Abstract
Kinetic models for bioethanol production from waste sorghum leaves by Saccharomyces cerevisiae BY4743 are presented. Fermentation processes were carried out at varied initial glucose concentrations (12.5–30.0 g/L). Experimental data on cell growth and substrate utilisation fit the Monod kinetic model with a coefficient of determination (R2) of 0.95. A maximum specific growth rate (μmax) and Monod constant (KS) of 0.176 h−1 and 10.11 g/L, respectively, were obtained. The bioethanol production data fit the modified Gompertz model with an R2 value of 0.98. A maximum bioethanol production rate (rp,m) of 0.52 g/L/h, maximum potential bioethanol concentration (Pm) of 17.15 g/L, and a bioethanol production lag time (tL) of 6.31 h were observed. The obtained Monod and modified Gompertz coefficients indicated that waste sorghum leaves can serve as an efficient substrate for bioethanol production. These models with high accuracy are suitable for the scale-up development of bioethanol production from lignocellulosic feedstocks such as sorghum leaves.
1. Introduction
Ideal crops for commercial bioethanol production in South Africa include maize, grain sorghum, and sugar cane [1]; however, in order to completely utilise these materials, post-harvest field waste should be employed for biofuel production. Sweet sorghum (Sorghum bicolor (L.) Moench), in particular, yields significant amounts of biomass (leaves and pressed stalks) and sugar (found in stalks) [2]. Bioconversion of lignocellulosic material to renewable fuels is currently receiving great interest since it does not impact food security [2]. Several studies on the enhancement of fermentable sugar release from lignocellulosic substrates have been reported [3,4,5]. Microwave-assisted pre-treatment has received increased attention due to its lower energy demand and shorter process times [6]. Microwave radiation alters the structure of lignocellulose by emitting electromagnetic radiation, which results in the formation of thermal pockets. These pockets ultimately explode due to an increase in heat, leading to the relocation of crystalline structures within the lignocellulosic material [6]. Gabhane et al. [7] studied the individual and interactive effects of acid and alkali pre-treatments using an autoclave, microwave, and ultrasonicator, and obtained a maximal reducing sugar yield of 36.84% from acid pre-treated banana waste by using microwave radiation. Despite the vast information available on lignocellulosic pre-treatment, a significant knowledge gap exists between this and the kinetic assessment of the fermentation efficiency of pre-treated lignocellulosic substrates for biofuel production.
Bioethanol is one such fuel which exhibits several advantages over conventional fossil fuels. This includes its renewable nature, ease of storage, higher oxygen content, higher octane number, the fact that it is free of sulfur, and contributes less to global warming and air pollution [8,9]. In recent times, the application of bioethanol as a fuel replacement has become more appealing [9]. Subsequent to lignocellulosic pre-treatment, the fermentable sugars released are converted to bioethanol by the exploitation of microbial metabolism, with the simultaneous release of carbon dioxide. Numerous studies have focused on the production of bioethanol from sorghum [10,11,12]. Suryaningsih and Irhas [10] obtained an optimal ethanol concentration of 40 g/L after 48 h of fermentation using sorghum grain, while an ethanol content of 12.4 g/L was obtained by Massoud et al. [11] by using the lignocellulosic hydrolysate of pressed sorghum stalks. However, very few have determined the efficiency of bioethanol production from sorghum leaves alone. Globally, efforts are being made to further expedite the use of renewable fuel sources as an alternative. These efforts are being challenged by a significant increase in the cost of production [13]. This suggests that further modelling and optimisation studies are required for the development of biofuel from lignocellulosic substrates.
Kinetic modelling refers to a mathematical description of the changes in the properties of a system in which biochemical reactions take place [14]. These models assist in the design of a production process by representing the complex biochemistry of cells. Kinetic models can be used to understand, predict, and evaluate the effects of altering the components of a fermentation process [15]. Most commonly, these models are used to increase yield and productivity as well as minimise the formation of undesired by-products, ensuring that the product is of high quality [15]. Models capable of describing the kinetics of microbial growth, substrate utilisation, and product formation play a fundamental role in process optimisation and control [16] by providing a basis for process design, control, and scale-up [17].
Monod kinetics models are commonly used to describe biomass growth and product formation with respect to the limiting substrate [18], while the modified Gompertz models are used to determine production lag time, maximum production rate, and maximum product concentration on a given substrate [8,19]. The original Gompertz function has been applied in a wide range of research areas, such as ecology, marketing, actuarial sciences, medicine, and biology [20]. Although the modified Gompertz equation has been used in many studies for ethanol and hydrogen production [19,21,22], very few studies have reported on bioethanol fermentation kinetics using lignocellulosic biomass as a feedstock [2,17,18]. These studies include feedstocks such as populus hydrolysate [17], sweet sorghum stalks [2], and rice hulls [18]. Despite this, there is a scarcity of information regarding the fermentation kinetics of this fuel using waste sorghum leaves.
Knowledge from fermentation kinetic studies on waste sorghum leaves will provide fundamental information on process characteristics and behaviour. Furthermore, decisions involving process control and improvement can be made with relative ease when a bioprocess is fully understood, advancing its commercial application. In this study, the Monod and modified Gompertz models were used to assess the kinetic behaviour of a bioethanol fermentation process (in a batch system) using waste sorghum leaves.
2. Materials and Methods
2.1. Feedstock Preparation and Pre-Treatment
Sorghum leaves used in this study were harvested from Ukulinga Research Farm, Pietermaritzburg, South Africa (29°67′ E, 30°40′ S). Approximately five to eight sorghum leaves were cut off at the leaf collar of mature (approximately 100–120 days) plants. They were immediately oven-dried at 70 °C for 48 h and milled to particle sizes of 1–2 mm using a centrifugal miller (Retsch ZM-1, Durban, South Africa). Milled leaves were treated under previously optimised conditions [23]; i.e., a 3.83% (v/v) HCl (Merck, Durban, South Africa) solution at a solid-to-liquid (S:L) ratio of 16.66% for 2 min at 600 W in a 1000 W capacity microwave oven (Samsung, Model: ME9114S1, Durban, South Africa).
2.2. Enzymatic Hydrolysis
Pre-treated biomass was rinsed with distilled water until a pH of 4.0 was achieved. The biomass was then oven-dried at 60 °C overnight and enzymatically hydrolysed using powdered cellulase enzyme, Onozuka R-10 (Merck, Durban, South Africa) in 500 mL Erlenmeyer flasks. A solid loading rate of 20 g dry biomass in 200 mL 0.05 M citrate buffer, with an enzyme loading rate of 50 mg/g of dry biomass was employed. The pH during enzymatic hydrolysis was 4.8, and the temperature was maintained at 50 °C using a water bath at 120 rpm for 72 h. The hydrolysate was filtered, and the filtrate was analysed for glucose concentration.
2.3. Fermentation Medium Formulation
A mineral salt solution (pH 4.5) containing (in g/L); yeast extract, 1.0; (NH4)2SO4, 2.0 and MgSO4, 1.0 was autoclaved at 121 °C for 15 min. All reagents purchased from Merck (Durban, South Africa). Filter-sterilised enzymatic hydrolysate was then added to the mineral salts. Initial glucose concentrations within a range of 12.5–30.0 g/L were obtained by diluting or—where needed—supplementing with pure glucose.
2.4. Microorganism and Inoculum Preparation
The S. cerevisiae BY4743 used in this study was obtained from the Department of Genetics, University of KwaZulu-Natal, Pietermaritzburg, South Africa. A single flask containing 100 mL Yeast-Peptone-Dextrose (YPD) medium was inoculated with a single colony and grown at 150 rpm, 30 °C overnight, until the exponential growth phase was reached. This culture was inoculated (10%) into the prepared fermentation medium (working volume of 100 mL) containing an initial glucose concentration of 12.5 g/L. The culture was then grown under the same conditions as previously described and then used as a starter culture for subsequent fermentation processes.
2.5. Fermentation Process and Analytical Methods
Fermentation processes were carried out in sterilised 250 mL flasks with a working volume of 100 mL. Aliquots of 10 mL (10% inoculation) S. cerevisiae were aseptically added to the fermentation flasks, and the cultures were incubated at 30 °C at 120 rpm for 24 h, or until glucose concentrations were depleted. Fermentations were aseptically sampled every two h and assessed for biomass concentration, sugar content, and bioethanol content.
The sugar content of filtered enzymatic hydrolysate and fermentation media was determined using a YSI 2700 Model Biochemical Analyser (YSI, Yellow Springs, OH, USA). Ethanol content was determined in the gas phase of the fermentation process, using an ethanol vapour sensor (ETH-BTA, Vernier Software and Technology, Beaverton, OR, USA). The absorbance of the culture broth was measured using a spectrophotometer (UV-Vis Spectrophotometer, UVmini-1240, Shimadzu, Kyoto, Japan) at 650 nm. Cell biomass quantification was achieved by using absorbance as a function of the concentration of yeast cells. A standard curve was prepared by determining the dry weights and corresponding absorbance values of yeast biomass at varied dilutions of a 24 h S. cerevisiae culture, grown in fermentation media containing 12.5 g/L glucose. Dry weights were determined by centrifuging 5 mL of each dilution (1, 1/2, 1/4, 1/8, and 1/10) for 10 min at 5000 rpm. The supernatant was removed, and the remaining biomass was dried at 60 °C until a constant mass was obtained.
2.6. Calculations of Kinetic Model Constants
The average specific growth rates (μ) of fermentation processes carried out in duplicate were calculated using Equation (1). The specific growth rate values (μ) and the substrate concentration data were subsequently used to estimate the maximum specific growth rate (μmax) and Monod constant (KS) by the double reciprocal Lineweaver–Burk plot. The Lineweaver–Burk plot has the possibility of distorting the error structure of the data, but it is still used for representation of kinetic data because the double reciprocal plot usually automatically and conveniently provides a considerably improved weighting for linear graphs of most kinetic parameters as a function of substrate concentration [24].
where X2 and X1 are biomass concentrations (g/L) at time instants t2 and t1, respectively.
The linear form of this equation is as follows:
where S represents substrate concentration. In addition, experimental data on bioethanol production over time were used to fit the modified Gompertz model (Equation (3)) using the least squares method, (CurveExpert V1.5.5), which showed the lag time, maximum bioethanol production rate, and the potential maximum product concentration.
where P is bioethanol concentration (g/L), Pm is potential maximum bioethanol concentration (g/L), rp,m is maximum bioethanol production rate (g/L/h), and tL is the time from the beginning of fermentation to exponential bioethanol production (h).
Sugar utilisation, ethanol yield, ethanol productivity, and fermentation efficiency were calculated using the following Equations (4)–(7) respectively [25]:
3. Results and Discussion
3.1. Monod Kinetic Model of S. cerevisiae on Waste Sorghum Leaves
The microwave-assisted acid pre-treatment and subsequent enzymatic hydrolysis of the biomass of sorghum leaves resulted in a glucose yield of 0.153 g/g sorghum leaves. Cell biomass, bioethanol production, and glucose consumption were monitored throughout the fermentation process. The correlation between absorbance and dry weight of yeast biomass was determined by linear regression, which gave a correlation coefficient (r) of 0.96. The specific growth rate (μ) values were calculated using the exponential (log) phase of microbial growth. The values obtained were 0.096, 0.104, 0.114, 0.122, and 0.123 h−1 at initial substrate concentrations of 12.5, 13.3, 19.4, 21.8, and 23.1 g/L, respectively (Figure 1). In comparison, Echegaray et al. [26] obtained a range of specific growth rates between 0.019 and 0.240 h−1 using diluted sugarcane molasses as a substrate (170–270 g/L total reducing sugar range) under anaerobic cultivation of S. cerevisiae. In addition, an increase in μ values from 0.096 to 0.123 h−1 was observed when the initial glucose concentration increased from 12.5 to 23.0 g/L. A similar trend was reported by Laopaiboon et al. [27], whereby an increase in glucose concentration from 10 to 150 g/L resulted in an increase of μ value from 0.43 to 0.49 h−1. These findings suggest that the specific growth rate of a culture increases with increasing substrate concentration, until substrate saturation is reached [28].
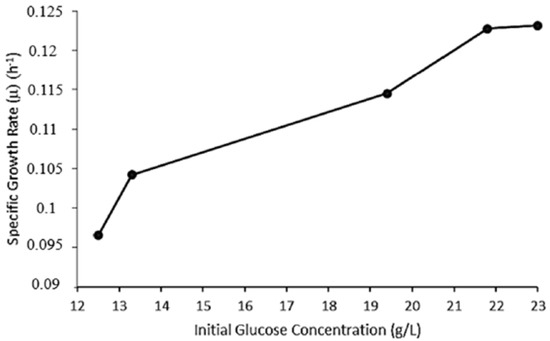
Figure 1.
Specific growth rates (μ) of S. cerevisiae BY4743 at varied initial glucose concentrations.
Data on the specific growth rate (μ) values and initial substrate concentrations were used to estimate KS and μmax (Figure 2). A maximum specific growth rate (μmax) value of 0.176 h−1 was obtained, which was close to the value of 0.169 h−1 previously reported by Dodić et al. [19] using S. cerevisiae cells grown on raw sugar beet juice. As cell growth rate is largely dependent on substrate concentration, it is expected that a higher initial sugar concentration will result in higher Monod coefficients [28]. The KS value obtained (10.11 g/L) was in line with values previously reported from several studies on lignocellulosic substrates (Table 1). Using citrus pulp waste as a substrate, a KS value of 10.690 g/L was reported by Raposo et al. [29], while Srimachai et al. [16] obtained a KS value of 10.210 g/L using oil palm frond juice. These observations imply that S. cerevisiae has a similar affinity (1/KS) to sorghum leaves as oil palm frond juice, glucose, and citrus waste pulp. In contrast to this, Ariyajaroenwong et al. [2] reported a Monod constant (KS) of 47.510 g/L when using sweet sorghum juice as a substrate. This decreased affinity may be due to the presence of more than one type of sugar in sweet sorghum juice [2]. Singh and Sharma [30] reported a KS value of 3.700 g/L using glucose, which is much lower than the range observed in previous studies; however, this corresponds to a higher affinity constant, which is expected as glucose is metabolised with ease.
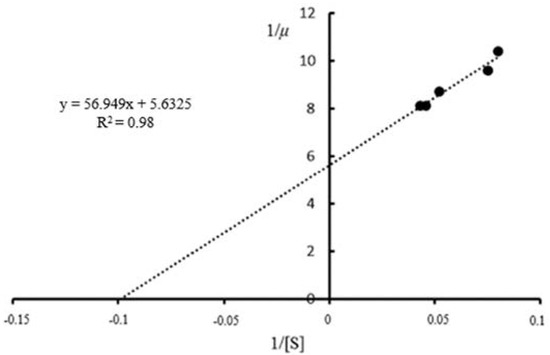
Figure 2.
Lineweaver–Burk plot used to estimate Monod constants for batch ethanol production from waste sorghum leaves (SL).

Table 1.
Comparison of the obtained Monod model coefficients with previous studies.
Variations in KS values (from 3.7 to 213.6 g/L) can be attributed to substrate type and concentration, strains of yeast employed, or the fermentation process itself [2]. These data demonstrate that the suitability of waste sorghum leaves as a substrate for S. cerevisiae growth is similar to that of raw sugar beet juice and oil palm frond juice. Furthermore, the fermentation volume size may impact the KS value. This is illustrated by the vast differences in substrate affinity for glucose obtained by Shafaghat et al. [22] using a working volume of less than 250 mL and Ahmad et al. [9] with a working volume of 8 L. The differences observed between the aforementioned studies may be attributed to additional process challenges encountered in large volume, such as poor agitation, low mass transfer, and inhomogeneity.
3.2. Bioethanol Production
The bioethanol production trend of S. cerevisiae cultivated on fermentation medium prepared from sorghum leaves is shown in Figure 3. A rapid depletion of glucose was observed from 0 to 32 h. A lag phase in bioethanol production of 6 h was obtained. This corresponds to cell adaptation and synthesis of key nutrients required for biomass or product (bioethanol) formation [14]. Ardestani and Shafiei [33] reported exponential growth of S. cerevisiae after 7 h of incubation. A rapid increase in ethanol concentration was observed from 6 to 28 h, corresponding to the exponential stage (Figure 4). This is expected, as ethanol is a primary metabolite and is therefore produced during the exponential phase of cell growth. A similar observation was reported by Lin et al. [34], where a steady increase in ethanol was observed over a duration of 48 h at 30 and 40 °C. An average ethanol yield of 0.49 g-ethanol/g-glucose was obtained, corresponding to a 96% fermentation efficiency during this period. Fermentation efficiencies between 72.78% and 78.43% have been reported by Srimachai et al. [25] using oil palm frond juice as a substrate, whilst ethanol yields between 0.40 and 0.49 g/g have been obtained from raw sugar beet juice [19]. Waste sorghum leaves show excellent potential for lignocellulosic bioethanol production. A productivity of 0.345 g/L/h was observed in this study. Ethanol productivities on other lignocellulosic substrates in the range of 0.25 to 1.01 g/L/h have been reported [35,36,37,38], further pointing to the relative higher potential of waste sorghum leaves for bioethanol production.
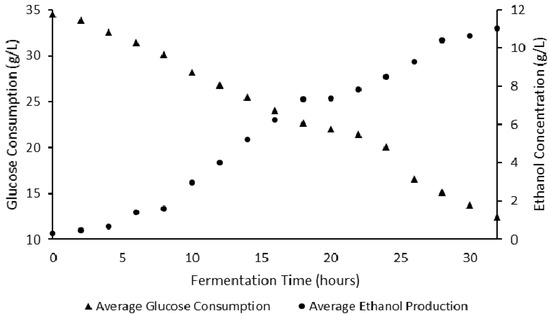
Figure 3.
Average glucose utilisation and ethanol formation during batch fermentation by S. cerevisiae BY4743.
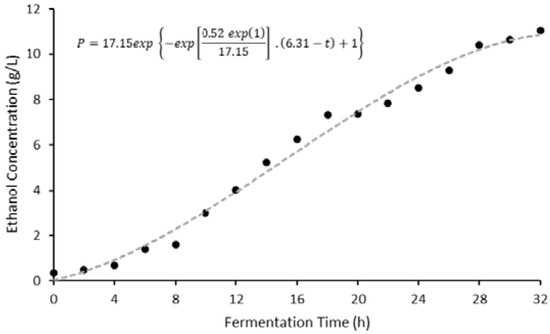
Figure 4.
Plot illustrating the fitted modified Gompertz curve.
The modified Gompertz model was fitted to the experimental data, and kinetic coefficients were determined (Equation (8)).
The fitted regression curve exhibited an R2 value of 0.98 and a correlation coefficient (r) of 0.99, suggesting that this model is able to efficiently describe bioethanol production during the fermentation of sorghum leaf wastes. The Gompertz coefficients for maximum potential bioethanol concentration (Pm), maximum bioethanol production rate (rp,m), and lag time were 17.15 g/L, 0.52 g/L/h, and 6.31 h, respectively, from waste sorghum leaves. Very few studies have reported a lag time of longer than one hour [19]. This suggests that a duration of at least 6 h was required for yeast cells to adapt to fermentation medium derived from waste sorghum leaves. Additionally, the maximum potential bioethanol concentration of 17.15 g/L—which corresponds to 2.17% (v/v)—illustrates that the impact of ethanol concentration within the medium may have a slight effect on the specific growth rate of S. cerevisiae. This is supported by an earlier study by Dinh et al. [39], which showed that a higher initial ethanol concentration within fermentation media resulted in an increase in the time required for cells to reach the optimal bioethanol production rate as well as a reduction in the maximum ethanol concentration.
Table 2 shows a comparison of the Gompertz coefficients obtained from this study using sorghum leaves and those reported from oil palm frond juice and sugar beet raw juice. From sorghum leaves, a higher maximum potential bioethanol concentration was achieved. In addition to this, an observed bioethanol production rate of 0.52 g/L/h was two times that achieved by Srimachai et al. [25] from oil palm frond juice. This illustrates the higher potential of waste sorghum leaves to accommodate a higher production rate.
Table 2.
Comparison of modified Gompertz model parameters with previous studies.
4. Conclusions
This study developed two kinetic models to describe the growth of S. cerevisiae BY4743 on pre-treated waste sorghum leaves for bioethanol production. Experimental data fitted the Monod and modified Gompertz model with high accuracy giving R2 values of 0.95 and 0.98, respectively. From the Monod model, a maximum specific growth rate and Monod constant of 0.176 h−1 and 10.11 g/L were obtained, respectively. These findings show that waste sorghum leaves have a greater potential for bioethanol production with a higher production rate and productivity than several lignocellulosic substrates. Furthermore, a maximum yield of 0.49 g-ethanol/g-glucose was achieved after 32 h of fermentation. The generated kinetic knowledge of S. cerevisiae growth on sorghum leaves and bioethanol formation in this study is of high importance for process optimisation and scale-up towards commercialisation of this fuel.
Acknowledgments
The financial assistance of the National Research Foundation of South Africa (NRF) towards this research is hereby acknowledged.
Author Contributions
Daneal C. S. Rorke and Evariste Bosco Gueguim Kana conceived and designed the experiments; Daneal C. S. Rorke performed the experiments and analysed the data; Daneal C. S. Rorke wrote the article with the help and guidance of Evariste Bosco Gueguim Kana.
Conflicts of Interest
The authors declare no conflict of interest.
References
- DoE. Biofuels Pricing and Manufacturing Economics. Department of Energy: South Africa, 2012. Available online: http://www.energy.gov.za/files/esources/renewables/BiofuelsPricingAndManufacturingEconomics.pdf (accessed on 10 November 2016).
- Ariyajaroenwong, P.; Laopaiboon, P.; Salakkam, A.; Srinophakun, P.; Laopaiboon, L. Kinetic models for batch and continuous ethanol fermentation from sweet sorghum juice by yeast immobilized on sweet sorghum stalks. J. Taiwan Inst. Chem. Eng. 2016, 66, 210–216. [Google Scholar] [CrossRef]
- Diaz, A.B.; de Souza Moretti, M.M.; Bezerra-Bussoli, C.; da Costa Carreira Nunes, C.; Blandino, A.; da Silva, R.; Gomes, E. Evaluation of microwave-assisted pretreatment of lignocellulosic biomass immersed in alkaline glycerol for fermentable sugars production. Bioresour. Technol. 2015, 185, 316–323. [Google Scholar] [CrossRef] [PubMed]
- Mai, N.L.; Ha, S.H.; Koo, Y. Efficient pretreatment of lignocellulose in ionic liquids/co-solvent for enzymatic hydrolysis enhancement into fermentable sugars. Process Biochem. 2014, 49, 1144–1151. [Google Scholar] [CrossRef]
- Vani, S.; Sukumaran, R.K.; Savithri, S. Prediction of sugar yields during hydrolysis of lignocellulosic biomass using artificial neural network modeling. Bioresour. Technol. 2015, 188, 128–135. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Reynosa, A.; Romaní, A.; Rodríguez-Jasso, R.M.; Aguilar, C.N.; Garrote, G.; Ruiz, H.A. Microwave heating processing as alternative of pretreatment in second-generation biorefinery: An overview. Energy Convers. Manag. 2017, 136, 50–65. [Google Scholar] [CrossRef]
- Gabhane, J.; Prince William, S.P.M.; Gadhe, A.; Rath, R.; Vaidya, A.N.; Wate, S. Pretreatment of banana agricultural waste for bio-ethanol production: Individual and interactive effects of acid and alkali pretreatments with autoclaving, microwave heating and ultrasonication. Waste Manag. 2014, 34, 498–503. [Google Scholar] [CrossRef] [PubMed]
- Putra, M.D.; Abasaeed, A.E.; Atiyeh, H.K.; Al-Zahrani, S.M.; Gaily, M.H.; Sulieman, A.K.; Zeinelabdeen, M.A. Kinetic Modeling and Enhanced Production of Fructose and Ethanol from Date Fruit Extract. Chem. Eng. Commun. 2015, 202, 1618–1627. [Google Scholar] [CrossRef]
- Ahmad, F.; Jameel, A.T.; Kamarudin, M.H.; Mel, M. Study of growth kinetic modeling of ethanol production by Saccharomyces cerevisiae. Afr. J. Biotechnol. 2011, 16, 18842–18846. [Google Scholar]
- Massoud, M.I.; El-Razek, A.M.A. Suitability of Sorghum bicolor L. stalks and grains for bioproduction of ethanol. Ann. Agric. Sci. 2011, 56, 83–87. [Google Scholar] [CrossRef][Green Version]
- Suryaningsih, R. Bioenergy Plants in Indonesia: Sorghum for Producing Bioethanol as an Alternative Energy Substitute of Fossil Fuels. Energy Procedia 2014, 47, 211–216. [Google Scholar] [CrossRef][Green Version]
- Shen, F.; Zeng, Y.; Deng, S.; Liu, R. Bioethanol production from sweet sorghum stalk juice with immobilized yeast. Procedia Environ. Sci. 2011, 11, 782–789. [Google Scholar] [CrossRef]
- Martins, F.; Gay, J.C. Biofuels: From Boom to Bust? Bain Brief, Bain and Company, 17 September 2014. Available online: http://www.bain.com/publications/articles/biofuels-from-boom-to-bust.aspx# (accessed on 23 October 2016).
- Lee, S.Y. Kinetic Modeling and Simulation. Encycl. Syst. Biol. 2013. [Google Scholar] [CrossRef]
- Almquist, J.; Cvijovic, M.; Hatzimanikatis, V.; Nielsen, J.; Jirstrand, M. Kinetic models in industrial biotechnology—Improving cell factory performance. Metab. Eng. 2014, 24, 38–60. [Google Scholar] [CrossRef] [PubMed]
- Ordoñez, M.C.; Raftery, J.P.; Jaladi, T.; Chen, X.; Kao, K.; Karim, M.N. Modelling of batch kinetics of aerobic carotenoid production using Saccharomyces cerevisiae. Biochem. Eng. J. 2016, 114, 226–236. [Google Scholar] [CrossRef]
- Linville, J.L.; Rodriguez, M., Jr.; Mielenz, J.R.; Cox, C.D. Kinetic modeling of batch fermentation for Populus hydrolysate tolerant mutant and wild type strains of Clostridium thermocellum. Bioresour. Technol. 2013, 147, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Imamoglu, E.; Sukan, F.V. Scale-up and kinetic modeling for bioethanol production. Bioresour. Technol. 2013, 144, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Dodić, J.M.; Vučurović, D.G.; Dodić, S.N.; Grahovac, J.A.; Popov, S.D.; Nedeljković, N.M. Kinetic modelling of batch ethanol production from sugar beet raw juice. Appl. Energy 2012, 99, 192–197. [Google Scholar] [CrossRef]
- Jukić, D.; Kralik, G.; Scitovski, R. Least-squares fitting Gompertz curve. J. Comput. Appl. Math. 2004, 169, 359–375. [Google Scholar] [CrossRef][Green Version]
- Mu, Y.; Wang, G.; Yu, H.Q. Kinetic modelling of batch hydrogen production process by mixed anaerobic cultures. Bioresour. Technol. 2006, 97, 1302–1307. [Google Scholar] [CrossRef] [PubMed]
- Shafaghat, H.; Najafpour, G.D.; Rezaei, P.S.; Sharifzadeh, M. Growth Kinetics and Ethanol Productivity of Saccharomyces cerevisiae PTCC 24860 on Various Carbon Sources. World Appl. Sci. J. 2009, 7, 140–144. [Google Scholar]
- Rorke, D.C.S.; Suinyuy, T.N.; Gueguim Kana, E.B. Microwave-assisted chemical pre-treatment of waste sorghum leaves: Process optimization and development of an intelligent model for determination of volatile compound fractions. Bioresour. Technol. 2017, 224, 590–600. [Google Scholar] [CrossRef] [PubMed]
- Burk, D. Enzyme kinetic constants: The double reciprocal plot. Trends Biochem. Sci. 1984, 9, 202–204. [Google Scholar] [CrossRef]
- Srimachai, T.; Nuithitikul, K.; O-thong, S.; Kongjan, P.; Panpong, K. Optimization and Kinetic Modeling of Ethanol Production from Oil Palm Frond Juice in Batch Fermentation. Energy Procedia 2015, 79, 111–118. [Google Scholar] [CrossRef]
- Echegaray, O.F.; Carvalho, J.C.M.; Fernandes, A.N.R.; Satos, S.; Aquarone, E.; Vitolo, M. Fed-batch culture of Saccharomyces cerevisiae in sugar-cane blackstrap molasses: Invertase activity of intact cells in ethanol fermentation. Biomass Bioenergy 2000, 19, 39–50. [Google Scholar] [CrossRef]
- Laopaiboon, L.; Nuanpeng, S.; Srinophakun, P.; Klanrit, P.; Laopaiboon, P. Selection of Saccharomyces cerevisiae and Investigation of its Performance for Very High Gravity Ethanol Fermentation. Biotechnology 2008, 7, 493–498. [Google Scholar] [CrossRef]
- Okpokwasili, G.C.; Nweke, C.O. Microbial growth and substrate utilization kinetics. Afr. J. Biotechnol. 2005, 5, 305–317. [Google Scholar]
- Raposo, S.; Pardão, J.M.; Díaz, I.; Lima-Costa, M.E. Kinetic modelling of bioethanol production using agro-industrial by-products. Int. J. Energy Environ. 2009, 1, 1–8. [Google Scholar]
- Singh, J.; Sharma, R. Growth kinetic and modeling of ethanol production by wilds and mutant Saccharomyces cerevisiae MTCC 170. Eur. J. Exp. Biol. 2015, 5, 1–6. [Google Scholar]
- Govindaswamy, S.; Vane, L.M. Kinetics of growth and ethanol production on different carbon substrates using genetically engineered xylose-fermenting yeast. Bioresour. Technol. 2007, 98, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Manikandan, K.; Saravanan, V.; Viruthagiri, T. Kinetics studies on ethanol production from banana peel waste using mutant strain of Saccharomyces cerevisiae. Indian J. Biotechnol. 2008, 7, 83–88. [Google Scholar]
- Ardestani, F.; Shafiei, S. Non-Structured Kinetic Model for the Cell Growth of Saccharomyces cerevisiae in a Batch Culture. Iran. J. Energy Environ. 2014, 5, 8–12. [Google Scholar] [CrossRef]
- Lin, Y.; Zhang, W.; Li, C.; Sakakibara, K.; Tanaka, S.; Kong, H. Factors affecting ethanol fermentation using Saccharomyces cerevisiae BY4742. Biomass Bioenergy 2012, 47, 395–401. [Google Scholar] [CrossRef]
- Abdullah, S.S.S.; Shirai, Y.; Bahrin, E.K.; Hassan, M.A. Fresh oil palm frond juice as a renewable, non-food, non-cellulosic and complete medium for direct bioethanol production. Ind. Crops Prod. 2015, 63, 357–361. [Google Scholar] [CrossRef]
- Ramos, C.L.; Duarte, W.F.; Freire, A.L.; Dias, D.R.; Eleutherio, E.C.; Schwan, R.F. Evaluation of stress tolerance and fermentative behaviour of indigenous Saccharomyces cerevisiae. Braz. J. Microbiol. 2013, 44, 935–944. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Staggenborg, S.; Propheter, J.L.; Rooney, W.L.; Yu, J.; Wang, D. Features of sweet sorghum juice and their performance in ethanol fermentation. Ind. Crops Prod. 2010, 31, 164–170. [Google Scholar] [CrossRef]
- Pavlečič, M.; Vrana, I.; Vibovec, K.; Šantek, M.I.; Horvat, P.; Šantek, B. Ethanol Production from Different Intermediates of Sugar Beet Processing. Food Technol. Biotechnol. 2010, 48, 362–367. [Google Scholar]
- Dinh, T.N.; Nagahisa, K.; Hirasawa, T.; Furusawa, C.; Shimizu, H. Adaptation of Saccharomyces cerevisiae Cells to High Ethanol Concentration and Changes in Fatty Acid Composition of Membrane and Cell Size. PLoS ONE 2008, 3, e2623. [Google Scholar] [CrossRef] [PubMed]
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).