Abstract
White shrimp Litopenaeus vannamei is an important species of farmed shrimp. Intestinal bacterial composition and immune activity play important roles in regulating the health condition of shrimp. Lactic acid bacteria Limosilactobacillus fermentum SWP-AFFS02 was isolated from the intestine of sea fish Rachycentron canadum, and the potential of its effect on growth, immunity, and intestinal microbiota of L. vannamei shrimp was investigated. Shrimps received feed with or without the addition of 8 log CFU/g L. fermentum SWP-AFFS02 thrice a day for 8 weeks. After 8-week treatment, weight gain, feed conversion rate, and survival rate of shrimp were greater in the L. fermentum SWP-AFFS02-feed group than in the control group. L. fermentum SWP-AFFS02 treatment increased the number of granular cells and semi-granular cells and decreased hyaline cell number when compared to the control group. L. fermentum SWP-AFFS02 promoted prophenoloxidase (PO) activity through increasing immune-associated gene expression in the hepatopancreas of shrimp. In addition, administration of feed containing L. fermentum SWP-AFFS02 regulated intestinal microbiota via decreasing the ratio of pathogenic bacteria, such as Vibrionaceae and Enterobacteriaceae, in the intestine of shrimp. This study demonstrated that administration of L. fermentum SWP-AFFS02 effectively prevented infection of L. vannamei shrimp by regulating intestinal microbiota and enhancing immunity in shrimp to increase the growth and improve their health status, which acted as a probiotic and provided beneficial effects on shrimp.
1. Introduction
Litopenaeus vannamei, commonly known as white shrimp, is native to Central and South America, which has strong tolerance to the environment, disease resistance, and osmotic pressure regulation in low-salinity waters. It is characterized by its feed conversion rate, fast growth rate, low residual food rate, and suitability for high breeding density, which have made it an internationally important species of farmed shrimp. Developing an intensive farming model to enable an increase in stocking density per unit area has caused problems, such as water contamination. Chemicals, growth of pathogenic bacteria, and immunity decline lead to the occurrence of diseases in the aquaculture industry [1]. However, the use of antibiotics causes the pathogenic bacteria to develop drug resistance, which makes it more difficult to treat diseases during the aquaculture period.
There are many types of bacteria in the intestinal tract. These bacteria can successfully adsorb on the surface of intestinal mucosal cells and proliferate in the intestinal tract. Moreover, intestinal bacteria can be roughly divided into three categories: (i) normal flora that often exists in the intestine of the host, (ii) probiotics that are beneficial to the health of the host, and (iii) opportunistic or pathogenic bacteria. The interactions (bacterial interference, bacterial antagonism, barrier effect, and competitive exclusion) between probiotics and pathogenic bacteria play an important role in stabilizing the intestinal physiological functions of the host, including intestinal immune activity and intestinal bacterial composition [2].
Feeding grass shrimp (Penaeus monodon) with Lactobacillus spp. as a probiotic can increase the resistance to the pathogenic bacteria Vibrio harveyi [3], and feeding with the shrimp Bacillus subtilis BP11 can increase their survival rate, improve immune activity, and reduce V. harveyi infection [4,5]; adding Lactobacillus plantarum to the feed can improve the intestinal function and enzyme secretion capacity of L. vannamei to elevate the metabolic utilization of feed and increase the growth rate [6]. In addition, studies have shown that the use of prebiotics to maintain water quality during shrimp farming can effectively resist the growth of pathogenic bacteria and increase the growth rate and production of shrimp [7]. Sha et al. (2016) found that adding different probiotics (L. pentosus and Enterococcus faecium) or the bacteria-free supernatant of a L. pentosus culture to the feed can significantly increase Actinobacteria in the intestine and maintain physiological health in L. vannamei shrimp [8]. Therefore, B. subtilis and lactic acid bacteria (LAB) are gradually being used in aquaculture. According to a recent study, the benefits of probiotics isolated from aquaculture species (fish and shrimp) are greater than those obtained from non-aquaculture species [9]. Dash et al. (2014) showed that another kind of LAB (approximately 3–9 cfu/mL) can also be determined in pond water after adding L. plantarum to the pond water of Macrobrachium rosenbergii [10], indicating that probiotic treatment may improve the quality and microbiota of the water environment, which promote the growth of other types of probiotics. These results suggest that LAB are adaptable to the environment. We isolated the bacterium Limosilactobacillus fermentum SWP-AFFS02 from the intestine of the sea fish Rachycentron canadum. The aim of this study was to evaluate the potential of LAB feed containing L. fermentum SWP-AFFS02 to regulate the growth, immunity, and intestinal microbiota of L. vannamei shrimp. The results in this study are the first to prove that L. fermentum SWP-AFFS02 has the potential to act as a probiotic.
2. Materials and Methods
2.1. Culture of L. fermentum SWP-AFFS02
LAB L. fermentum SWP-AFFS02 was isolated from the intestine of the sea fish R. canadum via MRS selection medium with CaCO3. After the Gram stain and catalase test, the strain was identified by 16S ribosomal DNA sequencing. LAB were cultured in MRS broth under anaerobic equipment (Oxoid, Basingstoke, Hampshire, England) at 37 °C, and the numbers of the LAB were detected on MRS agar plates (Difco Laboratories, Detroit, MI, USA). The commercial feed (young eel feed No. F; Tung Li Feed Industrial Co., LTD, PingTung, Taiwan) was inoculated with precultured LAB in MRS for 30 min (feed: MRS = 2:1; w/w). Finally, the LAB-dietary feed was freeze-dried by a freeze drier system (Rebers, Taoyuan, Taiwan) and stored at −20 °C.
2.2. The Number of LAB in Feed
According to the method of Wirunpan et al. (2016) [11], the feed with added LAB (1 g) was crushed, mixed with 9 mL of PBS, and diluted to appropriate concentration by serial dilution, then 0.1 mL of the suspension was inoculated on MRS agar. After cultivation for 24 h at 37 °C, counting colony numbers was carried out.
2.3. Husbandry Conditions and Feeding Trial
Shrimps used for this study were collected from an aquafarm at Kaohsiung, Taiwan. A total of 180 L. vannamei shrimps (initial body weight: 8.22 ± 0.38 g and 8.27 ± 0.53 g in blank and L. fermentum SWP-AFFS02 feed, respectively) were evenly distributed into 3 groups (60 shrimps/group), and they were stocked in a fiber reinforced plastics (FRP) bucket with a flow-through sea water system and continuous aeration. The indoor cement pool was maintained under natural light/dark regime. During the experimental period, the monitored water quality parameters (mean ± S.D.) were as follows: water temperature was 28.3 ± 2.4 °C, pH was 8.4 ± 0.5, and the salinity was 33.6 ± 0.2 ppt. All shrimp groups were fed feed with or without LAB to apparent satiation thrice a day (at 6:00, 17:00, and 22:00) for 8 weeks. The remaining feed residues after feeding were removed by siphoning and dried by using a freeze drier system to subtract from total feed intake. Body weight was also recorded once every two weeks.
Weight gain = ((final weight – initial weight)/initial weight) × 100%
Feed conversion rate (FCR) = feed supplied/(final weight – initial weight)
Survival (%) = (number of individuals at the end of the evaluation period/initial number of individuals stocked) × 100%
2.4. Detection of Total Bacteria and Vibrio spp. Counts in Seawater Surrounding
The 1 mL of aquaculture breeding water was diluted to the appropriate ratio by a 10-fold serial dilution method, and the 100 μL dilution solutions were smeared on 3% tryptone soy agar (TSA) plate medium (BD, Franklin Lakes, NJ, USA) and thiosulfate-citrate-bile salts-sucrose (TCBS) agar (Sigma–Aldrich, St. Louis, MO, USA) for total bacteria and Vibrio spp. cultivation, respectively. After cultivation for 24 h at 30 °C, the bacterial numbers between 25–250 colonies were counted [12].
2.5. Assay for Immune Activity (Hymolymph Parameters)
2.5.1. Preparation of Hemolymph Solution
The 0.8 mL of hemolymph was withdrawn from the ventral sinus in the first abdominal segment using a 26-gauge hypodermic needle. Each syringe was pre-filled with 0.2 mL of anticoagulant (10 mM Tris–HCl, 250 mM sucrose, 100 mM sodium citrate, pH 7.6). More anticoagulant was added to make an equal volume ratio of hemolymph to anticoagulant. The hemolymph samples of 180 L. vannamei shrimps (60 shrimps/group) from each treatment were analyzed individually [13].
2.5.2. Total Hemocyte Count (THC) and Differential Hemocyte Count (DHC)
A volume of 50 μL anticoagulated hemolymph was fixed with an equal volume of neutral buffered formalin (10%) for 30 min to measure the THC. Fixed hemolymph was smeared on a slide and stained with Giemsa solution (10%) for 10 min. The DHC was then characterized according to Tsing et al. (1989) [14], and 250–300 cells from each smear were counted. The remaining anticoagulated hemolymph was centrifuged at 300× g for 10 min at 4 °C to separate the hemocytes from plasma. Hemocytes were suspended and adjusted to 5 × 106 cells/mL in ice-cold cacodylate buffer (0.01 M sodium cacodylate, 0.45 M NaCl, 10 mM CaCl2, 26 mM MgCl2, pH 7.0). A volume of 2.5 mL (5 × 106 cells/mL) hemocyte suspension was respectively separated for superoxide anion and phenoloxidase activity [13].
2.5.3. Measurement of Superoxide Anion Respiratory Burst
According to recent study [15], after adding 100 μL (5 × 106 cells/mL) of diluted hemolymph to the wells of a 96-well microplate, the plate was centrifuged at 800× g at 4 °C for 20 min. The supernatant was discarded, and the pellets were rinsed with MCHBSS buffer (Sigma-Aldrich, St. Louis, MO, USA). Subsequently, he mocytes were mixed with 100 μL of DCFH-DA (Sigma-Aldrich, St. Louis, MO, USA) and zymosan (1 mg/mL) (Sigma-Aldrich, St. Louis, MO, USA) in the 96-well plate. After a 30-min incubation in the dark, cells were washed twice with MCHBSS buffer. The supernatant was discarded 30 min post-incubation and the pellets were rinsed once using MCHBSS buffer. Finally, the fluorescent intensity was measured using a spectrofluorometer with an excitation wavelength of 480 nm and an emission wavelength of 523 nm.
2.5.4. Assay for Phenoloxidase (PO) Activity
The PO activity was assayed, as described by Sung et al. (1994) [16]. using L-3,4-dihydroxyphenyl-alanine (L-dopa) (Sigma-Aldrich) as a substrate. To obtain lysates from hemolymph (200 μL; 5 × 106 cells/mL), frozen hemocytes were thawed and frozen several times. Hemocyte lysate supernatants (HLS) and plasma were assayed for PO activity using a spectrophotometer (Hitachi U-2000) to measure the OD490 nm. The concentration of total plasma protein was determined by the Bradford method [17] using bovine serum albumin (Bio-Rad Protein assay Kit II) as a standard. One unit of PO activity was defined as an increase in absorbance of 0.001 min/mg protein [18].
2.5.5. Reverse Transcription Quantitative PCR (RT-qPCR)
Total RNA from hepatopancreas was obtained using the Trizol reagent (Gibco BRL Life Technologies, Inc., Gaithersburg, MD, USA), according to the manufacturer’s instructions. Primers were synthesized by MD-Bio, Inc. (Taipei, Taiwan). The gene expression level was determined by relative quantification using RT-qPCR (CFX Cycler System, Bio-Rad Laboratories, Inc., Hercules, CA, USA). Primers: HSP70 (F): CAACGATTCTCAGCGTCAGG and HSP70 (R): ACCTTCTTGTCGAGGCCGTA; beta-actin (F): GCCCTGTTCCAGCCCTCATT and beta-actin (R): ACGGATGTCCACGTCGCACT [19]. PGx (F): GGCACCAGGAGAACACTAC and GPx (R): CGACTTTGCCGAACATAAC; LGBP (F): CCATGTCCGGCGGTGGAA and LGBP (R): GTCATCGCCCTTCCAGTTG; PPAF (F): GAGAAGGAGCTGAACCTGTAC and PPAF (R): AGCGCCTGAGTTGTAGTTAG [20]. PCR conditions were according to the literature [20].
2.6. Microbiota Sequencing
Each intestine of shrimp was collected and immediately soaked in liquid nitrogen and stored at −80 °C for subsequent use. Total genomic DNA from samples was extracted using QIAamp PowerFecal DNA Kit (Qiagen) for 16S rRNA gene sequencing. V3-V4 regions (515F-806R) were amplified, and PCR reactions were carried out with Phusion® High-Fidelity PCR Master Mix (New England Biolabs). The sequencing library was prepared and sequenced on an Illumina MiSeq platform.
2.7. Statistical Analysis
Experimental results were analyzed in five repeats and expressed as the mean ± standard error of mean. The results were examined by using one-way analysis of variance (ANOVA) and Duncan’s multiple range tests, and the significance of differences between sample means was calculated. A p value ≤ 0.05 was considered significant.
3. Results
3.1. Variation of LAB Feed-Contained L. fermentum SWP-AFFS02 after Stroage
L. fermentum SWP-AFFS02 was collected after cultivation for 48 h (approximately 109 CFU/mL) in MRS medium at 37 °C. The standard feed (sterilized) was mixed with a culture solution or deionized water (1/1: w/v) under laminar airflow and stored at 4 °C or 25 °C for 7 days after freeze-drying and vacuum-packaging. Finally, the LAB numbers in the L. fermentum SWP-AFFS02-rich feed were measured and calculated on MRS agar under anaerobic conditions. As shown in Table 1, the bacterial counts were 4.47 × 108 CFU/g on day 0. However, a decrease in bacterial levels was observed with increasing storage days (during days 0–7) in both 25 °C and 4 °C treatments. However, maintaining a low temperature could effectively retain the number of viable bacteria.

Table 1.
The variation of L. fermentum SWP-AFFS02 level in LAB feed after storage for 7 days at 25 °C or 4 °C.
3.2. Regulations of Growth, Environmental Microbe, and Immunity in L. vannamei Shrimp by LAB Feed-Contained L. fermentum SWP-AFFS02
3.2.1. Growth, FCR, and Survival Rate of Shrimp
The initial body weights of L. vannamei shrimp were 8.22 and 8.27 g in the control and LAB feed groups, respectively. The weight gain (WG), feed conversion rate (FCR), and survival of L. vannamei shrimp in the two groups were evaluated after 8 weeks. We found that the final weight of L. vannamei shrimp in the LAB group was higher than that in the control group. WG, FCR, and survival rate were greater in the LAB feed group than in the control group (Table 2).
Table 2.
The effects of LAB feed on the growth, FCR, and survival rate of L. vannamei shrimp.
3.2.2. Environmental Microbial Population (Total Bacteria Level and Vibrio spp.)
The results indicated that the total bacterial levels in environmental seawater were higher in the LAB feed group than in the control group between weeks 2 and 8 (Table 3). Vibrio spp. resulted in growth interference and yield reduction in brine shrimp (Artemia franciscana) [21]. Therefore, the level of Vibrio spp. in the environment was monitored from week 0 to week 8. We found that the number of Vibrio spp. increased with increasing aquaculture period in environmental seawater in both the control and LAB feed groups, but no significant difference was found between these two groups (Table 4).
Table 3.
The total bacterial numbers in environmental seawater.
Table 4.
The Vibrio spp. numbers in environmental seawater.
3.2.3. Investigation for Immunity Index
Total hemocyte count (THC) is a marker to indicate shrimp immunity, and the THC ratio approximately reaching 2.0–4.0 × 107 cells/mL was reported [22]. Another study demonstrated that THC levels were affected by internal factors (including sex, shelling cycle, age, food intake, season, and body temperature), as well as external environmental factors (including pollutants, temperature, pH, salinity, and dissolved oxygen) in crustaceans [23]. Moreover, the THC level was markedly reduced in shrimp, thereby leading to infections caused by pathogenic bacteria or viruses [24]. We found that the LAB feed markedly elevated THC levels in shrimp when compared to the control group (Figure 1A).
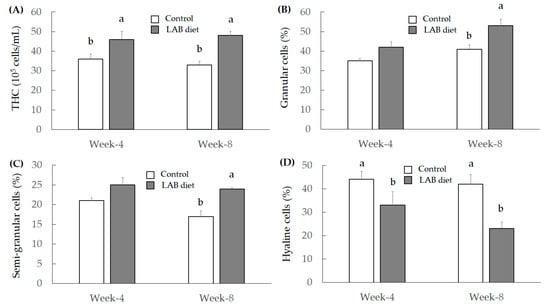
Figure 1.
The variations of THC (A), granular cells (B), semi-granular cells (C), and hyaline cells (D) in shrimp feed with control or LAB feed. Significant difference is shown by different letters (a, b) (p < 0.05) between control and LAB feed groups.
Differential hemocyte count (DHC) was divided into three types of immunocytes, including granular cells, semi-granular cells, and hyaline cells, and these cells constituted THC. In addition, DHC plays different roles in regulating immune function. Granular cells and semi-granular cells exert cytotoxicity and activate the prophenoloxidase system (PO system), but hyaline cells do not activate the PO system. Phagocytosis and respiratory burst (production of superoxide anions) are the major immune abilities of hyaline cells [25]. We found that the LAB feed markedly increased the number of granular cells (Figure 1B) and semi-granular cells (Figure 1C) after 8 weeks of administration. However, the number of hyaline cells treated with the LAB feed was reduced both in week 4 and week 8, compared to the control group (Figure 1D).
Respiratory burst was monitored to identify the body defense ability mediated by superoxide anion production; respiratory burst activation has been found to improve the shrimp defense system [26]. The results indicated that LAB feed administration did not promote respiratory burst (Figure 2A), which may be because the LAB feed could not promote hyaline cells. The activation of the PO system led to quinonoid production mediated by tyrosine oxidation, and quinonoid compounds exerted cytotoxicity [27]. As shown in Figure 2B, the LAB feed markedly promoted the PO activity of L. vannamei shrimp in both week 4 and week 8. These results demonstrate that L. fermentum SWP-AFFS02 could be developed as a probiotic to improve immunity and growth in L. vannamei shrimp, thereby elevating FCR efficacy.
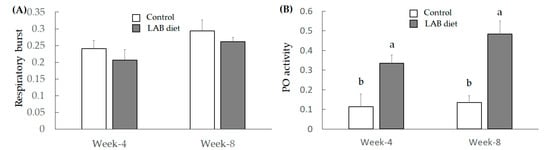
Figure 2.
The regulation of LAB feed on respiratory burst (index of hyaline cell) (A) and PO activity (granular cells) (B). Significant difference is shown by different letters (a, b) (p < 0.05) between control and LAB feed groups.
The lipopolysaccharide and beta-1,3-glucan binding protein (LGBP) and the cascade via PO-activating factor (PPAF) both activate the PO system, thereby promoting immune function in shrimp [28]. In addition, heat-shock protein 70 (HSP70) and glutathione peroxidase (GPx) are defense agents associated with antioxidant activity against stress [29]. The LAB feed containing L. fermentum SWP-AAS02 significantly increased LGBP and PPAF expression in the hepatopancreas of L. vannamei shrimp (Figure 3A,B), but the levels of HSP70 and GPx were not elevated by LAB feed administration (Figure 3C,D). These results indicate that L. fermentum SWP-AFFS02 markedly elevated immune activity in L. vannamei shrimp.
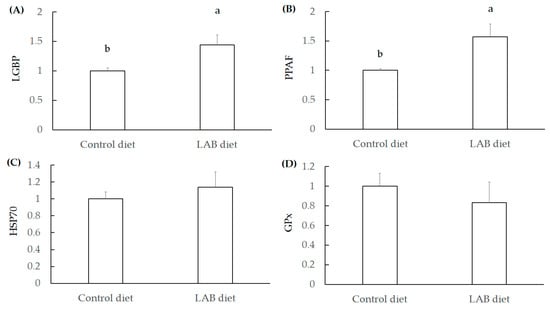
Figure 3.
Expression levels of LGBP (A), PPAF (B), HSP70 (C), and GPx (D) in the hepatopancreas of L. vannamei shrimp fed with LAB feed containing L. fermentum SWP-AFFS02 as determined by RT qPCR. Significant difference is shown by different letters (a, b) (p < 0.05) between control and LAB feed groups.
3.3. Protection of Intestinal Microbiota in L. vannamei Shrimp by LAB Feed Containing L. fermentum SWP-AFFS02
There are 391 different types of bacteria within the intestinal stool of L. vannamei shrimp fed a control feed or LAB feed. However, the operational taxonomic unit (OTU) numbers were 126 and 201 in the intestinal stool of L. vannamei shrimp fed a control feed or a LAB feed, respectively (Figure 4A). These results suggested that the LAB feed containing L. fermentum SWP-AFFS02 may potentially regulate microbial beta diversity in the intestine, compared to the control group, as shown by PCA. Furthermore, the microbial beta-diversity of seawater was different from that of the stools of L. vannamei shrimp fed with control and LAB feed (Figure 4B).
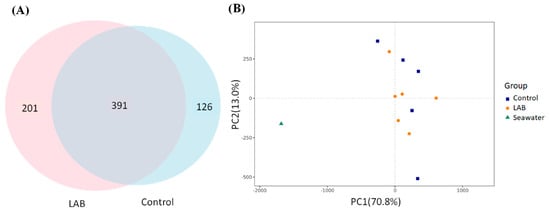
Figure 4.
Alpha and Beta diversity of intestinal microbiota in L. vannamei shrimp. (A) Venn diagram. The overlapping part indicates the number of OTUs shared between control and LAB treatment groups, and the non-overlapping part indicates the number of unique OTUs between two groups. (B) Principal component analysis (PCA) for species. Each point in the figure represents one sample, and the same grouped samples are represented by the same color.
We analyzed the most abundant microbes (top 35) at the genus and family levels in the stool of L. vannamei shrimp fed a control feed or a LAB feed. The results indicated that Pseudomonas, Bacillus, Idiomarina, NS5_marine_group, Aliidiomarina, and Alcanivorax at genus level (Figure 5A) and Flavobacteriaceae, Pseudomonadaceae, Bacillaceae, Idiomarinaceae, Rhodobacteraceae, Legionellaceae, Halieaceae, Alcanivoracaceae, Paenibacillaceae, Halomonadaceae, and Shingomonadaceae at family level (Figure 5B) in seawater were higher than in the stool of L. vannamei shrimp fed with control or LAB feed; however, other bacteria in seawater were markedly lower than those in the stool of L. vannamei shrimp, suggesting that some mechanisms may regulate special microbes remaining in the shrimp intestine, compared to seawater bacteria staying in shrimp. In addition, the levels of Paraprevotella, Diallister, Oscillibacter, Lachnoclostridium, and Sutterella at the genus level were elevated by LAB feed-containing L. fermentum SWP-AFFS02, compared to the control group; however, the levels of Acinetobacter, Ruminococcaceae_UCG_002, and Escherichia_Shigella were reduced by LAB feed-containing L. fermentum SWP-AFFS02, compared to the control group. Moreover, we found that the levels of Vibrionaceae, Corynebacteriaceae, Marinifilaceae, Tannerellaceae, and Enterobacteriaceae were suppressed by the LAB feed. These results indicate that the LAB feed potentially regulates the intestinal microbiota to improve host health in L. vannamei shrimp.
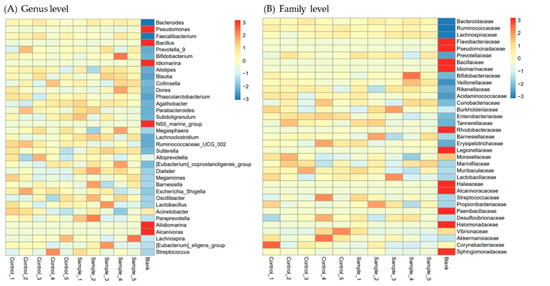
Figure 5.
Heatmap of intestinal microbiota in L. vannamei shrimp. According to the species annotation and abundance information of all samples in the (A) genus level and (B) family level, we selected the top 35 in abundance.
Moreover, short-chain fatty acids, including acetic acid, propionic acid, and butyric acid in the stools of L. vannamei shrimp fed the LAB feed were measured, as shown in Table 5. The results showed that acetic acid and butyric acid were elevated in the stools of L. vannamei shrimp fed the LAB feed, compared to the control group.
Table 5.
The level of intestinal short chain fatty acid in L. vannamei shrimp fed with LAB feed-contained L. fermentum SWP-AFFS02.
4. Discussion
A study has found that a Bacillus subtilis-fermented soybean meal could improve intestinal microbiota and disease prevention in shrimp [30]. Xie et al. (2017) indicated that piglets fed feed fermented by Lactobacillus, Bacillus, and Saccharomyces cerevisiae potentially improved growth performance [31]. In a similar study in fish, Lactobacillus and Pediococcus were abundant bacteria in the intestines of Atlantic salmon fed the fermented feed, and the expression of mucin 2 (muc2) and aquaporin 8ab (aqp8ab) significantly increased in the fermented feed group, compared to the control group [32].
Vibrio spp. are widely distributed in marine environments and are among the most abundant flora in shrimp digestive systems, and vibriosis is the most serious disease in worldwide aquaculture that causes shrimp death [33,34]. Huang et al. (2016) demonstrated that pathogenic Photobacterium and Vibrio were the predominant bacteria at the post-larval stage (80%) and juvenile stage (89.1% ~ 94.2%), resulting in shrimp infection [35]. Similar to this study, Vibrio spp. were predominantly found in the intestines of shrimp, but these pathogenic bacteria were reduced in shrimp fed a synbiotic feed [34], thereby reducing disease arising from Vibrio infection by promoting immunity [36,37], as well as by avoiding hepatopancreatic damage [38]. A previous study suggested that activation of the antioxidant system avoided these negative effects caused by Vibrio infection in shrimp [39]. The immune and antioxidant systems are two primary physiological mechanisms that protect aquatic animal health against Vibrio spp. infection [40]. Non-specific immunity is the major defense system for stress resistance and protection against pathogen infection in crustaceans, and the PO system is also one of the primary response pathways in shrimp [41]. PPAF and LGBP are the key regulators of the immune response in shrimp [28]. In our study, we found that the LAB feed containing L. fermentum SWP-AFFS02 potentially suppressed intestinal Vibrionaceae in shrimp (Figure 5B) and promoted immune-associated gene expression in the hepatopancreas of L. vannamei shrimp, including LGBP and PPAF (Figure 3A,B). Acinetobacter spp. is a pathogenic bacterium of L. vannamei shrimp [42]. Corynebacterium is a gram-positive (non-spore forming) and aerobic bacterium that opportunistically reduces immunity and defense systems in hosts associated with systemic infection [43]. Escherichia_Shigella (Enterobacteriaceae family) is a pathogenic bacterium that causes gut leakiness and infection [44]. We found that administration of LAB feed that containing L. fermentum SWP-AFFS02 markedly inhibited these pathogenic intestinal bacteria in L. vannamei shrimp.
The animal intestine plays an important role in nutrient absorption and metabolism and acts as the first line of defense against pathogen infection and environmental stress [45]. Intestinal bacteria and bacterial metabolites directly influence the physiology of the host, including absorption and metabolism, intestinal barrier integrity, and immune function [46]. We believe that the LAB feed containing L. fermentum SWP-AFFS02 could limit pathogenic bacteria in the intestine of L. vannamei shrimp, thereby improving growth, FCR, and survival (Table 2).
In a recent study, feeding L. vannamei with feed supplemented with bacteria (Bacillus subtilis and L. rhamnosus) enhanced the antioxidative status by increasing the activity of antioxidant enzymes in shrimp [47]. Thus, shrimp feed with bio-friendly agents. such as Lactobacillus spp., could be used as effective alternatives to antibiotics for treating bacterial infections, especially V. harveyi infection in shrimp aquaculture [48]. Pooljun et al. (2020) confirmed that, after feeding juvenile shrimp (Penaeus vannamei) with L. acidophilus and Saccharomyces cerevisiae mixture (1:1, at 108 and 109 CFU/kg feed), the adverse effects of acute hepatopancreas necrosis disease induced by V. parahaemolyticus infection were ameliorated by improving hemocyte parameters, including the THC, granular hemocyte percentage, and phenoloxidase activity, as well as by highly upregulating hemocyte genes (crustin and penaeidin), which lead to high shrimp survival rate [49].
5. Conclusions
This study aimed to evaluate the effects of feeding L. vannamei shrimp with a feed rich in the LAB L. fermentum SWP-AFFS02. The effect of L. fermentum SWP-AFFS02 on the immunity and microbiota, as well as the changes in size of white shrimp, were investigated. Our study makes a significant contribution to the literature due to L. vannamei being an economically important species of shrimp, which are often plagued by pathogenic bacteria that cause their death. We noticed a marked inhibition of pathogenic bacteria in the shrimp intestine, which could in turn promote growth and improve immunity in L. vannamei. This study provided useful insights for the use of L. fermentum SWP-AFFS02 in shrimp. Enhancing the health status of cultured shrimp by using beneficial microbes as a probiotic is a good approach to control pathogens.
Author Contributions
B.-H.L. and W.-H.H. performed the design for the framework of the study. B.-H.L. determined most of the experimental assays. Y.-Z.C. analyzed the data. T.-M.P. and W.-H.H. revised the manuscript. T.-M.P., B.-H.L., W.-H.H., and K.-T.H. participated in the study design and finished the draft. The corresponding author T.-M.P. was responsible for financial resources and funds for the project, supervision of the research activities, and the manuscript submission. The corresponding author T.-M.P. led the research group. All authors have read and agreed to the published version of the manuscript.
Funding
This research was founded by SunWay Biotech Co., Ltd., Taipei, Taiwan, grant number NCKU B110-K032.
Ethical Approval
It is not required to get an ‘Affidavit of Approval of Animal use Protocol’ when using invertebrates, including shrimp, as an experimental animal in Taiwan.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
This research work and subsidiary spending were mainly supported by SunWay Biotech Co., Ltd., Taipei, Taiwan.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ashley, P.J. Fish welfare: Current issues in aquaculture. Appl. Anim. Behav. Sci. 2007, 104, 199–235. [Google Scholar] [CrossRef]
- Rasheeda, M.K.; Rangamaran, V.R.; Srinivasan, S.; Ramaiah, S.K.; Gunasekaran, R.; Jaypal, S.; Gopal, D.; Ramalingam, K. Comparative profiling of microbial community of three economically important fishes reared in sea cages under tropical offshore environment. Mar. Genom. 2017, 34, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Utiswannakul, P.; Sangchai, S.; Rengpipat, S. Enhanced growth of black tiger shrimp Penaeus monodon by dietary supplementation with Bacillus (BP11) as a probiotic. J. Aquac. Res. Dev. 2011, 2, 6. [Google Scholar] [CrossRef]
- Rengpipat, S.; Tunyanun, A.; Fast, A.W.; Piyatiratitivorakul, S.; Menasveta, P. Enhanced growth and resistance to Vibrio challenge in pond-reared black tiger shrimp Penaeus monodon fed a Bacillus probiotic. Dis. Aquat. Org. 2003, 55, 169–173. [Google Scholar] [CrossRef] [PubMed]
- Vaseeharan, B.; Ramasamy, P. Control of pathogenic Vibrio spp. Bacillus subtilis BT23, by a possible probiotic treatment for black tiger shrimp Penaeus monodon. Lett. Appl. Microbiol. 2003, 36, 83–87. [Google Scholar] [CrossRef]
- Zheng, X.; Duan, Y.; Dong, H.; Zhang, J. Effects of dietary Lactobacillus plantarum on growth performance, digestive enzymes and gut morphology of Litopenaeus vannamei. Probiotics Antimicrob Proteins 2018, 10, 504–510. [Google Scholar] [CrossRef] [PubMed]
- Prabhu, N.M.; Nazar, A.R.; Rajagopal, S.; Khan, S.A. Use of probiotics in water quality management during shrimp culture. J. Aquac. Trop. 1999, 14, 227–236. [Google Scholar]
- Sha, Y.; Liu, M.; Wang, B.; Jiang, K.; Qi, C.; Wang, L. Bacterial population in intestines of Litopenaeus vannamei fed different probiotics or probiotic supernatant. J. Microbiol. Biotechnol. 2016, 26, 1736–1745. [Google Scholar] [CrossRef]
- Li, C.; Zhang, B.; Wang, X.; Pi, X.; Wang, X.; Zhou, H.; Mai, K.; He, G. Improved utilization of soybean meal through fermentation with commensal Shewanella sp. MR-7 in turbot (Scophthalmus maximus L.). Microb. Cell Fact. 2019, 18, 214. [Google Scholar] [CrossRef]
- Dash, G.; Raman, R.P.; Prasad, K.P.; Makesh, M.; Pradeep, M.A.; Sen, S. Evaluation of Lactobacillus plantarum as feed supplement on host associated microflora, growth, feed efficiency, carcass biochemical composition and immune response of giant freshwater prawn, Macrobrachium rosenbergii (de Man, 1879). Aquaculture 2014, 432, 225–236. [Google Scholar] [CrossRef]
- Wirunpan, M.; Savedboworn, W.; Wanchaitanawong, P. Survival and shelf life of Lactobacillus lactis 1464 in shrimp feed pellet after fluidized bed drying. Agric. Nat. Resour. 2016, 50, 1–7. [Google Scholar] [CrossRef][Green Version]
- Ngo, H.V.T.; Huang, H.T.; Lee, P.T.; Liao, Z.H.; Chen, H.Y.; Nan, F.H. Effects of Phyllanthus amarus extract on nonspecific immune responses, growth, and resistance to Vibrio alginolyticus in white shrimp Litopenaeus vannamei. Fish Shellfish Immunol. 2020, 107, 1–8. [Google Scholar] [CrossRef]
- Song, Y.L.; Yu, C.I.; Lien, T.W.; Huang, C.C.; Lin, M.N. Haemolymph parameters of Pacific white shrimp (Litopenaeus vannamei) infected with Taura syndrome virus. Fish Shellfish Immunol. 2003, 14, 317–331. [Google Scholar] [CrossRef] [PubMed]
- Tsing, A.; Arcier, J.M.; Brehelin, M. Hemocytes of penaeid and Palaemonid shrimps: Morphology, cytochemistry and hemegram. J. Invertebr. Pathol. 1989, 53, 64–77. [Google Scholar] [CrossRef]
- Lee, P.T.; Tran, H.T.Q.; Huang, H.T.; Nan, F.H.; Lee, M.C. Sargassum horneri extracts stimulate innate immunity, enhance growth performance, and upregulate immune genes in the white shrimp Litopenaeus vannamei. Fish Shell Immunol. 2020, 102, 276–285. [Google Scholar] [CrossRef]
- Sung, H.H.; Kou, G.H.; Song, Y.L. Vibriosis resistance induced by glucan treatment in tiger shrimp (Penaeus monodon). Fish Pathol. 1994, 29, 11–17. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of proteins utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Söderhäll, K.; Unestam, T. Activation of serum prophenoloxidase in arthropod immunity: The specificity of cell wall glucan activation and activation by purified fungal glycoproteins of crayfish phenoloxidase. Can. J. Microbiol. 1979, 25, 406–414. [Google Scholar] [CrossRef]
- Duan, Y.F.; Wang, Y.; Zhang, J.S.; Sun, X.X.; Wang, J. Dietary effects of succinic acid on the growth, digestive enzymes, immune response and resistance to ammonia stress of Litopenaeus vannamei. Fish Shellfish Immunol. 2018, 8, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Xie, S.W.; Zheng, L.; Wan, M.G.; Niu, J.; Liu, Y.J.; Tain, L.X. Effect of deoxynivalenol on growth performance, histological morphology, anti-oxidative ability and immune response of juvenile Pacific white shrimp, Litopenaeus vannamei. Fish Shellfish Immunol. 2018, 82, 442–452. [Google Scholar] [CrossRef]
- Vanmaele, S.; Defoirdt, T.; Cleenwerck, I.; De Vos, P.; Bossier, P. Characterization of the virulence of Harveyi clade vibrios isolated from a shrimp hatchery in vitro and in vivo, in a brine shrimp (Artemia franciscana) model system. Aquaculture 2015, 435, 28–32. [Google Scholar] [CrossRef]
- Moullac, G.L.; Groumellec, M.L.; Ansquer, D.; Froissard, S.; Levy, P. Haematological and phenoloxidase activity changes in the shrimp Penaeus stylirostris in relation with the moult cycle: Protection against Vibrio. Fish Shellfish Immunol. 1997, 7, 227–234. [Google Scholar] [CrossRef]
- Cheng, W.; Chen, J.C. Effects of environmental factors on the immune responses of freshwater prawn Macrobrachium rosenbergii and other decapod crustaceans. J.-Fisher Soc. Taiwan 2002, 29, 1–20. [Google Scholar]
- Cheng, W.; Chen, J.C. The virulence of Enterococcus to freshwater prawn Macrobrachium rosenbergii and its immune resistance under ammonia stress. Fish Shellfish Immonol. 2002, 12, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Risjani, Y.; Mutmainnah, N.; Manurung, P.; Wulan, S.N.; Yunianta. Exopolysaccharide from Porphyridium cruentum (purpureum) is not toxic and stimulates immune response against Vibriosis: The assessment using zebrafish and white shrimp Litopenaeus vannamei. Mar. Drugs 2021, 19, 133. [Google Scholar] [CrossRef]
- Rodriguez, J.; Le Moullac, G. State of the art of immunological tools and health control of Penaeid shrimp. Aquaculture 2000, 191, 109–119. [Google Scholar] [CrossRef]
- Chase, M.R.; Raina, K.; Bruno, J.M. Purification, characterization and molecular cloning of prophenol oxidases from Sarcophaga bullata. Insect Biochem. Mol. Biol. 2000, 30, 953–967. [Google Scholar] [CrossRef]
- Cerenius, L.; Soderhall, K. The prophenoloxidase-activating system in invertebrates. Immunol. Rev. 2004, 77, 21–26. [Google Scholar] [CrossRef]
- Guo, H.; Xian, J.A.; Li, B.; Ye, C.X.; Wang, A.L.; Miao, Y.T.; Liao, S.A. Gene expression of apoptosis-related genes, stress protein and antioxidant enzymes in hemocytes of whit shrimp Litopenaeus vannamei under nitrite stress. Comp. Biochem. Phys. C 2013, 157, 366–371. [Google Scholar]
- Cheng, A.C.; Yeh, S.P.; Hu, S.Y.; Lin, H.L.; Liu, C.H. Intestinal microbiota of white shrimp, Litopenaeus vannamei, fed diets containing Bacillus subtilis E20-fermented soybean meal (FSBM) or an antimicrobial peptide derived from B. subtilis E20-FSBM. Aquac. Res. 2019, 51, 41–50. [Google Scholar] [CrossRef]
- Xie, Z.; Hu, L.; Li, Y.; Geng, S.; Cheng, S.; Fu, X.; Zhao, S.; Han, X. Changes of gut microbiota structure and morphology in weaned piglets treated with fresh fermented soybean meal. World J. Microbiol. Biotechnol. 2017, 33, 213. [Google Scholar] [CrossRef]
- Catalán, N.; Villasante, A.; Wacyk, J.; Ramirez, C.; Romero, J. Fermented soybean meal increases lactic acid bacteria in gut microbiota of Atlantic salmon (Salmo salar). Probiotics Antimicrob Proteins 2018, 10, 566–576. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.Y.; Santos, H.M.; Hu, S.Y.; Sang, C.Y.; Yanuaria, C.A.S.; Lola, E.N.G.; Tayo, L.L.; Nituram, K.M.A.; Liu, C.H.; Chuang, K.P. LpxD gene knockout elicits protection to Litopenaeus vannamei, white shrimp, against Vibrio parahaemolyticus infection. Aquac. Int. 2019, 27, 1383–1393. [Google Scholar] [CrossRef]
- Huynh, T.G.; Hu, S.Y.; Chiu, C.H.; Truong, Q.P.; Liu, C.H. Bacterial population in intestines of white shrimp, Litopenaeus vannamei fed a synbiotic containing Lactobacillus plantarum and galactooligosaccharide. Aquac. Res. 2019, 50, 807–817. [Google Scholar] [CrossRef]
- Huang, Z.; Li, X.; Wang, L.; Shao, Z. Changes in the intestinal bacterial community during the growth of white shrimp, Litopenaeus vannamei. Aquac. Res. 2016, 47, 1737–1746. [Google Scholar] [CrossRef]
- Huynh, T.G.; Cheng, A.C.; Chi, C.C.; Chiu, K.H.; Liu, C.H. A synbiotic improves the immunity of white shrimp, Litopenaeus vannamei: Metabolomics analysis reveal compelling evidence. Fish Shellfish Immunol. 2018, 79, 284–293. [Google Scholar] [CrossRef]
- Li, E.; Chen, L.Q.; Zeng, C.; Yu, N.; Xiong, Z.Q.; Chen, X.F.; Qin, J.G. Comparison of digestive and antioxidant enzymes activities, haemolymph oxyhemocyanin contents and hepatopancreas histology of white shrimp, Litopenaeus vannamei, at various salinities. Aquaculture 2008, 274, 80–86. [Google Scholar] [CrossRef]
- Lavilla Pitogo, C.R.; Leano, E.M.; Paner, M.G. Mortalities of pond-cultured juvenile shrimp, Penaeus monodon, associated with dominance of luminescent Vibrios in the rearing environment. Aquaculture 1998, 164, 337–349. [Google Scholar] [CrossRef]
- Zhu, X.F.; Guo, H.; Li, G.L.; Zhu, C.H. Effects of dietary hydrolysable tannins on growth performance, antioxidant capacity, intestinal microflora and resistance against Vibrio parahaemolyticus of juvenile Pacific white shrimp, Litopenaeus vannamei (Boone, 1931). Aquac. Rep. 2021, 19, 100601. [Google Scholar] [CrossRef]
- Chen, S.J.; Zhuang, Z.X.; Yin, P.; Chen, X.; Zhang, Y.M.; Tain, L.X.; Niu, J.; Liu, Y.J. Changes in growth performance, haematological parameters, hepatopancreas histopathology and antioxidant status of pacific white shrimp (Litopenaeus vannamei) fed oxidized fish oil: Regulation by dietary myo-inositol. Fish Shellfish Immunol. 2019, 88, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Bachere, E.; Gueguen, Y.; Gonzalez, M.; De Lorgeril, J.; Garnier, J.; Romestand, B. Insights into the anti-microbial defense of marine invertebrates: The penaeid shrimps and the oyster Crassostrea gigas. Immunol. Rev. 2004, 198, 149–168. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Gu, Y.; Zhou, H.; Xu, L.; Cao, H.; Gai, C. Acinetobacter venetianus, a potential pathogen of red leg disease in freshwater-cultured whiteleg shrimp Penaeus vannamei. Aquac. Rep. 2020, 18, 100543. [Google Scholar] [CrossRef]
- Ott, L. Adhesion properties of toxigenic corynebacterial. AIMS Microbiol. 2018, 4, 85–103. [Google Scholar] [CrossRef]
- Shen, F.; Zheng, R.D.; Sun, X.Q.; Ding, W.J.; Wang, X.Y.; Fan, J.G. Gut microbiota dysbiosis in patients with non-alcoholic fatty liver disease. Hepatobillary Pancreat Dis. Int. 2017, 16, 375–381. [Google Scholar] [CrossRef]
- Sonnenburg, J.L.; Backhed, F. Diet-microbiota interactions as moderators of human metabolism. Nature 2016, 535, 56–64. [Google Scholar] [CrossRef]
- de Vadder, F.; Mithieux, G. Gut-brain signaling in energy homeostasis: The unexpected role of microbiota-derived succinate. J. Endocrinol. 2018, 236, R105–R108. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.N.J.; Jyothsna, R.; Reddy, M.H.; Sreevani, S.P.R. Effect of Bacillus subtilis and Lactobacillus rhamnosus incorporated probiotic diet on growth pattern and enzymes in Penaeus vannamei. Int. J. Life Sci. Pharm. Res. 2013, 3, 6–11. [Google Scholar]
- Ahmmed, F.; Ahmmed, M.K.; Shah, M.S.; Banu, G.R. Use of indigenous beneficial bacteria (Lactobacillus spp.) as probiotics in shrimp (Penaeus monodon) aquaculture. Res. Agric. Livest. Fish. 2018, 5, 127–135. [Google Scholar] [CrossRef][Green Version]
- Pooljun, C.; Daorueang, S.; Weerachatyanukul, W.; Direkbusarakom, S.; Jariyapong, P. Enhancement of shrimp health and immunity with diets supplemented with combined probiotics: Application to Vibrio parahaemolyticus infections. Dis. Aquac. Org. 2020, 140, 37–46. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).