
The Metabolites Produced by Lactic Acid Bacteria and Their Role in the Microbiota–Gut–Brain Axis

 , , and
, , and
Abstract
1. Introduction
2. Lactic Acid Bacteria
2.1. Main LAB Genera and Species
2.2. Fermentation Mechanisms and Metabolite Production
2.3. Factors Influencing Metabolite Production
3. Metabolites Produced by LAB and the Microbiota–Gut–Brain Axis
3.1. Lactic Acid
3.2. Short-Chain Fatty Acids (SCFAs)
3.3. Tryptophan Metabolites
3.4. Gamma-Aminobutyric Acid (GABA)
3.5. Exopolysaccharides
4. Health Implications and Therapeutic Applications
5. Therapeutic Application of LAB
6. Future Perspectives
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cryan, J.F.; O’Riordan, K.J.; Cowan, C.S.; Sandhu, K.V.; Bastiaanssen, T.F.; Boehme, M.; Codagnone, M.G.; Cussotto, S.; Fulling, C.; Golubeva, A.V.; et al. The microbiota–gut–brain axis. Physiol. Rev. 2023, 103, 277–320. [Google Scholar] [CrossRef] [PubMed]
- Morais, L.H.; Schreiber, H.L.; Mazmanian, S.K. The gut microbiota–brain axis in behaviour and brain disorders. Nat. Rev. Microbiol. 2021, 19, 241–255. [Google Scholar] [CrossRef] [PubMed]
- Heeney, D.D.; Gareau, M.G.; Marco, M.L. Intestinal Lactobacillus in health and disease, a driver or just along for the ride? Curr. Opin. Biotechnol. 2018, 49, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Silva, Y.P.; Bernardi, A.; Frozza, R.L. The role of short-chain fatty acids from gut microbiota in gut-brain communication. Front. Endocrinol. 2020, 11, 508738. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, J.; Lv, M.; Shao, Z.; Hungwe, M.; Wang, J.; Bai, X.; Xie, J.; Wang, Y.; Geng, W. Metabolism Characteristics of Lactic Acid Bacteria and the Expanding Applications in Food Industry. Front. Bioeng. Biotechnol. 2021, 9, 612285. [Google Scholar] [CrossRef]
- Santander-Cortés, A.I.; Cástro-Rosas, J. Isolation of lactic acid bacteria with probiotic potential from typical fermented Mexican food: A review. Pädi Boletín Científico Cienc. Básicas E Ing. Del ICBI 2024, 11, 59–68. [Google Scholar] [CrossRef]
- Yang, X.; Hong, J.; Wang, L.; Cai, C.; Mo, H.; Wang, J.; Fang, X.; Liao, Z. Effect of Lactic Acid Bacteria Fermentation on Plant-Based Products. Fermentation 2024, 10, 48. [Google Scholar] [CrossRef]
- Abdul Hakim, B.N.; Xuan, N.J.; Hazwani Oslan, S.N. A Comprehensive Review of Bioactive Compounds from Lactic Acid Bacteria: Potential Functions as Functional Food in Dietetics and the Food Industry. Foods 2023, 12, 2850. [Google Scholar] [CrossRef]
- Liu, Y.; Gao, S.; Cui, Y.; Wang, L.; Duan, J.; Yang, X.; Liu, X.; Zhang, S.; Sun, B.; Yu, H.; et al. Characteristics of Lactic Acid Bacteria as Potential Probiotic Starters and Their Effects on the Quality of Fermented Sausages. Foods 2024, 13, 198. [Google Scholar] [CrossRef]
- Ran, J.; Tang, Y.; Mao, W.; Meng, X.; Jiao, L.; Li, Y.; Zhao, R.; Zhou, H. Optimization of the fermentation process and antioxidant activity of mixed lactic acid bacteria for honeysuckle beverage. Front. Microbiol. 2024, 15, 1364448. [Google Scholar] [CrossRef]
- Asha, M.N.; Chowdhury, M.S.R.; Hossain, H.; Rahman, M.A.; Emon, A.A.; Tanni, F.Y.; Islam, M.R.; Hossain, M.M.; Rahman, M.M. Antibacterial potential of lactic acid bacteria isolated from raw cow milk in Sylhet district, Bangladesh: A molecular approach. Vet. Med. Sci. 2024, 10, e1463. [Google Scholar] [CrossRef] [PubMed]
- Du, R.; Li, W.; Li, J.; Zeng, S.; Chen, Z.; Gao, J.; Zheng, H.; Lin, H.; Zhu, G.; Qin, X.; et al. Dynamic changes of zinc chemical speciation and zinc-containing peptides release in oysters (Crassostrea hongkongensis) during enzymatic hydrolysis. Food Biosci. 2024, 58, 103649. [Google Scholar] [CrossRef]
- Wang, H.; He, X.; Li, J.; Wu, J.; Jiang, S.; Xue, H.; Zhang, J.; Jha, R.; Wang, R. Lactic acid bacteria fermentation improves physicochemical properties, bioactivity, and metabolic profiles of Opuntia ficus-indica fruit juice. Food Chem. 2024, 453, 139646. [Google Scholar] [CrossRef] [PubMed]
- Razola-Díaz, M.D.C.; De Montijo-Prieto, S.; Aznar-Ramos, M.J.; Jiménez-Valera, M.; Ruiz-Bravo, A.; Verardo, V.; Gómez-Caravaca, A.M. Effect of lactic acid bacteria fermentation on the polar compounds content with antioxidant and antidiabetic activity of avocado seed extracts. Fermentation 2023, 9, 420. [Google Scholar] [CrossRef]
- He, Y.; Zhu, Y.; Lv, J.; Gu, Y.; Wang, T.; Chen, J. Effects of lactic acid bacteria fermentation on the bioactive composition, volatile compounds and antioxidant activity of Huyou (Citrus aurantium ‘Changshan-huyou’) peel and pomace. Food Qual. Saf. 2023, 7, fyad003. [Google Scholar] [CrossRef]
- Leksono, B.Y.; Cahyanto, M.N.; Rahayu, E.S.; Yanti, R.; Utami, T. Enhancement of antioxidant activities in black soy milk through isoflavone aglycone production during indigenous lactic acid bacteria fermentation. Fermentation 2022, 8, 326. [Google Scholar] [CrossRef]
- Zhang, P.; Tang, F.; Cai, W.; Zhao, X.; Shan, C. Evaluating the effect of lactic acid bacteria fermentation on quality, aroma, and metabolites of chickpea milk. Front. Nutr. 2022, 9, 1069714. [Google Scholar] [CrossRef]
- Abd Al-Halim, L.R.; Nasr, N.M. Quality of new Lactobaillus rhamnosus like-yoghurt and the effect of using gum Arabic as prebiotic. Fayoum J. Agric. Res. Dev. 2024, 38, 381–391. [Google Scholar] [CrossRef]
- Colmenares-Cuevas, S.I.; Contreras-Oliva, A.; Salinas-Ruiz, J.; Hidalgo-Contreras, J.V.; Flores-Andrade, E.; García-Ramírez, E.J. Development and study of the functional properties of marshmallow enriched with bee (Apis mellifera) honey and encapsulated probiotics (Lactobacillus rhamnosus). Front. Nutr. 2024, 11, 1353530. [Google Scholar] [CrossRef]
- Shori, A.B. Comparative analysis of Lactobacillus starter cultures in fermented camel milk: Effects on viability, antioxidant properties, and sensory characteristics. Foods 2024, 13, 2711. [Google Scholar] [CrossRef]
- Ngamsamer, C.; Muangnoi, C.; Tongkhao, K.; Sae-Tan, S.; Treesuwan, K.; Sirivarasai, J. Potential health benefits of fermented vegetables with additions of Lacticaseibacillus rhamnosus GG and polyphenol vitexin based on their antioxidant properties and prohealth profiles. Foods 2024, 13, 982. [Google Scholar] [CrossRef] [PubMed]
- Karimkhani, M.M.; Jamshidi, A.; Nasrollahzadeh, M.; Armin, M.; Jafari, S.M.; Zeinali, T. Fermentation of Rubus dolichocarpus juice using Lactobacillus gasseri and Lacticaseibacillus casei and protecting phenolic compounds by Stevia extract during cold storage. Sci. Rep. 2024, 14, 5711. [Google Scholar] [CrossRef] [PubMed]
- Lima, W.D.L.; Monteiro, S.S.; Pasquali, M.A.D.B. Study of fermentation strategies by Lactobacillus gasseri for the Production of probiotic food using passion fruit juice combined with green tea as raw material. Foods 2022, 11, 1471. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Jiang, Y.; Zhang, X.; Cui, S.; Liu, X.; Zhao, J.; Zhang, H.; Chen, W. Comparative peptidomics analysis of Milk Fermented by Lactobacillus helveticus. Foods 2022, 11, 3885. [Google Scholar] [CrossRef]
- Skrzypczak, K.; Gustaw, W.; Fornal, E.; Kononiuk, A.; Michalak-Majewska, M.; Radzki, W.; Waśko, A. Functional and technological potential of whey protein isolate in production of milk beverages fermented by new strains of lactobacillus helveticus. Appl. Sci. 2020, 10, 7089. [Google Scholar] [CrossRef]
- Hovhannisyan, H.G.; Danielyan, L.V.; Chichoyan, N.B.; Pashayan, M.M.; Baghdasaryan, L.G.; Melkumyan, I.E.; Barseghyan, A.H. Improvement of functional and sensory properties of fermented dairy drink Narine using raw apricot gum. Funct. Foods Health Dis. 2024, 14, 600–614. [Google Scholar] [CrossRef]
- Gugel, I.; Marchetti, F.; Costa, S.; Gugel, I.; Baldini, E.; Vertuani, S.; Manfredini, S. 2G-lactic acid from olive oil supply chain waste: Olive leaves upcycling via Lactobacillus casei fermentation. Appl. Microbiol. Biotechnol. 2024, 108, 379. [Google Scholar] [CrossRef]
- Lugo-Zarate, L.; Delgado-Olivares, L.; Cruz-Cansino, N.D.S.; González-Olivares, L.G.; Castrejón-Jiménez, N.S.; Estrada-Luna, D.; Jiménez-Osorio, A.S. Blackberry juice fermented with two consortia of lactic acid bacteria and isolated whey: Physicochemical and antioxidant properties during storage. Int. J. Mol. Sci. 2024, 25, 8882. [Google Scholar] [CrossRef]
- Jeong, D.-E.; Kim, J.; Kim, B.; Cho, Y. Effects on antioxidant, whitening, and anti-wrinkle omprovement of ethanol extracts from fermented Trigonotis radicans var. sericea by Lactobacillus brevis. Food Sci. Preserv. 2024, 31, 173–182. [Google Scholar] [CrossRef]
- Wang, Z.; Zhao, C.; Guo, Z.; Li, S.; Zhu, Z.; Grimi, N.; Xiao, J. Fermentation of Betaphycus gelatinum using lactobacillus brevis: Growth of probiotics, total polyphenol content, polyphenol profile, and antioxidant capacity. Foods 2023, 12, 3334. [Google Scholar] [CrossRef]
- De Montijo-Prieto, S.; Razola-Díaz, M.D.C.; Barbieri, F.; Tabanelli, G.; Gardini, F.; Jiménez-Valera, M.; Ruiz-Bravo, A.; Verardo, V.; Gómez-Caravaca, A.M. Impact of lactic acid bacteria fermentation on phenolic compounds and antioxidant activity of avocado leaf extracts. Antioxidants 2023, 12, 298. [Google Scholar] [CrossRef] [PubMed]
- Neylon, E.; Nyhan, L.; Zannini, E.; Monin, T.; Münch, S.; Sahin, A.W.; Arendt, E.K. Food ingredients for the future: In-depth analysis of the effects of lactic acid bacteria fermentation on spent barley rootlets. Fermentation 2023, 9, 78. [Google Scholar] [CrossRef]
- Sionek, B.; Szydłowska, A.; Trząskowska, M.; Kołożyn-Krajewska, D. The impact of physicochemical conditions on lactic acid bacteria survival in food products. Fermentation 2024, 10, 298. [Google Scholar] [CrossRef]
- Fazio, A.; La Torre, C.; Caroleo, M.C.; Caputo, P.; Cannataro, R.; Plastina, P.; Cione, E. Effect of addition of pectins from jujubes (Ziziphus jujuba Mill.) on vitamin C production during heterolactic fermentation. Molecules 2020, 25, 2706. [Google Scholar] [CrossRef]
- Hou, F.; Cai, Y.; Wang, J. Antioxidant capacity changes and untargeted metabolite profile of broccoli during lactic acid bacteria fermentation. Fermentation 2023, 9, 474. [Google Scholar] [CrossRef]
- Abdel-Salam, S.W.; Abdel-Aziz, M.H.; Abdel-Razik, M.; Mattar, Z.A.; Abdel-Aziz, A.B.; Makharitta, R.R. Advanced in the production of L-Ascorbic acid using biotechnological processes. Int. J. Sci. Res. Arch. 2024, 3, 546–556. [Google Scholar] [CrossRef]
- Xie, Z.; McAuliffe, O.; Jin, Y.S.; Miller, M.J. Invited review: Genomic modifications of lactic acid bacteria and their applications in dairy fermentation. J. Dairy Sci. 2024, 107, 8749–8764. [Google Scholar] [CrossRef]
- Dong, H.; Wang, H.; Fu, S.; Zhang, D. CRISPR/Cas tools for enhancing the biopreservation ability of lactic acid bacteria in aquatic products. Front. Bioeng. Biotechnol. 2022, 10, 1114588. [Google Scholar] [CrossRef]
- Icer, M.A.; Sarikaya, B.; Kocyigit, E.; Atabilen, B.; Celik, M.N.; Capasso, R.; Agagündüz, D.; Budán, F. Contributions of Gamma-Aminobutyric Acid (GABA) produced by lactic acid bacteria on food quality and human health: Current applications and futes prospects. Foods 2024, 13, 2437. [Google Scholar] [CrossRef]
- Gagneten, M.; Passot, S.; Cenard, S.; Ghorbal, S.; Schebor, C.; Fonseca, F. Mechanistic study of the differences in lactic acid bacteria resistance to freeze or spray drying and storage. Appl. Microbiol. Biotechnol. 2024, 108, 361. [Google Scholar] [CrossRef]
- Yu, Y.; Yang, W.; Li, Y.; Cong, Y. Enteroendocrine cells: Sensing gut microbiota and regulating inflammatory bowel diseases. Inflamm. Bowel Dis. 2019, 26, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Scavuzzo, C.J.; Rakotovao, I.; Dickson, C.T. Differential effects of L-and D-lactate on memory encoding and consolidation: Potential role of HCAR1 signaling. Neurobiol. Learn. Mem. 2020, 168, 107151. [Google Scholar] [CrossRef] [PubMed]
- Luiz, J.; Manassi, C.; Magnani, M.; Cruz, A.; Pimentel, T.; Verruck, S. Lactiplantibacillus plantarum as a promising adjuvant for neurological disorders therapy through the brain-gut axis and related action pathways. Crit. Rev. Food Sci. Nutr. 2025, 65, 715–727. [Google Scholar] [CrossRef]
- Watanabe, T.; Nishio, H.; Tanigawa, T.; Yamagami, H.; Okazaki, H.; Watanabe, K.; Arakawa, T. Probiotic Lactobacillus casei strain Shirota prevents indomethacin-induced small intestinal injury: Involvement of lactic acid. Am. J. Physiol.-Gastrointest. Liver Physiol. 2009, 297, G506–G513. [Google Scholar] [CrossRef] [PubMed]
- Muto, M.; Kaji, T.; Onishi, S.; Yano, K.; Yamada, W.; Ieiri, S. An overview of the current management of short-bowel syndrome in pediatric patients. Surg. Today 2022, 52, 12–21. [Google Scholar] [CrossRef]
- Bamigbade, G.B.; Abdin, M.; Subhash, A.; Arachchi, M.P.; Ullah, N.; Gan, R.Y.; Ali, A.; Kamal-Eldin, A.; Ayyash, M. Plant polysaccharide-capped nanoparticles: A sustainable approach to modulate gut microbiota and advance functional food applications. Compr. Rev. Food Sci. Food Saf. 2025, 24, e70156. [Google Scholar] [CrossRef]
- Averina, O.V.; Zorkina, Y.A.; Yunes, R.A.; Kovtun, A.S.; Ushakova, V.M.; Morozova, A.Y.; Kostyuk, G.P.; Danilenko, V.N.; Chekhonin, V.P. Bacterial metabolites of human gut microbiota correlating with depression. Int. J. Mol. Sci. 2020, 21, 9234. [Google Scholar] [CrossRef]
- Dalile, B.; Van Oudenhove, L.; Vervliet, B.; Verbeke, K. The role of short-chain fatty acids in microbiota–gut–brain communication. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 461–478. [Google Scholar] [CrossRef]
- Erny, D.; Dokalis, N.; Mezö, C.; Castoldi, A.; Mossad, O.; Staszewski, O.; Frosch, M.; Villa, M.; Fuchs, V.; Mayer, A.; et al. Microbiota-derived acetate enables the metabolic fitness of the brain innate immune system during health and disease. Cell Metab. 2021, 33, 2260–2276.e7. [Google Scholar] [CrossRef]
- Solsona, R. Impact of the Short-Chain Fatty Acids on the Microbiota-Gut-Brain Axis. 2021. Available online: https://diposit.ub.edu/dspace/handle/2445/179149 (accessed on 13 June 2025).
- Stilling, R.M.; Van De Wouw, M.; Clarke, G.; Stanton, C.; Dinan, T.G.; Cryan, J.F. The neuropharmacology of butyrate: The bread and butter of the microbiota-gut-brain axis? Neurochem. Int. 2016, 99, 110–132. [Google Scholar] [CrossRef]
- Qian, X.H.; Xie, R.Y.; Liu, X.L.; Chen, S.D.; Tang, H.D. Mechanisms of short-chain fatty acids derived from gut microbiota in Alzheimer’s disease. Aging Dis. 2022, 13, 1252. [Google Scholar] [CrossRef] [PubMed]
- Barki, N.; Bolognini, D.; Börjesson, U.; Jenkins, L.; Riddell, J.; Hughes, D.I.; Ulven, T.; Hudson, B.D.; Ulven, E.R.; Dekker, N.; et al. Chemogenetics defines a short-chain fatty acid receptor gut–brain axis. eLife 2022, 11, e73777. [Google Scholar] [CrossRef] [PubMed]
- Majumdar, A.; Venkatesh, I.; Basu, A. Short-Chain Fatty Acids in the Microbiota-Gut-Brain Axis: Role in Neurodegenerative Disorders and Viral Infections. ACS Chem. Neurosci. 2023, 14, 1045–1062. [Google Scholar] [CrossRef]
- Huang, Y.; Wang, Y.F.; Miao, J.; Zheng, R.F.; Li, J.Y. Short-chain fatty acids: Important components of the gut-brain axis against AD. Biomed. Pharmacother. 2024, 175, 116601. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Noto, D.; Hoshino, Y.; Mizuno, M.; Miyake, S. Butyrate suppresses demyelination and enhances remyelination. J. Neuroinflamm. 2019, 16, 165. [Google Scholar] [CrossRef]
- Hoyles, L.; Snelling, T.; Umlai, U.; Nicholson, J.; Carding, S.; Glen, R.; McArthur, S. Microbiome–host systems interactions: Protective effects of propionate upon the blood–brain barrier. Microbiome 2018, 6, 55. [Google Scholar] [CrossRef]
- Wang, Z.; Yang, S.; Liu, L.; Mao, A.; Kan, H.; Yu, F.; Zhou, T. The gut microbiota-derived metabolite indole-3-propionic acid enhances leptin sensitivity by targeting STAT3 against diet-induced obesity. Clin. Transl. Med. 2024, 14, e70053. [Google Scholar] [CrossRef]
- Pan, T.; Pei, Z.; Fang, Z.; Wang, H.; Zhu, J.; Zhang, H.; Lu, W. Uncovering the specificity and predictability of tryptophan metabolism in lactic acid bacteria with genomics and metabolomics. Front. Cell. Infect. Microbiol. 2023, 13, 1154346. [Google Scholar] [CrossRef]
- Flannigan, K.L.; Nieves, K.M.; Szczepanski, H.E.; Serra, A.; Lee, J.W.; Alston, L.A.; Ramay, H.; Mani, S.; Hirota, S.A. The pregnane X receptor and indole-3-propionic acid shape the intestinal mesenchyme to restrain inflammation and fibrosis. Cell. Mol. Gastroenterol. Hepatol. 2023, 15, 765–795. [Google Scholar] [CrossRef]
- Sun, J.; Zhang, Y.; Kong, Y.; Ye, T.; Yu, Q.; Satyanarayanan, S.K.; Su, K.P.; Liu, J. Microbiota-derived metabolite Indoles induced aryl hydrocarbon receptor activation and inhibited neuroinflammation in APP/PS1 mice. Brain Behav. Immun. 2022, 106, 76–88. [Google Scholar] [CrossRef]
- Zheng, L.; Xin, J.; Ye, H.; Sun, N.; Gan, B.; Gong, X.; Zhang, T. Lactobacillus Johnsonii YH1136 alleviates schizophrenia-like behavior in mice: A gut–microbiota–brain axis hypothesis study. BMC Microbiol. 2025, 25, 191. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhao, P.; Cai, Z.; He, P.; Wang, J.; He, H.; Zhao, J. Buqi-Huoxue-Tongnao decoction drives gut microbiota-derived indole lactic acid to attenuate ischemic stroke via the gut-brain axis. Chin. Med. 2024, 19, 126. [Google Scholar] [CrossRef] [PubMed]
- Sahab, N.R.; Subroto, E.; Balia, R.L.; Utama, G.L. γ-Aminobutyric acid found in fermented foods and beverages: Current trends. Heliyon 2020, 6, e05526. [Google Scholar] [CrossRef] [PubMed]
- Jena, R.; Choudhury, P.K. Lactic acid bacteria in fermented dairy foods: Gamma-aminobutyric acid (GABA) production and its therapeutic implications. Food Biosci. 2024, 62, 105276. [Google Scholar] [CrossRef]
- Gutiérrez, L.C.D. Development of a Microencapsulated Functional Ingredient Formulated with the Neurotransmitter GABA and the Lactiplantibacillus plantarum K16 Produced Through Biotechnological. Ph.D. Thesis, Universidad del País Vasco-Euskal Herriko Unibertsitatea, Vitoria-Gasteiz, Spain, 2023. [Google Scholar]
- Salvador, A.G.; Antolinez, S.Q.; Furundarena, I.H.; Aróstegui, S.; de Cerio, F.G. Mental illness and healthy nutrition. New alternatives for its treatment. Rev. Esp. Nutr. Comunitaria 2021, 27, 70–81. [Google Scholar]
- Ikegami, M.; Narabayashi, H.; Nakata, K.; Yamashita, M.; Sugi, Y.; Fuji, Y.; Matsufuji, H.; Harata, G.; Yoda, K.; Miyazawa, K.; et al. Intervention in gut microbiota increases intestinal γ-aminobutyric acid and alleviates anxiety behavior: A possible mechanism via the action on intestinal epithelial cells. Front. Cell. Infect. Microbiol. 2024, 14, 1421791. [Google Scholar] [CrossRef]
- Martínez Blanco, P. Microorganismos Como Medicamentos, ¿Podemos Gestionar la Microbiota? 2020. Available online: http://hdl.handle.net/10902/19434 (accessed on 13 June 2025).
- Lacouture, A.T. Relación de la Microbiota Intestinal con el Desarrollo de Depresión y Ansiedad. Bachelor’s Thesis, Universidad Vasco de Quiroga, Morelia, Michoacán, 2023. [Google Scholar]
- Conn, K.; Borsom, E.; Cope, E. Implications of microbe-derived ɣ-aminobutyric acid (GABA) in gut and brain barrier integrity and GABAergic signaling in Alzheimer’s disease. Gut Microbes 2024, 16, 2371950. [Google Scholar] [CrossRef]
- Nguyen, H.-T.; Pham, T.-T.; Nguyen, P.-T.; Le-Buanec, H.; Rabetafika, H.N.; Razafindralambo, H.L. Advances in microbial exopolysaccharides: Present and future applications. Biomolecules 2024, 14, 1162. [Google Scholar] [CrossRef]
- Yegorenkova, I.V.; Tregubova, K.V.; Matora, L.Y.; Burygin, G.L.; Ignatov, V.V. Biofilm formation by Paenibacillus polymyxa strains differing in the production and rheological properties of their exopolysaccharides. Curr. Microbiol. 2011, 62, 1554–1559. [Google Scholar] [CrossRef]
- Li, S.; Huang, R.; Shah, N.P.; Tao, X.; Xiong, Y.; Wei, H. Antioxidant and antibacterial activities of exopolysaccharides from Bifidobacterium bifidum WBIN03 and Lactobacillus plantarum R315. J. Dairy Sci. 2014, 97, 7334–7343. [Google Scholar] [CrossRef]
- Sirin, S.; Aslim, B. Characterization of lactic acid bacteria derived exopolysaccharides for use as a defined neuroprotective agent against amyloid beta1–42-induced apoptosis in SH-SY5Y cells. Sci. Rep. 2020, 10, 8124. [Google Scholar] [CrossRef]
- Gientka, I.; Błażejak, S.; Stasiak-Różańska, L.; Chlebowska-Śmigiel, A. Exopolysaccharides from yeast: Insight into optimal conditions for biosynthesis, chemical composition and functional properties-review. Acta Sci. Pol. Technol. Aliment. 2015, 14, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Ayyash, M.; Abu-Jdayil, B.; Itsaranuwat, P.; Galiwango, E.; Tamiello-Rosa, C.; Abdullah, H.; Esposito, G.; Hunashal, Y.; Obaid, R.S.; Hamed, F. Characterization, bioactivities, and rheological properties of exopolysaccharide produced by novel probiotic Lactobacillus plantarum C70 isolated from camel milk. Int. J. Biol. Macromol. 2020, 144, 938–946. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-C.; Hsieh, Y.-H.; Hsu, C.-C.; Lin, C.-H.; Chen, Y.-H. Exopolysaccharide is the potential effector of Lactobacillus fermentum PS150, a hypnotic psychobiotic strain. Front. Microbiol. 2023, 14, 1209067. [Google Scholar] [CrossRef]
- Flores-Maciel, M.G.; Ochoa-Martínez, A.; Rutiaga-Quiñones, O.M.; Rutiaga-Quiñones, J.G. Importance of lactic acid bacteria as producers of exopolysaccharides. Agraria 2024, 1, 2. [Google Scholar]
- Zhang, J.; He, J.; Hu, J.; Ji, Y.; Lou, Z. Exploring the role of gut microbiota in depression: Pathogenesis and therapeutic insights. Asian J. Psychiatry 2025, 105, 104411. [Google Scholar] [CrossRef]
- Utama, G.L.; Sahab, N.R.M.; Nurmilah, S.; Yarlina, V.P.; Subroto, E.; Balia, R.L. Unveiling microbial dynamics in terasi spontaneous fermentation: Insights into glutamate and GABA production. Curr. Res. Food Sci. 2025, 10, 100950. [Google Scholar] [CrossRef]
- Chintakovid, N.; Singkhamanan, K.; Yaikhan, T.; Nokchan, N.; Wonglapsuwan, M.; Jitpakdee, J.; Kantachote, D.; Surachat, K. Probiogenomic analysis of Lactiplantibacillus plantarum SPS109: A potential GABA-producing and cholester-ol-lowering probiotic strain. Heliyon 2024, 10, e33823. [Google Scholar] [CrossRef]
- Karpiński, P.; Żebrowska-Różańska, P.; Kujawa, D.; Łaczmański, Ł.; Samochowiec, J.; Jabłoński, M.; Plichta, P.; Piotrowski, P.; Bielawski, T.; Misiak, B. Gut microbiota alterations in schizophrenia might be related to stress exposure: Findings from the machine learning analysis. Psychoneuroendocrinology 2023, 155, 106335. [Google Scholar] [CrossRef]
- Malan-Müller, S.; Martín-Hernández, D.; Caso, J.R.; Matthijnssens, J.; Rodríguez-Urrutia, A.; Lowry, C.A.; Leza, J.C. Metagenomic symphony of the intestinal ecosystem: How the composition affects the mind. Brain Behav. Immun. 2025, 123, 510–523. [Google Scholar] [CrossRef]
- Feng, J.; Cen, Q.; Cui, Y.; Hu, X.; Li, M.; Wang, L.; Wei, J.; Sun, N.; Wang, J.; Zhang, A. Lactobacillus rhamnosus: An emerging probiotic with therapeutic potential for depression. Pharmacol. Res. 2025, 211, 107541. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.W.; Vemuganti, V.; Kuehn, J.F.; Ulland, T.K.; Rey, F.E.; Bendlin, B.B. Gut microbial metabolism in Alzheimer’s disease and related dementias. Neurotherapeutics 2024, 21, e00470. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Nasser, A.; Hathout, A.S.; Badr, A.N.; Barakat, O.S.; Fathy, H.M. Extraction and characterization of bioactive secondary metabolites from lactic acid bacteria and evaluating their antifungal and antiaflatoxigenic activity. Biotechnol. Rep. 2023, 38, e00799. [Google Scholar] [CrossRef] [PubMed]
- Salvatore, M.M.; Maione, A.; Buonanno, A.; Guida, M.; Andolfi, A.; Salvatore, F.; Galdiero, E. Biological activities, biosynthetic capacity and metabolic interactions of lactic acid bacteria and yeast strains from traditional home-made kefir. Food Chem. 2025, 470, 142657. [Google Scholar] [CrossRef]
- De Luca, L.; Pizzolongo, F.; Calabrese, M.; Blaiotta, G.; Aponte, M.; Romano, R. Addition of glutamine to milk during fermentation by individual strains of lactic acid bacteria and the effects on pyroglutamic and butyric acid. J. Food Compos. Anal. 2024, 130, 106175. [Google Scholar] [CrossRef]
- Singh, H.; Chopra, C.; Singh, H.; Malgotra, V.; Khurshid Wani, A.; Singh Dhanjal, D.; Sharma, I.; Nepovimova, E.; Alomar, S.; Singh, R.; et al. Gut-brain axis and Alzheimer’s disease: Therapeutic interventions and strategies. J. Funct. Foods 2024, 112, 105915. [Google Scholar] [CrossRef]
- Liang, Y.; Cui, L.; Gao, J.; Zhu, M.; Zhang, Y.; Zhang, H.-L. Gut Microbial Metabolites in Parkinson’s Disease: Implications of Mitochondrial Dysfunction in the Pathogenesis and Treatment. Mol. Neurobiol. 2021, 58, 3745–3758. [Google Scholar] [CrossRef]
- Alli, S.R.; Gorbovskaya, I.; Liu, J.C.W.; Kolla, N.J.; Brown, L.; Müller, D.J. The gut microbiome in depression and potential benefit of prebiotics, probiotics and synbiotics: A systematic review of clinical trials and observational studies. Int. J. Mol. Sci. 2022, 23, 4494. [Google Scholar] [CrossRef]
- Misera, A.; Kaczmarczyk, M.; Łoniewski, I.; Liśkiewicz, P.; Podsiadło, K.; Misiak, B.; Skonieczna-Żydecka, K.; Samochowiec, J. Comparative analysis of gut microbiota in major depressive disorder and schizophrenia during hospitalisation—The case-control, post hoc study. Psychoneuroendocrinology 2025, 171, 107208. [Google Scholar] [CrossRef]
- Merchak, A.R.; Wachamo, S.; Brown, L.C.; Thakur, A.; Moreau, B.; Brown, R.M.; Rivet-Noor, C.R.; Raghavan, T.; Gaultier, A. Lactobacillus from the Altered Schaedler Flora maintain IFNγ homeostasis to promote behavioral stress resilience. Brain Behav. Immun. 2024, 115, 458–469. [Google Scholar] [CrossRef]
- Zhang, P.; Jin, W.; Lyu, Z.; Lyu, X.; Li, L. Study on the mechanism of gut microbiota in the pathogenetic interaction between depression and Parkinson ‘s disease. Brain Res. Bull. 2024, 215, 111001. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Chen, Y.; Wang, Z.; Wang, R.; Dong, Y. Lactiplantibacillus plantarum CR12 attenuates chronic unforeseeable mild stress induced anxiety and depression-like behaviors by modulating the gut microbiota-brain axis. J. Funct. Foods 2023, 107, 105710. [Google Scholar] [CrossRef]
- Qian, X.; Li, Q.; Zhu, H.; Chen, Y.; Lin, G.; Zhang, H.; Chen, W.; Wang, G.; Tian, P. Bifidobacteria with indole-3-lactic acid-producing capacity exhibit psychobiotic potential via reducing neuroinflammation. Cell Rep. Med. 2024, 5, 101798. [Google Scholar] [CrossRef] [PubMed]
- Sai, Y.; Ge, W.; Zhong, L.; Zhang, Q.; Xiao, J.; Shan, Y.; Ye, W.; Liu, H.; Liu, S.; Ye, F.; et al. The role of the gut microbiota and the nicotinate/nicotinamide pathway in rotenone-induced neurotoxicity. Curr. Res. Toxicol. 2025, 8, 100212. [Google Scholar] [CrossRef]
- Wang, L.; Tsai, C.; Chou, W.; Kuo, H.; Huang, Y.; Lee, S.; Dai, H.; Yang, C.; Li, C.; Yeh, Y. Add-On Bifidobacterium bifidum supplement in children with attention-deficit/hyperactivity disorder: A 12-Week randomized double-blind placebo-controlled clinical trial. Nutrients 2024, 16, 2260. [Google Scholar] [CrossRef]
- Tanure, Y.C.B.; Mafra, A.C.M.; Guimarães, B.L.M.; Magalhães, R.C.; Fagundez, C.; Nascimento, I.J.B.D.; Brito, J.C.M. Potential benefits of kefir and its compounds on Alzheimer’s Disease: A systematic review. Brain Behav. Immun. Integr. 2025, 10, 100115. [Google Scholar] [CrossRef]
- Dhami, M.; Raj, K.; Singh, S. Relevance of gut microbiota to Alzheimer’s Disease (AD): Potential effects of probiotic in management of AD. Aging Health Res. 2023, 3, 100128. [Google Scholar] [CrossRef]
- Kong, Q.; Chen, Q.; Mao, X.; Wang, G.; Zhao, J.; Zhang, H.; Chen, W. Bifidobacterium longum CCFM1077 ameliorated neurotransmitter disorder and neuroinflammation closely linked to regulation in the kynurenine pathway of autistic-like rats. Nutrients 2022, 14, 1615. [Google Scholar] [CrossRef]
- Miyanishi, H.; Nitta, A. A role of BDNF in the depression pathogenesis and a potential target as antidepressant: The modulator of stress sensitivity “Shati/Nat8l-BDNF system” in the dorsal striatum. Pharmaceuticals 2021, 14, 889. [Google Scholar] [CrossRef]
- Vergel Hernández, J.; Barrera Robledo, M.E. Management of depressive disorder: Which treatment should I choose? Med. J. Risaralda 2021, 27, 24637. [Google Scholar] [CrossRef]
- Miyaoka, T.; Kanayama, M.; Wake, R.; Hashioka, S.; Hayashida, M.; Nagahama, M.; Okazaki, S.; Yamashita, S.; Miura, S.; Miki, H.; et al. Clostridium butyricum MIYAIRI 588 as adjunctive therapy for treatment-resistant major depressive disorder: A prospective open-label trial. Clin. Neuropharmacol. 2018, 41, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Carrizo, S.L.; de Moreno de LeBlanc, A.; LeBlanc, J.G.; Rollán, G.C. Quinoa pasta fermented with lactic acid bacteria prevents nutritional deficiencies in mice. Food Res. Int. 2020, 127, 108735. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Liu, Z.; Xie, F.; Bilal, M.; Liu, L.; Yang, R.; Wang, Z. Microbial production of gamma-aminobutyric acid: Applications, state-of-the-art achievements, and future perspectives. Crit. Rev. Biotechnol. 2021, 41, 491–512. [Google Scholar] [CrossRef] [PubMed]
- Di Salvo, C.; D’Antongiovanni, V.; Benvenuti, L.; d’Amati, A.; Ippolito, C.; Segnani, C.; Pierucci, C.; Bellini, G.; Annese, T.; Virgintino, D.; et al. Lactiplantibacillus plantarum HEAL9 attenuates cognitive impairment and progression of Alzheimer’s disease and related bowel symptoms in SAMP8 mice by modulating microbiota-gut-inflammasome-brain axis. Food Funct. 2024, 15, 10323–10338. [Google Scholar] [CrossRef]
- Bravo, J.; Forsythe, P.; Chew, M.; Escaravage, E.; Savignac, H.; Dinan, T.; Bienenstock, J.; Cryan, J. Ingestion of Lactobacillus strain regulates emotional behavior and central GABA receptor expression in a mouse via the vagus nerve. Proc. Natl. Acad. Sci. USA 2011, 108, 16050–16055. [Google Scholar] [CrossRef]
- Van Son, J.; Koekkoek, L.L.; La Fleur, S.E.; Serlie, M.J.; Nieuwdorp, M. The role of the gut microbiota in the gut–brain axis in obesity: Mechanisms and future implications. Int. J. Mol. Sci. 2021, 22, 2993. [Google Scholar] [CrossRef]
- Chakrabarti, A.; Geurts, L.; Hoyles, L.; Iozzo, P.; Kraneveld, A.D.; La Fata, G.; Miani, M.; Patterson, E.; Pot, B.; Shortt, C.; et al. The microbiota–gut–brain axis: Pathways to better brain health. Perspectives on what we know, what we need to investigate and how to put knowledge into practice. Cell. Mol. Life Sci. 2022, 79, 80. [Google Scholar] [CrossRef]
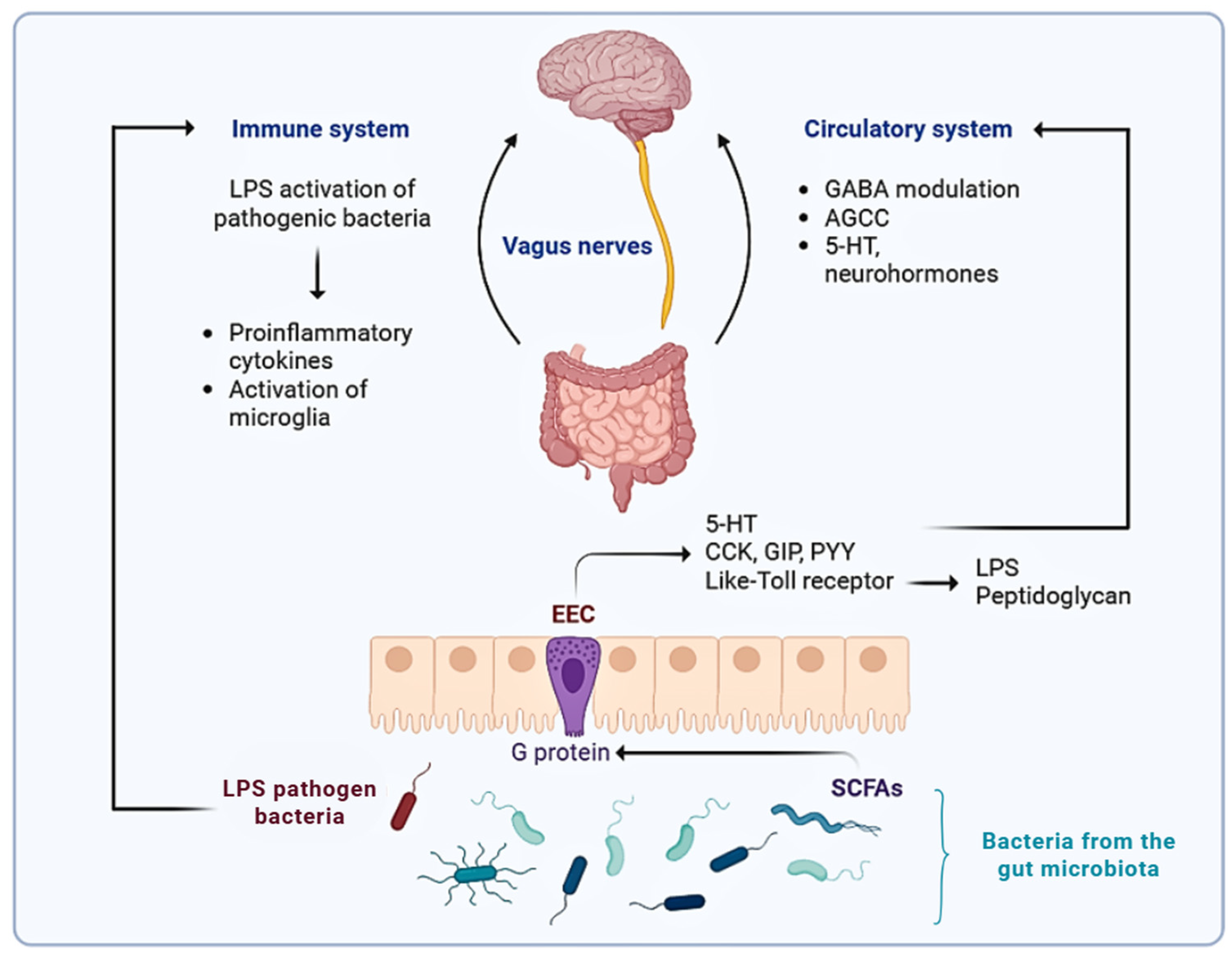

{kind=link}
| LAB | Matrix | Fermentation Effects | References |
|---|---|---|---|
| Lactiplantibacillus plantarum | Opuntia ficus-indica juice (OFIJI) | Significant increase in total phenols and anthocyanins. Antioxidant capacity increased by 16.81% (ABTS+) and 23.62% (DPPH). Sugar content reduced by 30.77%. | [13] |
| L. plantarum CET 9567 | Avocado seed | Inhibitory activity of the alpha-amylase enzyme. | [14] |
| L. plantarum JYLP-375 | Huyou peel and pomace (Citrus aurantium) | Bioconversion of organic acids, flavonoid production, and enhanced absorption. | [15] |
| L. plantarum | Black soybean seed | Increased acid production, enhanced antioxidant activity, and higher levels of aglycone isoflavones. | [16] |
| L. Plantarum | Chickpea milk | Improved color and lactic acid content, reduced beany flavor, enhanced overall flavor. | [17] |
| Latilactobacillus sakei | Lean pork and pork fat sausages (8:2) | Probiotic starter potential with antibacterial activity against E. coli and S. aureus. | [4] |
| Lacticaseibacillus rhamnosus | Yogurt-type drink with Arabic gum (GA) | Improvement in chemical, sensory, and rheological properties. | [18] |
| L. rhamnosus | Marshmallows with honey | Enhanced food stability; limited probiotic viability. | [19] |
| L. rhamnosus | Camel milk | Greater pH reduction during storage; significant increase in antioxidant activity and improved cell preservation. | [20] |
| L. rhamnosus GG | Vegetables | Suppression of pathogenic bacteria and enhancement of antioxidant activity. | [21] |
| Lactobacillus gasseri SM 05 | Fermented black raspberry juice | Increased gallic, ferulic, and cinnamic acid levels; enhanced antioxidant activity. | [22] |
| L. gasseri | Passion fruit juice with green tea | Functional activity. | [23] |
| Lactobacillus helveticus DQHXN-Q32M42 | Fermented milk | Peptide production and diversity of bioactive compounds. | [24] |
| L. helveticus B734 | Drinkable dairy beverage | Release of bioactive peptides. | [25] |
| L. helveticus MDC 9602 | Dairy product with apricot gum | Increased shelf life, higher antioxidant activity, increased acidity, and improved sensory properties. | [26] |
| Lacticaseibacillus casei DSM 20011 (ATCC-393) | Olive leaves | Lactic acid production. | [27] |
| L. casei | Blackberry juice | Increased phenolic content and antioxidant activity. | [28] |
| Levilactobacillus brevis | Trigonotis radicans var. sericea | Bioconversion of bioactive compounds, enhanced antioxidant activity (DPPH and ABTS); inhibitory activity against tyrosinase, elastase, and collagenase. | [29] |
| L. brevis | Betaphycus gelatinum | Significant increases in absorbable polyphenol and antioxidant activity. | [30] |
| Pediococcus acidilactici | Avocado leaves | Significant increase in bioactive compound content. | [31] |
| Pediococcus pentosaceas | Lean pork and pork fat sausages (8:2) | Probiotic starter potential; enhanced antioxidant and enzymatic antioxidant activity. | [4] |
| Limosilactobacillus reuteri R29 | Barley rootlets | Greater conversion of fructose to mannitol, reducing sugar contents; increased fiber content. | [32] |
| Leuconostoc citreum TR116 | Barley rootlets | Greater conversion of fructose to mannitol, reducing sugar contents. | [32] |
| L. plantarum | Lean pork and pork fat sausages (8:2) | Probiotic starter potential with antibacterial (against E. coli and S. aureus) and antioxidant activity; enzymatic antioxidant activity. | [4] |
| SCFA | Role in the Microbiota–Gut–Brain Axis | References |
|---|---|---|
| Acetate | Modulation of neuroimmune and neuroendocrine signaling pathways. Supports microglial development and functional maturation. Reduces pro-inflammatory cytokine activity and supports anti-inflammatory signaling. Maintains intestinal epithelial homeostasis and barrier integrity. Regulates central acid–base balance and supports CNS homeostatic mechanisms. | [49,53,54,55] |
| Butyrate | Promotes microglial homeostasis and anti-inflammatory phenotype. Stimulates intestinal hormone secretion (GLP-1, PYY). Exhibits neuroprotective effects via inhibition of oxidative stress and apoptosis. Restores intestinal permeability and enhances tight junction protein expression. Upregulates brain-derived neurotrophic factor (BDNF), supporting memory and synaptic plasticity. | [54,55,56] |
| Propionate | Contributes to blood–brain barrier (BBB) integrity and protection. Modulates neuroinflammatory responses by downregulating microglial activation and Th17 proliferation. Participates in metabolic regulation, including glucose and lipid metabolism. Enhances intestinal barrier function and reduces oxidative stress. Regulates neuronal excitability and synaptic signaling. | [4,55,57] |
| Lactate | Neuroimmune–endocrine regulation. Gut barrier integrity. | [4,58] |
| Metabolite | Properties | Disease Control | LAB That Produce It | Reference |
|---|---|---|---|---|
| GABA | Inhibitory neurotransmitter, improves vagus nerve conduction velocity and neuronal activity. | L. rhamnosus | [80] | |
| Antidepressant, antidiabetic, and antihypertensive. | Epilepsy, schizophrenia, depression, anxiety, chronic pain, and neurodevelopmental disorders. | [81] | ||
| Calming effects on the nervous system. | L. plantarum SPS109 | [82] | ||
| SCFAs | They stimulate the secretion of glucagon 1 and peptide YY (PYY), along with GABA and serotonin. | [83] | ||
| Propionate | Antidepressant | Depression | [84] | |
| IAA | Tryptophan metabolite produced by the intestinal microbiota; helps to alleviate mood disorders. | Depression | L. rhamnosus | [85] |
| Indole | No accumulation of Aβ and hyperphosphorylation of Tau. Inhibits the activation of the NF-κB signaling pathway, NLRP3, and reduces the release of inflammatory cytokines (TNF-α, IL-6, IL-1β and IL-18). | Alzheimer’s | [61] | |
| Attenuates neuroinflammation and improves neuronal survival, neuropsychological symptoms and cognitive impairment. | [86] | |||
| Acetate | Improved cognitive health in mice, increased neuronal activation (c-FOS) in mice. | [86] | ||
| Serotonin | Decreases the effect of antidepressants on depression-like behavior. | [86] |
| Neuropsychiatric Disorder | Individuals | Factors | Ingested Bacteria | Findings | References |
|---|---|---|---|---|---|
| Schizophrenia | Gastrointestinal disorders and low cognitive test scores. | L. rhamnosus strain GG and Bifidobacterium animalis subsp. lactis Bb12. | Reduced Candida albicans. | [84] | |
| 36 patients with the conditions and 29 healthy controls (HC). | Hypothalamic–pituitary–adrenal axis dysfunction. | Patients with the condition presented with alterations in the microbiota, attributed to bacteria associated with inflammation. | [93] | ||
| Total 142 participants (89 schizophrenia and 52 controls). | Psychosocial stress, malnutrition, lipid profile alterations, and cognitive impairment. | Microbiota (Major Lactobacillus and Limosilactobacillus, minor Faecalibacterium and Paraprevotella). | [83] | ||
| Depression | Patients with myocardial infarction. | L. rhamnosus (12 weeks). | Improvements in depressive symptoms and quality of life. Beneficial effects on objective markers such as 5-HT, PCR, TIC, LPS, and TNF-α. | [85] | |
| Depressed rats | L. rhamnosus and Bifidobacterium longum | Increased 5-HT and TPH2 levels in frontal cortex, IAA and indole production, and decreased IDO levels. | [85] | ||
| Gnotobiotic mouse colonized with Schaedler’s altered flora. | Lactobacillus | Protective against environmental stressors, maintains homeostatic levels of IFNγ related to behavior, increased numbers and activity of Th1 cells. | [94] | ||
| Intestinal microbial dysbiosis. | Increased intestinal permeability, secretion of inflammatory cytokines (TNF-α and IL-6), pathological changes in the prefrontal cortex and hippocampal region. | [95] | |||
| Mice | Exogenous sodium butyrate alters the expression of the brain-derived neurotrophic factor gene related to depression. | Lactobacillus | Increased production of neurotransmitters (GABA and serotonin), due to the action of their secondary metabolites, including SCFAs, indole, p-aminobenzoate, α-tocopherol, secondary bile acids, and tyramine. | [90] | |
| Mice | Mild stress environments. | L. plantarum CR12 | Improvement in cognitive level and spatial memory in open field tests, attenuation of anxious and obsessive behaviors by increasing the formation of butyrate. In addition, there was an increase in Lactobacillus and a reduction in Helicobacter pylori in the intestinal microbiota. | [96] | |
| Mice | Depressed mice with chronic stress. | Bifidobacterium breve CCFM1025 | Increased levels of indole-3-lactic acid (ILA) in the circulatory system, which may alleviate neuroinflammation and improve mood disorders. | [97] | |
| Parkinson’s disease | Rats (20: Control; 20: Rotenona; R) | Rotenone generates neurotoxic effects in humans. | Group R: Acetic acid and butyric acid in intestinal tissues decreased, as did NAD+, while IL-6, TNF-α, and VIP increased. | [98] | |
| Attention deficit hyperactivity disorder (ADHD) | Children with ADHD on stable methylphenidate treatment. | Bifidobacterium bifidum Bf-688 | Improves neuropsychological performance in children with ADHD by altering gut microbiota and reducing N-glycan biosynthesis. | [99] | |
| Alzheimer’s disease | Rats | Lactobacillus | Decreased cognitive deficits, anxious behaviors, and neuronal degeneration. Reduced Aβ accumulation in the brain and microglia activation, and also generated inhibitory effects on Aβ42 aggregation and amyloid-induced cytotoxicity. | [100] | |
| Mice | Lactobacillus | Increased memory, learning, and hippocampal level of IDE, decreased hippocampal Aβ levels, neutrophil infiltration, nitrotyrosine formation, NF-κB activation, and inducible nitric oxide synthase expression. | [100] | ||
| Albino rats | Administration of d-galactose to induce AD. | Lactobacillus | Regulation of brain metabolites (GABA and glutamate) associated with the cortex and hippocampus, and beneficial for propagating neuronal signaling and improving memory. They reduce the accumulation of Aβ. | [101] | |
| Mice | Lactobacillus | Decreased inflammation and intestinal permeability, with minimal effects on cytokines and beta-amyloid in the brain. | [101] | ||
| Autism spectrum | Rats | Bifidobacterium logum CCFM077 | Improves gut microbiota composition and regulates neurotransmitter levels and neuroinflammation in autistic rats. | [102] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aguirre-Garcia, Y.L.; Cerda-Alvarez, N.C.; Santiago-Santiago, R.M.; Chantre-López, A.R.; Rangel-Ortega, S.D.C.; Rodríguez-Herrera, R. The Metabolites Produced by Lactic Acid Bacteria and Their Role in the Microbiota–Gut–Brain Axis. Fermentation 2025, 11, 378. https://doi.org/10.3390/fermentation11070378
Aguirre-Garcia YL, Cerda-Alvarez NC, Santiago-Santiago RM, Chantre-López AR, Rangel-Ortega SDC, Rodríguez-Herrera R. The Metabolites Produced by Lactic Acid Bacteria and Their Role in the Microbiota–Gut–Brain Axis. Fermentation. 2025; 11(7):378. https://doi.org/10.3390/fermentation11070378
Chicago/Turabian StyleAguirre-Garcia, Yulma Lizbeth, Neftiti Carolina Cerda-Alvarez, Rosa María Santiago-Santiago, Adriana Rocío Chantre-López, Sarahi Del Carmen Rangel-Ortega, and Raúl Rodríguez-Herrera. 2025. "The Metabolites Produced by Lactic Acid Bacteria and Their Role in the Microbiota–Gut–Brain Axis" Fermentation 11, no. 7: 378. https://doi.org/10.3390/fermentation11070378
APA StyleAguirre-Garcia, Y. L., Cerda-Alvarez, N. C., Santiago-Santiago, R. M., Chantre-López, A. R., Rangel-Ortega, S. D. C., & Rodríguez-Herrera, R. (2025). The Metabolites Produced by Lactic Acid Bacteria and Their Role in the Microbiota–Gut–Brain Axis. Fermentation, 11(7), 378. https://doi.org/10.3390/fermentation11070378