Abstract
Gas fermentation by acetogens of the genus Clostridium is an attractive technology since it affords the production of biochemicals and biofuels from industrial waste gases while contributing to mitigate the carbon cycle alterations. The acetogenic model organisms C. ljungdahlii and C. autoethanogenum have already been used in large scale industrial fermentations. Among the natural products, ethanol production has already attained industrial scale. However, some acetogens are also natural producers of 2,3-butanediol (2,3-BDO), a platform chemical of relevant industrial interest. Here, we have developed a lab-scale screening campaign with the aim of enhancing 2,3-BDO production. Our study generated comparable data on growth and 2,3-BDO production of several batch gas fermentations using C. ljungdahlii and C. autoethanogenum grown on different gas substrates of primary applicative interest (CO2 · H2, CO · CO2, syngas) and on different media featuring different compositions as regards trace metals, mineral elements and vitamins. CO · CO2 fermentation was found to be preferable for the production of 2,3-BDO, and a fair comparison of the strains cultivated in comparable conditions revealed that C. ljungdahlii produced 3.43-fold higher titer of 2,3-BDO compared to C. autoethanogenum. Screening of different medium compositions revealed that mineral elements, Zinc and Iron exert a major positive influence on 2,3-BDO titer and productivity. Moreover, the CO2 influence on CO fermentation was explored by characterizing C. ljungdahlii response with respect to different gas ratios in the CO · CO2 gas mixtures. The screening strategies undertaken in this study led to the production of 2.03 ± 0.05 g/L of 2,3-BDO, which is unprecedented in serum bottle experiments.
1. Introduction
Gas fermentation mediated by acetogenic bacteria of the genus Clostridium enables carbon capture and conversion of greenhouse gases into useful products [1,2,3,4]. Therefore, the gas fermentation technology meets a two-fold objective, contributing to counteract the effects of the current carbon cycle alterations and, simultaneously, embodying carbon circular industrial biomanufacturing models [5]. Gas fermentation technology offers a unique level of feedstock flexibility. In fact, the gaseous feedstock may be the direct waste streams of industrial processes, such as steel manufacturing and oil refineries, or syngas of variable composition generated from gasification of an ample range of carbon-containing wastes [6]. To reflect such a high feedstocks’ flexibility, the synthetic gas mixtures used to grow acetogens in lab-scale gas fermentations are highly diversified and include CO · CO2 [7,8,9], CO2 · H2 [10,11,12] and syngas [6,13,14]. Compared to conventional chemocatalytic or thermochemical processes, gas fermentation benefits include milder process conditions, higher product selectivity owing to high enzymatic conversion specificities, higher tolerance to impurities which are known to poison inorganic catalysts [2], and higher flexibility to the employed feedstocks [15,16]. Nevertheless, the gas fermentation primary limitations are the poor gas to liquid mass transfer, due to the low solubility of the gas substrates [17], by the slow growth of acetogens that strive at the thermodynamic edge of life [18] and by low product yields [19].
Nonetheless, these challenging aspects do not represent unsolvable barriers to the use of gas fermentation as a commercial production system. In fact, industrial-scale processes have already been developed for ethanol production [20,21,22]. Although industrially relevant process parameters have not been achieved yet, process scalability has also been proven for the production of 2,3-butanediol (2,3-BDO), with a productivity of up to 45 g/L/d achieved by Simpson and co-workers [23]. 2,3-BDO is a valuable platform chemical [24], which can be used to produce butadiene, acetoin, diacetyl, methyl ethyl ketone and as a novel chain initiator and extender of polyurethane foams [25]. Acetogens of the genus Clostridium are common biocatalysts for gas fermentation applications since they feature among the highest growth rates and are natural producers of metabolites of commercial interest, such as ethanol and 2,3-BDO [26,27]. Clostridium ljungdahlii [28,29] and Clostridium autoethanogenum [30,31] are the most intensively investigated acetogenic biocatalysts.
The identification of the biocatalyst(s) and gaseous substrate(s) suitable for the production of the target product is paramount to ensure the viability of the entire process. Some studies were reliant on a single acetogen and explored its growth and product profile against several gaseous substrates. This is the case for C. autoethanogenum [12,32,33,34,35] and for C. ljungdahlii [7,8,9,36]. In some other studies, several strains were comparatively screened against a certain gaseous substrate [26,27,37,38,39]. Moreover, the identification of process conditions that act to enhance culture growth rates and/or productivities is necessary to improve the biological conversion profitability. Therefore, several studies focused on the screening of different media using a single strain [40,41] or multiple strains [37]. In spite of the undoubtful value of each study, gaining a thorough understanding of the sensitivity of C. autoethanogenum and C. ljungdahlii growth and product profile to variations in the gaseous substrate and/or medium is puzzling. This is severely hindered by the fact that the data on autotrophic growth and product formation are rarely acquired in comparable cultivation conditions [38].
Our study acquired comparable data on the growth and metabolite profile of a wide range of batch gas fermentations carried out by C. autoethanogenum and C. ljungdahlii in order to individuate the gas substrates and medium compositions that afford enhanced 2,3-BDO production. To this aim, both the model acetogens were grown on several gaseous substrates, which explored the inclusion of different gases or the usage of different relative gas ratios, and on four media featuring different compositions as regards trace metals, mineral elements and vitamins. Further provided is a pre-screening aimed at individuating a cultivation technique able to enhance 2,3-BDO titer and productivity. More precisely, the screening campaign was structured as follows. Firstly, we explored the cultivation technique for autotrophic batch experiments in serum bottles, by comparing the results obtained using two different volumetric gas to liquid ratios (4:1 and 9:1) and two different bottle orientations during agitation (vertical and horizontal orientation). We then identified the gas substrate and biocatalyst which promoted the production of 2,3-BDO by screening growth and product profiles of C. autoethanogenum and C. ljungdahlii on three gas substrates—CO2 · H2, CO · CO2 and simulated syngas. Subsequently, we investigated the influence of medium composition on 2,3-BDO titers and productivities by testing different media with the previously selected biocatalyst and gas substrate. Finally, the study of the 2,3-BDO production process encompassed a further level of investigation of the gas substrate, by exploring how different gas ratios impact on the metabolites’ profile.
2. Materials and Methods
2.1. Bacterial Strains and Medium Compositions
C. autoethanogenum DSM 10061 and C. ljungdahlii DSM 13582 were purchased from DSMZ (Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH, Braunschweig, Germany) and cultivated under strictly anaerobic conditions. To characterize growth and production profiles of the two acetogenic bacteria, four different cultivation media were employed. The “Tanner_mod” medium is a modified Tanner medium [42] as it is based on the medium proposed by Bengelsdorf and co-workers [27] and foresees modified concentrations of 2-(N-morpholino)ethanesulfonic acid (MES), ZnSO4, Na2SeO3, vitamin B12 and omits sodium-2-mercaptoethansulfonate (MESNA) (Table 1). The “Valgepea_mod” medium is a modified Valgepea medium as it is based on the medium introduced by Valgepea and co-workers [13] but foresees the addition of MES and different cysteine and resazurin concentrations (Table 1). The “Tan_Val” medium is introduced for the first time in this study and results from the combination of the mineral solution of the “Tanner_mod” medium with the trace element solution and vitamin solution of the “Valgepea_mod” medium. Finally, the “Tan_Val + Fe’’ medium differs from the “Tan_Val” medium only by the addition of 0.017 g/L of FeCl3 × 6 H2O. The detailed recipes and compositions of the four media used in this study are reported in Table 1. The pH of each medium employed was adjusted to 5.9 with NaOH 10 M. Under heterotrophic growth conditions, 5 g/L of fructose was added in each medium. All chemicals employed in this study were purchased from Sigma-Aldrich (St. Louis, MO, USA), Alpha Aesar (Haverhill, MA, United States), Fisher BioReagents by Thermo Fisher Scientific (Waltham, MA, USA), Carl Roth GmbH + Co. KG (Karlsruhe, Germany), Janssen Pharmaceuticals (Geel, Belgium), and Merck KGaA (Damstadt, Germany).

Table 1.
Overview of media compositions. In the table are reported the compositions of all the media tested in this study, in gram (g), milligram (mg) of milliliter (mL) per liter of solutions. * These elements have been added through the mineral solution. ** These elements, in the “Valgepea_mod” medium, have been added directly to the medium.
2.2. Batch Gas Fermentation Experiments
Autotrophic growth experiments were carried out in batch conditions using 250 mL serum bottles. The working volume was 50 mL for heterotrophic cultivation and 25 mL for autotrophic cultivation, unless differently specified. The gas mixtures considered in our screening consisted of CO2 and H2, or CO and CO2 or simulated syngas. In the CO · CO2 case, we also considered different gas ratios. Therefore, overall six different gas compositions were tested: CO2 · H2 in the ratio 1:4 (20% CO2 and 80% H2), CO · CO2 in the ratio 4:1 (80% CO and 20% CO2), CO · CO2 in the ratio 1:1 (50% CO and 50% CO2), CO · CO2 in the ratio 1:4 (20% CO and 80% CO2), pure CO (100% CO) and simulated syngas consisting of CO, CO2, H2, N2 in the ratio 5:2:2:1 (50% CO, 20% CO2, 20% H2 and 10% N2) which was inspired by the gas mixture adopted by Valgepea and co-workers [13]. The above gas mixtures were prepared in situ by using a customized gas station equipped with four Mass Flow Controllers (MFCs) (Burkert, Germany) and a gas mixer (Tecnodelta Srl, Turin, Italy). Serum bottles were pressurized to a final overpressure of 0.75 bar. For each gas fermentation experiment the same inoculation procedure was adopted. Glycerol stocks of C. autoethanogenum or C. ljungdahlii were reactivated in the “Tanner_mod” medium added with 5 g/L of fructose. Strain adaptation to a specific medium (“Tanner_mod”, “Valgepea_mod”, “Tan_Val” or “Tan_Val + Fe”) was carried out in heterotrophy. A mid-exponential grown culture was used as inoculum for autotrophic pre-culture preparation. The autotrophic pre-culture step made use of the specific gaseous substrate that needed to be evaluated. Gas fermentation experiments were inoculated with the autotrophic pre-culture in mid-late exponential growth phase. Tests were run at least in triplicate, at 37 °C and horizontally (or vertically, when specified) agitated at 100 rpm. Sampling for analysis of growth, pH and products’ profile occurred daily by removing 1 mL of liquid from the bottles, using a sterile syringe. Depending on the experiment, from 10 to 15 sampling points were evaluated. Experiments’ duration ranged between 10 and 15 days. Cell growth was constantly monitored by measuring optical density at 600 nm (OD600 nm), using a cell density meter model 40 (Thermo Fisher Scientific, Waltham, MA, USA), the pH of the cultures was monitored via pH meter Accumet® AB150 pH/mV pH meter (Thermo Fisher Scientific, Waltham, MA, USA), and the concentrations of products of interest were measured by high performance liquid chromatography (HPLC).
2.3. Calculation of Biomass Concentration
Biomass concentration (gCDW/L) was estimated through two different OD to cell dry weight (CDW) correlation coefficients. A correlation coefficient of 0.21 was employed for C. autoethanogenum as previously described by Valgepea and co-workers [13], whereas a correlation coefficient of 0.30 was employed for C. ljungdahlii as previously described by Infantes and co-workers [43].
2.4. Calculation of Batch Fermentation Metrics
Acetate, ethanol and 2,3-BDO productivities were calculated in grams/liter/hour (g/L/h) by dividing the difference of the titer (g/L) of a product at two consecutive sampling points by the number of hours between the two sampling points. This calculation was carried out for every sampling point in an experiment and only the maximum productivity was reported in the Supplementary Tables. The specific growth rates were calculated in grams of cell dry weight (gCDW)/liter/hour (gCDW/L/h), considering only the exponential growth phases and by dividing the difference of the natural logarithm of the CDW values at two different growth points by the number of hours between the two growth points. Finally, products’ ratios were calculated by dividing the titer (g/L) of two different products.
2.5. Analytics
Quantification of fructose, acetate, ethanol, R,R-2,3-butanediol, meso-2,3-butanediol and lactate was carried out using a “Dionex UltiMate 3000” HPLC system (Thermo Fisher Scientific, Waltham, MA, USA) coupled with a refractive index detector “RefractorMax 520” (RID) (ERC, Tokyo, Japan) operating at 55 °C and a “Dionex UltiMate 3000” variable wavelength detector (Thermo Fisher Scientific, Waltham, MA, USA) set at 210 nm. The “Metab-AAC BF’’ series column (length = 300 mm; ID = 7.8 mm) (Isera, Düren, Germany) with a pre-column was eluted isocratically with 9 mM H2SO4 at a flow rate of 0.6 mL/min and an oven temperature of 40 °C. Slightly acidified water (0.009 M H2SO4) was employed as the mobile phase. Prior to HPLC analysis, culture samples were centrifuged for 10 min at 10,000 rpm in a MicroCL 17R Centrifuge (Thermo Fisher Scientific, Waltham, MA, USA) and filtered through 0.22 μm filters (Scharlab, Barcelona, Spain).
2.6. Data Analysis
Data analysis and graphs were carried out via the GraphPad Prism 9.2.0 (GraphPad, San Diego, CA, USA) software. All data derived from triplicate experiments are shown as mean values ± standard error of the mean (SEM). ANOVA and the follow-up Tukey test were performed in order to compare the means of the distribution of the CDW or of the titer of a certain product among multiple conditions. Multiplicity adjusted Tukey’s test p-values are shown in relation to each comparison displayed in the plots. Single, paired, and one-tailed t-tests were carried out to compare the means of the distributions of a certain observable obtained in two different conditions.
3. Results and Discussion
3.1. Cultivation Technique Optimization for the Autotrophic Growth of a Model Acetogen
Prior to the gaseous substrate screening, we tested different cultivation conditions in the attempt to improve the gas to liquid mass transfer and, consequently, the autotrophic C. ljungdahlii growth and 2,3-butanediol (2,3-BDO) production [3]. Ensuring that the gas substrate is available for cellular uptake is fundamental in the microbial conversion of gaseous substrates to bioproducts. Gas substrate availability is influenced by the transport of the gas from the gaseous phase to the liquid phase and then to the cell surface [36,44,45]. At the lab-scale, microbial gas fermentation is generally carried out in serum bottles [32,34] and, in this case, the primary factor influencing the gas to liquid mass transfer is the gas to liquid volumetric ratio [37].
Here, we compared three different cultivation methods to enhance the gas to liquid mass transfer in serum bottles. C. ljungdahlii was grown on the “Tanner_mod”, a modified Tanner medium [27,42], according to the specifications provided in the Materials and Methods section. We adopted the cultivation method consisting of vertically incubated 250 mL serum bottles containing 50 mL of liquid medium [34,46] as the control. The second condition tested foresaw an increase in the gas to liquid volumetric ratio from 4:1 to 9:1. In the third condition tested, serum bottles with a gas to liquid volumetric ratio of 9:1 were placed horizontally to verify whether the increased gas-liquid contact surface could positively influence cellular growth or products’ profile. Thus, the three conditions tested can be summarized as follows, (i) vertically incubated bottles with a gas to liquid ratio of 4:1; (ii) vertically incubated bottles with a gas to liquid ratio of 9:1 and (iii) horizontally incubated bottles with a gas to liquid ratio of 9:1. C. ljungdahlli was autotrophically cultivated in parallel using each cultivation condition and CO · CO2 4:1 (80% CO and 20% CO2) as the gaseous substrate. C. ljungdahlii growth was found to take advantage of increasing the gas to liquid volumetric ratio but was not influenced by the orientation used during incubation. Indeed, maximal OD600 nm values of 0.83 ± 0.02, 1.46 ± 0.12 and 1.39 ± 0.03 were obtained using the 4:1 vertical, 9:1 vertical and 9:1 horizontal cultivation conditions, respectively (Figure 1A). As concerns products’ formation, the choice of the cultivation method mildly influenced the formation of acetate (Figure S1) and ethanol, where the maximal titer obtained with the 9:1 vertical condition (0.29 ± 0.01 g/L) was 1.52-fold higher than the minimal one, attained using the 4:1 vertical condition (0.19 ± 0.00 g/L) (Figure 1B). Instead, the most pronounced influence of the cultivation mode was observed on 2,3-BDO production. In particular, adopting a gas to liquid ratio of 9:1 and a horizontal orientation of the serum bottles afforded a 2,3-BDO titer of 1.25 ± 0.04 g/L, which is 1.26-fold and 5-fold higher than the titers obtained when C. ljungdahlii was cultivated with a gas to liquid ratio of 9:1 and vertically incubated serum bottles (0.99 ± 0.04 g/L) or with a gas to liquid ratio of 4:1 and vertically incubated serum bottles (0.25 ± 0.01 g/L), respectively (Figure 1C). These results confirmed the observation made by Lee and co-workers [37] whereby supplying a higher amount of gaseous substrate is necessary in order to obtain 2,3-BDO autotrophic production. Interestingly, also 2,3-BDO productivities and ethanol:2,3-BDO ratio were greatly influenced by the cultivation technique (Tables S1 and S2). In fact, the 2,3-BDO productivity increased by 5.75-fold in the 9:1 vertical configuration (0.023 ± 0.002 g/L/h) compared to the 4:1 vertical one (0.004 ± 0.000 g/L/h). An additional increment in 2,3-BDO productivity was obtained by testing the 9:1 horizontal configuration, where 2,3-BDO productivity was 0.027 ± 0.001 g/L/h (Table S1). Correspondingly, the 9:1 horizontal cultivation condition improved the ethanol:2,3-BDO ratio which equated 1:4.90 (Table S2).

Figure 1.
Comparative analysis of growth and production profile of C. ljungdahlii cultivated in CO · CO2 4:1 and using different cultivation techniques. Evaluated configurations were vertically incubated bottles with a gas to liquid ratio of 4:1 (rosewood red), vertically incubated bottles with a gas to liquid ratio of 9:1 (Spanish orange), and horizontally incubated bottles with a gas to liquid ratio of 9:1 (Indian yellow), during 250 mL serum bottles batch experiments. (A) Growth profile in the three cultivation conditions. (B) Ethanol maximum titer, measured at day 5, 6 and 7 for the 4:1-vertical, 9:1-vertical and 9:1-horizontal conditions, respectively. (C) 2,3-butanediol maximum titer, measured at day 5 for the 4:1-vertical and 9:1-vertical conditions and at day 7 for the 9:1-horizontal condition. The mean of three independent replicates is plotted for each tested condition. Error bars indicate the standard error of the mean (SEM). ANOVA P: ethanol (p < 0.0001); 2,3-BDO (p < 0.0001). Tukey’s test p: ** p < 0.0021; *** p < 0.0002 and **** p < 0.0001.
It is plausible that 2,3-BDO production is more favored than acetate or ethanol by the increased turbulence created by the agitation in horizontal mode since increasingly reduced products such as 2,3-BDO particularly benefit from an excess supply of energy (e.g., electrons from CO) [47]. Due to the increased 2,3-BDO titer obtained by using a gas to liquid ratio of 9:1 and a horizontal incubation, this cultivation configuration was used as the reference cultivation technique for performing all the subsequent experiments included in this work.
3.2. Screening of Gaseous Substrates for Enhanced 2,3-BDO Production Using Clostridium ljungdhalii and Clostridium autoethanogenum
The gaseous substrate composition alters the autotrophic metabolism and the production profile of a fermentation culture [7,8,10,33,48]. In spite of the breadth of studies focusing on different gaseous mixtures and strains [7,12,14], capitalizing on the reported outcomes is penalized by the scarce comparability between different studies [38]. Here, to overcome this limitation and ensure an accurate comparability of the experimental outcomes, we maintained the same experimental setup (inoculum, temperature, agitation speed, pH, and pressure) and sampling technique in each batch fermentation, carried out to explore the effects of different gas substrates on products’ formation. C. ljungdahlii and C. autoethanogenum were grown on the “Tanner_mod” medium” [27,42]. The three gaseous substrates explored included: (i) CO2 · H2 in the ratio 1:4 (20% CO2 and 80% H2); (ii) CO · CO2 in the ratio 4:1 (80% CO and 20% CO2) and (iii) simulated syngas consisting of CO, CO2, H2, N2 in the ratio 5:2:2:1 (50% CO, 20% CO2, 20% H2 and 10% N2). Gas ratios and percentages will no longer be reported explicitly, except for the CO · CO2 case for which the gas ratio influence was investigated in Section 3.4.
Growth and production metrics corresponding to C. ljungdahlii cultures inoculated with a headspace of each of the aforementioned gaseous mixture were comparatively assessed (Figure 2 and Figure S2, Tables S3 and S4). As shown in Figure 2A, the CO2 · H2 gas mixture, allowed to reach the least OD600nm, compared to CO · CO2 4:1 and syngas. Indeed, the maximum OD600nm attained under CO2 · H2 was only 0.53 ± 0.02, in accordance with previous observations [8,49]. When looking into the effects of the choice of the gaseous substrate on products’ formation, the CO2 · H2 gas mixture was slightly preferable for the formation of ethanol. Indeed, the maximum ethanol titers obtained using CO2 · H2, CO · CO2 4:1 and syngas as gaseous substrates were 0.29 ± 0.01, 0.23 ± 0.00 and 0.20 ± 0.00 g/L, respectively (Figure 2B). The 2,3-BDO titer was the most sensitive to this choice. In fact, C. ljungdahlii produced 1.27 ± 0.02 g/L of 2,3-BDO when grown on CO · CO2 4:1, which is 2.8-fold higher than the titer attained when C. ljungdahlii was grown on syngas, whereas no 2,3-BDO production was detected when CO2 · H2 was employed (Figure 2C). Our results neatly indicate that the CO2 · H2 gas mixture is not appropriate for 2,3-BDO production, confirming previous observations [7,8], and point at CO · CO2 4:1 as the preferable gaseous substrate for 2,3-BDO autotrophic production. Notably, growing C. ljungdahlii on CO · CO2 4:1 allows an ethanol:2,3-BDO ratio of 1:6.03 (Table S4). The favorable 2,3-BDO titer obtained growing C. ljungdahlii on CO · CO2 4:1 depends on the energetic favorable reduction power of CO, which enhances the ATP yield of 2,3-BDO production, compared to H2 [10,36]. In autotrophically grown C. ljungdahlii, all the produced pyruvate, a key precursor for 2,3-BDO production, is derived from acetyl-CoA through a single step catalyzed by a pyruvate:ferredoxin oxidoreductase (PFOR). As acetyl-CoA is a C2 compound and pyruvate is a C3 compound, a molecule of CO2 needs to be incorporated. The energy for this reaction is provided by reduced ferredoxin [50]. The usage of the gas mixture, consisting of both CO and CO2, can favor 2,3-BDO production by increasing the levels of both CO2 and reduced ferredoxin, the reactants in the reaction catalyzed by the PFOR. Indeed, this gas mixture can directly provide CO2 and, owing to the presence of CO, can increase the level of reduced ferredoxin by CO oxidation via the ferredoxin-dependent carbon monoxide dehydrogenase (CODH). The 2,3-BDO production allows C. ljungdahlii to neutralize pyruvic acid whose excess could threaten the transmembrane proton gradient, which is required for ATP formation [50].
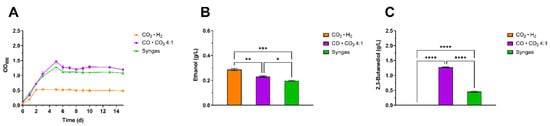
Figure 2.
Comparative analysis of growth and production profile of C. ljungdahlii using different gaseous substrates. The headspace of 250 mL serum bottles was filled with either CO2 · H2 (orange), CO · CO2 4:1 (violet) or syngas (CO, CO2, H2, N2 5:2:2:1) (green). (A) Growth profile characterization with the three gas mixtures. (B) Ethanol maximum titer, measured at day 15 for the three conditions tested. (C) 2,3-butanediol maximum titer, measured at day 5 and 3 for the CO2 · H2 and CO · CO2 4:1 conditions, respectively. The mean of three independent replicates is plotted for each tested condition. Error bars indicate the standard error of the mean (SEM). ANOVA p: ethanol (p = 0.0002); 2,3-BDO (p < 0.0001). Tukey’s test p: * p < 0.0332; ** p < 0.0021; *** p < 0.0002 and **** p < 0.0001.
As for C. ljungdahlii, growth and production specificities were assessed also in C. autoethanogenum grown on each gas mixture, using identical cultivation and sampling techniques to enable fair comparison of the outcomes (Figure 3 and Figure S3). We confirmed that CO · CO2 4:1 afforded the highest OD600nm, as shown in Figure 3A and, as a side result, that CO2 · H2 afforded the highest ethanol titer (Figure 3B). The highest 2,3-BDO titer was achieved by the CO · CO2 4:1 gas substrate, when compared to the other two gas substrates (Figure 3C). Indeed, the 2,3-BDO titer obtained using CO · CO2 4:1 (0.37 ± 0.03 g/L) was found statistically significantly higher than that obtained using syngas (0.20 ± 0.01 g/L), whereas CO2 · H2 led to no 2,3-BDO. Collectively, the outcomes of the gas substrates’ screening, conducted in full similarity using either C. ljungdahlii or C. autoethanogenum, allowed us to establish CO · CO2 4:1 as the preferable substrate for 2,3-BDO production.
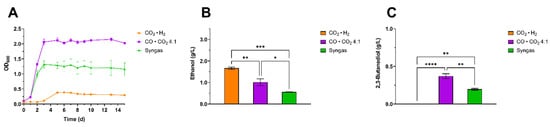
Figure 3.
Comparative analysis of growth and production profile of C. autoethanogenum using different gaseous substrates. The headspace of 250 mL serum bottles was filled with either CO2 · H2 (orange), CO · CO2 4:1 (violet) or syngas (CO, CO2, H2, N2 5:2:2:1) (green). (A) Growth profile characterization with the three gas mixtures. (B) Ethanol maximum titer, measured at day 8 for the CO2 · H2 condition and at day 15 for the CO · CO2 4:1 and syngas conditions. (C) 2,3-butanediol maximum titer, measured at day 3 and 5 for the CO · CO2 4:1 and syngas conditions, respectively. The mean of three independent replicates is plotted for each tested condition. Error bars indicate the standard error of the mean (SEM). ANOVA p: ethanol (p = 0.0006); 2,3-BDO (p < 0.0001). Tukey’s test p: * p < 0.0332; ** p < 0.0021; *** p < 0.0002 and **** p < 0.0001.
Upon identification of the preferable gas substrate for producing 2,3-BDO, we selected the appropriate acetogenic strain by comparing the maximal titer and productivity achieved by C. ljungdahlii and C. autoethanogenum grown on CO · CO2 4:1. This comparison revealed an interesting product specificity of these strains. C. autoethanogenum is preferable to C. ljungdahlii for ethanol production, by affording a 5.72-fold higher titer than C. ljungdahlii under the CO2 · H2 condition (Figure 4A). More importantly, we found that C. ljungdahlii is preferable to C. autoethanogenum for 2,3-BDO production. Indeed, growing C. ljungdahlii on CO · CO2 4:1 afforded a 3.43-fold higher titer compared to C. autoethanogenum (Figure 4B), grown on the same gas substrate. It is laterally noted that C. ljungdahlii was found to be preferential for 2,3-BDO production also when using syngas (Figure 4C). Moreover, the maximum 2,3-BDO productivity in C. ljungdahlii (0.024 ± 0.001 g/L/h) was 2.66-fold higher than that in C. autoethanogenum (0.009 ± 0.001 g/L/h), when CO · CO2 4:1 was used as gas substrate (Table S3). According to these findings, we set out to select C. ljungdahlii and CO · CO2 4:1 as the preferable strain and gas substrate to be used in the subsequent medium screening study to enhance 2,3-BDO production.
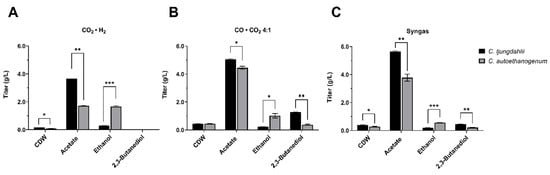
Figure 4.
Comparative analysis of growth and production profiles of C. ljungdahlii (black) and C. autoethanogenum (grey) using different gaseous substrates. The headspace of 250 mL serum bottles was filled with either (A) CO2 · H2, (B) CO · CO2 4:1 or (C) syngas (CO, CO2, H2, N2 5:2:2:1). Maximum values of cell dry weight (CDW), acetate, ethanol and 2,3-butanediol are reported. The mean of three independent replicates for each gaseous substrate is plotted. Error bars indicate the standard error of the mean (SEM). For each gaseous substrate tested, shown are the single, paired one-tailed t-tests, corresponding to statistically significant differences of the observable (CDW, titer of each product) between C. ljungdahlii and C. autoethanogenum (* p < 0.0332; ** p < 0.0021; *** p < 0.0002).
3.3. Influence of Cultivation Medium on 2,3-BDO Production from CO · CO2 using Clostridium ljungdahlii
In order to determine the preferable medium composition for 2,3-BDO production, we screened out three additional media featuring different concentrations of trace metals, mineral elements and vitamins, as shown in Table 2.
Table 2.
Summary of the final concentration of the main elements in the media tested in this study. Final concentrations of mineral elements (mM), trace elements (metal) (µM), and vitamins (µM) are reported in the table.
Although previous studies have explored medium definition to support ethanol production [37,40,41,51], the literature lacks a systematic characterization of the medium composition favoring 2,3-BDO acetogenic production. In full similarity to the screening on gaseous substrates, to enable an appropriate comparison of the results, we carried out the media screening by using the same inoculation, cultivation condition, sampling technique, gas substrate (CO · CO2 4:1) and biocatalyst (C. ljungdahlii), previously selected.
In addition to the “Tanner_mod” medium, our screening included three alternative media that are referred to as “Valgepea_mod”, “Tan_Val” and “Tan_Val + Fe” hereafter and that are detailed in the Materials and Methods section. The “Valgepea_mod” medium, slightly modified from Valgepea and co-workers [13], differs from the “Tanner_mod” one for the concentrations of mineral elements, trace metals, vitamins and Iron, as shown in Table 2. The “Tan_Val” medium has been conceived in this work and foresees the mineral elements of the “Tanner_mod” medium and the trace metals and vitamins of the “Valgepea-mod” medium. Subsequently, in order to investigate the Iron influence on 2,3-BDO production, we formulated the “Tan_Val + Fe” medium that is identical to the “Tan_Val” one but foresees the same Iron concentration of the “Valgepea_mod” medium–86.89 µM (Table 2). Yeast extract (YE) was removed from the formulation of all the media different from the “Tanner_mod” one, which included 0.5 g/L of YE, in the drive to reduce the costs, to favor the formation of non-growth-coupled products and to disambiguate the products’ profile interpretation [40,41,52].
Growth of C. ljungdahlii on CO · CO2 4:1 using the “Tanner_mod” medium did not show the lag phase, compared to other conditions, probably due to the presence of the optimal YE concentration [41,53]. The “Tan_Val” medium afforded a maximum OD600nm (1.56 ± 0.04 at day 6) similar to the one reached using the “Tanner_mod” medium (1.47 ± 0.02 at day 5) and led to the highest specific growth rate (0.0346 ± 0.0007 gCDW/L/h, Table S5), whereas growth on the “Valgepea_mod” and on the “Tan_Val + Fe” media, led to lower maximum OD600nm values (1.10 ± 0.13 at day 7 and 1.33 ± 0.04 at day 10, respectively), as shown in Figure 5A. We attributed the negative effect of the latter media on growth to the higher Iron concentration, which distinguishes the “Valgepea_mod” and “Tan_Val + Fe” media from the “Tanner_mod” and “Tan_Val” ones. In fact, the “Tan_Val” and the “Tanner_mod” media have similar Iron concentration (23.99 µM and 20.40 µM, respectively), while the “Valgepea_mod” and “Tan_Val + Fe” media are characterized by the same Iron concentration (86.89 µM) which is 4.25-fold higher than that of the “Tanner_mod” medium and 3.62-fold higher than that of the “Tan_Val” medium (Table 2).
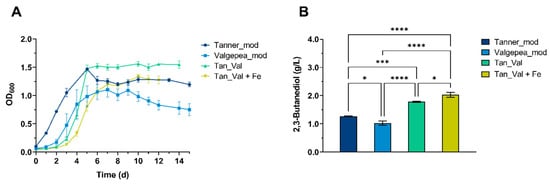
Figure 5.
Comparative analysis of growth and 2.3-butanediol production of C. ljungdahlii grown in CO · CO2 4:1 and in different media. “Tanner_mod” (Yale blue); “Valgepea_mod” (Munsell blue); “Tan_Val” (Malachite green) and “Tan_Val + Fe” (Chartreuse yellow) media were employed during 250 mL serum bottles batch experiments. (A) Growth profile with the four cultivation media. (B) Maximum 2,3-butanediol titer, measured at day 5, 10, 5 and 10 for the “Tanner_mod”, “Valgepea_mod”, “Tan_Val” and “Tan_val + Fe” conditions, respectively. The mean of three independent replicates is plotted for each cultivation medium. Error bars indicate the standard error of the mean (SEM). ANOVA P: 2,3-BDO (p < 0.0001). Tukey’s test p: * p < 0.0332; *** p < 0.0002 and **** p < 0.0001.
Medium formulation drastically influenced 2,3-BDO titer and productivity. In fact, 1.27 ± 0.02 g/L, 1.02 ± 0.08 g/L, 1.78 ± 0.03 g/L and 2.03 ± 0.05 g/L of 2,3-BDO were obtained by using the “Tanner_mod”, “Valgepea_mod”, “Tan_Val” and “Tan_Val + Fe” media after 5, 10, 5 and 10 days from the beginning of the test, respectively (Figure 5B). Whereas 2,3-BDO titers in the “Valgepea_mod” and “Tanner_mod” media were similar, 2,3-BDO productivity in the “Valgepea_mod” medium (0.012 ± 0.005 g/L/h) was 2-fold lower than using the “Tanner_mod” medium (0.024 ± 0.001 g/L/h) (Table S5). In addition to different mineral elements’ concentrations, the “Valgepea_mod’’ medium contains 200-, 1.5-, 1.5- and 4.25-fold higher concentrations of Zinc, Molybdenum, Selenium and Iron, respectively, compared to the “Tanner_mod‘‘ medium (Table 2). A higher concentration of Zinc, Molybdenum, Selenium and Iron has been previously reported to increase cell growth and ethanol production in C. ragsdalei grown on CO · CO2 [51]. Although a similar positive effect was not observed in our study on 2,3-BDO productivity and titer, our results do not rule out the possibility that increasing trace metals in the growth medium could be beneficial also to the 2,3-BDO production since the absolute amounts and relative increase of the trace metals in our study are neatly lower than in Saxena and Tanner [51]. Metal elements are essential to support the activity of metalloenzymes in the Wood-Ljungdahl pathway [54,55]. Moreover, the concentration of most constituents of the vitamin solution of the “Valgepea_mod” medium was increased by 10-fold, with the exception of vitamin B12, which was barely detectable in the “Tanner_mod” medium and was augmented by around 500-fold in the “Valgepea_mod” medium (Table 2). Although some studies did not provide evidence that manipulating vitamin concentrations could affect either the growth or the production profile of C. autoethanogenum [40,53], it was shown that increasing the concentration of specific vitamins, such as pantothenic acid and biotin, above the cellular requirements of an acetogen grown on a CO-rich gas mixture led to an increase in 2,3-BDO production [56]. Furthermore, a role in anaerobic CO2 fixation was described for vitamin B12, a cofactor for methyl group transfer reactions in the Wood-Ljungdahl pathway [57], whose concentration was substantially increased in the “Valgepea_mod” medium. Therefore, we speculated that it was the mineral elements defined in the “Valgepea_mod” medium (Table 2) which could exert a negative effect on 2,3-BDO production, despite the higher Zinc concentration present in the “Valgepea_mod” medium, compared with the “Tanner_mod” one.
Therefore, we tested the “Tan_Val” medium, which combines the trace metals’ and vitamins’ formulation of the “Valgepea_mod” medium with the mineral elements’ concentration used in the “Tanner_mod” medium (Table 2). We managed to enhance both 2,3-BDO titer (1.78 ± 0.03 g/L) and 2,3-BDO productivity (0.038 ± 0.005 g/L/h), which were found 1.4- and 1.58-fold higher, respectively, when compared to the “Tanner_mod” medium (Figure 5B, Table S5). The improvement, obtained using the “Tan_Val” medium, could be attributable to the effects exerted, in particular, by the combination of increased concentrations of trace metals and vitamins—derived from the “Valgepea_mod” medium—with the mineral element concentrations derived from the “Tanner_mod” medium. Zinc was one of the trace metals whose concentration mostly changed between the “Tanner_mod” and “Tan_Val” media with a 200-fold increase (Table 2). The positive effect observed could be plausible due to the fact that Zinc is a fundamental metal for acetogenic alcohol dehydrogenase enzymes [58]. In particular, the butanediol dehydrogenase (CLJU_c23220), which is the enzyme responsible for the conversion of acetoin into 2,3-BDO, is a Zinc-dependent enzyme [26]. The beneficial effect of replacing the “Valgepea_mod” mineral solution with the “Tanner_mod” one was confirmed when we compared the 2,3-BDO production obtained using the “Valgepea_mod” and “Tan_Val” media, which differ only for the mineral elements’ and Iron concentrations (Table 2). Here, the different concentration of mineral elements present in the “Tan_Val” medium, relative to the “Valgepea_mod” one, corresponded to a 1.74-fold increase in 2,3-BDO titer and to a 3.16-fold increase in 2,3-BDO productivity (Figure 5B).
Finally, we tested 2,3-BDO production in the “Tan_Val + Fe” medium which led to 2.03 ± 0.05 g/L of 2,3-BDO. The titer achieved here is unprecedented on the basis of the titers recorded in previous batch experiments on serum bottles level. Indeed, the highest titer by a pure culture was of 1 g/L but was obtained by growing C. autoethanogenum culture on syngas supplemented with fructose [14]. The titer obtained in our study was found to be similar to the titer previously obtained by a syntrophic cultivation of C. acetobutylicum and C. ljungdahlii which was equal to 1.98 g/L, but in this case, glucose was used as an additional carbon source [59].
The “Tan_Val + Fe” medium differs only for the concentration of mineral elements from the “Valgepea_mod” medium and differs only for the Iron concentration from the “Tan_Val” medium (Table 2). By comparing 2,3-BDO production titers obtained in the “Tan_Val + Fe” and “Valgepea_mod” media, we confirmed the positive influence of the mineral elements on 2,3-BDO production (Figure 5B). Moreover, by comparing 2,3-BDO production titers obtained in the “Tan_Val” and “Tan_Val + Fe” media, we found that increasing Iron concentration could have a slightly positive effect on 2,3-BDO production (Figure 5B). In fact, Iron serves as essential metal cofactor for the Iron-Sulphur proteins, which are related to the electron transfer pathway [18], the methyl-branch [60] and carbonyl-branch [55,61] of the Wood-Ljungdahl pathway. Moreover, Iron was found to be a limiting element for acetogens’ growth and productivity [41,62] and also an important coenzyme of alcohol dehydrogenase [63].
The results summarized as follows let us to conclude that trace elements and vitamins generally have only a minor effect on 2,3-BDO production while mineral elements, Zinc and Iron exert a major positive influence on 2,3-BDO titer and productivity. The higher titer and productivity achieved by the “Tan_Val + Fe” compared to the “Valgepea_mod” medium depends on the replacement of the “Valgepea_mod” mineral element concentrations with the “Tanner_mod” ones. The “Tanner_mod” mineral solution features 9.23- and 2-fold lower PO4 and KCl concentrations and 9.58-fold higher NaCl compared to the “Valgepea_mod” medium whereas Ca concentrations were similar. The higher titer achieved by the “Tan_Val + Fe” medium compared to the “Tan_Val” medium is due to the increased Fe supplementation. Finally, the similar titers of “Valgepea_mod” and “Tanner_mod” together with the lower productivity of the “Valgepea_mod” medium compared to the “Tanner_mod” one let us think that positive effect of increased Zn in the “Valgepea_mod” medium was reduced by the “Valgepea_mod” mineral element concentrations, which was found to be the only difference responsible of the lower performance of the “Valgepea_mod” medium with respect to the “Tan_Val + Fe” one. It is noted that the “Valgepea_mod” medium was specifically formulated to boost ethanol production of C. autoethanogenum using syngas [13], while we tested its influence on 2,3-BDO production in C. ljungdahlii grown on CO · CO2. Taken together, our results highlight the importance of undertaking studies on medium composition influence on 2,3-BDO production, as those already performed by Saxena and Tanner [51] and by Gao and co-workers [41] for ethanol production using C. ragsdalei grown on CO · CO2.
3.4. Characterization of the Effects of Different CO to CO2 Ratios on 2,3-BDO Production by Clostridium ljungdahlii
We investigated the growth and metabolites’ profile of C. ljungdahlii grown in the “ Tan_Val” medium and by using four CO · CO2 gas mixtures, differing in the CO to CO2 gas ratios: pure CO, CO · CO2 4:1 (80% CO and 20% CO2), CO · CO2 1:1 (50% CO and 50% CO2) and CO · CO2 1:4 (20% CO and 80% CO2) (Figure 6 and Figure S5, Tables S7 and S8).
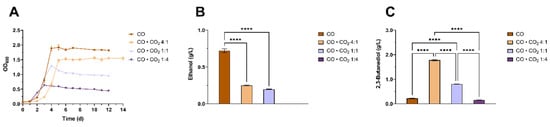
Figure 6.
Comparative analysis of growth and production profile of C. ljungdahlii using different CO:CO2 gas ratios. The headspace of 250 mL serum bottles was filled with either pure CO (brown) CO · CO2 4:1 (Sandy tan), CO · CO2 1:1 (Lavender blue) or CO · CO2 1:4 (dark purple). (A) Growth profiles with the four gas mixtures. (B) Ethanol maximum titer, measured at day 10, 14 and 4 for the pure CO, CO · CO2 4:1 and CO · CO2 1:1 conditions, respectively. (C) 2,3-butanediol maximum titer, measured at day 6, 5, 4 and 3 for the pure CO, CO · CO2 4:1, CO · CO2 1:1 and CO · CO2 1:4 conditions, respectively. The mean of three independent replicates is plotted for each tested condition. Error bars indicate the standard error of the mean (SEM). ANOVA P: ethanol (p < 0.0001); 2,3-BDO (p < 0.0001). Tukey’s test p: **** p < 0.0001.
C. ljungdahlii growth on pure CO gas substrate outperformed that on CO · CO2. In fact, C. ljungdahlii reached a maximal OD600nm of 1.92 ± 0.08, 1.56 ± 0.04, 1.30 ± 0.02 and 0.63 ± 0.00 using pure CO, CO · CO2 4:1; CO · CO2 1:1 and CO · CO2 1:4, respectively (Figure 6A). As expected, by reducing the amount of CO in the gaseous substrate, biomass and acetate, which is a growth-related metabolite, decreased (Figure 6 and Figure S5). Ethanol titers were negatively affected by increasing the percentage of CO2 at the expense of the CO present in the gaseous substrate. By comparing 2,3-BDO titers obtained in the pure CO and CO · CO2 4:1 conditions, it is interesting to note that the presence of CO2 positively influenced 2,3-BDO production at the expense of ethanol production. In fact, growing C. ljungdahlii on pure CO produced only 0.22 ± 0.01 g/L of 2,3-BDO whereas the 2,3-BDO titer increased up to 1.78 ± 0.03 g/L when C. ljungdahlii was grown on CO · CO2 4:1 (Figure 6B,C). The utilization of a gas mixture consisting of both CO and CO2 can favor 2,3-BDO production since it directly increases the level of CO2 and indirectly, via the CODH-catalyzed reaction, the levels of reduced ferredoxin, the two main reactants of the PFOR reaction [50]. Nevertheless, further increasing the amount of CO2 with respect to CO was revealed to be counterproductive. Indeed, 0.80 ± 0.01 g/L and 0.16 ± 0.00 g/L of 2,3-BDO were obtained on CO · CO2 1:1 and CO · CO2 1:4, respectively, corroborating the indication that the CO presence is a limiting element for 2,3-BDO production (Figure 6C). Indeed, providing sufficient levels of CO allows 2,3-BDO to act as an electron acceptor to relieve the microbial cell of excess of reducing power, in the form of NAD(P)H, thus restoring a favorable NAD(P):NAD(P)H equilibrium. In fact, it has been previously shown that where CO is supplied at sufficient levels, 2,3-BDO dehydrogenase expression is upregulated [64].
4. Conclusions
Our study presented a multifaceted screening campaign which, impinging on the selection of a suitable catalyst, gaseous substrate and medium composition, significantly enhanced 2,3-BDO production in batch experiments using serum bottles. The preferable configuration identified C. ljungdahlii as biocatalyst and CO · CO2 4:1 as gas substrate. Our medium screening unveiled that the “Tan_Val” medium outperformed the remaining ones with respect to 2,3-BDO productivity (0.038 ± 0.005 g/L/h) and that the “Tan_Val + Fe” medium ensured the highest 2,3-BDO titer (2.03 ± 0.05 g/L). We identified in the co-presence of optimized mineral elements, Zinc and Iron the major component to boost 2,3-BDO titer and productivity. Moreover, the screening of various CO · CO2 gaseous substrates, characterized by different gas ratios, revealed that CO2 addition to CO fermentation of C. ljungdahlii caused a metabolic shift from ethanol to 2,3-BDO production and that CO2 addition over the 20% drastically reduced 2,3-BDO titer. Key to the achievement of enhanced 2,3-BDO titer was the adoption of a rigorous comparative framework preceded by the selection of an improved cultivation configuration (horizontally incubated serum bottles with a gas to liquid ratio of 9:1).
The 2,3-BDO titer of 2.03 ± 0.05 g/L, which was achieved in this work by growing C. ljungdahlii on CO · CO2 4:1 and the “Tan_Val + Fe” medium, is superior to any 2,3-BDO titer previously observed in batch fermentation experiments carried out in serum bottles [26,32,36,37]. Similar or higher titers have been previously achieved only in batch [65] or fed-batch fermentation experiments carried out in reactor environments that ensure superior process control [7,8,9]. A similar 2,3-BDO titer was indeed obtained in a fed-batch fermentation in which a CO · CO2 gas substrate was constantly supplied for 276 h to a C. ljungdahlii culture hosted in a bioreactor (working volume, 2.5 L) where gas pressure and pH were constantly controlled at 1 bar and 6.0, respectively [9]. 2,3-BDO titers substantially higher than the value attained in the present study were observed in Zhu and co-workers [7]. In this case, C. ljungdahlii was grown in CO · CO2 in the presence of yeast extract in a bioreactor (working volume, 2.5 L) with pH and supplied gas pressure controlled at 6 and 1 bar, respectively, and the 2,3-BDO titer increased up to 16.94 ± 0.36 g/L.
Future development will involve further medium optimization by evaluating, for instance, the effects of supplementing specific amino acids to the growth medium on 2,3-BDO production performances [37]. However, we believe that a substantial turnaround in the development of the 2,3-BDO production process will be the optimization of the gaseous conversion into 2,3-BDO in fermentations carried out in gas and liquid continuous configuration. In this case, further medium and gas substrates’ optimization could be profitably combined with the optimization of equally relevant fermentation aspects, such as the pH control. Indeed, the pH of the broth affects the energy conservation system which relies on the establishment of a proton-dependent transmembrane ion gradient through the ATPase/Rnf system [7]. In fact, the highest titers of 2,3-BDO, present in literature, were obtained when a constant pH control was applied [7,9]. Furthermore, it is useful to study the effects of different pH profiles on the 2,3-BDO formation. For instance, cyclic pH shifts between a high level of 5.75 and a lower value of 4.75 were carried out in a bioreactor, continuously fed with carbon monoxide, for continuous ethanol and 2,3-BDO production, resulting in an overall concentration of 1.62 g/L of 2,3-BDO [66].
A yet further parameter that could be controlled, in experiments performed using bioreactors, is the flow rate with which the gas substrate is supplied. Indeed, a C. autoethanogenum fermentation carried out in continuous stirred tank reactor prepared at pH 5.5 has previously shown that 2,3-BDO productivity increased significantly from < 0.1 g/L/day to 1.2 g/L/day when the gas flow rate was increased in a such way that specific CO uptake increased from 0.3 to 0.6 mmol/g/min [64]. Finally, foreseeing cell recycling in the configuration of a continuous process for 2,3-BDO production could be beneficial. Aside from the possibility of achieving high cell density cultivation, cell retention could favor 2,3-BDO production since the age of the microbial cells and the growth phase of the microbial culture were found to have an effect on the amount of 2,3-BDO produced. Indeed, 2,3-BDO productivity and 2,3-BDO:ethanol ratio was found to increase with increased average cellular age [23].
In summary, our study enabled the generation of a notable dataset providing a potential guide for continuous gas fermentation processes development for 2,3-BDO production from CO · CO2 fermentation by C. ljungdahlii.
Supplementary Materials
The following are available online at www.mdpi.com/article/10.3390/fermentation7040264/s1, Figure S1: Comparative analysis of acetate production by C. ljungdahlii cultivated in CO · CO2 4:1 and using different cultivation techniques, Figure S2: Comparative analysis of acetate production by C. ljungdahlii using different gaseous substrates, Figure S3: Comparative analysis of acetate production by C. autoethanogenum using different gaseous substrates, Figure S4: Comparative analysis of acetate and ethanol production of C. ljungdahlii grown in CO · CO2 4:1 and different media, Figure S5: Comparative analysis of acetate production of C. ljungdahlii using different CO:CO2 gas ratios, Table S1: Specific growth rate, acetate, ethanol and 2,3-BDO productivities of C. ljungdahlii grown in CO · CO2 4:1 using three different cultivation conditions, Table S2: Product ratios of C. ljungdahlii grown in CO · CO2 4:1 using three different cultivation conditions, Table S3: Specific growth rate, acetate, ethanol and 2,3-BDO productivities of C. ljungdahlii and C. autoethanogenum grown using three different gaseous substrates, Table S4: Product ratios of C. ljungdahlii and C. autoethanogenum grown using three different gaseous substrates, Table S5: Specific growth rate, acetate, ethanol and 2,3-BDO productivities of C. ljungdahlii grown using four different cultivation media, Table S6: Product ratios of C. ljungdahlii grown using four different cultivation media, Table S7: Specific growth rate, acetate, ethanol and 2,3-BDO productivities of C. ljungdahlii grown using four different CO-based gaseous substrates, Table S8: Product ratios of C. ljungdahlii grown using four different CO-based gaseous substrates.
Author Contributions
Conceptualization, L.R. and V.A.; methodology, L.R.; software, L.R.; validation, L.R. and A.R.; formal analysis, L.R. and A.R.; investigation, L.R. and A.R.; resources, V.A. and A.R.; data curation, L.R.; writing—original draft preparation, L.R. and A.R.; writing—review and editing, L.R., V.A., D.F. and A.R.; visualization, L.R. and A.R.; supervision, A.R. and D.F.; project administration, L.R. and A.R.; funding acquisition, A.R. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the program P.O.R FESR 2014/2020—Action I.1b.2.2-Bioeconomy Technological Platform—under Grant 333-30 (project PRIME “Processi e pRodotti Innovativi di Chimica vErde”).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained with the article and Supplementary Materials.
Acknowledgments
We thank the master thesis student Giacomo Antonicelli for the useful help provided during the investigation and data acquisition processes.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Liew, F.M.; Martin, M.E.; Tappel, R.C.; Heijstra, B.D.; Mihalcea, C.; Köpke, M. Gas fermentation-A flexible platform for commercial scale production of low-carbon-fuels and chemicals from waste and renewable feedstocks. Front. Microbiol. 2016, 7, 694. [Google Scholar] [CrossRef]
- Köpke, M.; Simpson, S. Pollution to products: Recycling of ‘above ground’ carbon by gas fermentation. Curr. Opin. Biotechnol. 2020, 65, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Takors, R.; Kopf, M.; Mampel, J.; Bluemke, W.; Blombach, B.; Eikmanns, B.; Bengelsdorf, F.R.; Weuster-Botz, D.; Dürre, P. Using gas mixtures of CO, CO2 and H2 as microbial substrates: The do’s and don’ts of successful technology transfer from laboratory to production scale. Microb. Biotechnol. 2018, 11, 606–625. [Google Scholar] [CrossRef] [PubMed]
- Bae, J.; Song, Y.; Lee, H.; Shin, J.; Jin, S.; Kang, S.; Cho, B.K. Valorization of C1 gases to value-added chemicals using acetogenic biocatalysts. Chem. Eng. J. 2022, 428, 131325. [Google Scholar] [CrossRef]
- European Commission. Available online: https://ec.europa.eu/info/strategy/priorities-2019-2024/european-green-deal_en (accessed on 14 October 2021).
- Infantes, A.; Kugel, M.; Raffelt, K.; Neumann, A. Side-by-side comparison of clean and as substrate for acetogenic fermentation with Clostridium ljungdahlii. Fermentation 2020, 6, 84. [Google Scholar] [CrossRef]
- Zhu, H.F.; Liu, Z.Y.; Zhou, X.; Yi, J.H.; Lun, Z.M.; Wang, S.N.; Tang, W.Z.; Li, F.L. Energy conservation and carbon flux distribution during fermentation of CO or H2/CO2 by Clostridium ljungdahlii. Front. Microbiol. 2020, 11, 416. [Google Scholar] [CrossRef]
- Hermann, M.; Teleki, A.; Weitz, S.; Niess, A.; Freund, A.; Bengelsdorf, F.R.; Takors, R. Electron availability in CO2, CO and H2 mixtures constrains flux distribution, energy management and product formation in Clostridium ljungdahlii. Microb. Biotechnol. 2020, 13, 1831–1846. [Google Scholar] [CrossRef]
- Liu, Z.Y.; Jia, D.C.; Zhang, K.D.; Zhu, H.F.; Zhang, Q.; Jiang, W.H.; Gu, Y.; Li, F.L. Ethanol metabolism dynamics in Clostridium ljungdahlii grown on carbon monoxide. Appl. Environ. Microbiol. 2020, 86, 1–14. [Google Scholar] [CrossRef]
- Katsyv, A.; Müller, V. Overcoming energetic barriers in acetogenic C1 conversion. Front. Bioeng. Biotechnol. 2020, 8, 621166. [Google Scholar] [CrossRef]
- Mock, J.; Zheng, Y.; Mueller, A.P.; Ly, S.; Tran, L.; Segovia, S.; Nagaraju, S.; Köpke, M.; Dürre, P.; Thauer, R.K. Energy conservation associated with ethanol formation from H2 and CO2 in Clostridium autoethanogenum involving electron bifurcation. J. Bacteriol. 2015, 197, 2965–2980. [Google Scholar] [CrossRef]
- Heffernan, J.K.; Valgepea, K.; de Souza Pinto Lemgruber, R.; Casini, I.; Plan, M.; Tappel, R.; Simpson, S.; Köpke, M.; Nielsen, L.K.; Marcellin, E. Enhancing CO2-valorization using clostridium autoethanogenum for sustainable fuel and chemicals production. Front. Bioeng. Biotechnol. 2020, 8, 204. [Google Scholar] [CrossRef] [PubMed]
- Valgepea, K.; de Souza Pinto Lemgruber, R.; Meaghan, K.; Palfreyman, R.W.; Abdalla, T.; Heijstra, B.D.; Behrendorff, J.B.; Marcellin, E. Maintenance of ATP homeostasis triggers metabolic shifts in gas-fermenting acetogens. Cell Syst. 2017, 4, 505–515. [Google Scholar] [CrossRef] [PubMed]
- Marcellin, E.; Behrendorff, J.B.; Nagaraju, S.; DeTissera, S.; Segovia, S.; Palfreyman, R.W.; Daniell, J.; Licona-Cassani, C.; Quek, L.; Speight, R.; et al. Low carbon fuels and commodity chemicals from waste gases-systematic approach to understand energy metabolism in a model acetogen. Green Chem. 2016, 18, 3020–3028. [Google Scholar] [CrossRef]
- Oswald, F.; Zwick, M.; Omar, O.; Hotz, E.N.; Neumann, A. Growth and product formation of Clostridium ljungdahlii in presence of cyanide. Front. Microbiol. 2018, 9, 1213. [Google Scholar] [CrossRef]
- Rückel, A.; Hannemann, J.; Maierhofer, C.; Fuchs, A.; Weuster-Botz, D. Studies on syngas fermentation with Clostridium carboxidivorans in stirred-tank reactors with defined gas impurities. Front. Microbiol. 2021, 12, 655390. [Google Scholar] [CrossRef]
- Stoll, I.K.; Boukis, N.; Sauer, J. Syngas fermentation to alcohols: Reactor technology and application perspective. Chem. Ing. Tech. 2020, 92, 125–136. [Google Scholar] [CrossRef]
- Schuchmann, K.; Müller, V. Autotrophy at the thermodynamic limit of life: A model for energy conservation in acetogenic bacteria. Nat. Rev. Microbiol. 2014, 12, 809–821. [Google Scholar] [CrossRef]
- Bengelsdorf, F.R.; Straub, M.; Dürre, P. Bacterial synthesis gas (syngas) fermentation. Environ. Technol. 2013, 34, 1639–1651. [Google Scholar] [CrossRef]
- ArcelorMittal. Available online: https://belgium.arcelormittal.com/en/arcelormittal-and-lanzatech-break-ground-on-e150million-project-to-.revolutionise-blast-furnace-carbon-emissions-capture/ (accessed on 14 October 2021).
- Lanzatech 2020. Available online: https://www.lanzatech.com/2020/09/15/advanced-biofuel-facility-gets-green-light-in-india/ (accessed on 14 October 2021).
- Sekisui Chemical 2020. Available online: https://www.sekisuichemical.com/news/2020/1364066_38530.html (accessed on 14 October 2021).
- Simpson, S.; Collet, C.; Mihalcea, C.D.; Conolly, J.J.; Waters, G.W. Fermentation Process for Controlling Butanediol Production. U.S. Patent No. US 2012/0252082 A1, 4 October 2012. [Google Scholar]
- Song, C.W.; Park, J.M.; Chung, S.C.; Lee, S.Y.; Song, H. Microbial production of 2,3-butanediol for industrial applications. J. Ind. Microbiol. Biotechnol. 2019, 46, 1583–1601. [Google Scholar] [CrossRef]
- Paciorek-Sadowska, J.; Czupryn, B. New compounds for production of polyurethane foams. J. Appl. Polym. Sci. 2006, 102, 5918–5926. [Google Scholar] [CrossRef]
- Köpke, M.; Mihalcea, C.; Liew, F.M.; Tizard, J.H.; Ali, M.S.; Conolly, J.J.; Al-Sinawi, B.; Simpson, S. 2,3-butanediol production by acetogenic bacteria, an alternative route to chemical synthesis, using industrial waste gas. Appl. Environ. Microbiol. 2011, 77, 5467–5475. [Google Scholar] [CrossRef]
- Bengelsdorf, F.R.; Poehlein, A.; Linder, S.; Erz, C.; Hummel, T.; Hoffmeister, S.; Daniel, R.; Dürre, P. Industrial acetogenic biocatalysts: A comparative metabolic and genomic analysis. Front. Microbiol. 2016, 7, 1036. [Google Scholar] [CrossRef] [PubMed]
- Tanner, R.S.; Miller, L.M.; Yang, D. Clostridium ljungdahlii Sp-Nov, an acetogenic species in clostridial ribosomal-RNA homology group-I. Int. J. Syst. Bacteriol. 1993, 43, 232–236. [Google Scholar] [CrossRef] [PubMed]
- Köpke, M.; Held, C.; Hujer, S.; Liesegang, H.; Wiezer, A.; Wollherr, A.; Ehrenreich, A.; Liebl, W.; Gottschalk, G.; Durre, P. Clostridium ljungdahlii represents a microbial production platform based on syngas. Proc. Natl. Acad. Sci. USA 2010, 107, 13087–13092. [Google Scholar] [CrossRef] [PubMed]
- Abrini, J.; Naveau, H.; Nyns, E.J. Clostridium autoethanogenum, Sp. Nov., an anaerobic bacterium that produces ethanol from carbon monoxide. Arch. Microbiol. 1994, 161, 345–351. [Google Scholar] [CrossRef]
- Brown, S.D.; Nagaraju, S.; Utturkar, S.; De Tissera, S.; Segovia, S.; Mitchell, W.; Land, M.L.; Dassanayake, A.; Köpke, M. Comparison of single-molecule sequencing and hybrid approaches for finishing the genome of clostridium autoethanogenum and analysis of CRISPR systems in industrial relevant clostridia. Biotechnol. Biofuels 2014, 7, 40. [Google Scholar] [CrossRef] [PubMed]
- Liew, F.; Henstra, A.M.; Winzer, K.; Köpke, M.; Simpson, S.D.; Minton, N.P. Insights into CO2 fixation pathway of Clostridium autoethanogenum by targeted mutagenesis. MBio 2016, 7, e00427-16. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Liang, C.; Yuan, Z.; Xu, J.; Hua, Q.; Guo, Y. A study of CO/Syngas bioconversion by Clostridium autoethanogenum with a flexible gas-cultivation system. Enzyme Microb. Technol. 2017, 101, 24–29. [Google Scholar] [CrossRef]
- Liew, F.; Henstra, A.M.; Köpke, M.; Winzer, K.; Simpson, S.D.; Minton, N.P. Metabolic engineering of Clostridium autoethanogenum for selective alcohol production. Metab. Eng. 2017, 40, 104–114. [Google Scholar] [CrossRef]
- Valgepea, K.; De Souza Pinto Lemgruber, R.; Abdalla, T.; Binos, S.; Takemori, N.; Takemori, A.; Tanaka, Y.; Marcellin, E. H2 drives metabolic rearrangements in gas-fermenting Clostridium autoethanogenum. Biotechnol. Biofuels 2018, 11, 55. [Google Scholar] [CrossRef]
- Jack, J.; Lo, J.; Maness, P.-C.; Ren, Z.J. Directing Clostridium ljungdahlii fermentation products via hydrogen to carbon monoxide ratio in syngas. Biomass Bioenerg. 2019, 124, 95–101. [Google Scholar] [CrossRef]
- Lee, J.; Lee, J.W.; Chae, C.G.; Kwon, S.J.; Kim, Y.J.; Lee, J.H.; Lee, H.S. Domestication of the novel alcohologenic acetogen Clostridium sp. AWRP: From isolation to characterization for syngas fermentation. Biotechnol. Biofuels 2019, 12, 228. [Google Scholar] [CrossRef] [PubMed]
- Martin, M.E.; Richter, H.; Saha, S.; Angenent, L.T. Traits of selected clostridium strains for syngas fermentation to ethanol. Biotechnol. Bioeng. 2016, 113, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Cotter, J.L.; Chinn, M.S.; Grunden, A.M. Influence of process parameters on growth of Clostridium ljungdahlii and Clostridium autoethanogenum on synthesis gas. Enzyme Microb. Technol. 2009, 44, 281–288. [Google Scholar] [CrossRef]
- Abubackar, H.N.; Veiga, M.C.; Kennes, C. Carbon monoxide fermentation to ethanol by Clostridium autoethanogenum in a bioreactor with no accumulation of acetic acid. Bioresour. Technol. 2015, 186, 122–127. [Google Scholar] [CrossRef]
- Gao, J.; Atiyeh, H.K.; Phillips, J.R.; Wilkins, M.R.; Huhnke, R.L. Development of low cost medium for ethanol production from syngas by Clostridium ragsdalei. Bioresour. Technol. 2013, 147, 508–515. [Google Scholar] [CrossRef]
- Tanner, R.S. Cultivation of bacteria and fungi. In Manual of Environmental Microbiology; Hurst, C.J., Crawford, R.L., Garland, J.L., Lipson, D.A., Mills, A.L., Stetzenbach, L.D., Eds.; ASM Press: Washington, DC, USA, 2007; pp. 69–78. [Google Scholar]
- Infantes, A.; Kugel, M.; Neumann, A. evaluation of media components and process parameters in a sensitive and robust fed-batch syngas fermentation system with Clostridium ljungdahlii. Fermentation 2020, 6, 61. [Google Scholar] [CrossRef]
- Yasin, M.; Jeong, Y.; Park, S.; Jeong, J.; Lee, E.Y.; Lovitt, R.W.; Kim, B.H.; Lee, J.; Chang, I.S. Microbial synthesis gas utilization and ways to resolve kinetic and mass-transfer limitations. Bioresour. Technol. 2015, 177, 361–374. [Google Scholar] [CrossRef]
- Ribeiro, R.R.; Coelho, F.; Ferreira, T.F.; Amaral, P.F.F. A new strategy for acetogenic bacteria cell growth and metabolites production using syngas in lab scale. IOSR J. Biotechnol. Biochem. 2017, 3, 27–30. [Google Scholar] [CrossRef]
- Phillips, J.R.; Atiyeh, H.K.; Tanner, R.S.; Torres, J.R.; Saxena, J.; Wilkins, M.R.; Huhnke, R.L. Butanol and hexanol production in Clostridium carboxidivorans syngas fermentation: Medium development and culture techniques. Bioresour. Technol. 2015, 190, 114–121. [Google Scholar] [CrossRef]
- Norman, R.; Millat, T.; Schatschneider, S.; Henstra, A.M.; Breitkopf, R.; Pander, B.; Annan, F.; Piatek, P.; Hartman, H.B.; Hodgman, C. A genome-scale model of Clostridium autoethanogenum reveals optimal bioprocess conditions for high-value commodity chemical production from carbon monoxide. Eng. Biol. 2019, 3, 32–40. [Google Scholar] [CrossRef]
- Bertsch, J.; Müller, V. Bioenergetic constraints for conversion of syngas to biofuels in acetogenic bacteria. Biotechnol. Biofuels 2015, 8, 210. [Google Scholar] [CrossRef] [PubMed]
- Stoll, K.I.; Herbig, S.; Zwick, M.; Boukis, N.; Sauer, J.; Neumann, A.; Oswald, F. Fermentation of H2 and CO2 with Clostridium ljungdahlii at elevated process pressure—first experimental results. Chem. Eng. Trans. 2018, 64, 151–156. [Google Scholar] [CrossRef]
- Simpson, S.D.; Köpke, M.; Smart, K.F.; Tran, L.P.; Sechrist, P. A System and a Method for Controlling Metabolite Production in a Microbial Fermentation. World Intellectual Property. Organization Patent No. WO 2014/151158 A1, 25 September 2014. [Google Scholar]
- Saxena, J.; Tanner, R.S. Effect of trace metals on ethanol production from synthesis gas by the ethanologenic acetogen, Clostridium ragsdalei. J. Ind. Microbiol. Biotechnol. 2011, 38, 513–521. [Google Scholar] [CrossRef] [PubMed]
- Richter, H.; Martin, M.E.; Angenent, L.T. A two-stage continuous fermentation system for conversion of syngas into ethanol. Energies 2013, 6, 3987–4000. [Google Scholar] [CrossRef]
- Park, S.; Ahn, B.; Kim, Y.K. Growth enhancement of bioethanol-producing microbe Clostridium autoethanogenum by changing culture medium composition. Bioresour. Technol. Rep. 2019, 6, 237–240. [Google Scholar] [CrossRef]
- Ragsdale, S.W.; Pierce, E. Acetogenesis and the Wood-Ljungdahl pathway of CO2 fixation. Biochim. Biophys. Acta 2008, 1784, 1873–1898. [Google Scholar] [CrossRef]
- Bender, G.; Pierce, E.; Hill, J.A.; Darty, J.E.; Ragsdale, S.W. Metal centers in the anaerobic microbial metabolism of CO and CO2. Metallomics 2011, 3, 797–815. [Google Scholar] [CrossRef][Green Version]
- Smart, K.F.; Ly, B.S. Fermentation Process for the Production and Control of Pyruvate-Derived Products. U.S. Patent No. US 9,701,987 B2, 11 July 2017. [Google Scholar]
- Ragsdale, S.W. Catalysis of methyl group transfers involving tetrahydrofolate and B12. Vitam. Horm. 2008, 79, 293–324. [Google Scholar] [CrossRef]
- Korkhin, Y.; Kalb (Gilboa), A.J.; Peretz, M.; Bogin, O.; Burstein, Y.; Frolow, F. NADP-dependent bacterial alcohol dehydrogenases: Crystal structure, cofactor-binding and cofactor specificity of the ADHs of Clostridium Beijerinckii and Thermoanaerobacter Brockii. J. Mol. Biol. 1998, 278, 967–981. [Google Scholar] [CrossRef]
- Charubin, K.; Papoutsakis, E.T. Direct cell-to-cell exchange of matter in a synthetic Clostridium syntrophy enables CO2 fixation, superior metabolite yields, and an expanded metabolic space. Metab. Eng. 2019, 52, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Ragsdale, S.W. Enzymology of the wood-ljungdahl pathway of acetogenesis. Ann. N. Y. Acad. Sci. 2008, 1125, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Drennan, C.L.; Doukov, T.I.; Ragsdale, S.W. The metalloclusters of carbon monoxide Dehydrogenase/Acetyl-CoA Synthase: A Story in Pictures. J. Biol. Inorg. Chem. 2004, 9, 511–515. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.J.; Lee, J.; Lee, H.S. Enhanced alcohol production from synthesis gas using a CO-resistant mutant of Clostridium sp. AWRP. Microbiol. Biotechnol. Lett. 2019, 47, 581–584. [Google Scholar] [CrossRef]
- Scopes, R.K. An Iron-activated alcohol Dehydrogenase. FEBS Lett. 1983, 156, 303–306. [Google Scholar] [CrossRef]
- Simpson, S.; Tran, P.L.; Mihalcea, C.D.; Fung, J.M.Y.; Liew, F.M. Production of Butanediol by Anaerobic Microbial Fermentation. World Intellectual Property. Organization Patent No. WO/2009/151342, 17 December 2009. [Google Scholar]
- Simpson, S.; Fleming, S.E.; Havill, A.M.; Trevethick, S.R. Process for Producing Chemicals Using Microbial Fermentation of Substrates Comprising Carbon Monoxide. U.S. Patent No. US 2012/0045807 A1, 23 February 2012. [Google Scholar]
- Abubackar, H.N.; Veiga, M.C.; Kennes, C. Impact of cyclic pH shifts on carbon monoxide fermentation to ethanol by Clostridium autoethanogenum. Fuel 2016, 178, 56–62. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).