

The Effects of Solid-State Fermentation by Aspergillus spp. on the Nutritional Profile of Selected Agro-Industrial by-Products as Potential Feedstuffs for Weaner Rabbits

 , , , , ,
, , , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Feedstuffs Used as Substrate for Solid-State Fermentation
2.2. Source of Microorganism and Inoculum Preparation
2.2.1. Molecular Characterization of Selected Isolates
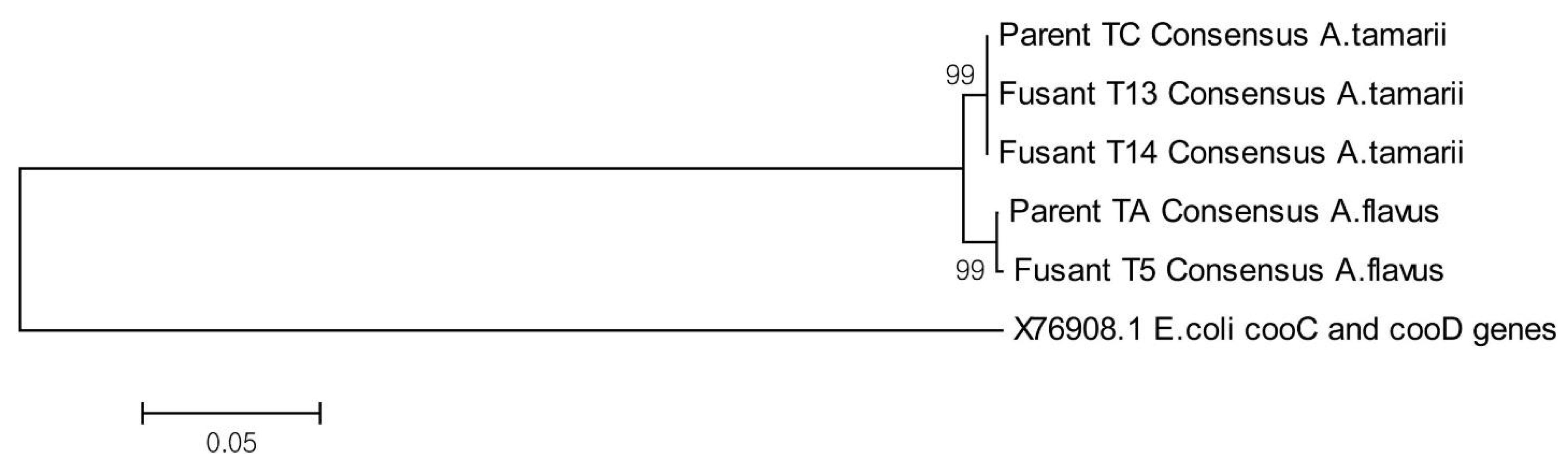
2.2.2. Molecular Identification and Phylogenetic Tree Representation of Sequences
2.2.3. Screening for Cellulolytic Properties of Isolates
2.3. Solid-State Fermentation of Agro-Industrial By-Products (AIBPs) by Selected Cellulolytic Fungi
2.4. Chemical Analyses
2.4.1. Mineral Composition
2.4.2. Anti-Nutritional Factors
2.4.3. Proximate Composition and Fibre Fractions
2.5. Statistical Analyses
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AIBP | Agro-industrial by-product |
| ANOVA | Analysis of variance |
| CP | Crude protein |
| CF | Crude fibre |
| NFE | Nitrogen-free extract |
| EE | Ether extract |
| ADF | Acid detergent fibre |
| NDF | Neutral detergent fibre |
| ADL | Acid detergent lignin |
| SSF | Solid-state fermentation |
| CS | Cowpea shell |
| GS | Groundnut shell |
| MS | Maize shaft |
| SH | Soybean hull |
| PCR | Polymerase chain reaction |
| DE | Digestible energy |
| GED | Gross energy digestibility coefficient |
| SDA | Sabouraud dextrose agar |
References
- Sadh, P.K.; Duhan, S.; Duhan, J.S. Agro-industrial wastes and their utilization using solid state fermentation: A review. BIOB 2018, 5, 1. [Google Scholar] [CrossRef]
- Dhingra, D.; Michael, M.; Rajput, H.; Patil, R.T. Dietary fibre in foods: A review. J. Food Sci. Technol. 2021, 49, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.H.; His, H.Y.; Leu, J.Y.; Lee, T.H. Cereal-based solid-state fermentation and anti-inflammatory potential of Paecilomyces cicadae SH1 (Ascomycota) isolated in China. Int. J. Med. Mushrooms. 2022, 24, 73–84. [Google Scholar] [CrossRef]
- Liu, Y.; Li, C.; Meng, X.; Yan, Y. Biodiesel synthesis directly catalyzed by the fermented solid of Burkholderia cenocepacia via solid state fermentation. Fuel Process. Technol. 2013, 106, 303–309. [Google Scholar] [CrossRef]
- Zhu, X.; Chen, Y.; Hao, S.; Jin, S.; Li, X. Improvement of the Nutritional Quality of Rapeseed Meal through Solid-State Fermentation with B. subtilis, S. cerevisiae, and B. amyloliquefaciens. Fermentation 2023, 9, 492. [Google Scholar] [CrossRef]
- Lawal, T.; Ademola, S.; Owoseni, A.; Atobatele, O.; Asuelimen, P. Aspergillus flavus degraded brewer dried grains for broiler chicken diet: Performance and nutrient digestibility parameters. J. Adv. Biol. Biotechnol. 2017, 11, 1–8. [Google Scholar] [CrossRef]
- Narsale, P.V.; Patel, S.R.; Acharya, P. Role of Aspergillus flavus on biodegradation of lignocellulosic waste millet straw and optimization parameters for enzyme hydrolysis and ethanol production under solid state fermentation. Int. J. Curr. Microbiol. 2018, 7, 429–445. [Google Scholar] [CrossRef]
- Sadh, P.K.; Saharan, P.; Duhan, S.; Duhan, J.S. Bio-enrichment of phenolics and antioxidant activity of combination of Oryza sativa and Lablab purpureus fermented with GRAS filamentous fungi. Resour.-Effic. Technol. 2017, 3, 347–352. [Google Scholar] [CrossRef]
- Manavalan, T.; Manavalan, A.; Heese, K. Characterization of lingo-cellulolytic enzymes from white-rot fungi. Curr. Trends Microbiol. 2015, 70, 485–498. [Google Scholar] [CrossRef]
- Fonseca, M.I.; Fariña, J.I.; Castrillo, M.L.; Rodríguez, M.D.; Nuñez, C.E.; Villalba, L.L.; Zapata, P.D. Bio pulping of wood chips with Phlebia brevispora BAFC 633 reduces lignin content and improves pulp quality. Int. Biodeterior. Biodegrad. 2014, 90, 29–35. [Google Scholar] [CrossRef]
- Matei, J.C.; Oliveira, J.A.; Pamphile, J.A.; Polonio, J.C. Agro-industrial wastes for biotechnological production as potential substrates to obtain fungal enzymes. Ciência Natura. 2021, 43, e72. [Google Scholar] [CrossRef]
- Fan, W.; Huang, X.; Liu, K.; Xu, Y.; Chi, Z. Advanced upcycling of agro-industrial co-products of corn via different microorganisms. Biomass Bioenerg. 2023, 168, 106669. [Google Scholar] [CrossRef]
- Adeleye, T.M.; Kareem, S.O.; Olufunmilayo, B.M.; Atanda, O.; Osho, M.B.; Dairo, O. Studies on Improved Amylases Developed by Protoplast Fusion of Aspergillus species. Microbiol. Biotechnol. Lett. 2020, 49, 45–56. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The Neighbor-Joining Method: A New Method for Reconstructing Phylogenetic Trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Coronado-Ruiz, C.; Avendaño, R.; Escudero-Leyva, E.; Conejo-Barboza, G.; Chaverri, P.; Chavarría, M. Two new cellulolytic fungal species isolated from a 19th-century art collection. Sci. Rep. 2018, 8, 7492. [Google Scholar] [CrossRef]
- Pensupa, N.; Jin, M.; Kokolski, M.; Archer, D.B.; Du, C.A. Solid state fungal fermentation-based strategy for the hydrolysis of wheat straw. Bioresour. Technol. 2013, 149, 261–267. [Google Scholar] [CrossRef] [PubMed]
- AOAC—Association of Analytical Chemists. Official Methods of Analysis, 15th ed.; Helrich, K., Ed.; AOAC: Arlington, VA, USA, 1990. [Google Scholar]
- Dahal, P.; Tamang, M.K. Effects of Different Processing Methods on Anti-Nutritional Factors of Cassava (Manihot esculenta crantz). J. Food Nutr. Disodr. 2021, 10, 5. [Google Scholar]
- AOAC—Association of Analytical Chemists. Official Methods of Analysis, 17th ed.; Horwitz, W., Ed.; AOAC: Washington, DC, USA, 2002. [Google Scholar]
- Prokopet, G.; Unlenbruck, K.W. Protectine eine nen kalsse anti-kowperahlich verbindungen dish. Ges. Heit. 2002, 23, 318. [Google Scholar]
- AOAC. Official Method of Analysis, 18th ed.; Association of Official Analytical Chemistry: Arlington, VA, USA, 2005. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for Dietary Fiber, Neutral Detergent Fiber, and Nonstarch Polysaccharides in Relation to Animal Nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Sun, D.; Chen, X.; Zhu, C. Physicochemical properties and antioxidant activity of pectin from hawthorn wine pomace: A comparison of different extraction methods. Int. J. Biol. Macromol. 2020, 158, 1239–1247. [Google Scholar] [CrossRef] [PubMed]
- De Blas, C.; Wiseman, J.; Fraga, M.J.; Villamide, M.J. Prediction of the digestible energy and digestibility of gross energy of feeds for rabbits. 2. Mixed diets. Anim. Feed Sci. Technol. 1992, 39, 39–59. [Google Scholar] [CrossRef]
- Wiseman, J.; Villamide, M.J.; De Blas, C.; Carabaño, M.J.; Carabaño, R. Prediction of the digestible energy and digestibility of gross energy of feeds for rabbits. 1. Individual classes of feeds. Anim. Feed Sci. Technol. 1992, 39, 27–38. [Google Scholar] [CrossRef]
- Chaves-Lopez, C.; Serio, A.; Grande-Tovar, C.D.; Cuervo-Mulet, R.; Delgado-Ospina, J.; Paparella, A. Traditional Fermented Foods and Beverages from a Microbiological and Nutritional Perspective: The Colombian Heritage. Compr. Rev. Food Sci. Food Saf. 2014, 13, 1031–1048. [Google Scholar] [CrossRef]
- Torre, M.; Rodriguez, A.R.; Saura-Calixto, F. Effects of dietary fiber and phytic acid on mineral availability. Crit. Rev. Food Sci. Nutr. 1991, 30, 1–22. [Google Scholar] [CrossRef]
- Sousa, D.; Moset, V.; López-Luján, M.C.; Salgado, J.M.; Dias, A.; Belo, I.; Pascual, J.J.; Cambra-López, M. Potential of solid-state fermentation to enhance the nutritional value of oilseed cakes for poultry. Anim. Feed Sci. Technol. 2024, 316, 116056. [Google Scholar] [CrossRef]
- Samtiya, M.; Aluko, R.E.; Dhewa, T. Plant food anti-nutritional factors and their reduction strategies: An overview. Food Prod. Process. Nutr. 2020, 2, 84–90. [Google Scholar] [CrossRef]
- Amos, A.T.; Idowu, O.M.O.; Oso, A.O.; Durojaiye, O.J.; Agazue, K.; Adebowale, A.A. The chemical composition, Anti-nutritional and Microbial Properties of Ensiled Cassava Root-Leaf Blends as a Potential Feed in Swine Diet. J. Trop. Agric. Sci. 2019, 44, 1219–1235. [Google Scholar]
- Terefe, Z.K.; Omwamba, M.N.; Nduko, J.M. Effect of solid state fermentation on proximate composition, antinutritional factors and in vitro protein digestibility of maize flour. Food Sci. Nutr. 2021, 9, 6343–6352. [Google Scholar] [CrossRef]
- Vikram, A.; Woolston, J.; Sulakvelidze, A. Phage biocontrol applications in food production and processing. Curr. Issues Mol. Biol. 2021, 40, 267–302. [Google Scholar] [CrossRef]
- Habtamu, F.G.; Negussie, R. Antinutritional factors in plant foods: Potential health benefits and adverse effects. Int. J. Food Nutr. Sci. 2014, 3, 284–289. [Google Scholar] [CrossRef]
- Erdaw, M.M.; Bhuiyan, M.M.; Iji, P.A. Enhancing the nutritional value of soybeans for poultry through supplementation with new-generation feed enzymes. J. World’s Poult. Sci. 2016, 72, 307–322. [Google Scholar] [CrossRef]
- Sokolowski, K.; Turner, P.V.; Lewis, E.; Wange, R.L.; Fortin, M.C. Exploring rabbit as a nonrodent species for general toxicology studies. Toxicol. Sci. 2024, 199, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Mashamaite, L.; Ng’ambi, J.W.; Norris, D.; Ndlovu, L.R.; Mbajiorgu, C.A. Relationship Between Tannin Contents and Short-Term Biological Responses in Male Rabbits Supplemented with Leaves of Different Acacia Tree Species Grown in Limpopo province of South Africa. Livest. Res. Rural. Dev. 2009, 21, 109. Available online: http://www.lrrd.org/lrrd21/7/mash21109.htm (accessed on 22 April 2025).
- Guan, L.; Wang, K.; Gao, Y.; Li, J.; Yan, S.; Ji, N.; Ren, C.; Wang, J.; Zhou, Y.; Li, B.; et al. Biochemical and Structural Characterization of a Novel Bacterial Tannase From Lachnospiraceae bacterium in Ruminant Gastrointestinal Tract. Front. Bioeng. Biotechnol. 2021, 9, 806788. [Google Scholar] [CrossRef] [PubMed]
- Dileep, N.; Pradhan, C.; Peter, N.; Kaippilly, D.; Sashidharan, A.; Sankar, T.V. Nutritive value of guar and copra meal after fermentation with yeast Saccharomyces cerevisiae in the diet of Nile tilapia, Oreochromis niloticus. Trop. Anim. Health Prod. 2021, 53, 416. [Google Scholar] [CrossRef]
- Amoah, K.O.; Boateng, M.; Okai, D.B.; Frimpong, Y.O. Anti-nutritional factors and their relevance to monogastric animal agri-509 culture in Ghana: Some important considerations. GJAS 2018, 9, 1–15. [Google Scholar]
- Olukomaiya, O.O.; Fernando, W.C.; Mereddy, R.; Li, X.; Sultanbawa, Y. Physicochemical, microbiological and functional properties of camelina meal fermented in solid-state using food grade Aspergillus fungi. Fermentation 2020, 6, 44. [Google Scholar] [CrossRef]
- Adedayo, M.R.; Sani, A.A. Mixed-Culture Fungal Fermentation for Protease and Amylase Production from Adansonia Digitata Seed Through Solid State Technique. Covenant J. Phys. Life Sci. 2019, 7, 37–46. [Google Scholar]
- Altop, A. Effect of solid-state fermentation on main nutritional components, some minerals, condensed tannin and phenolic compounds of olive leaves. Turk. J. Agric.-Food Sci. Technol. 2019, 7, 115–119. [Google Scholar] [CrossRef]
- Sianoun, N.; Pongyeela, P.; Chungsiriporn, J. Aspergillus niger fermentation of cocoa pod husk for increased total crude protein utilizing response surface methodology. Biomass Convers. Biorefin. 2024, 15, 5313–5325. [Google Scholar] [CrossRef]
- Mbuyane, L.L.; Bauer, F.F.; Bloem, A.; Camarasa, C.; Ortiz-Julien, A.; Divol, B. Species-dependent metabolic response to lipid mixtures in wine yeasts. Front. Microbiol. 2022, 3, 823581. [Google Scholar] [CrossRef] [PubMed]
- López, A.M.; Silva, A.L.; Santos, E.C. The fungal ability for biobleaching/biopulping/bioremediation of lignin-like compounds of agro-industrial raw material. Quím.Nova. 2017, 40, 916–931. [Google Scholar] [CrossRef]
- Altop, A.; Gungor, E.; Erener, E. Improvement of nutritional quality of some oilseed meals through solid-state fermentation. Turk. J. Agric.-Food Sci. Technol. 2019, 7, 1411–1414. [Google Scholar]
- Demirgül, K.; Ozturk, E. Changes in nutrients, energy, antioxidant and carotenoid levels of dried tomato (Lycopersicon esculentum) pomage treated with Aspergillus niger solid-state fermentation. Turk. J. Agric.-Food Sci. Technol. 2021, 9, 701–708. [Google Scholar] [CrossRef]

{kind=link}
| Macro Minerals (g/kg) | Micro Minerals (mg/kg) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Ca | Mg | K | Na | Fe | Cu | Zn | Mn | |||
| Interaction | ||||||||||
| Fermentation: | Fibre source: | |||||||||
| Unfermented | CS | 3.731 c | 3.889 c | 2.110 e | 3.956 f | 95.700 f | 8.10 c | 28.40 d | 84.50 c | |
| Unfermented | GS | 1.492 g | 1.287 g | 6.759 b | 4.225 c | 103.280 e | 7.10 d | 16.00 g | 87.70 b | |
| Unfermented | MS | 0.810 h | 0.963 h | 1.309 h | 3.842 g | 11.430 h | 2.30 g | 12.90 h | 60.00 f | |
| Unfermented | SH | 2.493 e | 2.629 e | 1.381 g | 3.057 h | 128.220 c | 5.90 f | 44.30 b | 70.30 e | |
| Fermented | CS | 6.462 a | 6.109 a | 2.838 d | 4.331 b | 122.420 d | 10.10 b | 33.50 c | 97.40 a | |
| Fermented | GS | 3.382 d | 3.279 d | 8.666 a | 4.577 a | 156.88 b | 12.00 a | 25.30 e | 97.10 a | |
| Fermented | MS | 1.578 f | 1.339 f | 3.791 c | 4.017 e | 19.23 g | 6.80 e | 22.45 f | 73.60 d | |
| Fermented | SH | 5.689 b | 5.290 b | 2.056 f | 4.129 d | 177.24 a | 10.80 b | 65.10 a | 83.90 c | |
| SEM | 0.0126 | 0.0597 | 0.1855 | 0.0845 | 0.7209 | 0.0647 | 0.1083 | 0.1698 | ||
| Main effects | ||||||||||
| Fermentation: | ||||||||||
| Unfermented | 2.132 | 2.192 | 2.890 | 3.769 | 84.658 | 5.85 | 25.40 | 75.62 | ||
| Fermented | 4.277 | 4.004 | 4.338 | 4.263 | 129.94 | 9.925 | 36.58 | 88.00 | ||
| SEM | 0.5563 | 0.2989 | 0.0927 | 0.0423 | 0.3604 | 0.0323 | 0.0542 | 0.0849 | ||
| Fibre source: | ||||||||||
| CS | 5.097 | 4.999 | 2.474 | 4.144 | 109.06 | 9.10 | 30.95 | 90.95 | ||
| GS | 2.437 | 2.283 | 7.712 | 4.401 | 130.08 | 9.55 | 20.65 | 92.40 | ||
| MS | 1.194 | 1.151 | 2.551 | 3.928 | 37.33 | 4.55 | 17.67 | 66.80 | ||
| SH | 4.091 | 3.959 | 1.718 | 3.593 | 152.73 | 8.35 | 54.70 | 77.10 | ||
| SEM | 0.7867 | 0.4227 | 1.3115 | 0.5976 | 0.5097 | 0.0457 | 0.0766 | 0.1200 | ||
| p-Value | ||||||||||
| Interaction | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | ||
| Fermentation | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | 0.0002 | <0.0001 | <0.0001 | ||
| Fibre source | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | 0.0013 | <0.0001 | <0.0001 | ||
| Anti-Nutritional Factors | |||||
|---|---|---|---|---|---|
| Tannin | Phytate | TI | Oxalate | ||
| Interaction | |||||
| Fermentation: | Fibre source: | ||||
| Unfermented | CS | 6.289 c | 8.068 a | 7.270 b | 1.785 a |
| Unfermented | GS | 8.031 b | 7.017 c | 8.708 a | 1.572 b |
| Unfermented | MS | 0.716 g | 0.704 g | 0.181 f | 0.221 g |
| Unfermented | SH | 8.938 a | 7.163 b | 8.730 a | 1.536 c |
| Fermented | CS | 2.270 f | 3.512 d | 3.243 e | 0.430 d |
| Fermented | GS | 2.863 e | 2.488 e | 3.490 c | 0.255 f |
| Fermented | MS | 0.423 h | 0.247 h | 0.082 g | 0.110 h |
| Fermented | SH | 3.019 d | 2.310 f | 3.408 d | 0.287 e |
| SEM | 0.0073 | 0.0085 | 0.0106 | 0.0003 | |
| Main effects | |||||
| Fermentation: | |||||
| Unfermented | 6.018 | 5.738 | 6.222 | 1.278 | |
| Fermented | 2.143 | 2.139 | 2.556 | 0.270 | |
| SEM | 0.0036 | 0.0043 | 0.0053 | 0.0014 | |
| Fibre source: | |||||
| CS | 4.279 | 5.790 | 5.256 | 1.1076 | |
| GS | 5.497 | 4.752 | 6.099 | 0.913 | |
| MS | 0.569 | 0.475 | 0.132 | 0.166 | |
| SH | 5.978 | 4.737 | 6.069 | 0.911 | |
| SEM | 0.0052 | 0.0060 | 0.0075 | 0.0020 | |
| p-Value | |||||
| Interaction | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |
| Fermentation | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |
| Fibre source | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |
| Proximate Composition (g/kg) | Energy | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| DM | CF | CP | Ash | EE | NFE | GED 1 | DE 2 (MJ/kg) | |||
| Interaction | ||||||||||
| Fermentation: | Fibre source: | |||||||||
| Unfermented | CS | 905.00 | 175.00 c | 105.60 d | 65.20 ab | 62.38 f | 496.83 e | 0.55 e | 11.09 e | |
| Unfermented | GS | 945.00 | 444.88 a | 102.00 e | 45.00 c | 82.50 d | 270.63 h | 0.24 h | 5.08 h | |
| Unfermented | MS | 880.00 | 65.10 e | 44.13 g | 10.00 d | 2.45 h | 758.33 b | 0.67 c | 11.87 d | |
| Unfermented | SH | 882.50 | 260.00 b | 128.78 b | 62.63 b | 95.13 b | 335.98 g | 0.40 f | 9.97 f | |
| Fermented | CS | 930.43 | 115.50 d | 111.75 c | 66.65 a | 70.50 e | 566.03 d | 0.63 d | 12.71 b | |
| Fermented | GS | 951.95 | 266.55 b | 108.23 d | 41.83 c | 87.65 c | 447.70 f | 0.39 g | 9.46 g | |
| Fermented | MS | 893.33 | 58.25 f | 54.25 f | 11.75 d | 8.45 g | 760.63 a | 0.81 a | 12.25 c | |
| Fermented | SH | 892.40 | 117.90 d | 150.76 a | 65.92 ab | 103.80 a | 454.02 c | 0.72 b | 13.71 a | |
| SEM | 4.5390 | 1.8370 | 0.7120 | 0.8225 | 0.6836 | 4.3590 | 0.0017 | 0.0544 | ||
| Main effects | ||||||||||
| Fermentation: | ||||||||||
| Unfermented | 903.13 b | 236.25 | 95.13 | 45.71 | 60.61 | 465.44 | 0.46 | 9.50 | ||
| Fermented | 917.03 a | 139.55 | 103.25 | 46.54 | 67.60 | 557.09 | 0.64 | 12.03 | ||
| SEM | 2.2700 | 0.9186 | 0.3560 | 0.4112 | 0.3418 | 2.1790 | 0.0009 | 0.0385 | ||
| Fibre source: | ||||||||||
| CS | 917.52 b | 145.25 | 108.67 | 65.92 | 66.44 | 531.43 | 0.59 | 11.90 | ||
| GS | 948.5 a | 355.71 | 105.11 | 43.41 | 85.08 | 359.16 | 0.31 | 7.27 | ||
| MS | 886.65 c | 61.68 | 49.19 | 10.88 | 5.45 | 759.48 | 0.74 | 12.06 | ||
| SH | 887.45 c | 188.95 | 139.77 | 64.28 | 99.46 | 395.00 | 0.56 | 11.84 | ||
| SEM | 3.2100 | 1.2990 | 0.5034 | 0.5816 | 0.4834 | 3.0820 | 0.0012 | 0.0272 | ||
| p-Value: | ||||||||||
| Interaction | 0.2161 | <0.0001 | <0.0001 | 0.0041 | 0.0490 | <0.0001 | <0.0001 | <0.0001 | ||
| Fermentation | 0.0002 | <0.0001 | <0.0001 | 0.1658 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | ||
| Fibre source | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | ||
| Fibre Fractions | |||||||
|---|---|---|---|---|---|---|---|
| NDF | ADF | ADL | Cellulose | Hemicellulose | Pectin | ||
| Interaction | |||||||
| Fermentation: | Fibre source: | ||||||
| Unfermented | CS | 485.00 d | 263.25 d | 120.10 c | 143.20 c | 221.8 b | 5.14 b |
| Unfermented | GS | 710.00 a | 525.00 a | 284.50 a | 240.50 b | 185.00 c | 6.07 a |
| Unfermented | MS | 420.00 e | 168.25 f | 47.50 f | 120.80 d | 251.80 a | 2.78 g |
| Unfermented | SH | 595.00 b | 390.00 c | 65.10 e | 324.90 a | 205.01 b | 4.18 d |
| Fermented | CS | 310.90 f | 199.51 e | 99.30 d | 100.23 e | 111.39 e | 3.25 e |
| Fermented | GS | 549.10 c | 400.13 b | 252.92 b | 147.19 c | 149.10 d | 4.48 c |
| Fermented | MS | 251.90 h | 46.85 h | 12.00 h | 94.90 f | 145.00 e | 1.96 h |
| Fermented | SH | 298.10 g | 122.91 g | 22.70 g | 100.26 e | 175.20 c | 3.22 f |
| SEM | 3.0620 | 1.4460 | 0.8950 | 2.0460 | 3.6690 | 0.0065 | |
| Main effects | |||||||
| Fermentation: | |||||||
| Unfermented | 552.50 | 336.63 | 129.30 | 207.35 | 215.86 | 4.54 | |
| Fermented | 352.50 | 192.35 | 96.73 | 110.61 | 145.16 | 3.23 | |
| SEM | 1.5310 | 0.7229 | 0.4470 | 1.0230 | 1.8340 | 0.0032 | |
| Fibre source: | |||||||
| CS | 397.95 | 231.38 | 109.70 | 121.70 | 166.55 | 4.19 | |
| GS | 629.60 | 462.56 | 268.71 | 193.84 | 167.00 | 5.28 | |
| MS | 335.95 | 107.55 | 29.75 | 107.85 | 198.35 | 2.37 | |
| SH | 446.55 | 256.45 | 43.90 | 212.60 | 190.10 | 3.70 | |
| SEM | 2.1650 | 1.0220 | 0.6330 | 1.4470 | 2.5940 | 0.0046 | |
| p-Value | |||||||
| Interaction | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |
| Fermentation | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |
| Fibre source | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amos, A.T.; Kareem, D.U.; Adeleye, T.M.; Adeyeye, E.A.; Abatan, M.O.; Ayorinde, O.S.; Adeboye, E.O.; Sbardella, M.; Fafiolu, A.O.; Oso, A.O.; et al. The Effects of Solid-State Fermentation by Aspergillus spp. on the Nutritional Profile of Selected Agro-Industrial by-Products as Potential Feedstuffs for Weaner Rabbits. Fermentation 2025, 11, 356. https://doi.org/10.3390/fermentation11060356
Amos AT, Kareem DU, Adeleye TM, Adeyeye EA, Abatan MO, Ayorinde OS, Adeboye EO, Sbardella M, Fafiolu AO, Oso AO, et al. The Effects of Solid-State Fermentation by Aspergillus spp. on the Nutritional Profile of Selected Agro-Industrial by-Products as Potential Feedstuffs for Weaner Rabbits. Fermentation. 2025; 11(6):356. https://doi.org/10.3390/fermentation11060356
Chicago/Turabian StyleAmos, Adedoyin Titi, Damilola Uthman Kareem, Tolulope Modupe Adeleye, Emmanuel Abiodun Adeyeye, Munirat Olaide Abatan, Olusola Sarah Ayorinde, Esther Oluwasayo Adeboye, Maicon Sbardella, Adeboye Olusesan Fafiolu, Abimbola Oladele Oso, and et al. 2025. "The Effects of Solid-State Fermentation by Aspergillus spp. on the Nutritional Profile of Selected Agro-Industrial by-Products as Potential Feedstuffs for Weaner Rabbits" Fermentation 11, no. 6: 356. https://doi.org/10.3390/fermentation11060356
APA StyleAmos, A. T., Kareem, D. U., Adeleye, T. M., Adeyeye, E. A., Abatan, M. O., Ayorinde, O. S., Adeboye, E. O., Sbardella, M., Fafiolu, A. O., Oso, A. O., & Idowu, O. M. O. (2025). The Effects of Solid-State Fermentation by Aspergillus spp. on the Nutritional Profile of Selected Agro-Industrial by-Products as Potential Feedstuffs for Weaner Rabbits. Fermentation, 11(6), 356. https://doi.org/10.3390/fermentation11060356