
Pretreatment of Luzhou Distiller’s Grains with Crude Enzyme from Trichoderma harzianum for Feed Protein Production

 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Raw Materials and Microorganisms
2.2. Medium Screening
2.2.1. Spore Production Medium Screening
2.2.2. Enzyme-Producing Medium Screening
2.3. Experimental Design
2.3.1. Design of Enzyme-Producing Experiments
2.3.2. Design of Enzymatic Hydrolysis Optimization Experiments
2.3.3. Experimental Design to Produce Feed Protein from DGs Using Enzymatic Pretreatment
2.3.4. Design of In Vitro Simulated Rumen Fluid Digestion Experiment
2.3.5. Experimental Design for Lactic Acid Fermentation of Protein Feed
2.4. Methods of Measurement and Data Processing
2.4.1. Filter Paper Enzyme Activity Assay Method
2.4.2. Method for Determination of Lignocellulose Content
2.4.3. Methods for Determination of Sugar and Protein Content
2.4.4. Methods for Determination of Ammonia Nitrogen, Lactic Acid, and Volatile Fatty Acids (VFA)
2.4.5. Detection Methods Using Electronic Nose and Tongue
2.5. Statistical Analysis
3. Results
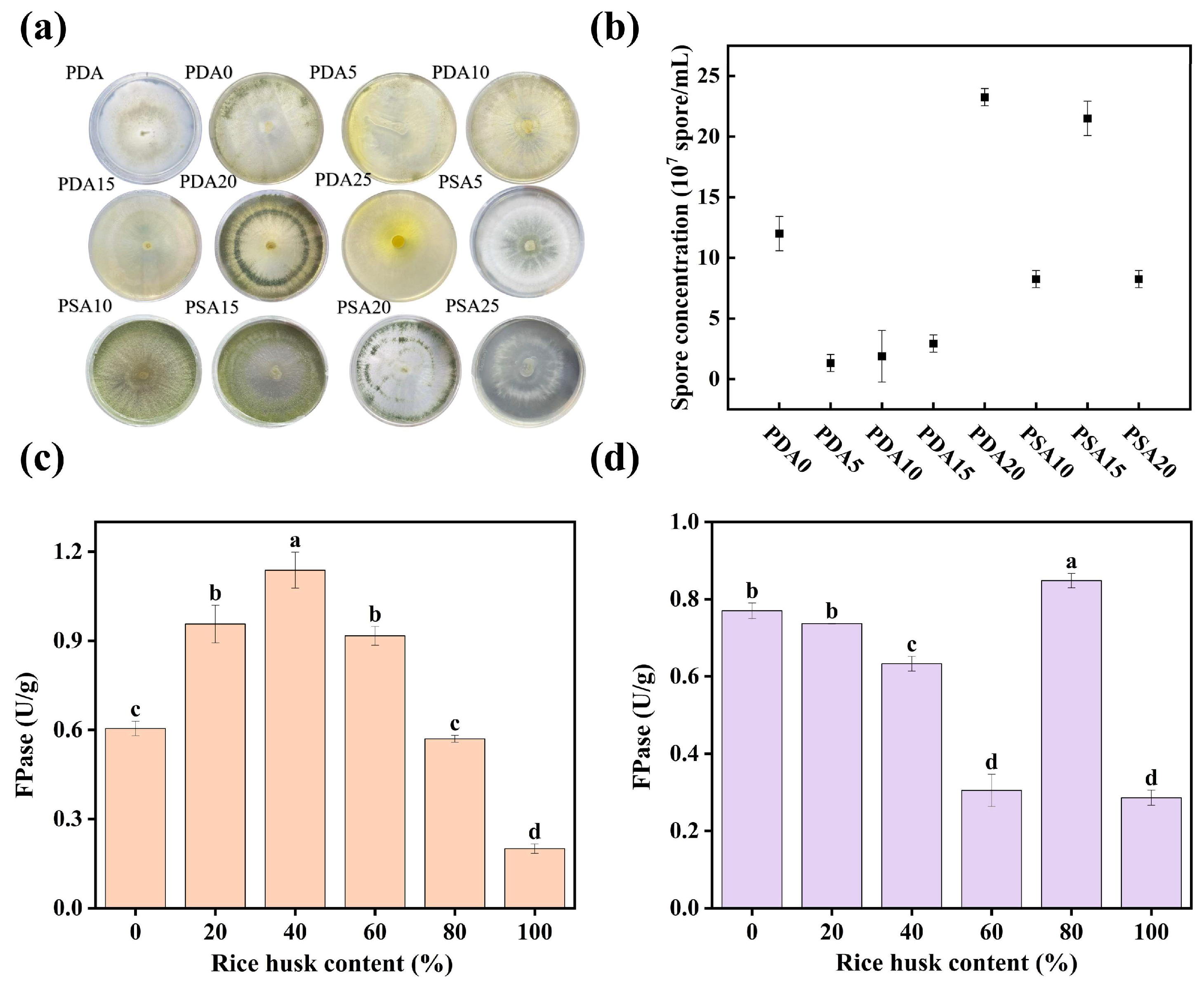
3.1. Screening of Culture Medium for Trichoderma harzianum
3.1.1. Screening of Growth Medium for Trichoderma harzianum
3.1.2. Screening of Enzyme-Producing Medium for Trichoderma harzianum
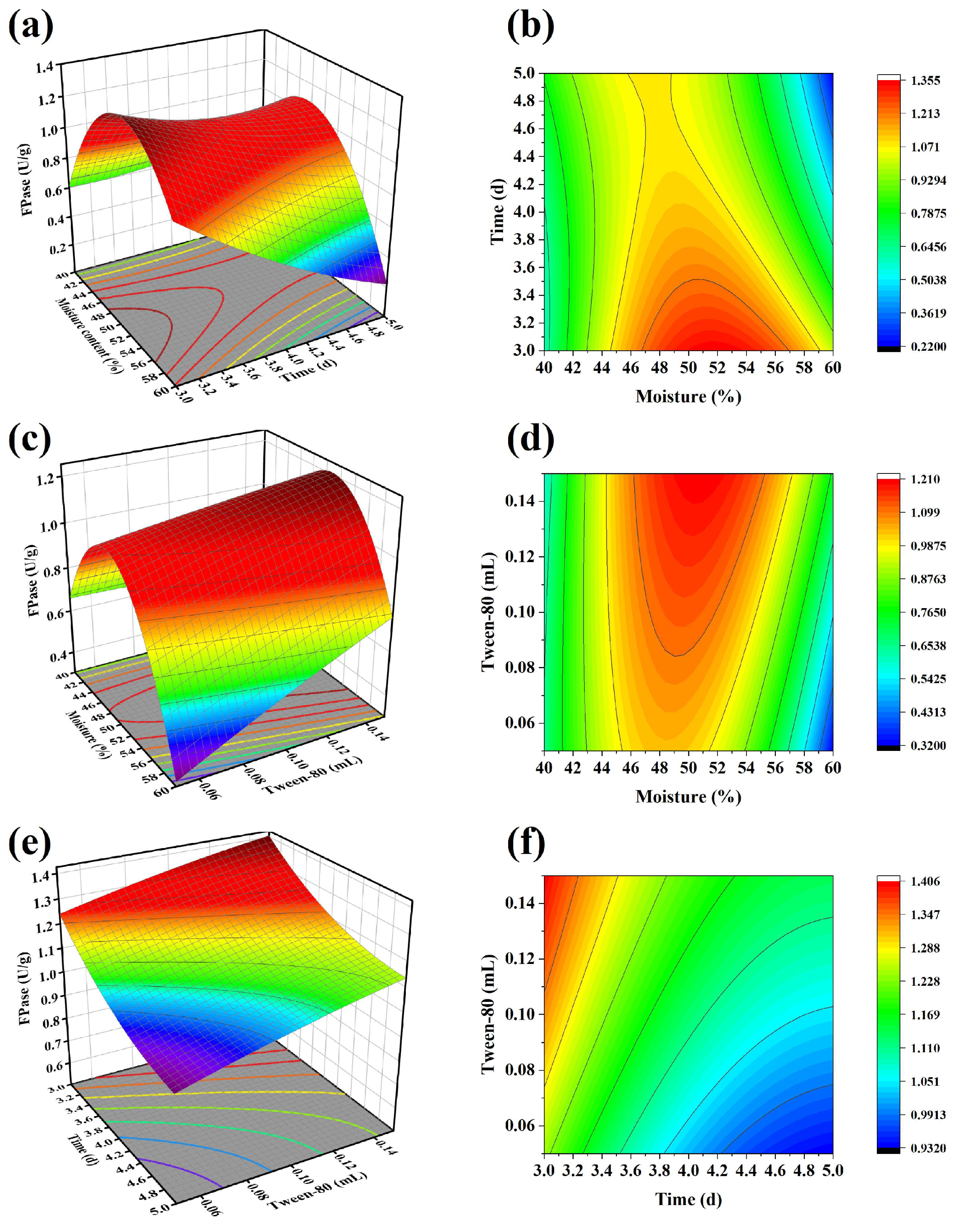
3.2. Response Surface Optimization of FPase Activity
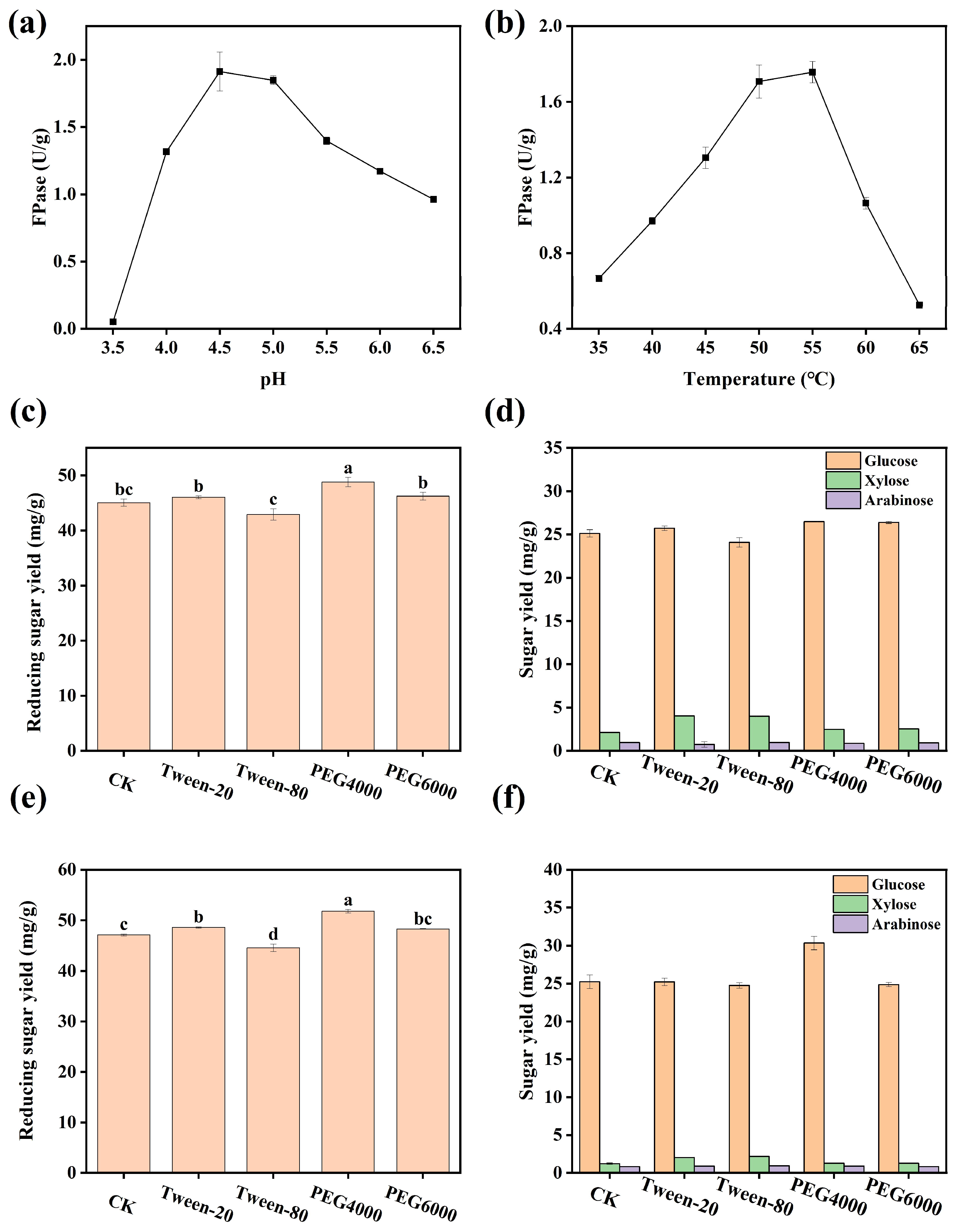
3.3. Exploration of Optimal Reaction Conditions for Enzymolysis
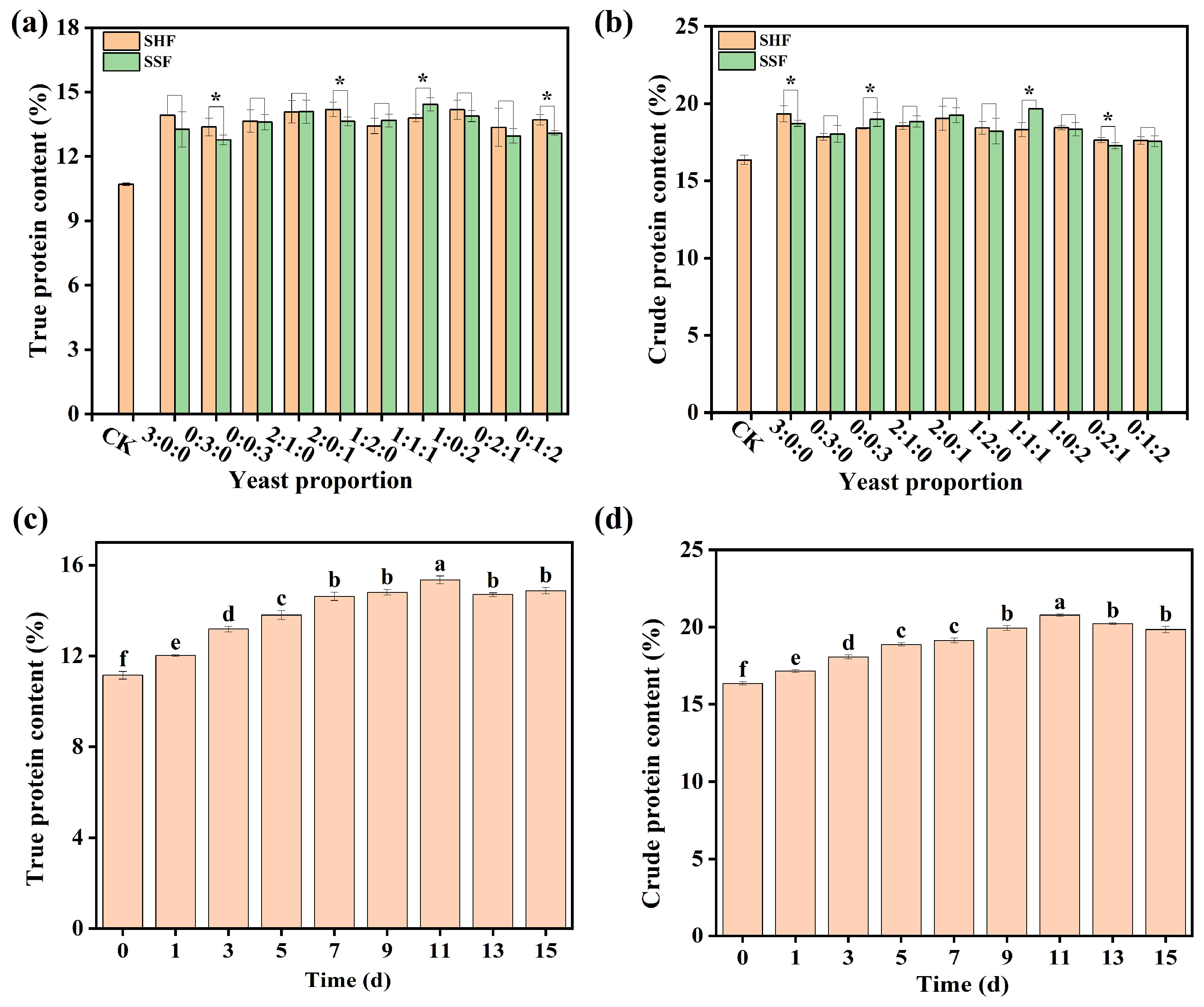
3.4. Effect of Trichoderma harzianum Fermentation on Protein Production from DGs
3.5. Electronic Sensory Characterization of Protein Feed
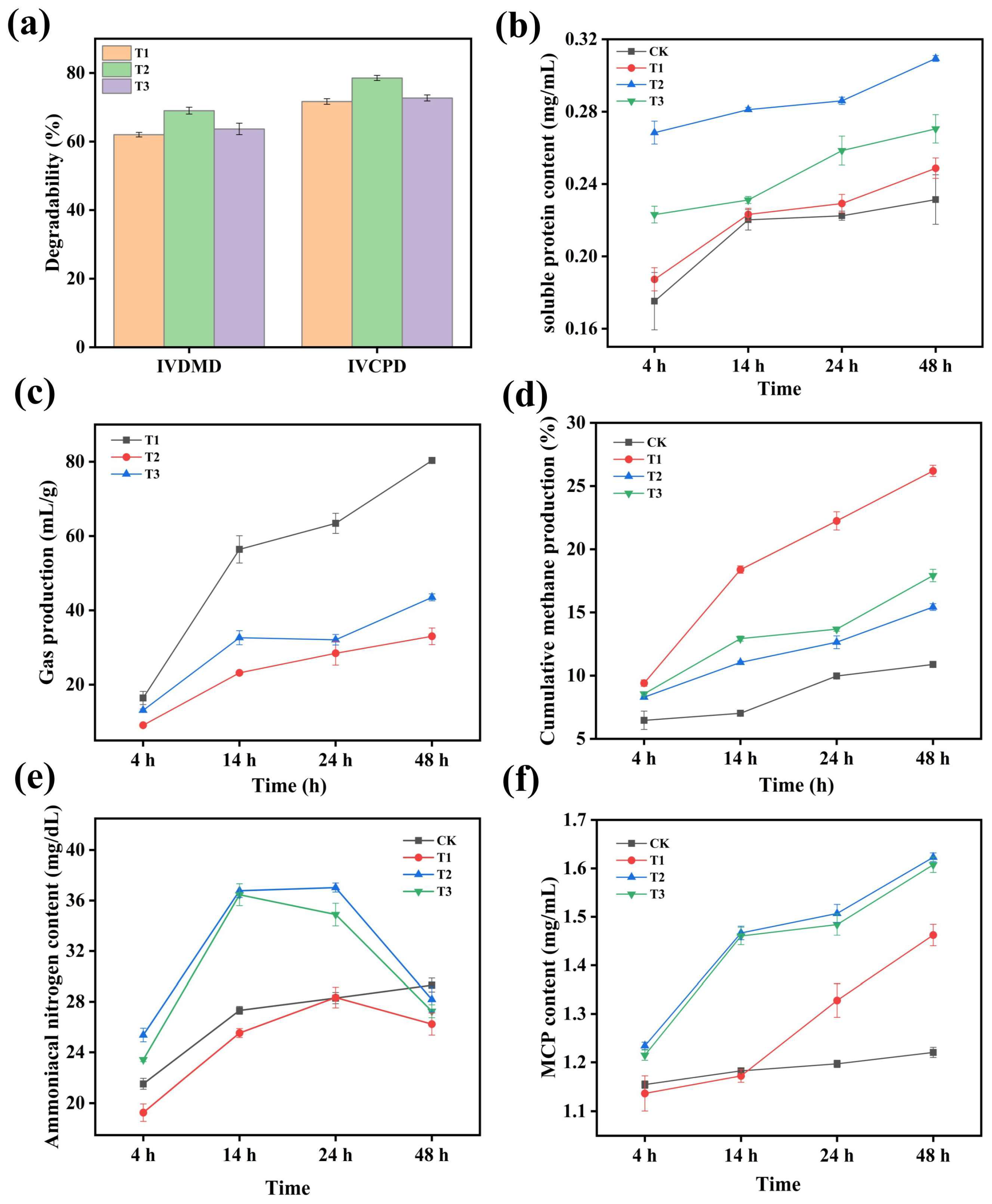
3.6. Digestibility of Protein Feed
4. Discussion
4.1. Feasibility Analysis of Cellulase Production by Trichoderma harzianum Fermentation
4.2. Feasibility Analysis of Protein Feed Production by Trichoderma harzianum–Yeast Fermentation
4.3. Performance Assessment of Protein Feed Produced by Trichoderma harzianum–Yeast Fermentation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, J.; Wang, Z.; Shen, X.; Chen, R.; Peng, Y.; Cai, Y.; Zeng, S.; Liu, D.; Yang, J.; Zhuang, W.; et al. Solid-state fermentation through synthetic microbiome: An effective strategy for converting Chinese distillers’ grains into functional protein feed. Int. J. Food Microbiol. 2025, 435, 111154. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, X.; Qin, H.; Huang, M.; Xi, B.; Mao, J.; Zhang, S. Comparing the antioxidation and bioavailability of polysaccharides from extruded and unextruded Baijiu vinasses via in vitro digestion and fecal fermentation. Int. J. Biol. Macromol. 2024, 276, 133681. [Google Scholar] [CrossRef]
- Andrade, L.C.L.; Putti, F.F.; Cremasco, C.P.; Gabriel Filho, L.R.A. New Paradigm for Vinasse Use as Fertilizer in Hydroponics. Sugar Tech 2022, 24, 1260–1271. [Google Scholar] [CrossRef]
- Fernández, F.J.; Sánchez-Arias, V.; Rodríguez, L.; Villaseñor, J. Feasibility of composting combinations of sewage sludge, olive mill waste and winery waste in a rotary drum reactor. Waste Manag. 2010, 30, 1948–1956. [Google Scholar] [CrossRef]
- Dennis, E.; Gertner, D.; Erickson, G. Economic Research on Ethanol Feed-Use Coproducts: A Review, Synthesis, and Path Forward. Animals 2024, 14, 1551. [Google Scholar] [CrossRef]
- Liu, J.; Wang, S.; Wang, Z.; Shen, C.; Liu, D.; Shen, X.; Weng, L.; He, Y.; Wang, S.; Wang, J.; et al. Pretreatment of Luzhou distiller’s grains for feed protein production using crude enzymes produced by a synthetic microbial consortium. Bioresour. Technol. 2023, 390, 129852. [Google Scholar] [CrossRef]
- Kong, S.; Wang, S.; He, Y.; Wang, N.; Wang, Z.; Weng, L.; Liu, D.; Zhao, X.; Chen, J.; Xu, J.; et al. Three-Stage Solid-State Fermentation Technology for Distillers’ Grain Feed Protein Based on Different Microorganisms Considering Oxygen Requirements. Fermentation 2024, 10, 550. [Google Scholar] [CrossRef]
- Braun, H.; Woitsch, L.; Hetzer, B.; Geisen, R.; Zange, B.; Schmidt-Heydt, M. Trichoderma harzianum: Inhibition of mycotoxin producing fungi and toxin biosynthesis. Int. J. Food Microbiol. 2018, 280, 10–16. [Google Scholar] [CrossRef]
- Lopez-Ramirez, N.; Volke-Sepulveda, T.; Gaime-Perraud, I.; Saucedo-Castañeda, G.; Favela-Torres, E. Effect of stirring on growth and cellulolytic enzymes production by Trichoderma harzianum in a novel bench-scale solid-state fermentation bioreactor. Bioresour. Technol. 2018, 265, 291–298. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, J.; Luo, L.; Wang, E.; Wang, R.; Liu, L.; Liu, J.; Yuan, H. Low-Cost Cellulase-Hemicellulase Mixture Secreted by Trichoderma harzianum EM0925 with Complete Saccharification Efficacy of Lignocellulose. Int. J. Mol. Sci. 2020, 21, 371. [Google Scholar] [CrossRef]
- Zhao, X.; Zheng, Z.; Cai, Y.; Zhao, Y.; Zhang, Y.; Gao, Y.; Cui, Z.; Wang, X. Accelerated biomethane production from lignocellulosic biomass: Pretreated by mixed enzymes secreted by Trichoderma viride and Aspergillus sp. Bioresour. Technol. 2020, 309, 123378. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Plowman, J.E.; Tian, B.; Clerens, S.; On, S.L.W. The influence of growth conditions on MALDI-TOF MS spectra of winemaking yeast: Implications for industry applications. J. Microbiol. Methods 2021, 188, 106280. [Google Scholar] [CrossRef]
- Zhang, H.; Wu, J. Statistical optimization of sodium hydroxide pretreatment and enzymatic hydrolysis of corn stover powder for enhancing sugar production using response surface methodology. Biomass Convers. Biorefinery 2023, 13, 7111–7125. [Google Scholar] [CrossRef]
- Wang, Y.; Qiao, H.; Tao, Y.; Ma, Z.; Zheng, Z.; Ouyang, J. Addressing two major limitations in high-solids enzymatic hydrolysis by an ordered polyethylene glycol pre-incubated strategy: Rheological properties and lignin adsorption for enzyme. Bioresour. Technol. 2023, 390, 129895. [Google Scholar] [CrossRef]
- Goshadrou, A.; Lefsrud, M. Synergistic surfactant-assisted [EMIM]OAc pretreatment of lignocellulosic waste for enhanced cellulose accessibility to cellulase. Carbohydr. Polym. 2017, 166, 104–113. [Google Scholar] [CrossRef]
- Valles, A.; Álvarez-Hornos, F.J.; Martínez-Soria, V.; Marzal, P.; Gabaldón, C. Comparison of simultaneous saccharification and fermentation and separate hydrolysis and fermentation processes for butanol production from rice straw. Fuel 2020, 282, 118831. [Google Scholar] [CrossRef]
- Mateos, I.; Ranilla, M.J.; Ramos, M.; Saro, C.; Carro, M.D. Influence of rumen contents’ processing method on microbial populations in the fluid and subsequent in vitro fermentation of substrates of variable composition. Anim. Feed Sci. Technol. 2016, 220, 109–120. [Google Scholar] [CrossRef]
- Krishnamoorthy, U.; Soller, H.; Steingass, H.; Menke, K.H. Energy and protein evaluation of tropical feedstuffs for whole tract and ruminal digestion by chemical analyses and rumen inoculum studies in vitro. Anim. Feed Sci. Technol. 1995, 52, 177–188. [Google Scholar] [CrossRef]
- Santamaría-Fernández, M.; Molinuevo-Salces, B.; Kiel, P.; Steenfeldt, S.; Uellendahl, H.; Lübeck, M. Lactic acid fermentation for refining proteins from green crops and obtaining a high quality feed product for monogastric animals. J. Clean. Prod. 2017, 162, 875–881. [Google Scholar] [CrossRef]
- Oesper, R.E. Kjeldahl and the determination of nitrogen. J. Chem. Educ. 1934, 11, 457. [Google Scholar] [CrossRef]
- Wang, G.; Tan, L.; Sun, Z.; Gou, Z.; Tang, Y.; Kida, K. Production of bioethanol from rice straw by simultaneous saccharification and fermentation of whole pretreated slurry using Saccharomyces cerevisiae KF-7. Environ. Prog. Sustain. Energy 2015, 34, 582–588. [Google Scholar] [CrossRef]
- Smith, V.R. A phenol-hypochlorite manual determination of ammonium-nitrogen in Kjeldahl digests of plant tissue. Commun. Soil Sci. Plant Anal. 1980, 11, 709–722. [Google Scholar] [CrossRef]
- He, Y.; Wang, S.; Shen, C.; Wang, Z.; Liu, Y.; Meng, X.; Li, X.; Zhao, X.; Chen, J.; Xu, J.; et al. Biochar accelerates methane production efficiency from Baijiu wastewater: Some viewpoints considering direct interspecies electron transfer. Chem. Eng. J. 2024, 497, 154527. [Google Scholar] [CrossRef]
- Wu, J.; Ren, L.; Zhao, N.; Wu, T.; Liu, R.; Sui, W.; Zhang, M. Solid-state fermentation by Rhizopus oryzae improves flavor of wheat bran for application in food. J. Cereal Sci. 2022, 107, 103536. [Google Scholar] [CrossRef]
- Miller, G.L. Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Xie, C.; Yuan, R.; Su, L.; Li, D.; Zhang, C.; Yin, Y.; Wang, P.; Yang, R. Improving nutritional and sensory properties of rice bran by germination and solid-state fermentation with fungi. Food Biosci. 2024, 59, 103992. [Google Scholar] [CrossRef]
- Klop, G.; van Laar-Van Schuppen, S.; Pellikaan, W.F.; Hendriks, W.H.; Bannink, A.; Dijkstra, J. Changes in in vitro gas and methane production from rumen fluid from dairy cows during adaptation to feed additives in vivo. Animal 2017, 11, 591–599. [Google Scholar] [CrossRef]
- Wang, W.; Wang, C.; Zahoor; Chen, X.; Yu, Q.; Wang, Z.; Zhuang, X.; Yuan, Z. Effect of a Nonionic Surfactant on Enzymatic Hydrolysis of Lignocellulose Based on Lignocellulosic Features and Enzyme Adsorption. Acs Omega 2020, 5, 15812–15820. [Google Scholar] [CrossRef]
- Deswal, D.; Khasa, Y.P.; Kuhad, R.C. Optimization of cellulase production by a brown rot fungus Fomitopsis sp. RCK2010 under solid state fermentation. Bioresour. Technol. 2011, 102, 6065–6072. [Google Scholar] [CrossRef]
- Ma, X.; Li, S.; Tong, X.; Liu, K. An overview on the current status and future prospects in Aspergillus cellulase production. Environ. Res. 2024, 244, 117866. [Google Scholar] [CrossRef]
- Liang, J.; Zheng, W.; Zhang, H.; Zhang, P.; Cai, Y.; Wang, Q.; Zhou, Z.; Ding, Y. Transformation of bacterial community structure in rumen liquid anaerobic digestion of rice straw. Environ. Pollut. 2021, 269, 116130. [Google Scholar] [CrossRef] [PubMed]
- Aderibigbe, A.O.; Cheeke, P.R. Comparison of in vitro digestion of feed ingredients by rabbit cecal and bovine rumen fluids. Anim. Feed Sci. Technol. 1993, 41, 329–339. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, M.; Wang, R.; Ma, Z.; Long, D.; Mao, H.; Wen, J.; Bernard, L.A.; Beauchemin, K.A.; Tan, Z. Urea plus nitrate pretreatment of rice and wheat straws enhances degradation and reduces methane production in in vitro ruminal culture. J. Sci. Food Agric. 2018, 98, 5205–5211. [Google Scholar] [CrossRef]
- Dai, X.; Tian, Y.; Li, J.; Su, X.; Wang, X.; Zhao, S.; Liu, L.; Luo, Y.; Liu, D.; Zheng, H.; et al. Metatranscriptomic Analyses of Plant Cell Wall Polysaccharide Degradation by Microorganisms in the Cow Rumen. Appl. Environ. Microbiol. 2015, 81, 1375–1386. [Google Scholar] [CrossRef]
- Saminathan, M.; Wan Mohamed, W.N.; Md Noh, A.; Ibrahim, N.A.; Fuat, M.A.; Kumari Ramiah, S.; Jusoh, S.; Mat Dian, N.L.H. Effects of urea-treated oil palm frond on nutrient composition and in vitro rumen fermentation using goat rumen fluid. J. Anim. Physiol. Anim. Nutr. 2022, 106, 1228–1237. [Google Scholar] [CrossRef]
- Schlageter-Tello, A.; Fahey, G.C.; Freel, T.; Koutsos, L.; Miller, P.S.; Weiss, W.P. ASAS-NANP symposium: Ruminant/nonruminant feed composition: Challenges and opportunities associated with creating large feed composition tables. J. Anim. Sci. 2020, 98, skaa240. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Cellulose | Hemicellulose | Lignin | Crude Protein | True Protein |
|---|---|---|---|---|---|
| DGs | 31% | 7.5% | 24% | 14% | 10% |
| WB | 29% | 22% | 2.9% | 15% | 12% |
| CS | 25% | 13% | 15% | 8.5% | 6.2% |
| RH | 30% | 7.1% | 26% | 10% | 7.1% |
| Culture Medium (200 mL) | Glucose (g) | Sucrose (g) | Agar (g) | Potato Extract (mL) | Potato Extract Powder (g) |
|---|---|---|---|---|---|
| PDA | 4 | 0 | 4 | 0 | 1.2 |
| PDA0 | 0 | 0 | 4 | 200 | 0 |
| PDA5 | 1 | 0 | 4 | 200 | 0 |
| PDA10 | 2 | 0 | 4 | 200 | 0 |
| PDA15 | 3 | 0 | 4 | 200 | 0 |
| PDA20 | 4 | 0 | 4 | 200 | 0 |
| PDA25 | 5 | 0 | 4 | 200 | 0 |
| PSA0 | 4 | 0 | 4 | 200 | 0 |
| PSA5 | 3 | 1 | 4 | 200 | 0 |
| PSA10 | 2 | 2 | 4 | 200 | 0 |
| PSA15 | 1 | 3 | 4 | 200 | 0 |
| PSA20 | 0 | 4 | 4 | 200 | 0 |
| PSA25 | 0 | 5 | 4 | 200 | 0 |
| Sensor | Taste Information | |
|---|---|---|
| Initial Taste | Aftertaste | |
| CS00 | Bitterness | Sourness and Bitterness (Aftertaste-B) |
| AE1 | Astringency | Astringency (Aftertaste-A) |
| CA0 | Sourness | |
| CT0 | Saltness | |
| AAE | Umami | Richness |
| Sensor Number | Sensor Code | Sensitive Substances |
|---|---|---|
| 1 | W1C | Aromatic Compounds |
| 2 | W5S | Nitrogen Oxides |
| 3 | W3C | Ammonia and Aromatic Molecules |
| 4 | W6S | Hydrides |
| 5 | W5C | Olefins, Aromatic Compounds, and Polar Molecules |
| 6 | W1S | Alkanes |
| 7 | W1W | Sulfides |
| 8 | W2S | Alcohols and Some Aromatic Compounds |
| 9 | W2W | Aromatic Compounds and Organic Sulfur Compounds |
| 10 | W3S | Alkanes and Aliphatic Compounds |
| RUN | X1: Moisture Content (%) | X2: Time (d) | X3: Tween-80 (mL) | FPase (U/gds) |
|---|---|---|---|---|
| 1 | 50 | 4 | 0.1 | 1.11 |
| 2 | 60 | 4 | 0.15 | 0.72 |
| 3 | 40 | 4 | 015 | 0.62 |
| 4 | 50 | 4 | 0.1 | 1.14 |
| 5 | 40 | 5 | 0.1 | 0.72 |
| 6 | 60 | 3 | 0.1 | 1.08 |
| 7 | 50 | 4 | 0.1 | 1.18 |
| 8 | 50 | 3 | 0.15 | 1.36 |
| 9 | 40 | 3 | 0.1 | 0.62 |
| 10 | 60 | 5 | 0.1 | 0.20 |
| 11 | 50 | 5 | 0.05 | 0.98 |
| 12 | 50 | 4 | 0.1 | 1.03 |
| 13 | 60 | 4 | 0.05 | 0.29 |
| 14 | 50 | 5 | 0.15 | 1.19 |
| 15 | 50 | 4 | 0.1 | 1.17 |
| 16 | 40 | 4 | 0.05 | 0.69 |
| 17 | 50 | 3 | 0.05 | 1.18 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bai, X.; Wang, J.; Wang, X.; Li, S.; Yang, Y.; Sun, R.; Wang, S.; Zhao, X.; Wang, Z.; Cai, Y.; et al. Pretreatment of Luzhou Distiller’s Grains with Crude Enzyme from Trichoderma harzianum for Feed Protein Production. Fermentation 2025, 11, 294. https://doi.org/10.3390/fermentation11050294
Bai X, Wang J, Wang X, Li S, Yang Y, Sun R, Wang S, Zhao X, Wang Z, Cai Y, et al. Pretreatment of Luzhou Distiller’s Grains with Crude Enzyme from Trichoderma harzianum for Feed Protein Production. Fermentation. 2025; 11(5):294. https://doi.org/10.3390/fermentation11050294
Chicago/Turabian StyleBai, Xueke, Jiaxin Wang, Xi Wang, Shuai Li, Yanni Yang, Ruoya Sun, Shilei Wang, Xiaoling Zhao, Zhi Wang, Yafan Cai, and et al. 2025. "Pretreatment of Luzhou Distiller’s Grains with Crude Enzyme from Trichoderma harzianum for Feed Protein Production" Fermentation 11, no. 5: 294. https://doi.org/10.3390/fermentation11050294
APA StyleBai, X., Wang, J., Wang, X., Li, S., Yang, Y., Sun, R., Wang, S., Zhao, X., Wang, Z., Cai, Y., Xu, J., & Ying, H. (2025). Pretreatment of Luzhou Distiller’s Grains with Crude Enzyme from Trichoderma harzianum for Feed Protein Production. Fermentation, 11(5), 294. https://doi.org/10.3390/fermentation11050294