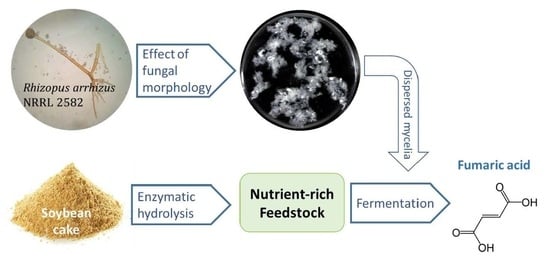
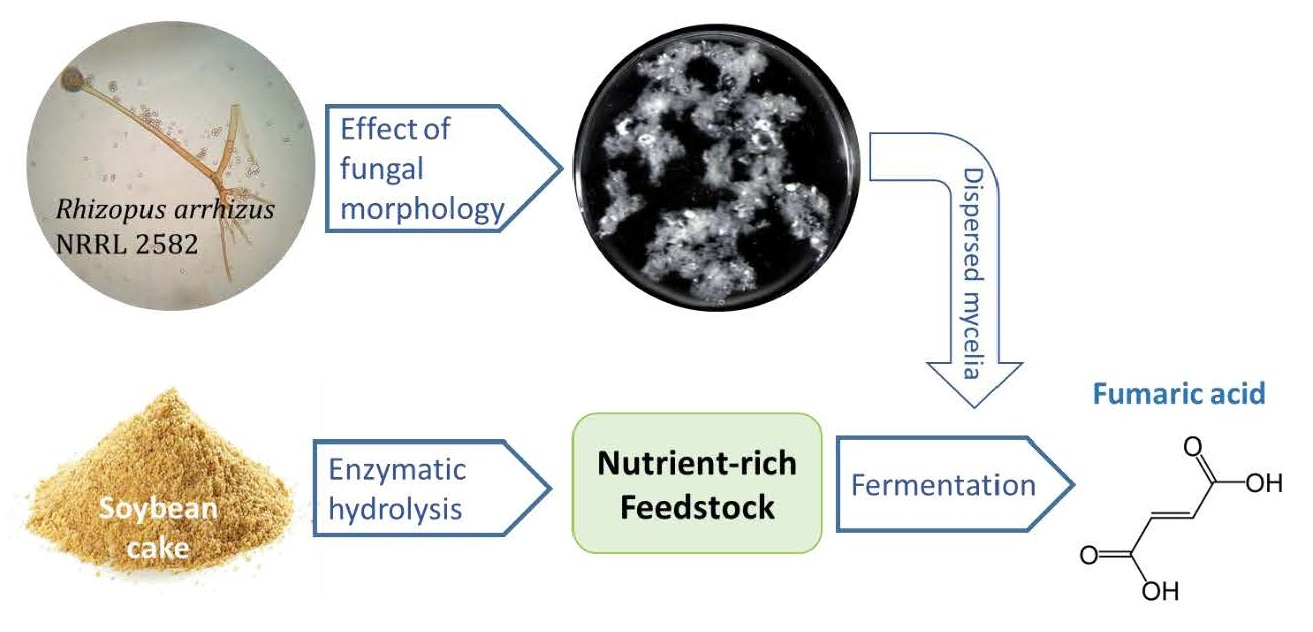
Biotechnological Production of Fumaric Acid: The Effect of Morphology of Rhizopus arrhizus NRRL 2582

 ,
, 
 ,
, 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Microorganisms and Growth Media
2.2. Raw Materials Used as Fermentation Media
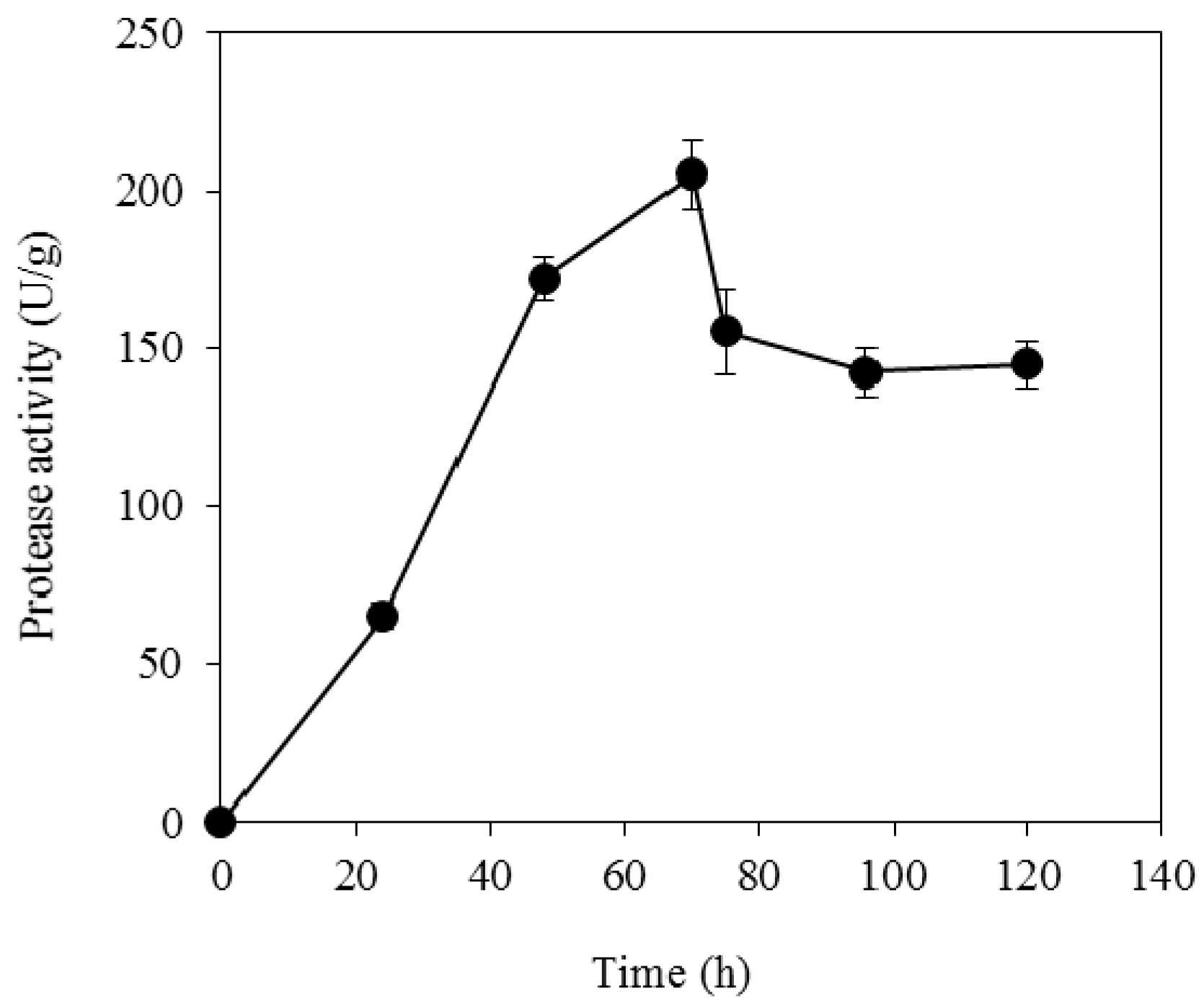
2.3. Crude Enzyme Production by Solid State Fermentation
2.4. Enzymatic Hydrolysis of Soybean Cake
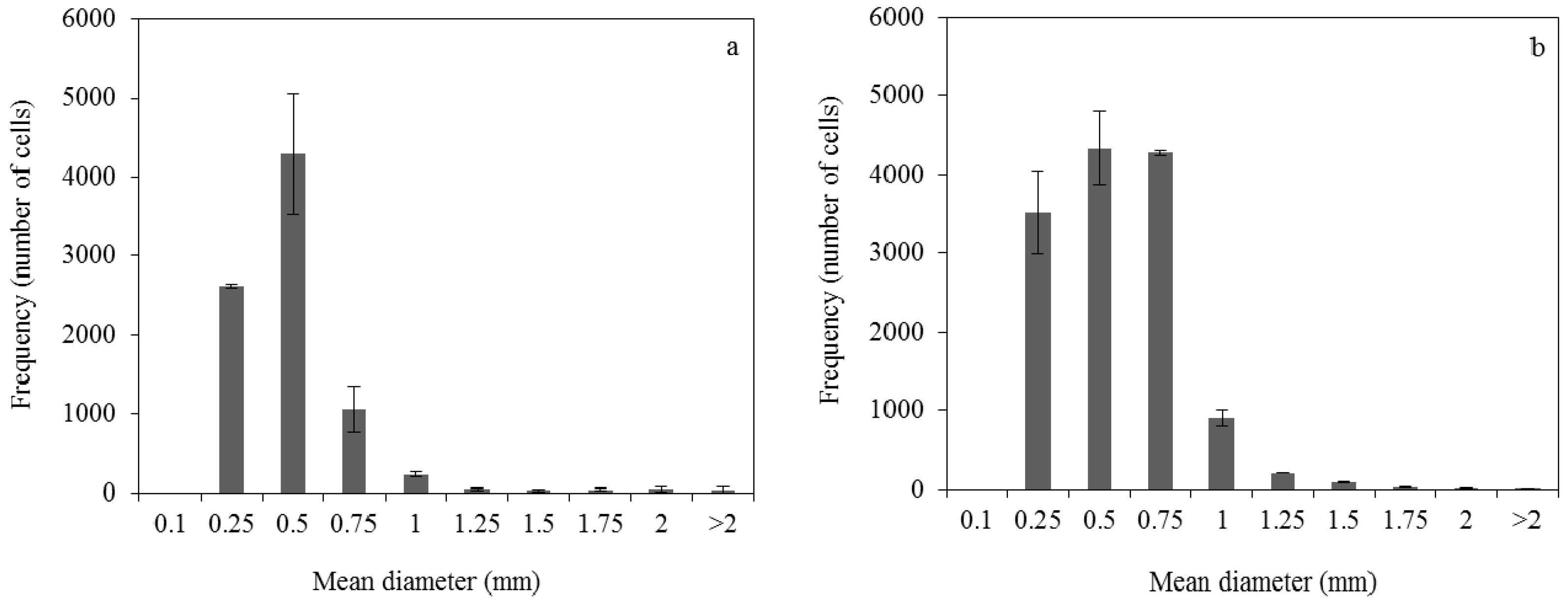
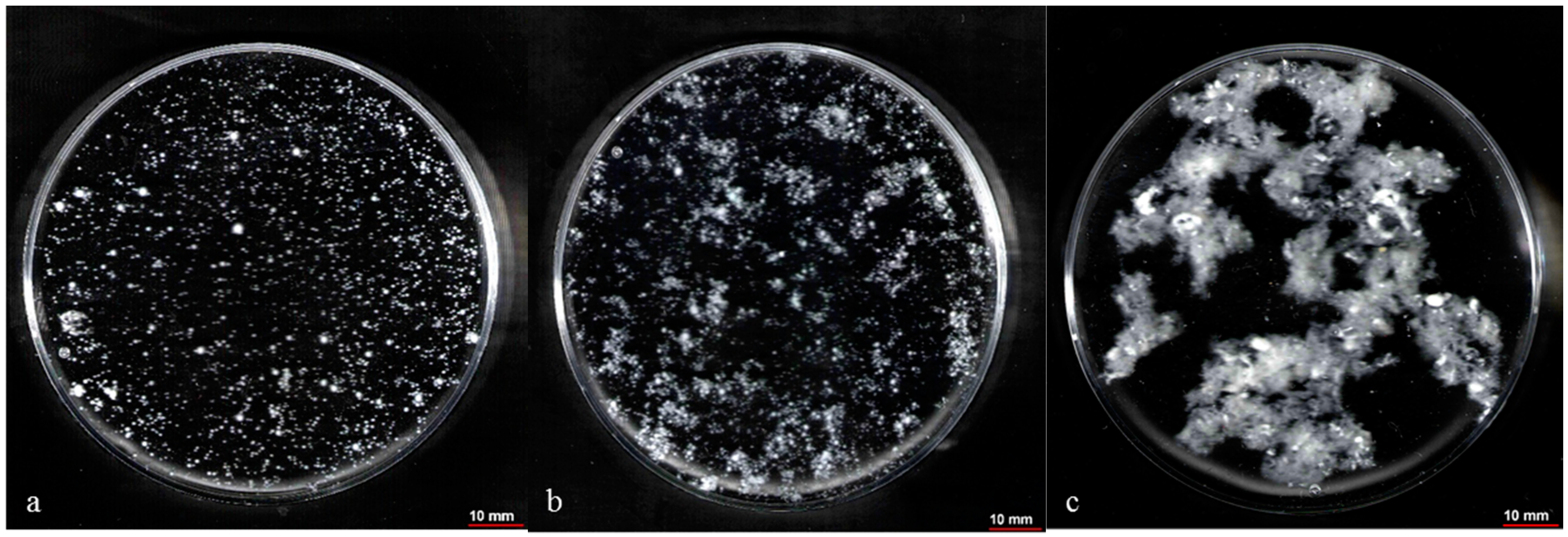
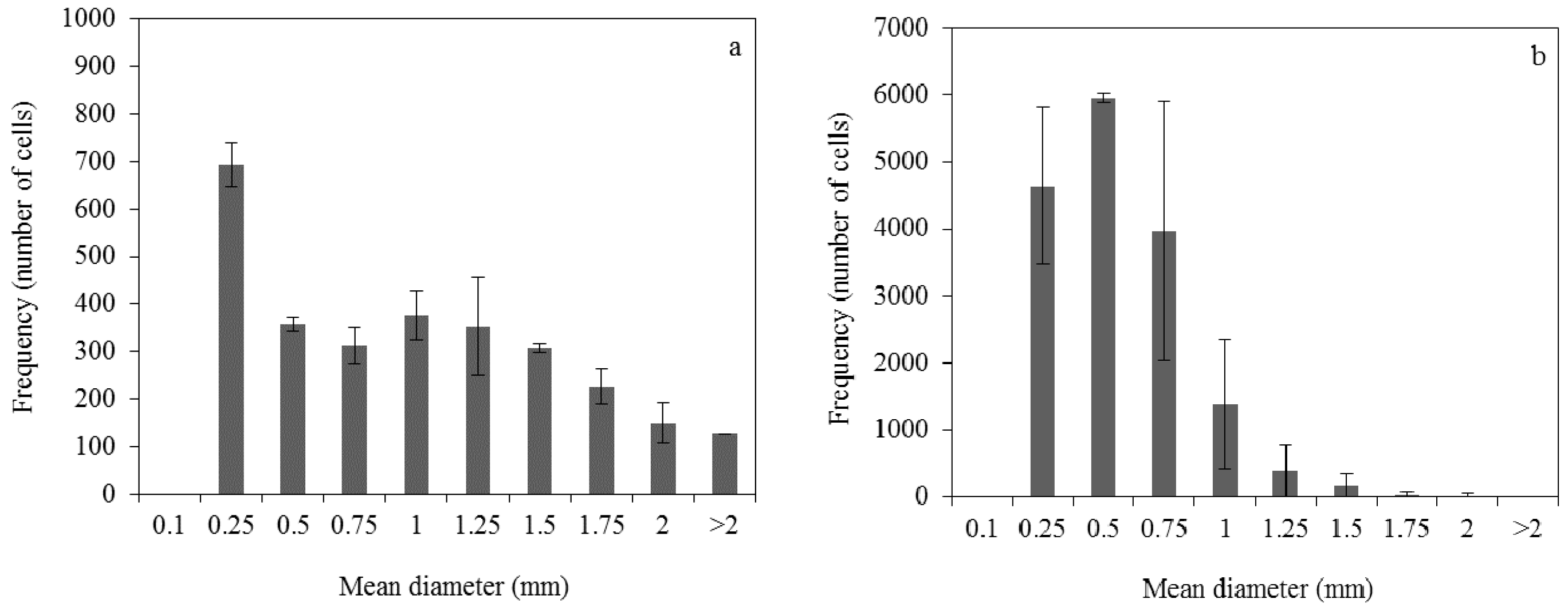
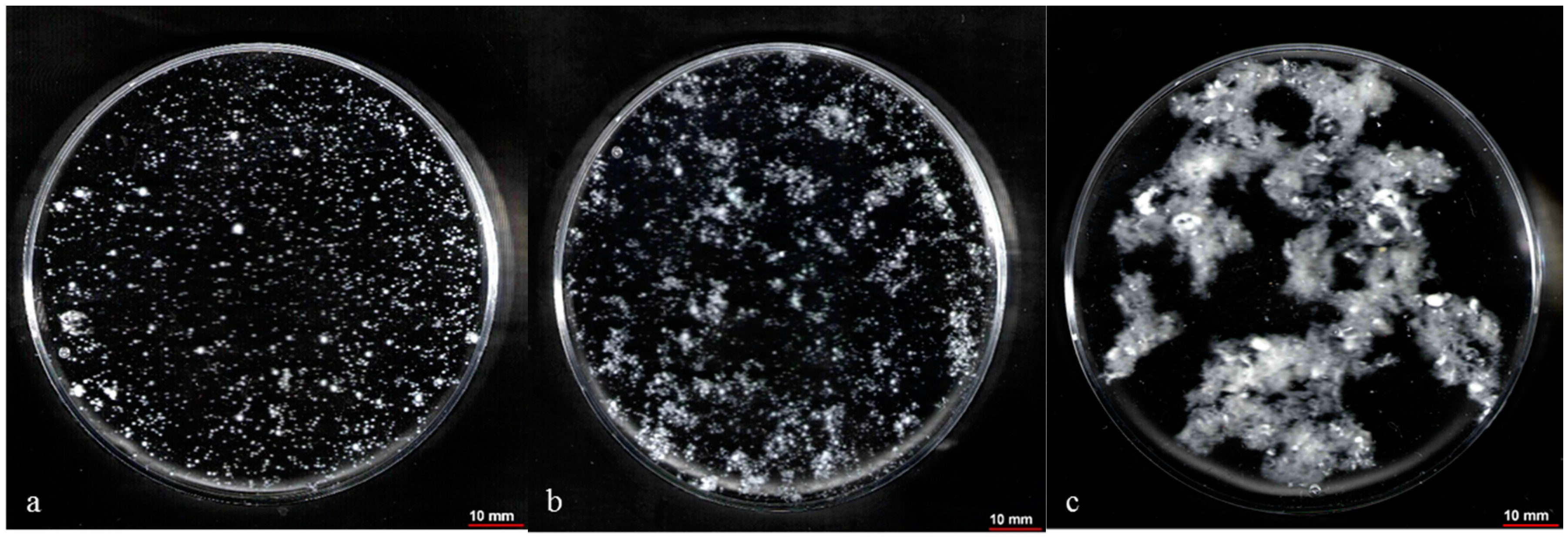
2.5. Control of Fungal Morphology
2.5.1. Production of Pelletized Biomass
2.5.2. Production of Dispersed Mycelia
2.6. Fumaric Acid Production
2.7. Fumaric Acid Production Using Soybean Cake Hydrolysate
2.8. Analytical Methods
3. Results and Discussion
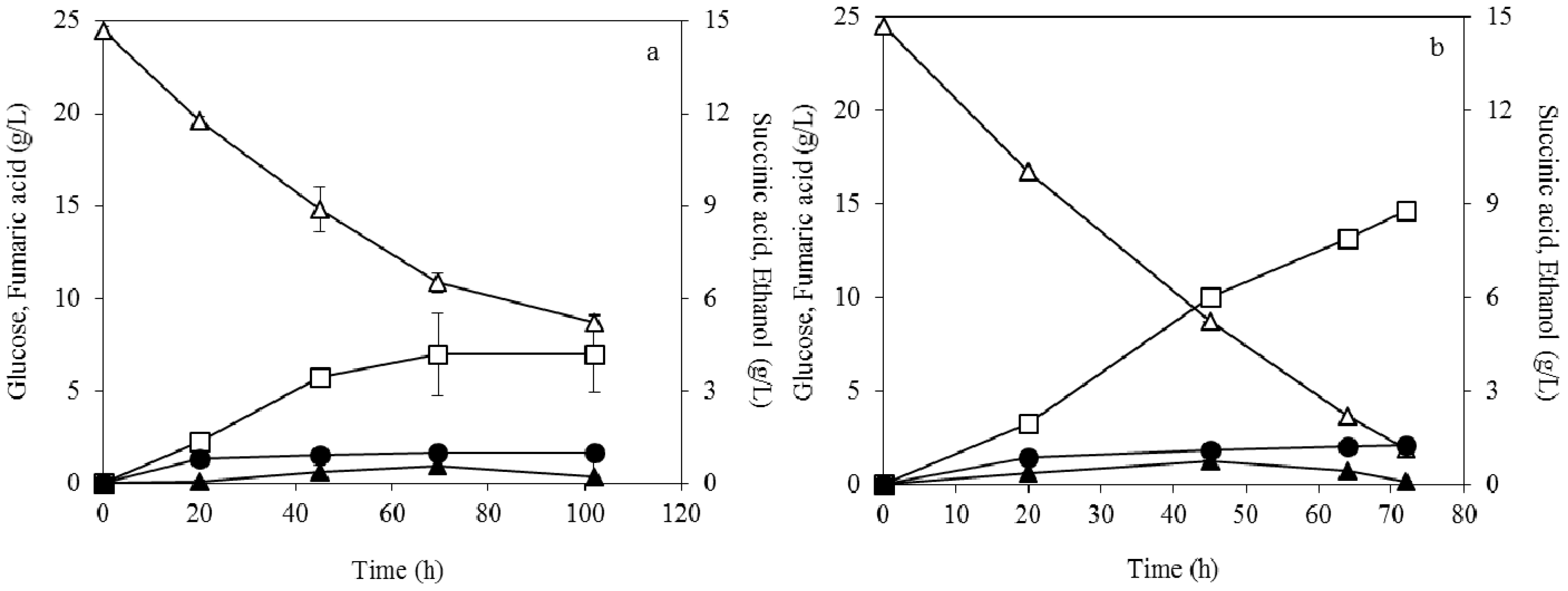
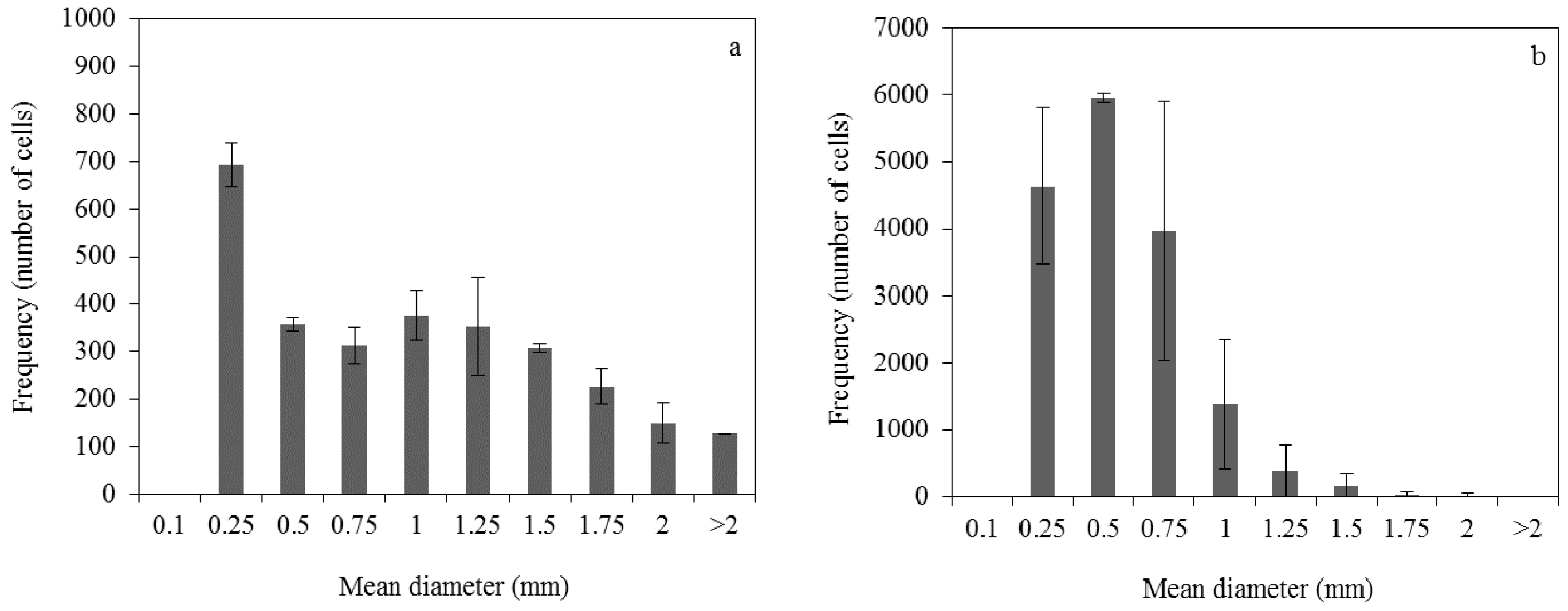
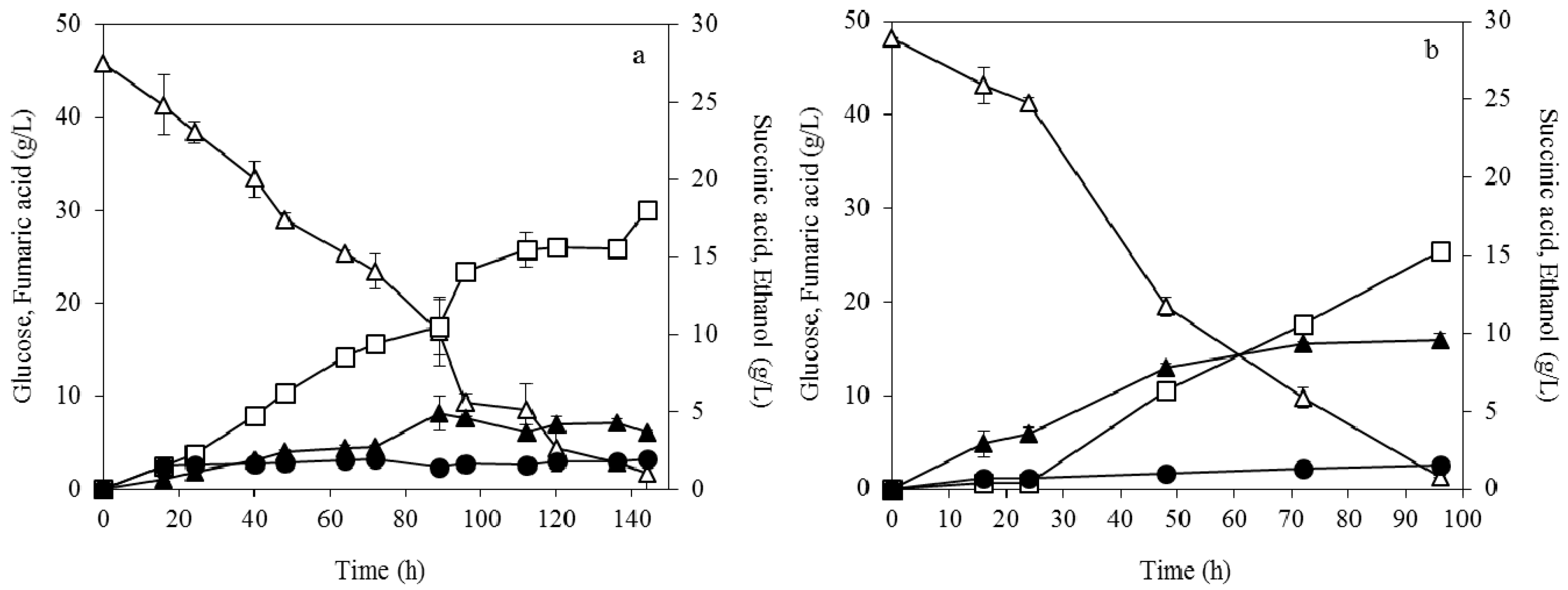
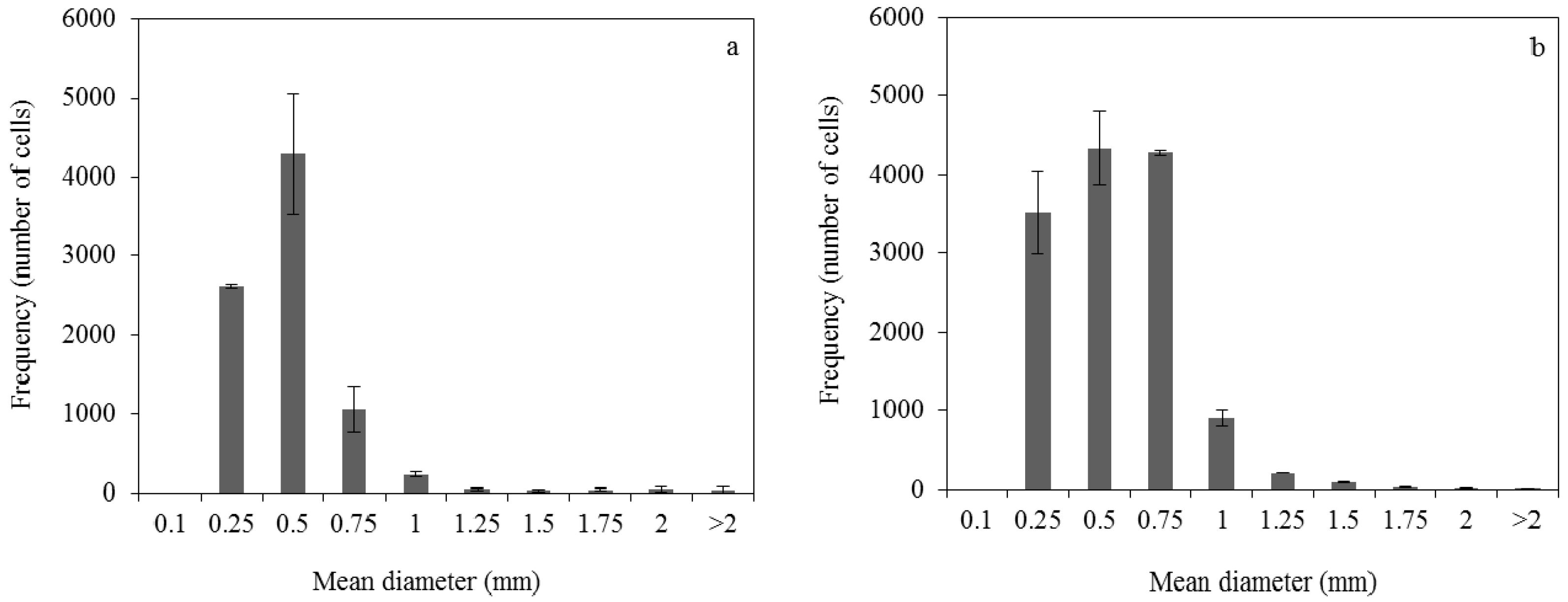
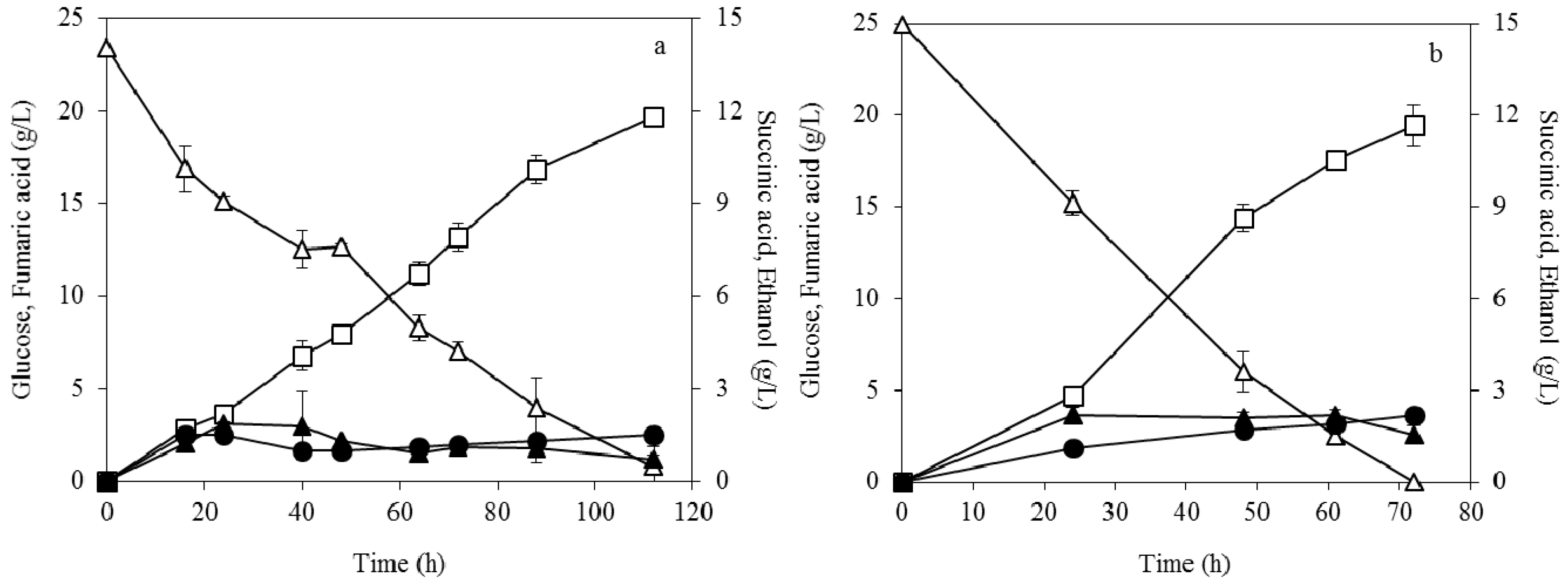
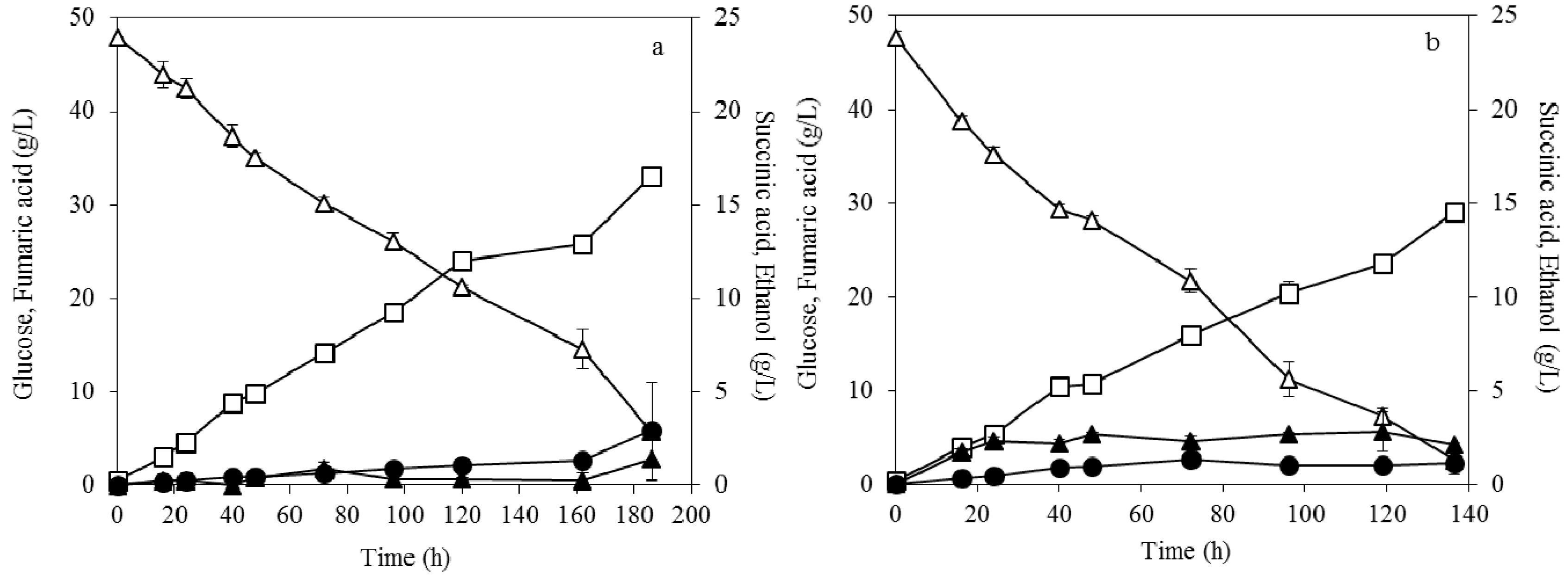
3.1. Effect of Pelletized Mycelia on Fumaric Acid Production
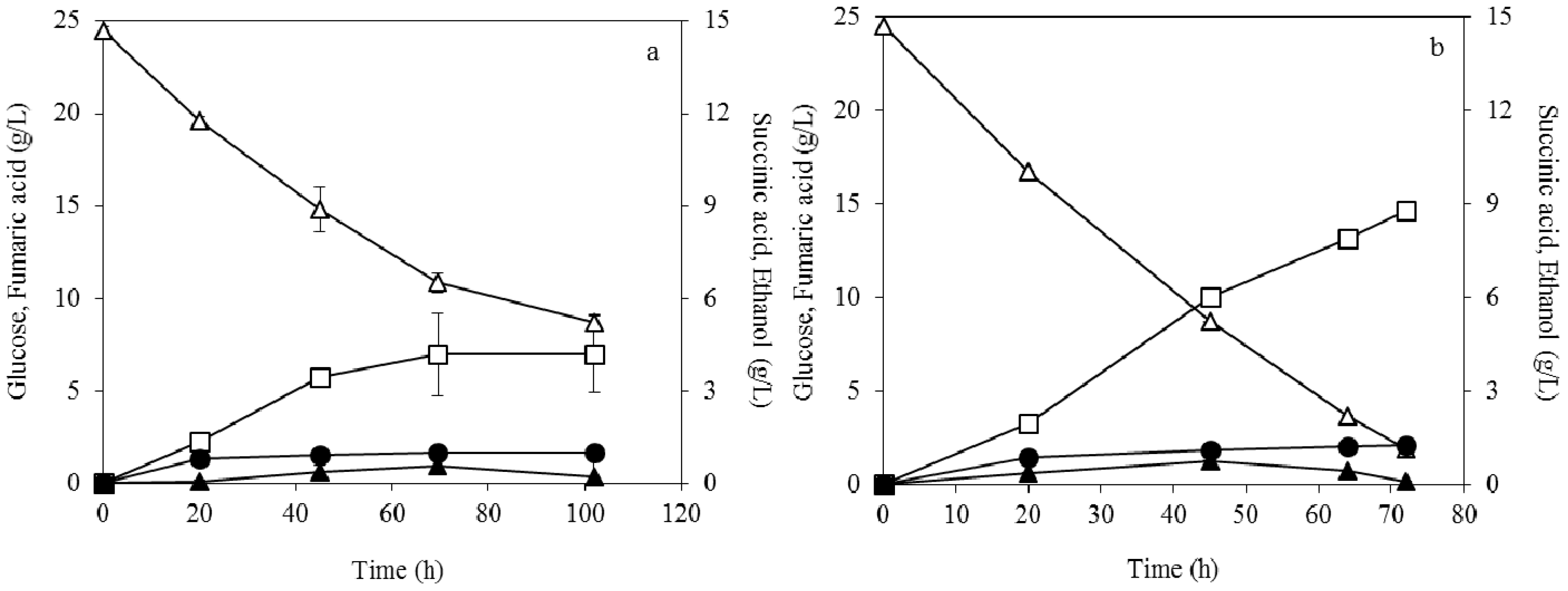
3.2. Effect of Dispersed Mycelia on Fumaric Acid Production
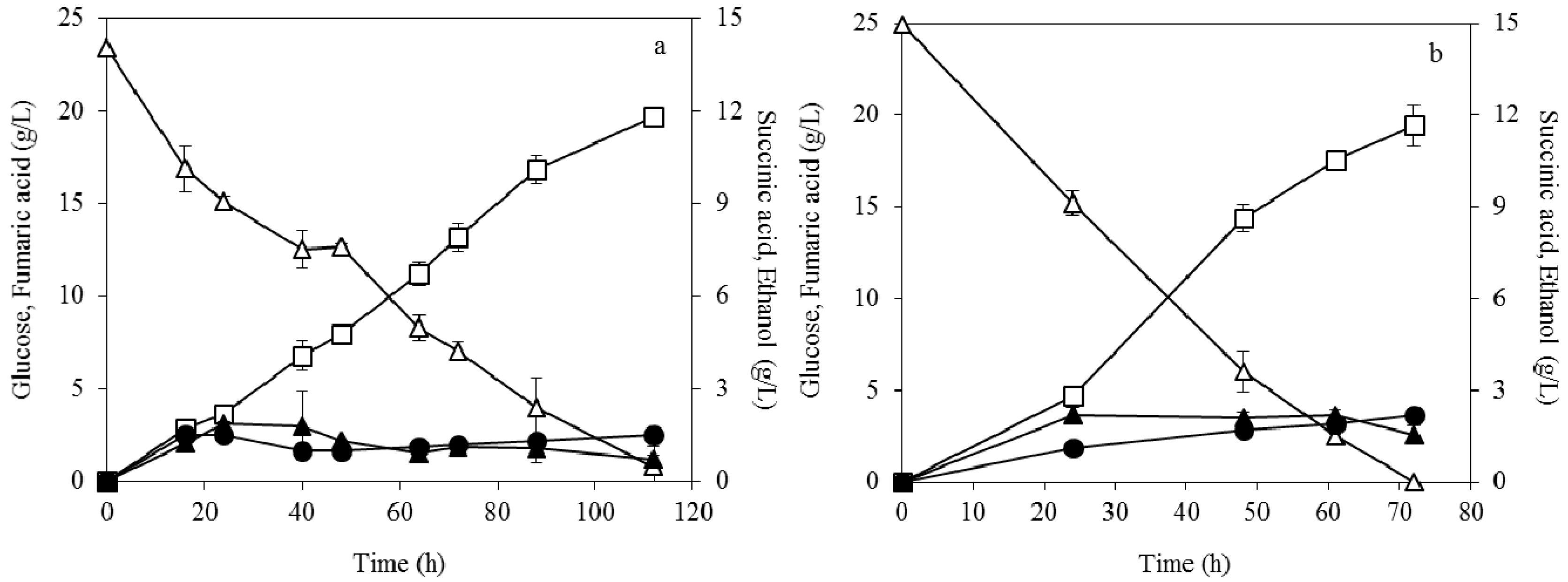
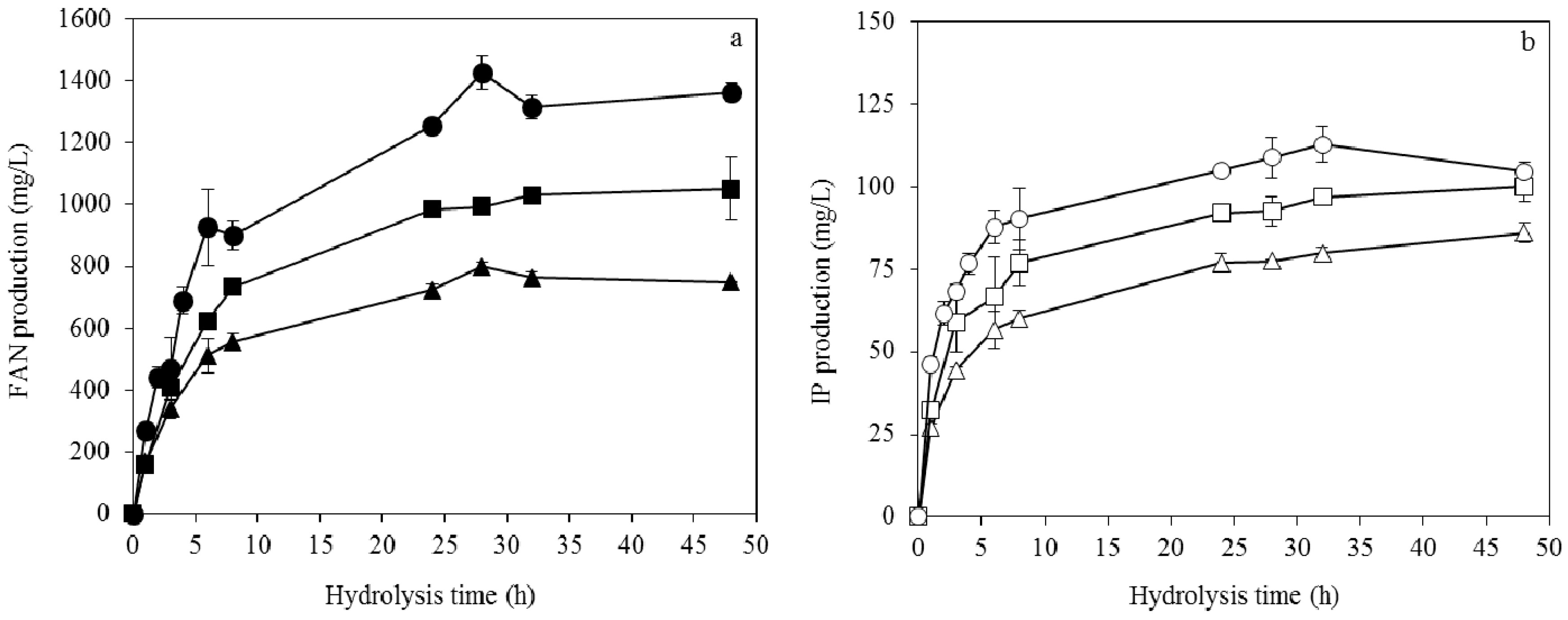
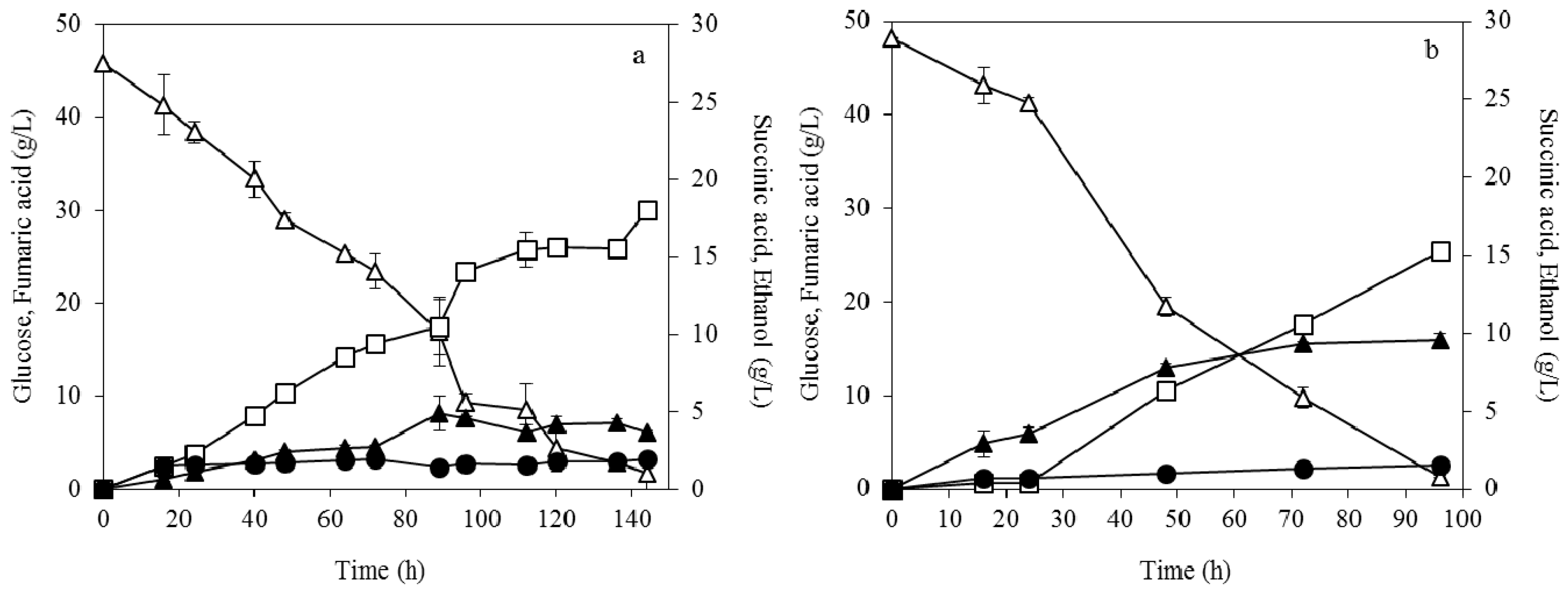
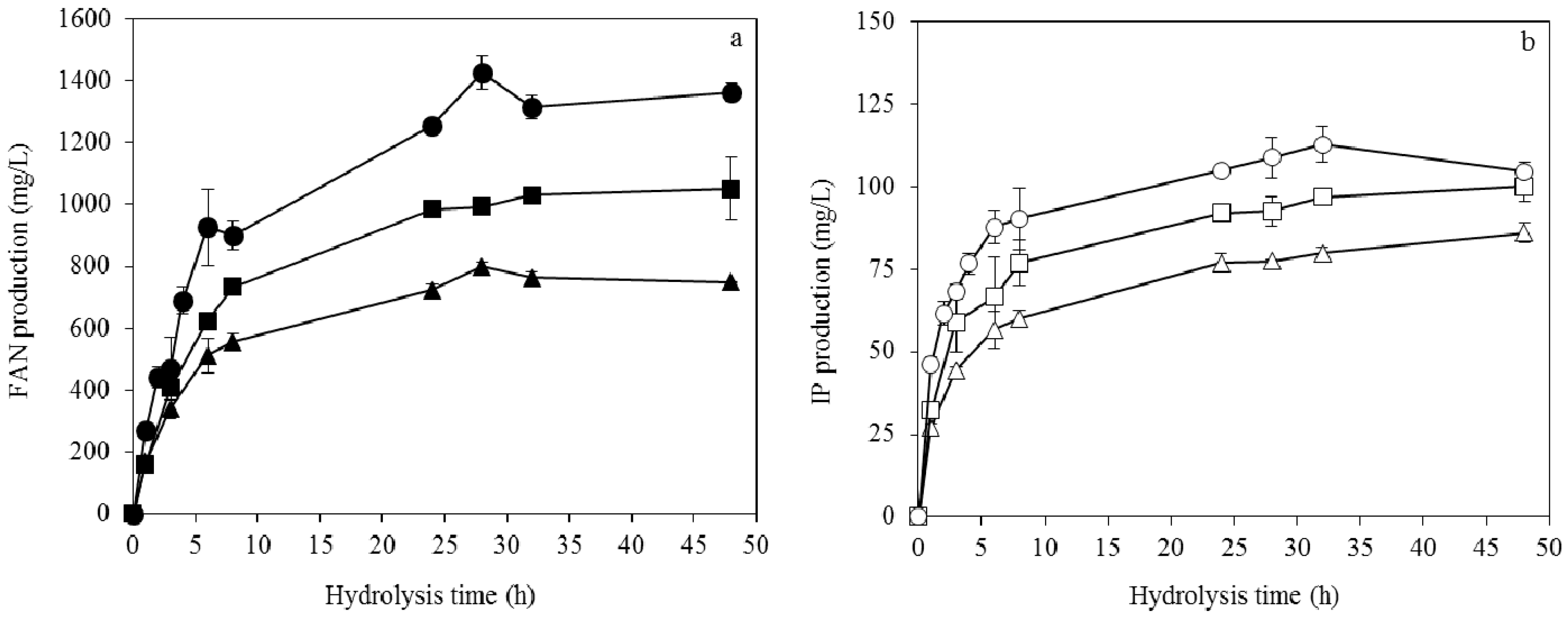
3.3. Enzymatic Hydrolysis of Soybean Cake
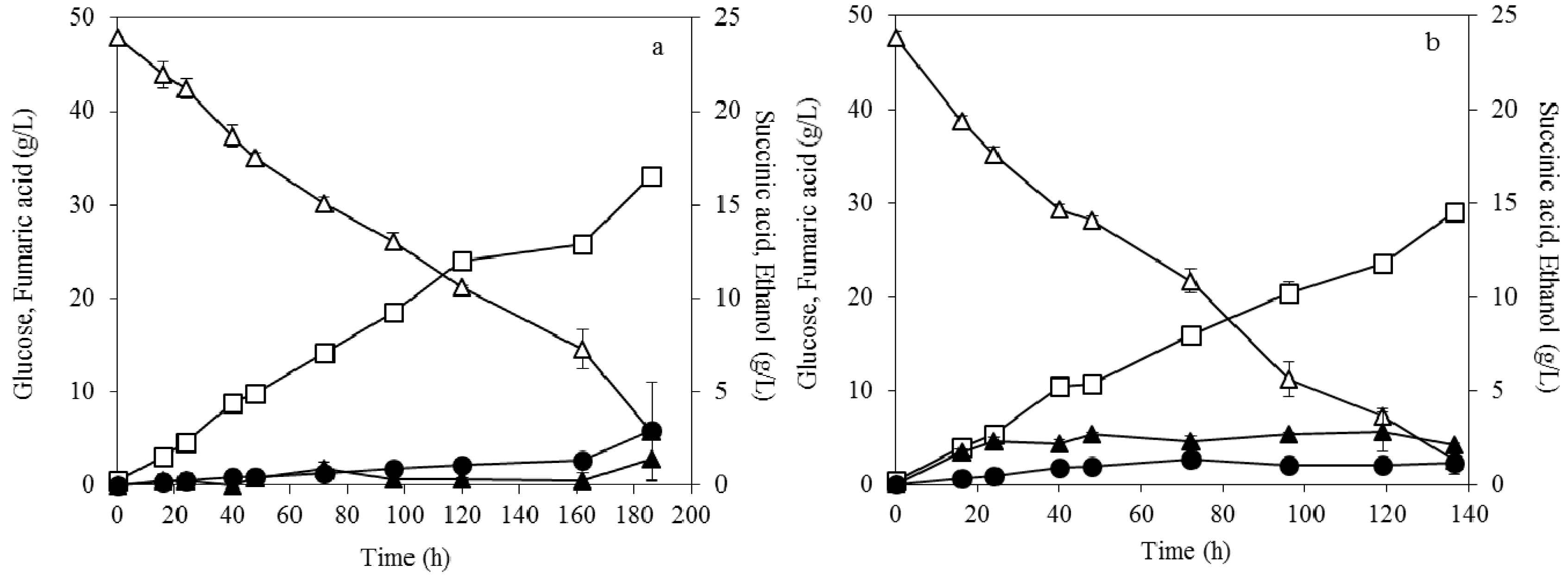
3.4. Fumaric Acid Production Using Soybean Cake Hydrolysate
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Koutinas, A.A.; Vlysidis, A.; Pleissner, D.; Kopsahelis, N.; Garcia, I.L.; Kookos, I.K.; Papanikolaou, S.; Kwan, T.H.; Lin, C.S. Valorization of industrial waste and by-product streams via fermentation for the production of chemicals and biopolymers. Chem. Soc. Rev. 2014, 43, 2587–2627. [Google Scholar] [CrossRef] [PubMed]
- Fumaric Acid Market Analysis by Application (Food & Beverages, Rosin Paper Sizes, UPR, Alkyd Resins) and Segment Forecasts to 2020. 2015. Available online: http://www.grandviewresearch.com/industry-analysis/fumaric-acid-market (accessed on 26 March 2017).
- Zhang, K.; Zhang, B.; Yang, S.-T. Production of citric, itaconic, fumaric and malic acids in filamentous fungal fermentation. In Bioprocessing Technologies in Biorefinery for Sustainable Production of Fuels, Chemicals, and Polymers, 1st ed.; Yang, S.-T., El-Enshasy, H.A., Thongchul, N., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2013. [Google Scholar]
- Roa Engel, C.A.; Straathof, A.J.J.; Zijlmans, T.W.; van Gulk, W.M.; van der Wielen, L.A.M. Fumaric acid production by fermentation. Appl. Microbiol. Biotechnol. 2008, 78, 379–389. [Google Scholar] [CrossRef] [PubMed]
- Magnuson, J.K.; Lasure, L.L. Organic acid production by filamentous fungi. In Advances in Fungal Biotechnology for Industry, Agriculture and Medicine; Tracz, J.S., Lange, L., Eds.; Kluwer/Plenum: New York, NY, USA, 2004; pp. 307–340. [Google Scholar]
- Rhodes, R.A.; Lagoda, A.A.; Jackson, R.W.; Misenhei, T.J.; Smith, M.L.; Anderson, R.F. Production of fumaric acid in 20 liter fermentors. Appl. Microbiol. 1962, 10, 9–15. [Google Scholar] [PubMed]
- Kenealy, W.; Zaady, E.; Dupreez, J.C.; Stieglitz, B.; Goldberg, I. Biochemical aspects of fumaric acid accumulation by Rhizopus arrhizus. Appl. Environ. Microbiol. 1986, 52, 128–133. [Google Scholar] [PubMed]
- Gangl, I.C.; Weigand, W.A.; Keller, F.A. Economic comparison of calcium fumarate and sodium fumarate production by Rhizopus arrhizus. Appl. Biochem. Biotechnol. 1990, 24–25, 663–677. [Google Scholar] [CrossRef]
- Petruccioli, M.; Angiani, E.; Federici, F. Semi continuous fumaric acid production by Rhizopus arrhizus immobilized in polyurethane sponge. Process Biochem. 1996, 31, 463–469. [Google Scholar] [CrossRef]
- Liao, W.; Liu, Y.; Frear, C.; Chen, S. A new approach of pellet formation of a filamentous fungus–Rhizopus oryzae. Bioresour. Technol. 2007, 98, 3415–3423. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Du, G.; Hua, Z.; Zhou, J.; Chen, J. Optimisation of fumaric acid production by Rhizopus delemar based on the morphology formation. Bioresour. Technol. 2011, 102, 9345–9349. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Yu, C.; Yang, S.-T. Effects of soybean meal hydrolysate as the nitrogen source on seed culture morphology and fumaric acid production by Rhizopus oryzae. Process Biochem. 2015, 50, 173–179. [Google Scholar] [CrossRef]
- Roa Engel, C.A.; van Gulk, W.M.; Marang, L.; van der Wielen, L.A.M.; Straathof, A.J.J. Development of low pH fermentation strategy for fumaric acid production by Rhizopus oryzae. Enzyme Microb. Technol. 2011, 48, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Byrne, G.S.; Ward, O.P. Effect of nutrition on pellet formation by Rhizopus arrhizus. Biotechnol. Bioeng. 1989, 33, 912–914. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Du, J.X.; Tsao, G.T. Mycelial pellet formation by Rhizopus oryzae ATCC 20344. Appl. Biochem. Biotechnol. 2000, 86, 779–789. [Google Scholar] [CrossRef]
- Das, R.K.; Brar, S.K. Enhanced fumaric acid production from brewery wastewater and insight into the morphology of Rhizopus oryzae 1526. Appl. Biochem. Biotechnol. 2014, 172, 2974–2988. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, R.A.; Moyer, A.J.; Smith, M.L.; Kelley, S.E. Production of fumaric acid by Rhizopus arrhizus. Appl. Microbiol. 1959, 7, 74–80. [Google Scholar] [PubMed]
- Kachrimanidou, V.; Kopsahelis, N.; Chatzifragkou, A.; Papanikolaou, S.; Yanniotis, S.; Kookos, I.; Koutinas, A.A. Utilisation of by-products from sunflower-based biodiesel production processes for the production of fermentation feedstock. Waste Biomass Valoriz. 2013, 4, 529–537. [Google Scholar] [CrossRef]
- Wang, R.; Law, R.C.S.; Webb, C. Protease production and conidiation by Aspergillus oryzae in flour fermentation. Proc. Biochem. 2005, 40, 217–227. [Google Scholar] [CrossRef]
- Dimou, C.; Kopsahelis, N.; Papadaki, A.; Papanikolaou, S.; Kookos, I.K.; Mandala, I.; Koutinas, A.A. Wine lees valorization: Biorefinery development including production of a generic fermentation feedstock employed for poly(3-hydroxybutyrate) synthesis. Food Res. Int. 2015, 73, 81–87. [Google Scholar] [CrossRef]
- American Association of Cereal Chemists Inc. (AACC). Approved Methods of the American Association of Cereal Chemists, 8th ed.; American Association of Cereal Chemists Inc.: St. Paul, MN, USA, 1983. [Google Scholar]
- Association of Official Analytical Chemists (AOAC). Official Method 973.18, Fiber (Acid Detergent) and Lignin in Animal Feed, 15th ed.; Association of Official Analytical Chemists, Inc.: Arlington, VA, USA, 1990. [Google Scholar]
- Lie, S. The EBC-ninhydrin method for determination of free alpha amino nitrogen. J. Inst. Brew. 1973, 79, 37–41. [Google Scholar] [CrossRef]
- Harland, B.F.; Harland, J. Fermentative reduction of phytate in rye, white and whole wheat breads. Cereal Chem. 1980, 57, 226–229. [Google Scholar]
- Goldberg, I.; Lonberg-Holm, K.; Bagley, E.A.; Stieglitz, B. Improved conversion of fumarate to succinate Escherichia coli strains amplified for fumarate reductase. Appl. Environ. Microbiol. 1983, 1838–1847. [Google Scholar]
- Ng, T.K.; Hesser, R.J.; Stieglitz, B.; Griffiths, B.S.; Ling, L.B. Production of tetrahydrofuran/1,4-butanediol by a combined biological and chemical process. Biotechnol. Bioeng. Symp. 1986, 17, 344–363. [Google Scholar]
- Tsouko, E.; Kachrimanidou, V.; Dos Santos, A.F.; do Nascimento Vitorino Lima, M.E.; Papanikolaou, S.; de Castro, A.M.; Freire, D.M.; Koutinas, A.A. Valorization of By-Products from Palm Oil Mills for the Production of Generic Fermentation Media for Microbial Oil Synthesis. Appl. Biochem. Biotechnol. 2016, 181, 1241–1256. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, S.; Sikander, A.; Haq, I. Acidic pre-treatment of sugarcane molasses for molasses for citric acid production by Aspergillus niger NG-4. Int. J. Curr. Microbiol. Appl. Sci. 2015, 4, 584–595. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Constituents | Pellets 1 | Dispersed Clumps 2 |
|---|---|---|
| Glucose (g/L) | 10 | 25 |
| Peptone (g/L) | 2 | 1.6 |
| Urea (g/L) | – | 1 |
| (NH4)2SO4 (g/L) | – | – |
| KH2PO4 (g/L) | – | 0.6 |
| MgSO4·7H2O (g/L) | – | 0.4 |
| ZnSO4·7H2O (g/L) | – | 0.044 |
| FeCl3·6H2O (g/L) | – | 0.016 |
| Tartaric acid (g/L) | – | – |
| Methanol (mL/L) | – | – |
| Corn steep liquor (mL/L) | – | 3 |
| Corn starch (g/L) | – | 30 |
| Agar (g/L) | – | 1 |
| Composition | % (Dry Basis) |
|---|---|
| Moisture (%, wet basis) | 13 ± 0.4 |
| Protein (TKN × 6.25) | 47 ± 1.2 |
| Ash | 6.5 ± 0.09 |
| Lipids | 2.2 ± 0.2 |
| Cellulose | 24.1 ± 0.9 |
| Hemicellulose | 18.1 ± 1.6 |
| Lignin | 2.1 ± 0.08 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Papadaki, A.; Androutsopoulos, N.; Patsalou, M.; Koutinas, M.; Kopsahelis, N.; Castro, A.M.d.; Papanikolaou, S.; Koutinas, A.A. Biotechnological Production of Fumaric Acid: The Effect of Morphology of Rhizopus arrhizus NRRL 2582. Fermentation 2017, 3, 33. https://doi.org/10.3390/fermentation3030033
Papadaki A, Androutsopoulos N, Patsalou M, Koutinas M, Kopsahelis N, Castro AMd, Papanikolaou S, Koutinas AA. Biotechnological Production of Fumaric Acid: The Effect of Morphology of Rhizopus arrhizus NRRL 2582. Fermentation. 2017; 3(3):33. https://doi.org/10.3390/fermentation3030033
Chicago/Turabian StylePapadaki, Aikaterini, Nikolaos Androutsopoulos, Maria Patsalou, Michalis Koutinas, Nikolaos Kopsahelis, Aline Machado de Castro, Seraphim Papanikolaou, and Apostolis A. Koutinas. 2017. "Biotechnological Production of Fumaric Acid: The Effect of Morphology of Rhizopus arrhizus NRRL 2582" Fermentation 3, no. 3: 33. https://doi.org/10.3390/fermentation3030033
APA StylePapadaki, A., Androutsopoulos, N., Patsalou, M., Koutinas, M., Kopsahelis, N., Castro, A. M. d., Papanikolaou, S., & Koutinas, A. A. (2017). Biotechnological Production of Fumaric Acid: The Effect of Morphology of Rhizopus arrhizus NRRL 2582. Fermentation, 3(3), 33. https://doi.org/10.3390/fermentation3030033