
Sustainable Production of Bacterial Cellulose in a Rotary Disk Bioreactor: Grape Pomace as a Replacement for the Carbon Source

 ,
,  , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Strain and Hestrin–Schramm S1717 Medium Preparation
2.3. Preparation of the Inoculum
2.4. Fermentation in Static Culture
2.5. Cellulose and Biomass Quantification
2.6. Quantification of Reducing Sugars
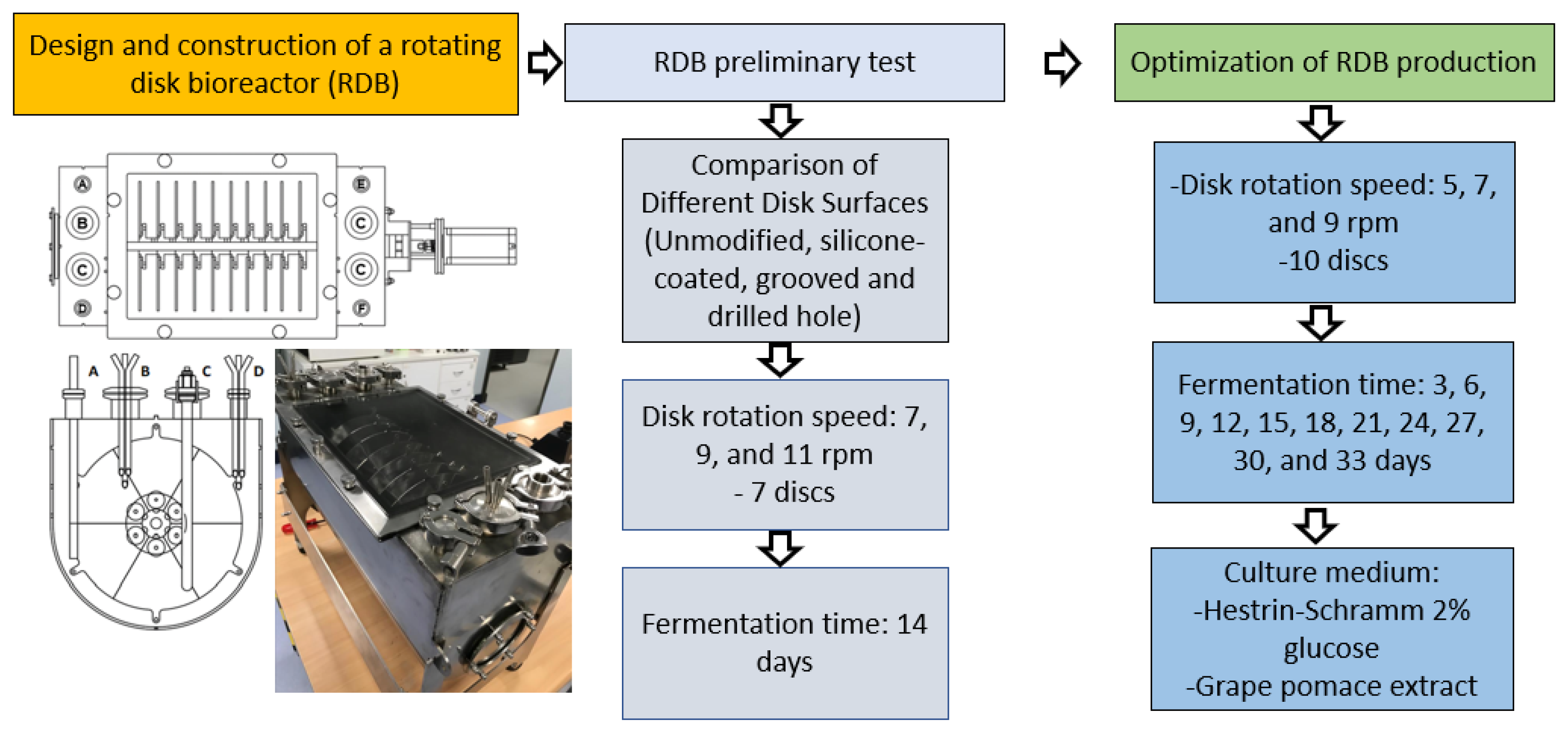
2.7. Optimization of BNC Production in Rotary Disk Bioreactor
2.7.1. Start-Up of the RDB
2.7.2. Comparison of Different Disk Surfaces
2.7.3. Comparing RDB Operating Parameters
2.8. Grape Pomace Extracts
2.9. Comparison Between Rotary Disk Bioreactor and Static Culture Using Grape Pomace
2.10. Determination of Productive Yields
- Yp/s: Product yield coefficient based on substrate
- Yp/x: Product yield coefficient based on biomass
- Yx/s: Cell yield coefficient based on substrate
- Q: Volumetric productivity (Q)
- ΔP: Mass of BNC produced (g)
- ΔS: Mass of substrate consumed (g)
- ΔX: Biomass generated (g)
- MP: Mass of BNC produced (mg)
- T: Cultivation time (days)
- V: Fermentation volume (L)
2.11. Characterization of the BNC Pellicles
3. Results and Discussion
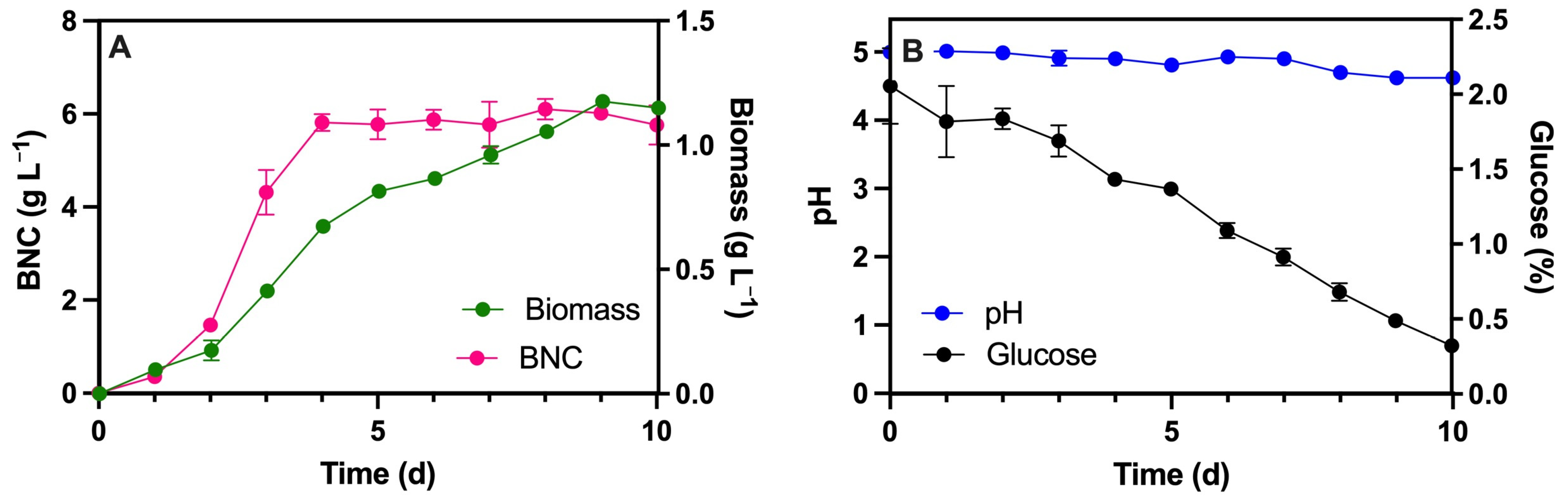
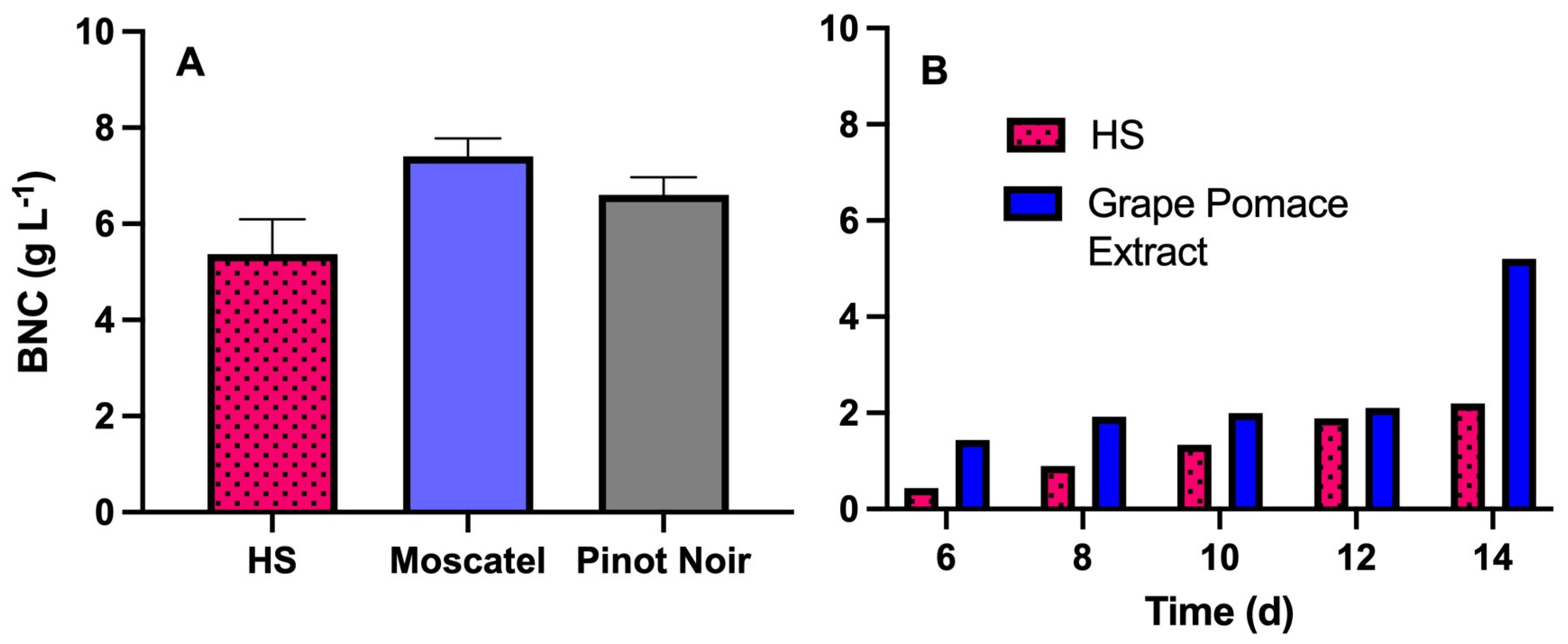
3.1. Fermentation in Static Culture
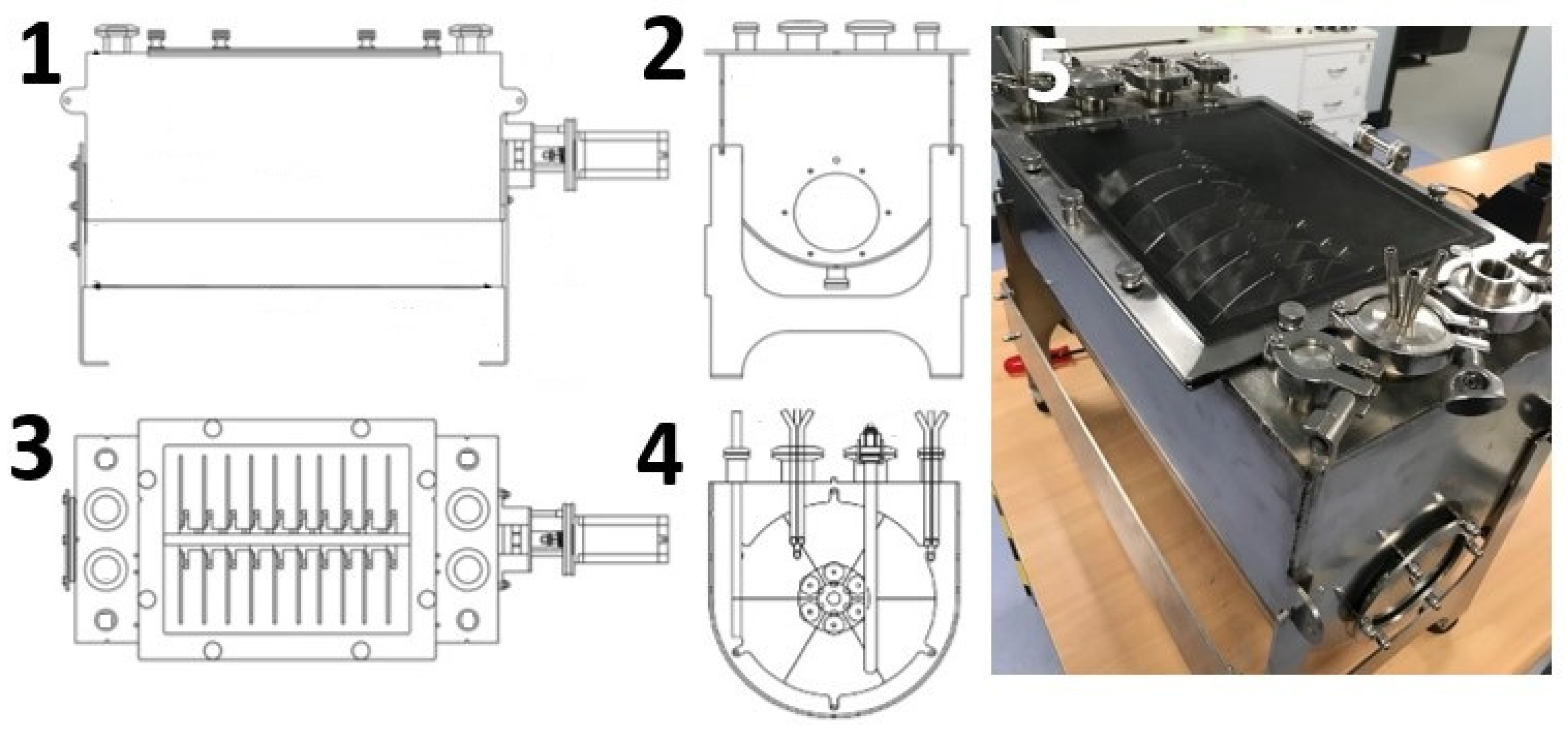
3.2. Rotary Disk Bioreactor Design
3.3. Optimization of BNC Production in Rotary Disk Bioreactor
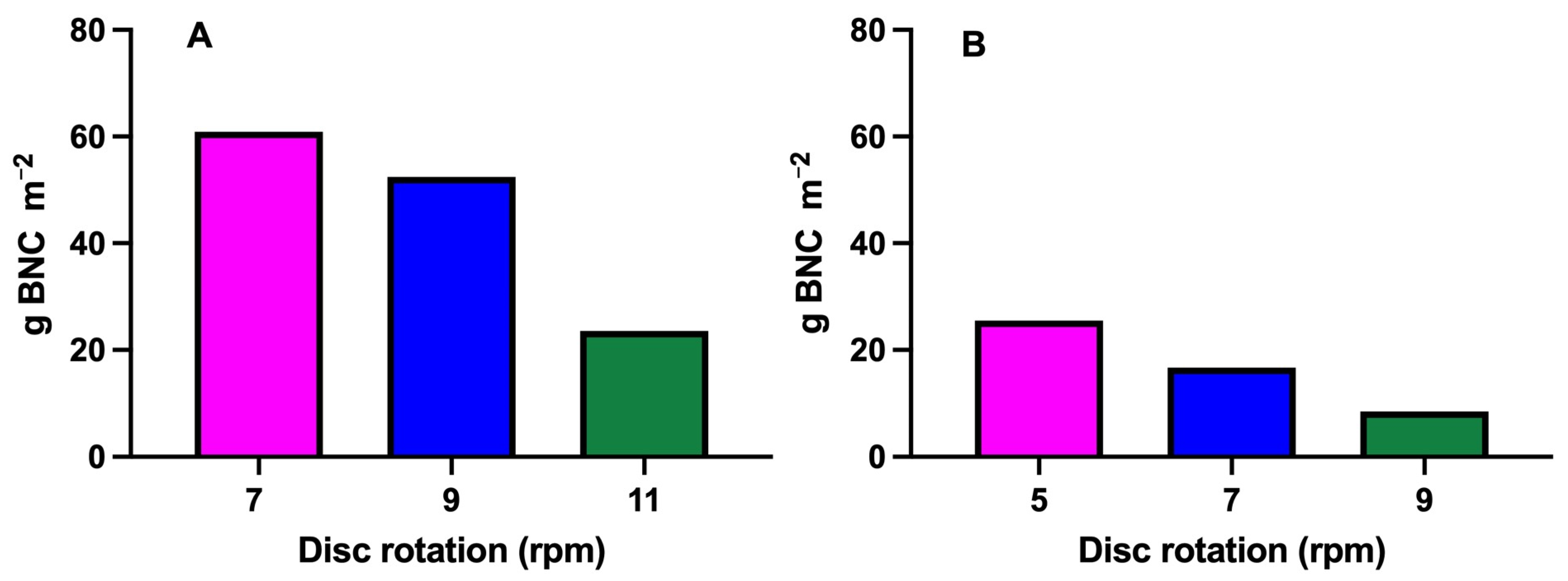
3.3.1. Comparison of Different Disk Surfaces
3.3.2. Comparison of Different Disk Rotation Speeds
3.3.3. Kinetics of BNC Production in a Rotary Disk Bioreactor
3.3.4. BNC Production in Rotary Disk Bioreactor Using Grape Pomace
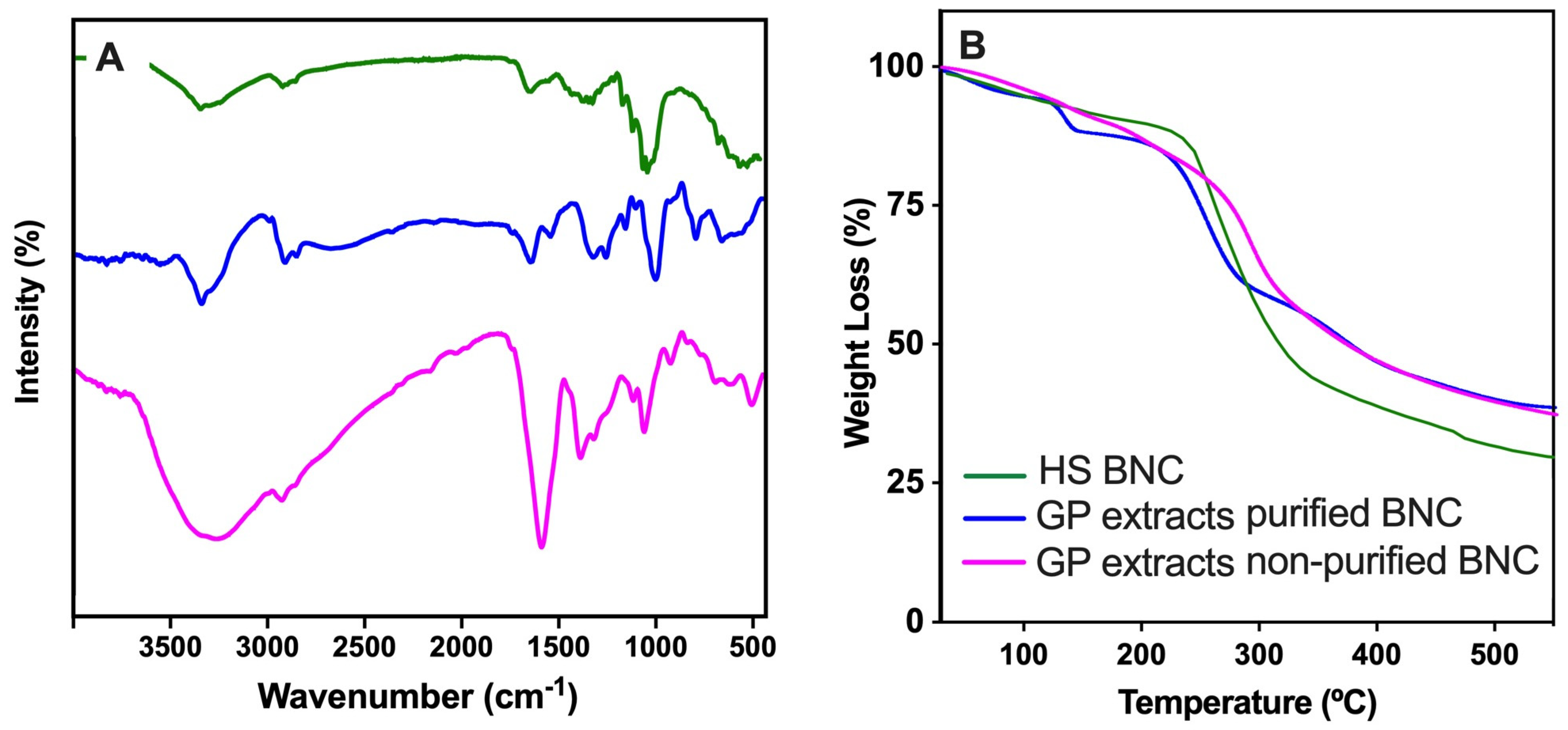
3.4. BNC Characterization
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BNC | Bacterial nanocellulose |
| RDB | Rotary disk bioreactor |
| TGA | Thermogravimetric analysis |
| ATCC | American Type Culture Collection |
| SEM | Scanning electron microscope |
| HS | Hestrin–Schramm medium |
| GRAS | Generally Recognized as Safe |
| FDA | Food and Drug Administration |
| DO | Dissolved oxygen |
| CCD | Central composite design |
| GP | Grape pomace |
References
- Soleimani, A.; Hamedi, S.; Babaeipour, V.; Rouhi, M. Design, Construction and Optimization a Flexible Bench-Scale Rotating Biological Contactor (RBC) for Enhanced Production of Bacterial Cellulose by Acetobacter Xylinium. Bioprocess. Biosyst. Eng. 2021, 44, 1071–1080. [Google Scholar] [CrossRef]
- Jagtap, A.; Dastager, S.G. Bacterial Nanocellulose: A Versatile Biopolymer Production Using a Cost-Effective Wooden Disc Based Rotary Reactor. Biopolymers 2024, 115, e23577. [Google Scholar] [CrossRef]
- Sharma, C.; Bhardwaj, N.K.; Pathak, P.; Dey, P.; Gautam, S.; Kumar, S.; Dutt Purohit, S. Bacterial Nanocellulose by Static, Static Intermittent Fed-Batch and Rotary Disc Bioreactor-Based Fermentation Routes Using Economical Black Tea Broth Medium: A Comparative Account. Int. J. Biol. Macromol. 2024, 277, 134228. [Google Scholar] [CrossRef]
- Sharma, C.; Bhardwaj, N.K.; Pathak, P. Rotary Disc Bioreactor-Based Approach for Bacterial Nanocellulose Production Using Gluconacetobacter Xylinus NCIM 2526 Strain. Cellulose 2022, 29, 7177–7191. [Google Scholar] [CrossRef]
- Dourado, F.; Leal, M.; Martins, D.; Fontão, A.; Cristina Rodrigues, A.; Gama, M. Chapter 7—Celluloses as Food Ingredients/Additives: Is There a Room for BNC? In Bacterial Nanocellulose; Gama, M., Dourado, F., Bielecki, S., Eds.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 123–133. ISBN 978-0-444-63458-0. [Google Scholar]
- Manan, S.; Ullah, M.W.; Ul-Islam, M.; Shi, Z.; Gauthier, M.; Yang, G. Bacterial Cellulose: Molecular Regulation of Biosynthesis, Supramolecular Assembly, and Tailored Structural and Functional Properties. Prog. Mater. Sci. 2022, 129, 100972. [Google Scholar] [CrossRef]
- Ul-Islam, M.; Ullah, M.W.; Khan, S.; Park, J.K. Production of Bacterial Cellulose from Alternative Cheap and Waste Resources: A Step for Cost Reduction with Positive Environmental Aspects. Korean J. Chem. Eng. 2020, 37, 925–937. [Google Scholar] [CrossRef]
- Zahan, K.A.; Pa’e, N.; Muhamad, I.I. Monitoring the Effect of pH on Bacterial Cellulose Production and Acetobacter Xylinum 0416 Growth in a Rotary Discs Reactor. Arab. J. Sci. Eng. 2015, 40, 1881–1885. [Google Scholar] [CrossRef]
- Zahan, K.; Paé, N.; Muhamad, I. An Evaluation of Fermentation Period and Discs Rotation Speed of Rotary Discs Reactor for Bacterial Cellulose Production. Sains Malays. 2016, 45, 393–400. [Google Scholar]
- Zahan, K.A.; Pa’E, N.; Seng, K.F.; Muhamad, I.I. Monitoring Initial Glucose Concentration for Optimum pH Control during Fermentation of Microbial Cellulose in Rotary Discs Reactor. Key Eng. Mater. 2014, 594–595, 319–324. [Google Scholar] [CrossRef]
- Cruz, M.A.; Flor-Unda, O.; Avila, A.; Garcia, M.D.; Cerda-Mejía, L. Advances in Bacterial Cellulose Production: A Scoping Review. Coatings 2024, 14, 1401. [Google Scholar] [CrossRef]
- Hosseinzadeh, A.; Chegeni, A.; Babaeipour, V. Mathematical Modeling and Simulation of Oxygen Mass Transfer in Rotating Biological Contactor (RBC) for Bacterial Cellulose Production. Biochem. Eng. J. 2023, 200, 109076. [Google Scholar] [CrossRef]
- Hestrin, S.; Schramm, M. Synthesis of Cellulose by Acetobacter Xylinum. 2. Preparation of Freeze-Dried Cells Capable of Polymerizing Glucose to Cellulose *. Biochem. J. 1954, 58, 345–352. [Google Scholar] [CrossRef]
- Núñez, D.; Oyarzún, P.; Cáceres, R.; Elgueta, E.; Gamboa, M. Citrate-Buffered Yamanaka Medium Allows to Produce High-Yield Bacterial Nanocellulose in Static Culture Using Komagataeibacter Strains Isolated from Apple Cider Vinegar. Front. Bioeng. Biotechnol. 2024, 12, 1375984. [Google Scholar] [CrossRef]
- Levenspiel, O. The Monod Equation: A Revisit and a Generalization to Product Inhibition Situations. Biotechnol. Bioeng. 1980, 22, 1671–1687. [Google Scholar] [CrossRef]
- Núñez, D.; Cáceres, R.; Ide, W.; Varaprasad, K.; Oyarzún, P. An Ecofriendly Nanocomposite of Bacterial Cellulose and Hydroxyapatite Efficiently Removes Lead from Water. Int. J. Biol. Macromol. 2020, 165, 2711–2720. [Google Scholar] [CrossRef] [PubMed]
- Lestari, P.; Elfrida, N.; Suryani, A.; Suryadi, Y. Study on the Production of Bacterial Cellulose from Acetobacter Xylinum Using Agro-Waste. Jordan. J. Biol. Sci. 2014, 7, 75–80. [Google Scholar] [CrossRef]
- Avirasdya, R.A.; Nursiwi, A.; Sari, A.M.; Zaman, M.Z.; Sanjaya, A.P. Kinetics Study of Bacterial Cellulose Production by Acetobacter Xylinum FNCC 0001 with Variation of Carbon Sources. E3S Web Conf. 2022, 344, 03002. [Google Scholar] [CrossRef]
- Guevara, K.M.; Martínez-Valenzuela, G.; Sánchez-Vásquez, V.; Guerrero-Ruiz, K.; Fiallos-Cárdenas, M. Trends and Perspectives on Bacterial Nanocellulose: A Comprehensive Analysis from the Three Helixes of Innovation. Mater. Today. Sustain. 2025, 30, 101090. [Google Scholar] [CrossRef]
- Kumaravel, A.; Lin, S.P.; Santoso, S.P.; Shanmugasundaram, S.; Hsu, H.Y.; Hsieh, C.W.; Chou, Y.C.; Lin, H.W.; Cheng, K.C. Unlocking the Potential of Bacterial Cellulose: Synthesis, Functionalization, and Industrial Impact. Int. J. Biol. Macromol. 2025, 311, 143951. [Google Scholar] [CrossRef]
- Serafica, G.; Mormino, R.; Bungay, H. Inclusion of Solid Particles in Bacterial Cellulose. Appl. Microbiol. Biotechnol. 2002, 58, 756–760. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yang, S.F.; Li, Y.; Xu, H.; Qin, L.; Tay, J.H. The Influence of Cell and Substratum Surface Hydrophobicities on Microbial Attachment. J. Biotechnol. 2004, 110, 251–256. [Google Scholar] [CrossRef]
- Pilafidis, S.; Vardaxi, A.; Kourmentza, K.; Pispas, S.; Dimopoulou, M.; Tsouko, E. From Bread Waste to Bacterial Cellulose Nanostructures: Development of a Novel Rotating Disk Bioreactor. Int. J. Biol. Macromol. 2025, 314, 144374. [Google Scholar] [CrossRef]
- Coyte, K.Z.; Tabuteau, H.; Gaffney, E.A.; Fostera, K.R.; Durham, W.M. Microbial Competition in Porous Environments Can Select against Rapid Biofilm Growth. Proc. Natl. Acad. Sci. USA 2017, 114, E161–E170. [Google Scholar] [CrossRef]
- Rendueles, O.; Ghigo, J.-M. Mechanisms of Competition in Biofilm Communities. Microbiol. Spectr. 2015, 3, 3. [Google Scholar] [CrossRef]
- Zahan, K.A.; Pa’, N.; Muhamad, I.I. Process Parameters for Fermentation in a Rotary Disc for Optimum Microbial Cellulose Production Response Surface Methodology. Bioresources 2014, 9, 1858–1872. [Google Scholar] [CrossRef]
- Choi, C.N.; Song, H.J.; Kim, M.J.; Chang, M.H.; Kim, S.J. Properties of Bacterial Cellulose Produced in a Pilot-Scale Spherical Type Bubble Column Bioreactor. Korean J. Chem. Eng. 2009, 26, 136–140. [Google Scholar] [CrossRef]
- Krsmanovic, M.; Biswas, D.; Ali, H.; Kumar, A.; Ghosh, R.; Dickerson, A.K. Hydrodynamics and Surface Properties Influence Biofilm Proliferation. Adv. Colloid. Interface Sci. 2021, 288, 102336. [Google Scholar] [CrossRef]
- Lahiri, D.; Nag, M.; Dutta, B.; Dey, A.; Sarkar, T.; Pati, S.; Edinur, H.A.; Kari, Z.A.; Noor, N.H.M.; Ray, R.R. Bacterial Cellulose: Production, Characterization and Application as Antimicrobial Agent. Int. J. Mol. Sci. 2021, 22, 12984. [Google Scholar] [CrossRef]
- Beluhan, S.; Herceg, F.; Leboš Pavunc, A.; Djaković, S. Preparation and Structural Properties of Bacterial Nanocellulose Obtained from Beetroot Peel Medium. Energies 2022, 15, 9374. [Google Scholar] [CrossRef]
- Chen, G.; Chen, L.; Wang, W.; Chen, S.; Wang, H.; Wei, Y.; Hong, F.F. Improved Bacterial Nanocellulose Production from Glucose without the Loss of Quality by Evaluating Thirteen Agitator Configurations at Low Speed. Microb. Biotechnol. 2019, 12, 1387–1402. [Google Scholar] [CrossRef]
- Páez, M.A.; Casa-Villegas, M.; Aldas, M.; Luna, M.; Cabrera-Valle, D.; López, O.; Fernández, D.; Cruz, M.A.; Flor-Unda, O.; García, M.D.; et al. Insights into Agitated Bacterial Cellulose Production with Microbial Consortia and Agro-Industrial Wastes. Fermentation 2024, 10, 425. [Google Scholar] [CrossRef]
- Aswini, K.; Gopal, N.O.; Uthandi, S. Optimized Culture Conditions for Bacterial Cellulose Production by Acetobacter Senegalensis MA1. BMC Biotechnol. 2020, 20, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Ruka, D.R.; Simon, G.P.; Dean, K.M. Altering the Growth Conditions of Gluconacetobacter Xylinus to Maximize the Yield of Bacterial Cellulose. Carbohydr. Polym. 2012, 89, 613–622. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-J.; Kim, J.-N.; Wee, Y.-J.; Park, D.-H.; Ryu, H.-W. Bacterial Cellulose Production by Gluconacetobacter Sp. RKY5 in a Rotary Biofilm Contactor. Appl. Biochem. Biotechnol. 2007, 137–140, 529–537. [Google Scholar] [CrossRef]
- Krystynowicz, A.; Czaja, W.; Wiktorowska-Jezierska, A.; Gonçalves-Miśkiewicz, M.; Turkiewicz, M.; Bielecki, S. Factors Affecting the Yield and Properties of Bacterial Cellulose. J. Ind. Microbiol. Biotechnol. 2002, 29, 189–195. [Google Scholar] [CrossRef]
- Lin, S.P.; Liu, C.T.; Hsu, K.D.; Hung, Y.T.; Shih, T.Y.; Cheng, K.C. Production of Bacterial Cellulose with Various Additives in a PCS Rotating Disk Bioreactor and Its Material Property Analysis. Cellulose 2016, 23, 367–377. [Google Scholar] [CrossRef]
- Lin, S.P.; Hsieh, S.C.; Chen, K.I.; Demirci, A.; Cheng, K.C. Semi-Continuous Bacterial Cellulose Production in a Rotating Disk Bioreactor and Its Materials Properties Analysis. Cellulose 2014, 21, 835–844. [Google Scholar] [CrossRef]
- Samarakoon, K.; Rupasinghe, H.P.V. Valorization of Grape Pomace by Microbial Fermentation: Composition, Biological Activities and Potential Applications for the Food Industry. J. Food Compos. Anal. 2025, 144, 107656. [Google Scholar] [CrossRef]
- Antonić, B.; Jančíková, S.; Dordević, D.; Tremlová, B. Grape Pomace Valorization: A Systematic Review and Meta-Analysis. Foods 2020, 9, 1627. [Google Scholar] [CrossRef]
- Karastergiou, A.; Gancel, A.L.; Jourdes, M.; Teissedre, P.L. Valorization of Grape Pomace: A Review of Phenolic Composition, Bioactivity, and Therapeutic Potential. Antioxidants 2024, 13, 1131. [Google Scholar] [CrossRef]
- Spinei, M.; Oroian, M. The Potential of Grape Pomace Varieties as a Dietary Source of Pectic Substances. Foods 2021, 10, 14. [Google Scholar] [CrossRef]
- Taokaew, S. Bacterial Nanocellulose Produced by Cost-Effective and Sustainable Methods and Its Applications: A Review. Fermentation 2024, 10, 316. [Google Scholar] [CrossRef]
- Katia Cury, R.; Yelitza Aguas, M.; Ana Martinez, M.; Rafael Olivero, V.; Linda Chams, C. Residuos Agroindustriales Su Impacto, Manejo y Aprovechamiento. RECIA 2017, 9, 122–132. [Google Scholar] [CrossRef]
- Keshk, S.; Sameshima, K. Evaluation of Different Carbon Sources for Bacterial Cellulose Production. Afr. J. Biotechnol. 2005, 4, 478–482. [Google Scholar]
- Gorgieva, S.; Jančič, U.; Cepec, E.; Trček, J. Production Efficiency and Properties of Bacterial Cellulose Membranes in a Novel Grape Pomace Hydrolysate by Komagataeibacter Melomenusus AV436T and Komagataeibacter Xylinus LMG 1518. Int. J. Biol. Macromol. 2023, 244, 125368. [Google Scholar] [CrossRef]
- Tsouko, E.; Pilafidis, S.; Kourmentza, K.; Gomes, H.I.; Sarris, G.; Koralli, P.; Papagiannopoulos, A.; Pispas, S.; Sarris, D. A Sustainable Bioprocess to Produce Bacterial Cellulose (BC) Using Waste Streams from Wine Distilleries and the Biodiesel Industry: Evaluation of BC for Adsorption of Phenolic Compounds, Dyes and Metals. Biotechnol. Biofuels Bioprod. 2024, 17, 1–17. [Google Scholar] [CrossRef]
- Fan, X.; Gao, Y.; He, W.; Hu, H.; Tian, M.; Wang, K.; Pan, S. Production of Nano Bacterial Cellulose from Beverage Industrial Waste of Citrus Peel and Pomace Using Komagataeibacter Xylinus. Carbohydr. Polym. 2016, 151, 1068–1072. [Google Scholar] [CrossRef]
- Coelho, C.C.S.; Michelin, M.; Cerqueira, M.A.; Gonçalves, C.; Tonon, R.V.; Pastrana, L.M.; Freitas-Silva, O.; Vicente, A.A.; Cabral, L.M.C.; Teixeira, J.A. Cellulose Nanocrystals from Grape Pomace: Production, Properties and Cytotoxicity Assessment. Carbohydr. Polym. 2018, 192, 327–336. [Google Scholar] [CrossRef]
- Cortina, J.L.; Nayak, A.; Bhushan, B.; Rosales, A.; Rodriguez-Turiel, L. Valorisation Potential of Cabernet Grape Pomace for the Recovery of Polyphenols: Process Intensification, Optimisation and Study of Kinetics. Food Bioprod. Process. 2018, 109, 74–85. [Google Scholar] [CrossRef]
- Andritsou, V.; Melo, E.; Tsouko, E.; Ladakis, D.; Maragkoudaki, S.; Koutinas, A.; Matharu, A. Synthesis and Characterization of Bacterial Cellulose from Citrus-Based Sustainable Resources. ACS Omega 2018, 3, 10365–10373. [Google Scholar] [CrossRef]
- Keshk, S. Vitamin C Enhances Bacterial Cellulose Production in Gluconacetobacter Xylinus. Carbohydr. Polym. 2014, 99, 98–100. [Google Scholar] [CrossRef] [PubMed]
- Fuller, M.E.; Andaya, C.; McClay, K. Evaluation of ATR-FTIR for Analysis of Bacterial Cellulose Impurities. J. Microbiol. Methods 2018, 144, 145–151. [Google Scholar] [CrossRef]
- Lopes, J.d.C.; Madureira, J.; Margaça, F.M.A.; Cabo Verde, S. Grape Pomace: A Review of Its Bioactive Phenolic Compounds, Health Benefits, and Applications. Molecules 2025, 30, 362. [Google Scholar] [CrossRef] [PubMed]
- Chawla, P.R.; Bajaj, I.B.; Survase, S.; Singhal, R.S. Microbial Cellulose: Fermentative Production and Applications (Review). Food Technol. Biotechnol. 2009, 47, 107–124. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Productivity (g L−1 d−1) | Nº Disks/ϕ cm/Disks Spacing (mm) | Rotation Speed (rpm) | Cultivation Time (d) | pH/T °C | Disk Material/Culture Medium | Reference | |
|---|---|---|---|---|---|---|---|
| 1.54 | 8/12/- | 15 | 4 | 6.0/30 | Polypropylene | /HS | [35] |
| 3.01 | 16/13.5/10 | 13 | 7 | 5.5/30 | Polyethylene | /HS | [1] |
| 0.82 | 24/20/4 | 4 | 4 | 5.7/30 | Plastic | /HS | [36] |
| 0.85 | 6/8/- | 5 | 5 | -/28 | Plastic composite | /Modified corn steep liquor with fructose | [37] |
| 0.34 | 8/10/- | - | 20 | 6.0/30 | - | /Brewed tea | [3] |
| 1.61 | - | 7 | 4 | 5.0(c)/28 | - | /Supplemented liquid pineapple waste | [8] |
| 0.24 | 3/8/- | 5 | 5 | -/28 | Plastic composite | /Corn steep liquor with fructose | [38] |
| 0.66 | 8/10/- | 7 | 10 | 5.0(c)/30 | Stainless steel with 16 mesh size | /HS | [4] |
| 0.24 | 5.0(c)/20 | ||||||
| 2.60 | 6/14/50 | 5 | 5 | 6.0/28 | Wood | /Modified HS | [2] |
| 3.77 * | 10/10/35 | 20 | 7 | 6.0/30 | Polycarbonate | /Bread waste enzymatic hydrolysates | [23] |
| 0.19 | 7/20/18 | 9 | 14 | 5.0/20 | Silicone-coated | /HS | This study |
| 0.18 | 10/20/13 | 7 | 33 | /HS | |||
| 0.37 | 7/20/18 | 7 | 14 | /Supplemented GP | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cáceres, R.; Oyarzún, P.; Vargas, J.P.; Cuevas, F.; Torres, K.; Elgueta, E.; Martínez, I.; Núñez, D. Sustainable Production of Bacterial Cellulose in a Rotary Disk Bioreactor: Grape Pomace as a Replacement for the Carbon Source. Fermentation 2025, 11, 441. https://doi.org/10.3390/fermentation11080441
Cáceres R, Oyarzún P, Vargas JP, Cuevas F, Torres K, Elgueta E, Martínez I, Núñez D. Sustainable Production of Bacterial Cellulose in a Rotary Disk Bioreactor: Grape Pomace as a Replacement for the Carbon Source. Fermentation. 2025; 11(8):441. https://doi.org/10.3390/fermentation11080441
Chicago/Turabian StyleCáceres, Rodrigo, Patricio Oyarzún, Juan Pablo Vargas, Francisca Cuevas, Kelly Torres, Elizabeth Elgueta, Irene Martínez, and Dariela Núñez. 2025. "Sustainable Production of Bacterial Cellulose in a Rotary Disk Bioreactor: Grape Pomace as a Replacement for the Carbon Source" Fermentation 11, no. 8: 441. https://doi.org/10.3390/fermentation11080441
APA StyleCáceres, R., Oyarzún, P., Vargas, J. P., Cuevas, F., Torres, K., Elgueta, E., Martínez, I., & Núñez, D. (2025). Sustainable Production of Bacterial Cellulose in a Rotary Disk Bioreactor: Grape Pomace as a Replacement for the Carbon Source. Fermentation, 11(8), 441. https://doi.org/10.3390/fermentation11080441