containing surfactants. Braz. J. Microbiol. 2016, 47, 417–423. [Google Scholar] [CrossRef] [Green Version]
- Cheng, H.; Wang, S.; Bilal, M.; Ge, X.; Zhang, C.; Fickers, P.; Cheng, H. Identification, characterization of two NADPH-dependent erythrose reductases in the yeast Yarrowia lipolytica and improvement of erythritol productivity using metabolic engineering. Microb. Cell Fact. 2018, 17, 133. [Google Scholar] [CrossRef]
- Diamantopoulou, P.; Filippousi, R.; Antoniou, D.; Varfi, E.; Xenopoulos, E.; Sarris, D.; Papanikolaou, S. Production of added-value microbial metabolites during growth of yeast strains on media composed of biodiesel-derived crude glycerol and glycerol/xylose blends. FEMS Microbiol. Lett. 2020, 367, fnaa063. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Diamantopoulou, P.; Blanchard, F.; Lambrinea, E.; Chevalot, I.; Stoforos, N.G.; Rondags, E. Physiological characterization of a novel wild-type Yarrowia lipolytica strain grown on glycerol: Effects of cultivation conditions and mode on polyols and citric acid production. Appl. Sci. 2020, 10, 7373. [Google Scholar] [CrossRef]
- Imandi, S.B.; Bandaru, V.V.R.; Somalanka, S.R.; Bandaru, S.R.; Garapati, H.R. Application of statistical experimental designs for the optimization of medium constituents for the production of citric acid from pineapple waste. Bioresour. Technol. 2008, 99, 4445–4450. [Google Scholar] [CrossRef]
- Morgunov, I.G.; Kamzolova, S.V.; Lunina, J.N. Citric acid production by Yarrowia lipolytica yeast on different renewable raw materials. Fermentation 2018, 4, 36. [Google Scholar] [CrossRef] [Green Version]
- Finogenova, T.V.; Morgunov, I.G.; Kamzolova, S.V.; Chernyavskaya, O.G. Organic acid production by the yeast Yarrowia lipolytica: A review of prospects. Appl. Biochem. Microbiol. 2005, 41, 418–425. [Google Scholar] [CrossRef]
- Kamzolova, S.V.; Morgunov, I.G. Microbial production of (2 R,3 S)-isocitric acid: State of the arts and prospects. Appl. Microbiol. Biotechnol. 2019, 103, 9321–9333. [Google Scholar] [CrossRef]
- Fickers, P.; Cheng, H.; Sze, K.; Lin, C. Sugar alcohols and organic acids synthesis in Yarrowia lipolytica: Where Are We? Microorganisms 2020, 8, 574. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Kim, M.J.; Yoon, W.; Kim, E.Y.; Kim, H.; Lee, Y.; Min, B.; Kang, K.S.; Son, J.H.; Park, H.T.; et al. Isocitrate protects DJ-1 null dopaminergic cells from oxidative stress through NADP+-dependent isocitrate dehydrogenase (IDH). PLoS Genet. 2017, 13, e1006975. [Google Scholar] [CrossRef] [PubMed]
- Heretsch, P.; Thomas, F.; Aurich, A.; Krautscheid, H.; Sicker, D.; Giannis, A. Syntheses with a chiral building block from the citric acid cycle: (2R,3S)-isocitric acid by fermentation of sunflower oil. Angew. Chem. Int. Ed. Engl. 2008, 47, 1958–1960. [Google Scholar] [CrossRef] [PubMed]
- Aurich, A.; Specht, R.; Müller, R.A.; Stottmeister, U.; Yovkova, V.; Otto, C.; Holz, M.; Barth, G.; Heretsch, P.; Thomas, F.A.; et al. Microbiologically produced carboxylic acids used as building blocks in organic synthesis. In Reprogramming Microbial Metabolic Pathways. Subcellular Biochemistry; Wang, X., Chen, J., Quinn, P., Eds.; Springer: Dordrecht, The Netherlands, 2012; Volume 64, pp. 391–423. [Google Scholar] [CrossRef]
- Aurich, A.; Hofmann, J.; Oltrogge, R.; Wecks, M.; Glaser, R.; Blömer, L.; Mauersberger, S.; Roland, A.; Müller, R.A.; Sicker, D.; et al. Improved isolation of microbiologically produced (2R,3S)-isocitric acid by adsorption on activated carbon and recovery with methanol. Org. Process Res. Dev. 2017, 21, 866–870. [Google Scholar] [CrossRef] [Green Version]
- Bullin, K.; Hennig, L.; Herold, R.; Krautscheid, H.; Richter, K.; Sicker, D. An optimized method for an (2R,3S)-isocitric acid building block. Mon. Chem. 2019, 150, 247–253. [Google Scholar] [CrossRef]
- Kulkarni, M.G.; Shaikh, Y.B.; Borhade, A.S.; Dhondge, A.P.; Chavhan, S.W.; Desai, M.P.; Birhade, D.R.; Dhatrak, N.R.; Gannimani, R. The efficient synthesis of (3R,3aS,6aR)-hexahydrofuro[2,3-b]furan-3-ol and its isomers. Tetrahedron Asymmetry 2010, 21, 2394–2398. [Google Scholar] [CrossRef]
- Moore, G.L.; Stringham, R.W.; Teager, D.S.; Yue, T.Y. Practical synthesis of the bicyclic darunavir side chain: (3R,3aS,6aR)-hexahydrofuro[2,3-b]furan-3-ol from monopotassium isocitrate. Org. Process Res. Dev. 2017, 21, 98–106. [Google Scholar] [CrossRef]
- Morgunov, I.G.; Karpukhina, O.V.; Kamzolova, S.V.; Samoilenko, V.A.; Inozemtsev, A.N. Investigation of the effect of biologically active threo-Ds-isocitric acid on oxidative stress in Paramecium caudatum. Prep. Biochem. Biotechnol. 2018, 48, 1–5. [Google Scholar] [CrossRef]
- Morgunov, I.G.; Kamzolova, S.V.; Karpukhina, O.V.; Bokieva, S.V.; Inozemtsev, A.N. Biosynthesis of isocitric acid in repeated-batch culture and testing of its stress-protective activity. Appl. Microbiol. Biotechnol. 2019, 103, 3549–3558. [Google Scholar] [CrossRef]
- Morgunov, I.G.; Kamzolova, S.V.; Karpukhina, O.V.; Bokieva, S.B.; Lunina, J.N.; Inozemtsev, A.N. Microbiological Production of Isocitric Acid from Biodiesel Waste and Its Effect on Spatial Memory. Microorganisms 2020, 8, 462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finogenova, T.V.; Shishkanova, N.V.; Fausek, E.A.; Eremina, S.S. Biosynthesis of isocitric acid from ethanol by yeasts. Appl. Microbiol. Biotechnol. 1991, 36, 231–235. [Google Scholar] [CrossRef]
- Kamzolova, S.V.; Shamin, R.V.; Stepanova, N.N.; Morgunov, G.I.; Lunina, J.N.; Allayarov, R.K.; Samoilenko, V.A.; Morgunov, I.G. Fermentation conditions and media optimization for isocitric acid production from ethanol by Yarrowia lipolytica. Biomed Res. Int. 2018, 2018, e2543210. [Google Scholar] [CrossRef] [Green Version]
- Kamzolova, S.V.; Dedyukhina, E.G.; Samoilenko, V.A.; Lunina, J.N.; Puntus, I.F.; Allayarov, R.K.; Chiglintseva, M.N.; Mironov, A.A.; Morgunov, I.G. Isocitric acid production from rapeseed oil by Yarrowia lipolytica yeast. Appl. Microbiol. Biotechnol. 2013, 97, 9133–9144. [Google Scholar] [CrossRef] [PubMed]
- Hapeta, P.; Rakicka-Pustułka, M.; Juszczyk, P.; Robak, M.; Rymowicz, W.; Lazar, Z. Overexpression of citrate synthase increases isocitric acid biosynthesis in the yeast Yarrowia lipolytica. Sustainability 2020, 12, e7364. [Google Scholar] [CrossRef]
- Kamzolova, S.V.; Samoilenko, V.A.; Lunina, J.N.; Morgunov, I.G. Effects of medium components on isocitric acid production by Yarrowia lipolytica yeast. Fermentation 2020, 6, 112. [Google Scholar] [CrossRef]
- Förster, A.; Jacobs, K.; Juretzek, T.; Mauersberger, S.; Barth, B. Overexpression of the ICL1 gene changes the product ratio of citric acid production by Yarrowia lipolytica. Appl. Microbiol. Biotechnol. 2007, 77, 861–869. [Google Scholar] [CrossRef]
- Holz, M.; Förster, A.; Mauersberger, S.; Barth, G. Aconitase overexpression changes the product ratio of citric acid production by Yarrowia lipolytica. Appl. Microbiol. Biotechnol. 2009, 81, 1087–1096. [Google Scholar] [CrossRef]
- Yuzbasheva, E.Y.; Scarcia, P.; Yuzbashev, T.V.; Messina, E.; Kosikhina, I.M.; Palmieri, L.; Shutov, A.V.; Taratynova, M.O.; Amaro, R.L.; Palmieri, F.; et al. Engineering Yarrowia lipolytica for the selective and high-level production of isocitric acid through manipulation of mitochondrial dicarboxylate-tricarboxylate carriers. Metab. Eng. 2021, 65, 156–166. [Google Scholar] [CrossRef]
- Da Silva, L.V.; Tavares, C.B.; Amaral, P.F.F.; Coehlo, M.A.Z. Production of citric acid by Yarrowia lipolytica in different crude oil concentrations and in different nitrogen sources. Chem. Eng. Trans. 2012, 27, 199–204. [Google Scholar]
- Rzechonek, D.A.; Dobrowolski, A.; Rymowicz, W.; Mirończuk, A.M. Aseptic production of citric and isocitric acid from crude glycerol by genetically modified Yarrowia lipolytica. Bioresour. Technol. 2019, 271, 340–344. [Google Scholar] [CrossRef]
- Kamzolova, S.V.; Samoilenko, V.A.; Lunina, J.N.; Morgunov, I.G. Isocitric acid production from ethanol industry waste by Yarrowia Lipolytica. Fermentation 2021, 7, 146. [Google Scholar] [CrossRef]
- Tsygankov, P.S.; Tsygankov, S.P. Handbook on Alcohol Rectification; Pishchepromizdat: Moscow, Russia, 2002; p. 400. (In Russian) [Google Scholar]
- Karklin, R.; Peltzmane, I.; Raminya, L.; Korde, G. Overproduction of isocitric acid by wild strain of Candida lipolytica. In Metabolism of n-Alkanes and Oversynthesis of Products by Microorganisms; USSR Academy of Sciences, Institute of Biochemistry and Physiology of Microorganisms: Pushchino, Russia, 1991; pp. 143–146. (In Russian) [Google Scholar]
- Kamzolova, S.V.; Allayarov, R.K.; Lunina, J.N.; Morgunov, I.G. The effect of oxalic and itaconic acids on threo-Ds-isocitric acid production from rapeseed oil by Yarrowia lipolytica. Bioresour. Technol. 2016, 206, 128–133. [Google Scholar] [CrossRef]
- Bradford, M.M. Rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Il’chenko, A.P.; Chernyavskaya, O.G.; Shishkanova, N.V.; Finogenova, T.V. The Induction of cytochrome P-450 and ethanol oxidation in Yarrowia lipolytica cells. Microbiology 2003, 72, 138–143. [Google Scholar] [CrossRef]
- Il’chenko, A.P.; Chernyavskaya, O.G.; Finogenova, T.V. Ethanol metabolism in the yeasts Yarrowia and Torulopsis: A Review. Appl. Biochem Microbiol. 2005, 41, 426–432. [Google Scholar] [CrossRef]
- Kujau, M.; Weber, H.; Barth, G. Characterization of mutants of the yeast Yarrowia lipolytica defective in acetyl-coenzyme A synthetase. Yeast 1992, 8, 193–203. [Google Scholar] [CrossRef]
- Gatter, M.; Ottlik, S.; Kövesi, Z.; Bauer, B.; Matthäus, F.; Barth, G. Three alcohol dehydrogenase genes and one acetyl-CoA synthetase gene are responsible for ethanol utilization in Yarrowia lipolytica. Fungal Genet. Biol. 2016, 95, 30–38. [Google Scholar] [CrossRef]
- Holdsworth, J.E.; Veenhuis, H.; Ratledge, C. Enzyme activities in oleaginous yeasts accumulating and utilizing exogenous or endogenous lipids. J. Gen. Microbiol. 1988, 134, 2907–2915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fickers, P.; Benetti, P.-H.; Waché, Y.; Marty, A.; Mauersberger, S.; Smit, M.S.; Nicaud, J.-M. Hydrophobic substrate utilisation by the yeast Yarrowia lipolytica, and its potential applications. FEMS Yeast Res. 2005, 5, 527–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papanikolaou, S.; Aggelis, G. Yarrowia lipolytica: A model microorganism used for the production of tailor-made lipids. Eur. J. Lipid Sci. Technol. 2010, 112, 639–654. [Google Scholar] [CrossRef]
- Sabra, W.; Bommareddy, R.R.; Maheshwari, G.; Papanikolaou, S.; Zeng, A.-P. Substrates and oxygen dependent citric acid production by Yarrowia lipolytica: Insights through transcriptome and fluxome analyses. Microb. Cell Fact. 2017, 16, 78. [Google Scholar] [CrossRef]
- Stahmann, K.P.; Revuelta, J.L.; Seulberger, H. Three biotechnical processes using Ashbya gossypii, Candida famata, or Bacillus subtilis compete with chemical riboflavin production. Appl. Microbiol. Biotechnol. 2000, 53, 509–516. [Google Scholar] [CrossRef]
- Berg, I.A.; Filatova, L.V.; Ivanovsky, R.N. Inhibition of acetate and propionate assimilation by itaconate via propionyl-CoA carboxylase in isocitrate lyase-negative purple bacterium Rhodospirillum rubrum. FEMS Microbiol. Lett. 2002, 216, 49–54. [Google Scholar] [CrossRef]
- Krátký, M.; Vinšová, J. Advances in mycobacterial isocitrate lyase targeting and inhibitors. Curr. Med. Chem. 2012, 19, 6126–6137. [Google Scholar] [CrossRef]
- Thomas, K.C.; Dawson, P.S.S. Relationship between iron-limited growth and energy limitation during phased cultivation of Candida utilis. Can. J. Microbiol. 1978, 24, 440–447. [Google Scholar] [CrossRef]
- Hoffmann, A.; Kupsch, C.; Walther, T.; Löser, C. Synthesis of ethyl acetate from glucose by Kluyveromyces marxianus, Cyberlindnera jadinii and Wickerhamomyces anomalus depending on the induction mode. Eng. Life Sci. 2021, 21, 154–168. [Google Scholar] [CrossRef]
- Laptev, I.A.; Filimonova, N.A.; Allayarov, R.K.; Kamzolova, S.V.; Samoilenko, V.A.; Sineoky, S.P.; Morgunov, I.G. New recombinant strains of the yeast Yarrowia lipolytica with overexpression of the aconitate hydratase gene for the obtainment of isocitric acid from rapeseed oil. Appl. Biochem. Microbiol. 2016, 52, 699–704. [Google Scholar] [CrossRef]
- Finogenova, T.V. Biosynthesis of Organic Acids by Yeast Organisms and its Regulation. Ph.D. Thesis, USSR Academy of Sciences, Institute of Biochemistry and Physiology of Microorganisms, Pushchino, Russia, 1982. [Google Scholar]
- Babel, W. The auxiliary substrate concept: From simple considerations to heuristically valuable knowledge. Eng. Life Sci. 2009, 9, 285–290. [Google Scholar] [CrossRef]
- Müller, R.; Markuske, K.D.; Babel, W. Improvement of Y-values of Hansenula polymorpha growth on methanol by simultaneous utilization of glucose. Z. Allg. Mikrobiol. 1983, 23, 375–384. [Google Scholar] [CrossRef] [PubMed]
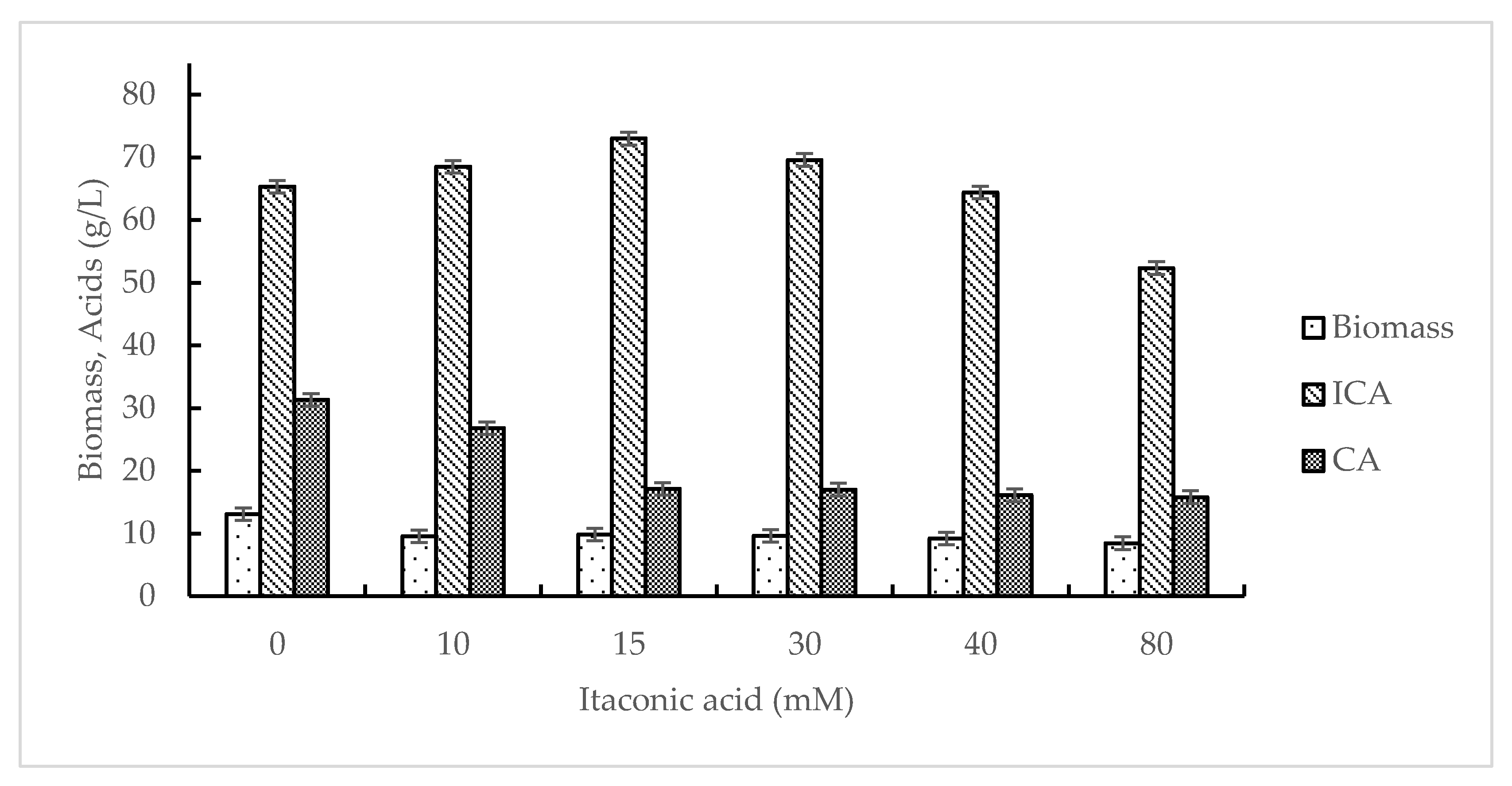


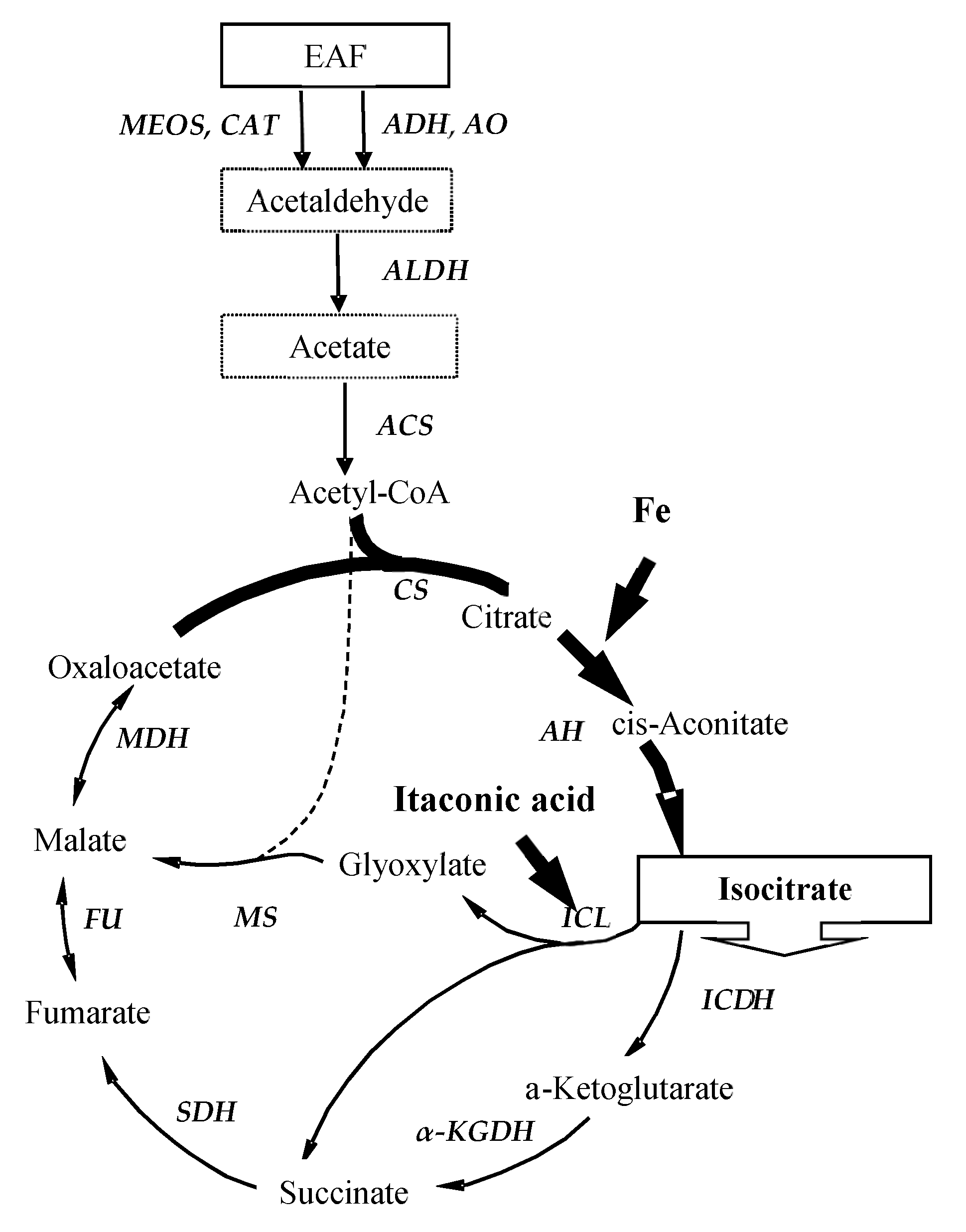

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Indices | Control | Itaconic Acid (mM) | ||||
|---|---|---|---|---|---|---|
| 10 | 15 | 30 | 40 | 80 | ||
| ICA/CA ratio | 2.1:1 | 2.6:1 | 4.3:1 | 4.1:1 | 4:1 | 3.3:1 |
| Ethanol consumed (g/L) | 890 | 800 | 757 | 779 | 670 | 570 |
| YICA (g/g) | 0.78 | 0.90 | 0.97 | 0.83 | 0.81 | 0.79 |
| qICA (g/g ·h) | 0.052 | 0.068 | 0.078 | 0.076 | 0.073 | 0.064 |
| QICA (g/L ·h) | 0.95 | 1.12 | 1.18 | 1.13 | 1.07 | 1.00 |
| Enzymes | Control | Itaconic Acid (mM) | ||||
|---|---|---|---|---|---|---|
| 10 | 15 | 30 | 40 | 80 | ||
| ADH | 0.075 ± 0.007 | 0.072 ± 0.007 | 0.075 ± 0.006 | 0.071 ± 0.007 | 0.071 ± 0.006 | 0.060 ± 0.005 |
| AlDH | 0.072 ± 0.007 | 0.072 ± 0.006 | 0.070 ± 0.006 | 0.072 ± 0.006 | 0.071 ± 0.006 | 0.071 ± 0.006 |
| Catalase | 310 ± 15 | 290 ± 10 | 300 ± 15 | 300 ± 15 | 285 ± 10 | 260 ± 8 |
| CS | 2.875 ± 0.100 | 2.520 ± 0.080 | 2.445 ± 0.100 | 2.315 ± 0.090 | 2.125 ± 0.080 | 2.025 ± 0.075 |
| AH | 0.405 ± 0.020 | 0.355 ± 0.015 | 0.345 ± 0.020 | 0.325 ± 0.020 | 0.285 ± 0.010 | 0.280 ± 0.010 |
| NAD-ICDH | 0.100 ±0.005 | 0.094 ± 0.005 | 0.091 ± 0.004 | 0.080 ± 0.005 | 0.078 ± 0.004 | 0.069 ± 0.003 |
| ICL | 0.231 ± 0.002 | 0.025 ± 0.001 | 0.013 ± 0.001 | 0.010 ± 0.001 | 0.012 ± 0.001 | 0.012 ± 0.001 |
| MS | 0.068 ± 0.005 | 0.030 ± 0.004 | 0.030 ± 0.004 | 0.030 ± 0.003 | 0.030 ± 0.002 | 0.030 ± 0.002 |
| Indices | Iron Concentration (mg/L) | |||||
|---|---|---|---|---|---|---|
| 0.001 | 0.3 | 0.6 | 1.2 | 2.4 | 5.0 | |
| ICA/CA ratio | n.d. | 1.8:1 | 3.3:1 | 4.1:1 | 4.1:1 | 3:1 |
| Ethanol consumed (g/L) | 60 | 750 | 760 | 757 | 840 | 320 |
| YICA (g/g) | n.d. | 0.66 | 0.92 | 0.97 | 1.1 | 0.35 |
| qICA (g/g·h) | n.d. | 0.054 | 0.069 | 0.078 | 0.088 | 0.038 |
| QICA (g/L·h) | n.d. | 0.85 | 1.16 | 1.18 | 1.25 | 0.44 |
| Enzymes | Iron Concentration (mg/L) | |||||
|---|---|---|---|---|---|---|
| 0.001 | 0.3 | 0.6 | 1.2 | 2.4 | 5 | |
| ADH | 0.04 ± 0.001 | 0.060 ± 0.006 | 0.075 ± 0.006 | 0.075 ± 0.007 | 0.070 ± 0.007 | 0.035± 0.008 |
| AlDH | 0.010 ± 0.001 | 0.030 ± 0.003 | 0.050 ± 0.006 | 0.070 ± 0.006 | 0.070 ± 0.006 | 0.080 ± 0.007 |
| Catalase | 48.5 ± 4 | 140 ± 10 | 200 ± 12 | 330 ± 10 | 340 ± 10 | 380 ± 15 |
| CS | 1.425 ± 0.112 | 2.000 ± 0.100 | 2.100± 0.1 | 2.445 ± 0.100 | 2.500 ± 0.120 | 1.625 ± 0.090 |
| AH | 0.040 ± 0.005 | 0.120 ± 0.015 | 0.290 ± 0.020 | 0.345 ± 0.020 | 0.600± 0.030 | 0.620 ± 0.040 |
| NAD-ICDH | 0.02 ± 0.003 | 0.07 ± 0.005 | 0.09 ± 0.004 | 0.091 ± 0.004 | 0.080 ± 0.006 | 0.050 ± 0.005 |
| ICL | 0.015 ± 0.001 | 0.015± 0.001 | 0.013 ± 0.001 | 0.013 ± 0.001 | 0.010 ± 0.001 | 0.010 ± 0.001 |
| MS | 0.025 ± 0.003 | 0.030 ± 0.003 | 0.030 ± 0.003 | 0.030 ± 0.003 | 0.025 ± 0.003 | 0.025 ± 0.003 |
| Indices | pO2 (% of Air Saturation in O2) | ||||
|---|---|---|---|---|---|
| 5 | 20 | 40 | 60 | 80 | |
| ICA/CA ratio | 5.1:1 | 4.1:1 | 4.1:1 | 4.1:1 | 4.1:1 |
| Ethanol consumed (g/L) | 260 | 590 | 738 | 840 | 570 |
| YICA (g/g) | 0.57 | 0.88 | 0.88 | 1.1 | 0.72 |
| qICA (g/g·h) | 0.061 | 0.061 | 0.070 | 0.088 | 0.080 |
| QICA (g/L·h) | 0.38 | 0.87 | 1.10 | 1.25 | 0.84 |
| Enzymes | pO2 (% of Air Saturation in O2) | ||||
|---|---|---|---|---|---|
| 5 | 20 | 40 | 60 | 80 | |
| ADH | 0.056± 0.005 | 0.070 ± 0.006 | 0.070 ± 0.006 | 0.070 ± 0.007 | 0.035 ± 0.005 |
| AlDH | 0.060 ± 0.006 | 0.070± 0.006 | 0.070 ± 0.006 | 0.070 ± 0.006 | 0.070 ± 0.006 |
| Catalase | 283 ± 12 | 340 ± 10 | 340 ± 12 | 340 ± 10 | 340 ± 15 |
| CS | 1.25 ± 0.100 | 1.625 ± 0.110 | 2.225 ± 0.110 | 2.500 ± 0.120 | 1.380 ± 0.110 |
| AH | 0.360 ± 0.030 | 0.486 ± 0.040 | 0.570 ± 0.050 | 0.600 ± 0.030 | 0.310 ± 0.030 |
| NAD-ICDH | 0.056 ± 0.005 | 0.070 ± 0.007 | 0.080 ± 0.004 | 0.080 ± 0.005 | 0.045 ± 0.004 |
| ICL | 0.002 ± 0.001 | 0.010± 0.001 | 0.010 ± 0.001 | 0.010 ± 0.001 | 0.012 ± 0.001 |
| MS | 0.005 ± 0.001 | 0.025± 0.002 | 0.025 ± 0.002 | 0.025 ± 0.003 | 0.022 ± 0.003 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kamzolova, S.V.; Morgunov, I.G. Effect of Metabolic Regulators and Aeration on Isocitric Acid Synthesis by Yarrowia lipolytica Grown on Ester-Aldehyde Fraction. Fermentation 2021, 7, 283. https://doi.org/10.3390/fermentation7040283
Kamzolova SV, Morgunov IG. Effect of Metabolic Regulators and Aeration on Isocitric Acid Synthesis by Yarrowia lipolytica Grown on Ester-Aldehyde Fraction. Fermentation. 2021; 7(4):283. https://doi.org/10.3390/fermentation7040283
Chicago/Turabian StyleKamzolova, Svetlana V., and Igor G. Morgunov. 2021. "Effect of Metabolic Regulators and Aeration on Isocitric Acid Synthesis by Yarrowia lipolytica Grown on Ester-Aldehyde Fraction" Fermentation 7, no. 4: 283. https://doi.org/10.3390/fermentation7040283
APA StyleKamzolova, S. V., & Morgunov, I. G. (2021). Effect of Metabolic Regulators and Aeration on Isocitric Acid Synthesis by Yarrowia lipolytica Grown on Ester-Aldehyde Fraction. Fermentation, 7(4), 283. https://doi.org/10.3390/fermentation7040283