Direct Succinic Acid Production from Minimally Pretreated Biomass Using Sequential Solid-State and Slurry Fermentation with Mixed Fungal Cultures

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Substrates
2.2. Fungi
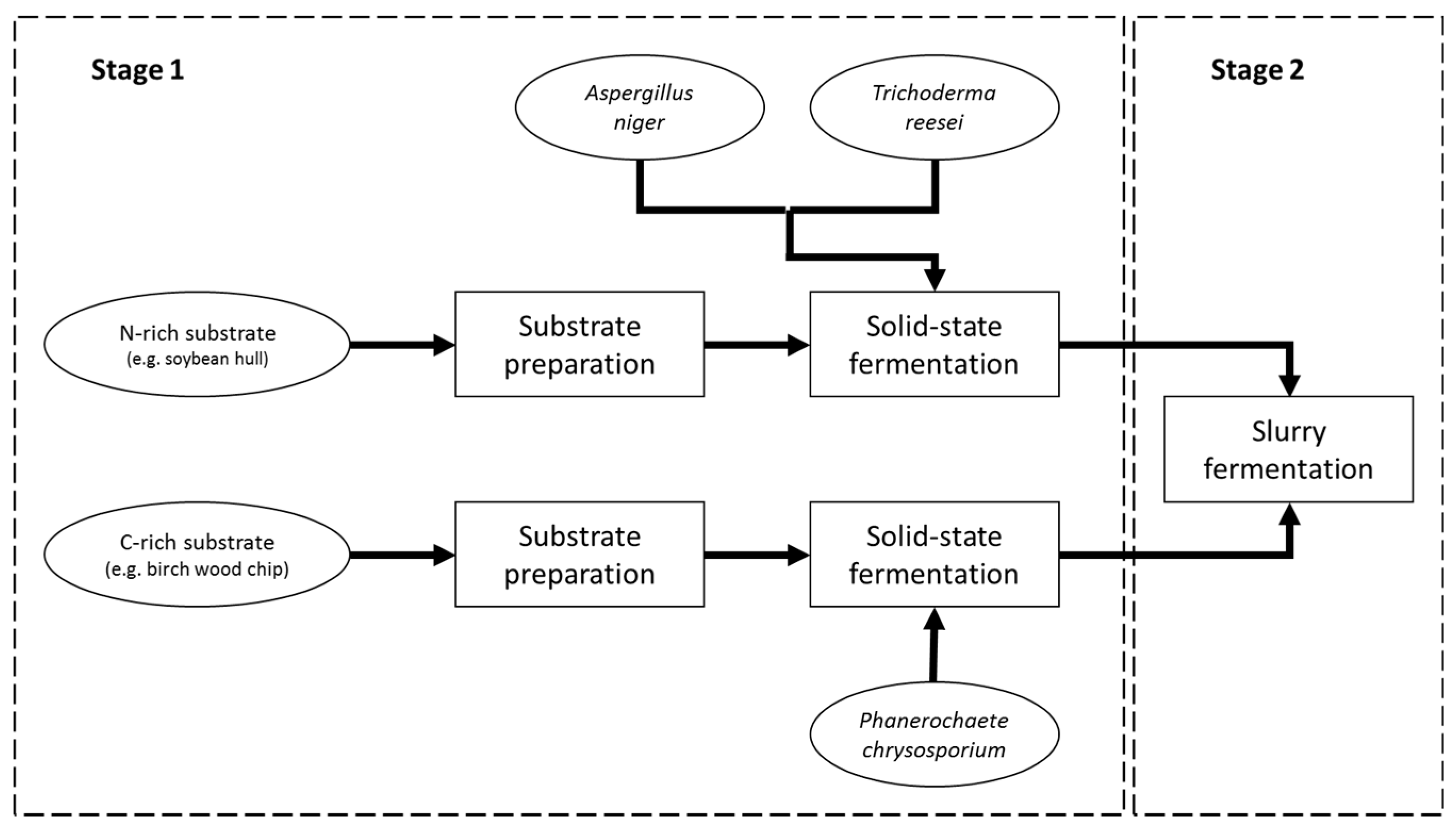
2.3. Solid-State Pre-Fermentation Stage
2.4. Slurry Fermentation Stage
2.5. Analyses
3. Results and Discussion
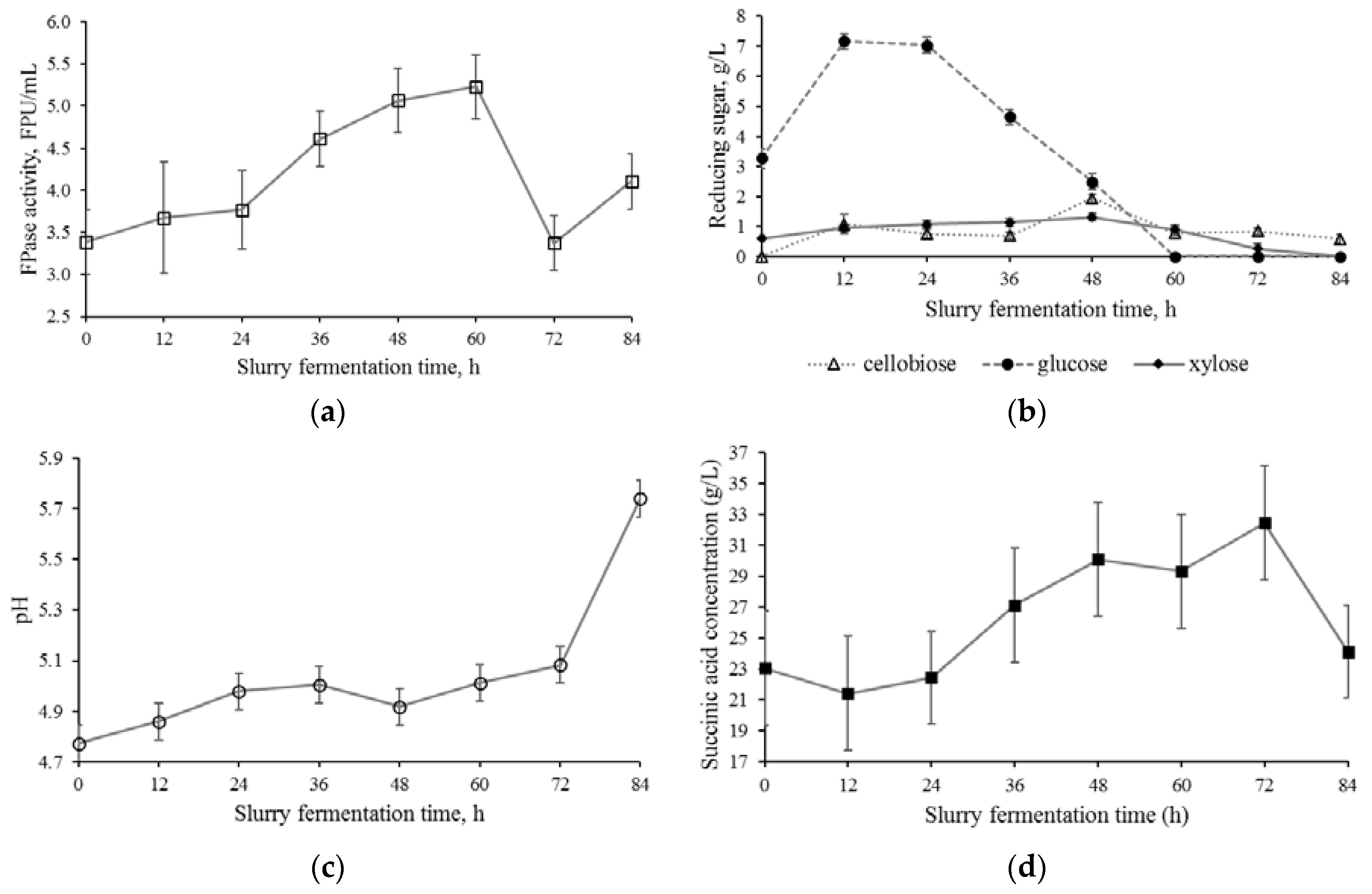
3.1. Solid-State Pre-Fermentation Stage
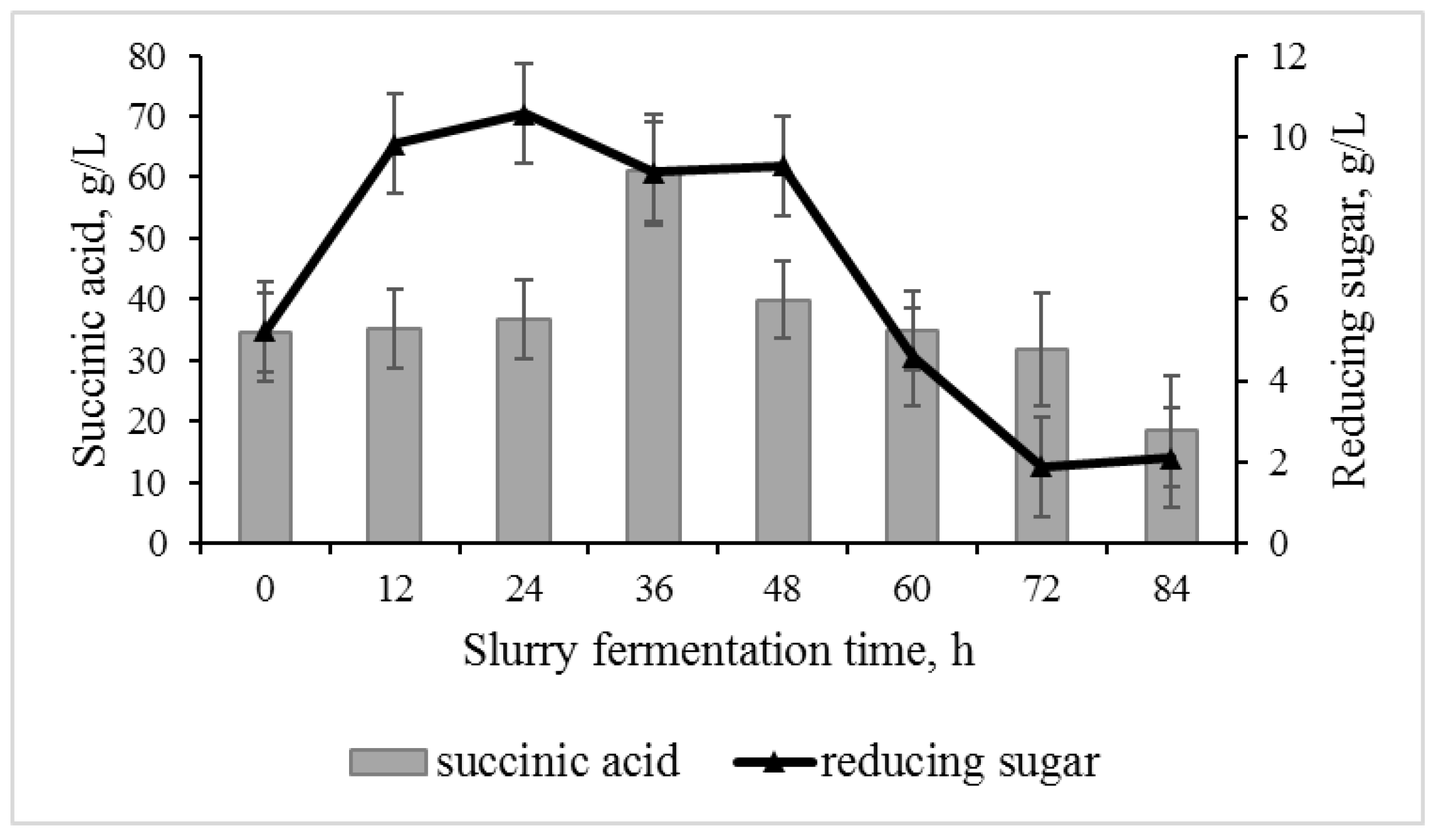
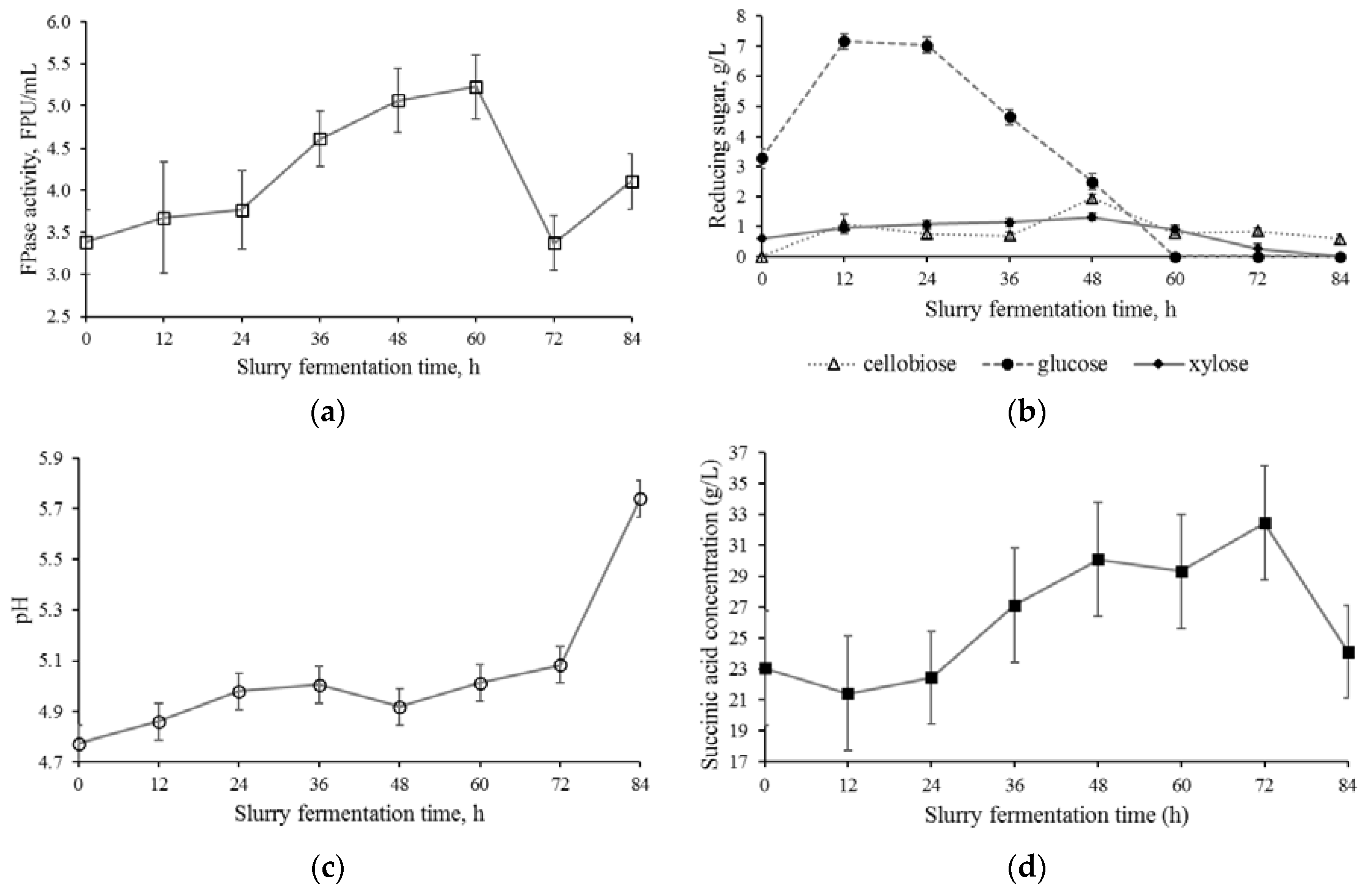
3.2. Slurry Fermentation Stage
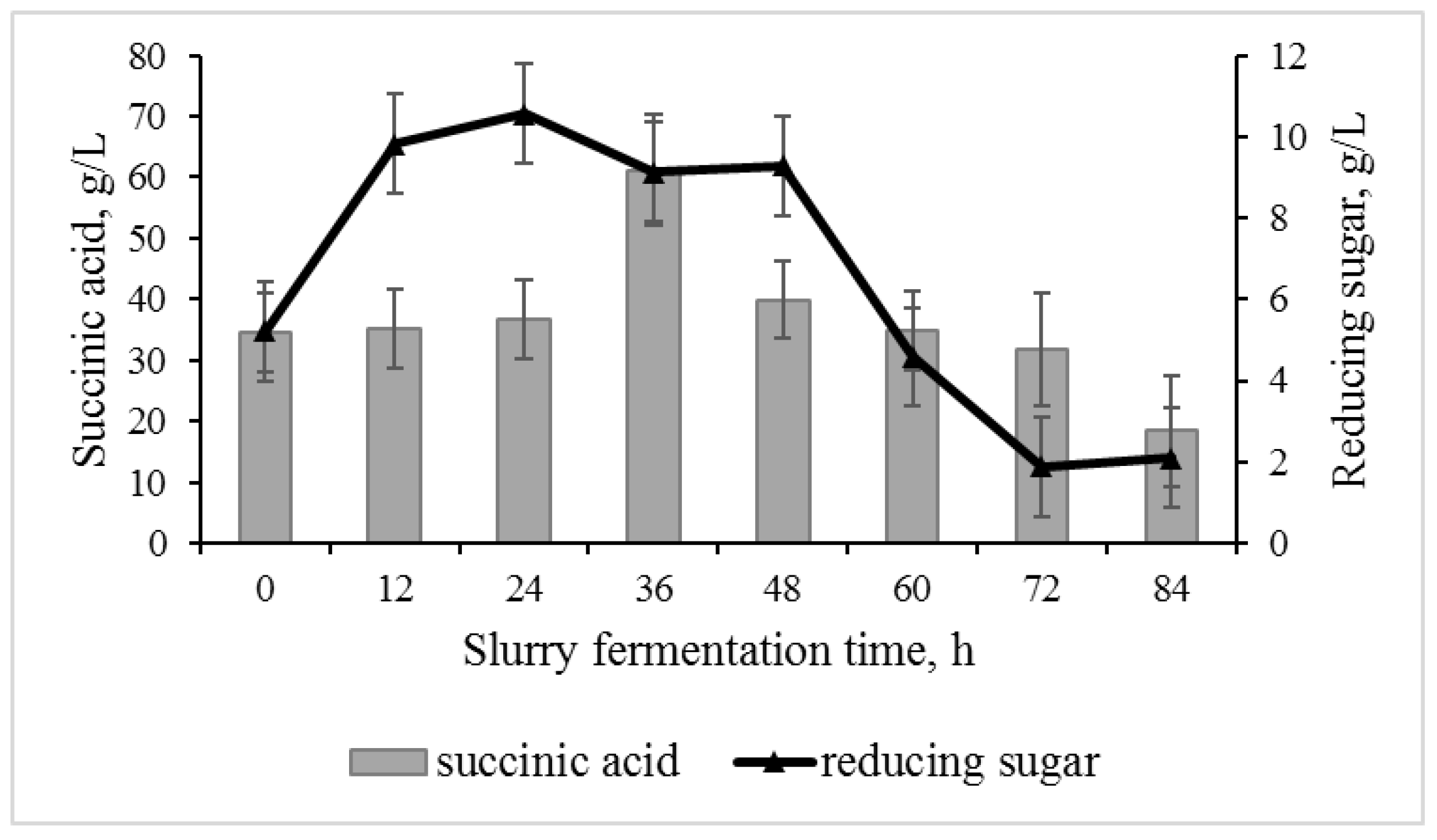
3.3. Effect of Adding Fresh Substrate
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Werpy, P.; Petersen, G. Top Value Added Chemicals From Biomass: Volume I—Results of sCreening Potential Candidates from Sugars And Synthesis Gas; US Department of Energy: Washington, DC, USA, 2004.
- Zeikus, J.G.; Jain, M.K.; Elankovan, P. Biotechnology of succinic acid production and markets for derived industrial products. Appl. Microbiol. Biotechnol. 1999, 51, 545–552. [Google Scholar] [CrossRef]
- Cornils, B.; Lappe, P. Dicarboxylic acids, aliphatic. In Ullmann’s Encyclopedia of Industrial Chemistry; Wiley-VCA: Weinheim, Germany, 2010. [Google Scholar] [CrossRef]
- Song, H.; Lee, S.Y. Production of succinic acid by bacterial fermentation. Enzym. Microb. Technol. 2006, 39, 352–361. [Google Scholar] [CrossRef]
- Thakker, C.; Martinez, I.; San, K.Y.; Bennett, G.N. Succinate production in Escherichia coli. Biotechnol. J. 2012, 7, 213–224. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Correa, M.; Portal, L.; Moreno, P.; Tengerdy, R.P. Mixed culture solid substrate fermentation of Trichoderma reesei with Aspergillus niger on sugar cane bagasse. Bioresour. Technol. 1999, 68, 173–178. [Google Scholar] [CrossRef]
- Dhillon, G.S.; Oberoi, H.S.; Kaur, S.; Bansal, S.; Brar, S.K. Value-addition of agricultural wastes for augmented cellulase and xylanase production through solid-state tray fermentation employing mixed-culture of fungi. Ind. Crops Prod. 2011, 34, 1160–1167. [Google Scholar] [CrossRef]
- Fang, H.; Zhao, C.; Song, X.; Chen, M.; Chang, Z.; Chu, J. Enhanced cellulolytic enzyme production by the synergism between Trichoderma reesei RUT-C30 and Aspergillus niger NL02 and by the addition of surfactants. Biotechnol. Bioprocess Eng. 2003, 18, 390–398. [Google Scholar] [CrossRef]
- Lu, J.; Weerasiri, R.R.; Liu, Y.; Wang, W.; Ji, S.; Lee, I. Enzyme production by the mixed fungal culture with nano-shear pretreated biomass and lignocellulose hydrolysis. Biotechnol. Bioeng. 2013, 110, 2123–2130. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Chinn, M.S.; Sharma-Shivappa, R.R. Microbial pretreatment of cotton stalks by solid state cultivation of Phanerochaete chrysosporium. Bioresour. Technol. 2008, 99, 6556–6564. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; Singh, D.; Chen, S. Biological pretreatment of wheat straw by Phanerochaete chrysosporium supplemented with inorganic salts. Bioresour. Technol. 2011, 102, 3206–3214. [Google Scholar] [CrossRef] [PubMed]
- Bercovitz, A.; Peleg, Y.; Battat, E.; Rokem, J.S.; Goldberg, I. Localization of pyruvate carboxylase in organic acid producing Aspergillus strains. Appl. Environ. Microbiol. 1990, 56, 1594–1597. [Google Scholar] [PubMed]
- Ikram-ul, H.; Ali, S.; Qadeer, M.A.; Iqbal, J. Citric acid production by selected mutants of Aspergillus niger from cane molasses. Bioresour. Technol. 2004, 93, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Mielenz, J.R.; Bardsley, J.S.; Wyman, C.E. Fermentation of soybean hulls to ethanol while preserving protein value. Bioresour. Technol. 2009, 100, 3532–3539. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.Z. Chemical degradation. In Wood and Cellulosic Chemistry; Hon, D.N.-S., Shiraishi, N., Eds.; Marcel Dekker Inc.: New York, NY, USA, 1991; pp. 455–524. [Google Scholar]
- Tangnu, S.K.; Blanch, H.W.; Wilke, C.R. Enhanced production of cellulase, hemicellulose, and β-glucosidase by Trichoderma reesei (Rut C-30). Biotechnol. Bioeng. 1981, 23, 1837–1849. [Google Scholar] [CrossRef]
- Ghose, T.K. Measurement of cellulase activities. Pure Appl. Chem. 1987, 59, 257–268. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- McMillan, J.D. Pretreatment of lignocellulosic biomass. In Enzymatic Conversion of Biomass for Fuels Production; Himmel, M.E., Baker, J.O., Overend, R.P., Eds.; American Chemical Society: Washington, DC, USA, 1994; Volume 566, pp. 292–324. [Google Scholar]
- Mosier, N.; Wyman, C.; Dale, B.; Elander, R.; Lee, Y.Y.; Holtzapple, M.; Ladisch, M. Features of promising technologies for pretreatment of lignocellulosic biomass. Bioresour. Technol. 2005, 96, 673–686. [Google Scholar] [CrossRef] [PubMed]
- Keller, F.A.; Hamilton, J.E.; Nguyen, Q.A. Microbial pretreatment of biomass: potential for reducing severity of thermochemical biomass pretreatment. Appl. Biochem. Biotechnol. 2003, 105, 27–41. [Google Scholar] [CrossRef]
- Duff, S.J.B.; Murray, W.D. Bioconversion of forest products industry waste cellulosics to fuel ethanol: A review. Bioresour. Technol. 1996, 55, 1–33. [Google Scholar] [CrossRef]
- Lee, P.C.; Lee, S.Y.; Hong, S.H.; Chang, H.N.; Park, S.C. Biological conversion of wood hydrolysate to succinic acid by Anaerobiospirillum succiniciproducens. Biotechnol. Lett. 2003, 25, 111–114. [Google Scholar] [CrossRef] [PubMed]
- Borges, E.R.; Pereira, N., Jr. Succinic acid production from sugarcane bagasse hemicellulose hydrolysate by Actinobacillus succinogenes. J. Ind. Microbiol. Biotechnol. 2011, 38, 1001–1011. [Google Scholar] [CrossRef] [PubMed]
- Magnuson, J.K.; Lasure, L.L. Organic acid production by filamentous Fungi. In Advances in Fungal Biotechnology for Industry, Agriculture, and Medicine; Tkacz, J.S., Lange, L., Eds.; Kluwer Academic/Plenum Publishers: New York, NY, USA, 2004; pp. 307–340. [Google Scholar]

© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alcantara, J.; Mondala, A.; Hughey, L.; Shields, S. Direct Succinic Acid Production from Minimally Pretreated Biomass Using Sequential Solid-State and Slurry Fermentation with Mixed Fungal Cultures. Fermentation 2017, 3, 30. https://doi.org/10.3390/fermentation3030030
Alcantara J, Mondala A, Hughey L, Shields S. Direct Succinic Acid Production from Minimally Pretreated Biomass Using Sequential Solid-State and Slurry Fermentation with Mixed Fungal Cultures. Fermentation. 2017; 3(3):30. https://doi.org/10.3390/fermentation3030030
Chicago/Turabian StyleAlcantara, Jerico, Andro Mondala, Logan Hughey, and Shaun Shields. 2017. "Direct Succinic Acid Production from Minimally Pretreated Biomass Using Sequential Solid-State and Slurry Fermentation with Mixed Fungal Cultures" Fermentation 3, no. 3: 30. https://doi.org/10.3390/fermentation3030030
APA StyleAlcantara, J., Mondala, A., Hughey, L., & Shields, S. (2017). Direct Succinic Acid Production from Minimally Pretreated Biomass Using Sequential Solid-State and Slurry Fermentation with Mixed Fungal Cultures. Fermentation, 3(3), 30. https://doi.org/10.3390/fermentation3030030