
Transcriptional and Physiological Responses of Saccharomyces cerevisiae CZ to Octanoic Acid Stress

Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents and Yeast Strains
2.2. Determination of Octanoic Acid Stress Concentration
- (1)
- OD600 measurement: cells were vortexed for 30 sec, and absorbance was measured against blank YPD.
- (2)
- Spot plating: serial 10-fold dilutions (100 to 10−4) in sterile water, plated (2 μL/drop) on YPD agar.
- (3)
- Viability assessment: methylene blue staining as per Section 2.4.
2.3. Determination of Spotting Count and Yeast Suspension Concentration
2.4. Methylene Blue Staining and Cell Counting
2.5. Growth Curve Determination
2.6. Determination of Trace Malondialdehyde (MDA) and H+/K+-ATPase Activity
2.7. Transcriptional Analysis of Cell Gene Expression Profiles
2.8. Data Analysis
3. Results and Discussion
3.1. Physiological Characteristics Analysis of S. cerevisiae CZ
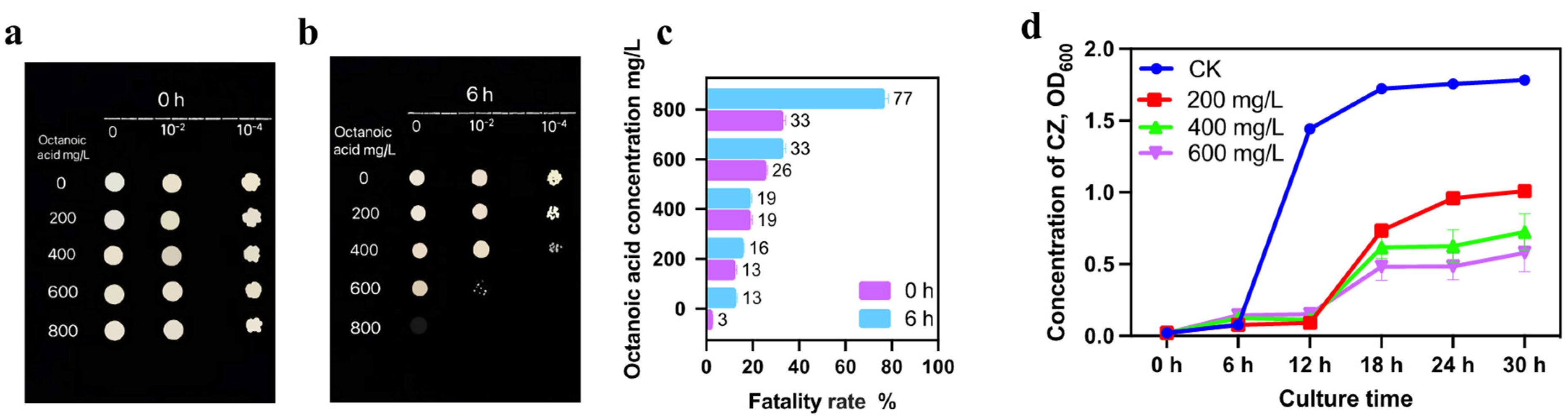
3.1.1. Effects of Octanoic Acid Stress on Growth and Lethality of S. cerevisiae CZ
3.1.2. Effects of Octanoic Acid Stress on H+/K+-ATPase Activity and Malondialdehyde Content in S. cerevisiae CZ
3.2. Analysis of Gene Expression Profiles in S. cerevisiae
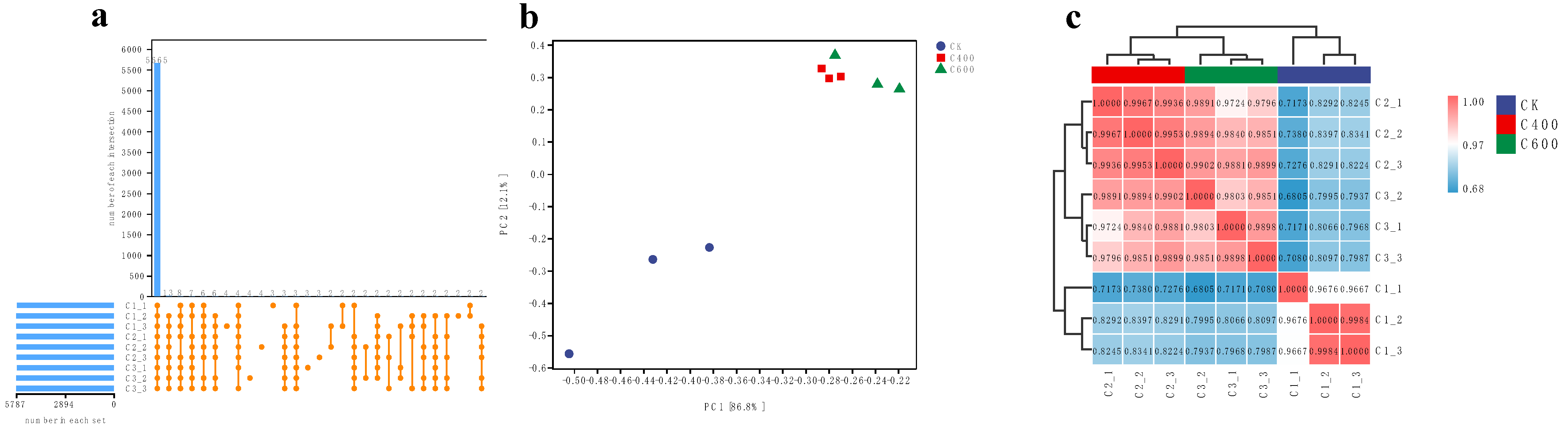
3.2.1. Quality Control Analysis of Transcriptomic Data
3.2.2. Gene Expression Analysis
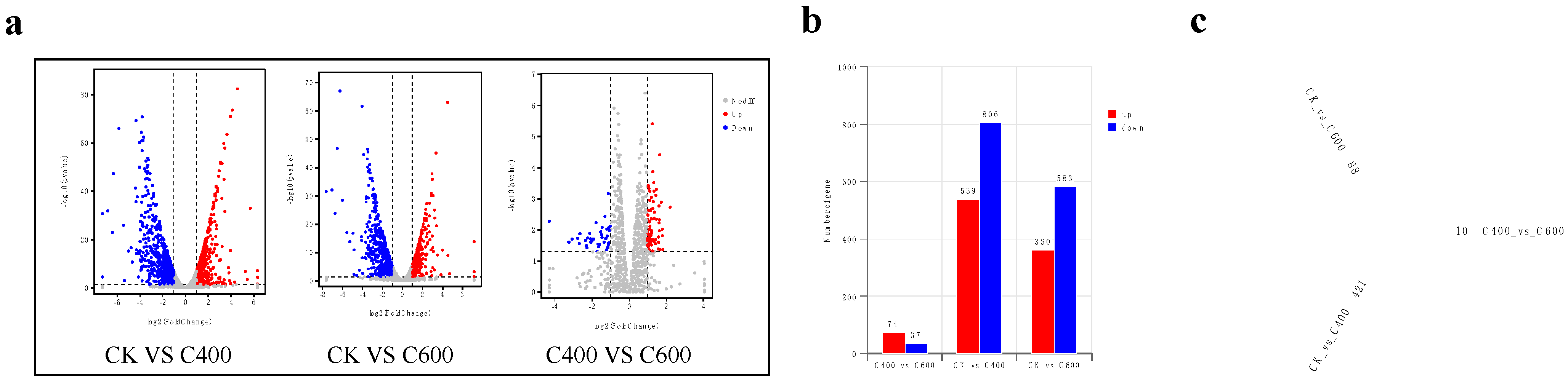
3.2.3. Statistical Analysis of Differentially Expressed Genes
3.2.4. Functional Enrichment Analysis of Differentially Expressed Genes
3.2.5. Potential Mechanisms of S. cerevisiae in Response to Octanoic Acid Stress
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vanderwaeren, L.; Dok, R.; Voordeckers, K.; Nuyts, S.; Verstrepen, K.J. Saccharomyces cerevisiae as a model system for eukaryotic cell biology, from cell cycle control to DNA damage response. Int. J. Mol. Sci. 2022, 23, 11665. [Google Scholar] [CrossRef] [PubMed]
- Jouhten, P.; Ponomarova, O.; Gonzalez, R.; Patil, K.R. Saccharomyces cerevisiae metabolism in ecological context. FEMS Yeast Res. 2016, 16, fow080. [Google Scholar] [PubMed]
- Nielsen, J. Yeast systems biology: Model organism and cell factory. Biotechnol. J. 2019, 14, e1800421. [Google Scholar]
- Berłowska, J.; Pielech-Przybylska, K.; Balcerek, M.; Dziekońska-Kubczak, U.; Patelski, P.; Dzi-ugan, P.; Kręgiel, D. Simultaneous Saccharification and fermentation of sugar beet pulp for efficient bioethanol production. Biomed. Res. Int. 2016, 2016, 3154929. [Google Scholar]
- Tao, Z.; Yuan, H.; Liu, M.; Liu, Q.; Zhang, S.; Liu, H.; Jiang, Y.; Huang, D.; Wang, T. Yeast extract: Characteristics, production, applications and future perspectives. J. Microbiol. Biotechnol. 2023, 33, 151–166. [Google Scholar]
- Wang, Z.; Zhang, R.; Yang, Q.; Zhang, J.; Zhao, Y.; Zheng, Y.; Yang, J. Recent advances in the biosynthesis of isoprenoids in engineered Saccharomyces cerevisiae. Adv. Appl. Microbiol. 2021, 114, 1–35. [Google Scholar]
- Kang, N.K.; Lee, J.W.; Ort, D.R.; Jin, Y.S. L-malic acid production from xylose by engineered Saccharomyces cerevisiae. Biotechnol. J. 2022, 17, e2000431. [Google Scholar]
- Swamy, K.B.S.; Zhou, N. Experimental evolution: Its principles and applications in developing stress-tolerant yeasts. Appl. Microbiol. Biotechnol. 2019, 103, 2067–2077. [Google Scholar] [PubMed]
- Cabral, M.G.; Viegas, C.A.; Sá-Correia, I. Mechanisms underlying the acquisition of resistance to octanoic-acid-induced-death following exposure of Saccharomyces cerevisiae to mild stress imposed by octanoic acid or ethanol. Arch. Microbiol. 2001, 175, 301–307. [Google Scholar]
- Viegas, C.A.; Supply, P.; Capieaux, E.; Van Dyck, L.; Goffeau, A.; Sá-Correia, I. Regulation of the expression of the H(+)-ATPase genes PMA1 and PMA2 during growth and effects of octanoic acid in Saccharomyces cerevisiae. Biochim. Biophys. Acta 1994, 1217, 74–80. [Google Scholar]
- Legras, J.L.; Erny, C.; Le Jeune, C.; Lollier, M.; Adolphe, Y.; Demuyter, C.; Delobel, P.; Blondin, B.; Karst, F. Activation of two different resistance mechanisms in Saccharomyces cerevisiae upon exposure to octanoic and decanoic acids. Appl. Environ. Microbiol. 2010, 76, 7526–7535. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Chernyshov, A.; Najdi, T.; Fu, Y.; Dickerson, J.; Sandmeyer, S.; Jarboe, L. Membrane stress caused by octanoic acid in Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2013, 97, 3239–3251. [Google Scholar] [PubMed]
- Rajvanshi, P.K.; Arya, M.; Rajasekharan, R. The stress-regulatory transcription factors Msn2 and Msn4 regulate fatty acid oxidation in budding yeast. J. Biol. Chem. 2017, 292, 18628–18643. [Google Scholar] [CrossRef]
- Zhang, X.; Nijland, J.G.; Driessen, A.J.M. Combined roles of exporters in acetic acid tolerance in Saccharomyces cerevisiae. Biotechnol. Biofuels Bioprod. 2022, 15, 67. [Google Scholar] [CrossRef]
- Guirimand, G.; Kulagina, N.; Papon, N.; Hasunuma, T.; Courdavault, V. Innovative tools and strategies for optimizing yeast cell factories. Trends Biotechnol. 2021, 39, 488–504. [Google Scholar] [CrossRef]
- Zhang, Z.; Lan, Q.; Yu, Y.; Zhou, J.; Lu, H. Comparative metabolome and transcriptome analyses of the properties of Kluyveromyces marxianus and Saccharomyces yeasts in apple cider fermentation. Food Chem. 2022, 4, 100095. [Google Scholar] [CrossRef]
- Liu, H.; Yuan, W.; Zhou, P.; Liang, G.; Gao, C.; Guo, L.; Hu, G.; Song, W.; Wu, J.; Chen, X.; et al. Engineering membrane asymmetry to increase medium-chain fatty acid tolerance in Saccharomyces cerevisiae. Biotechnol. Bioeng. 2022, 119, 277–286. [Google Scholar] [CrossRef]
- Besada-Lombana, P.B.; Fernandez-Moya, R.; Fenster, J.; Da Silva, N.A. Engineering Saccharomyces cerevisiae fatty acid composition for increased tolerance to octanoic acid. Biotechnol. Bioeng. 2017, 114, 1531–1538. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Yue, L.; Wang, Q.; Wu, J.; Zhong, X.; Liu, X.; Huang, M. Identification and brewing potential of a Saccharomyces cerevisiae strain from Rosa roxbunghii. Food Ferment. Indu. 2022, 48, 70–75. (In Chinese) [Google Scholar]
- Yu, Z.-H.; Huang, G.-D.; Huang, X.Y.; Pu, J.H.; Wu, J.S.; Yue, L.R.; Hardie, W.J.; Liu, X.Z.; Huang, M.Z. A Comparative Study of Yeasts for Rosa roxburghii Wine Fermentation. Fermentation 2022, 8, 311. [Google Scholar] [CrossRef]
- Li, Y.; Long, H.; Jiang, G.; Gong, X.; Yu, Z.; Huang, M.; Guan, T.; Guan, Y.; Liu, X. Analysis of the ethanol stress response mechanism in Wickerhamomyces anomalus based on transcriptomics and metabolomics approaches. BMC Microbiol. 2022, 22, 275. [Google Scholar]
- Chen, S.; Li, Y.; Wu, E.; Li, Q.; Xiang, L.; Qi, J. Arctigenin from Fructus arctii exhibits antiaging effects via autophagy induction, antioxidative stress, and increase in telomerase activity in yeast. Antioxidants 2024, 13, 684. [Google Scholar] [CrossRef] [PubMed]
- Grigore, D.; Meade, J.C. Functional complementation of the yeast P-type H-ATPase, PMA1, by the Pneumocystis carinii P-type H-ATPase, PCA1. J. Eukaryot. Microbiol. 2006, 53, 157–164. [Google Scholar] [PubMed]
- Viegas, C.A.; Rosa, M.F.; Sá-Correia, I.; Novais, J.M. Inhibition of yeast growth by octanoic and decanoic acids produced during ethanolic fermentation. Appl. Environ. Microbiol. 1989, 55, 21–28. [Google Scholar] [CrossRef]
- Borrull, A.; López-Martínez, G.; Poblet, M.; Cordero-Otero, R.; Rozès, N. New insights into the toxicity mechanism of octanoic and decanoic acids on Saccharomyces cerevisiae. Yeast 2015, 32, 451–460. [Google Scholar]
- Viegas, C.A.; Almeida, P.F.; Cavaco, M.; Sá-Correia, I. The H(+)-ATPase in the plasma membrane of Saccharomyces cerevisiae is activated during growth latency in octanoic acid-supplemented medium accompanying the decrease in intracellular pH and cell viability. Appl. Environ. Microbiol. 1998, 64, 779–783. [Google Scholar]
- Mohideen, K.; Chandrasekar, K.; Ramsridhar, S.; Rajkumar, C.; Ghosh, S.; Dhungel, S. Assessment of oxidative stress by the estimation of lipid peroxidation marker malondialdehyde (MDA) in patients with chronic periodontitis: A systematic review and meta-analysis. Int. J. Dent. 2023, 2023, 6014706. [Google Scholar]
- Gerçek, E.; Zengin, H.; Erdem Erişir, F.; Yılmaz, Ö. Biochemical changes and antioxidant capacity of naringin and naringenin against malathion toxicity in Saccharomyces cerevisiae. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2021, 241, 108969. [Google Scholar]
- James, T.C.; Campbell, S.; Donnelly, D.; Bond, U. Transcription profile of brewery yeast under fermentation conditions. J. Appl. Microbiol. 2003, 94, 432–448. [Google Scholar]
- Li, P.; Fu, X.; Chen, M.; Zhang, L.; Li, S. Proteomic profiling and integrated analysis with transcriptomic data bring new insights in the stress responses of Kluyveromyces marxianus after an arrest during high-temperature ethanol fermentation. Biotechnol. Biofuels 2019, 12, 49. [Google Scholar]
- Baumann, L.; Doughty, T.; Siewers, V.; Nielsen, J.; Boles, E.; Oreb, M. Transcriptomic response of Saccharomyces cerevisiae to octanoic acid production. FEMS Yeast Res. 2021, 21, foab011. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
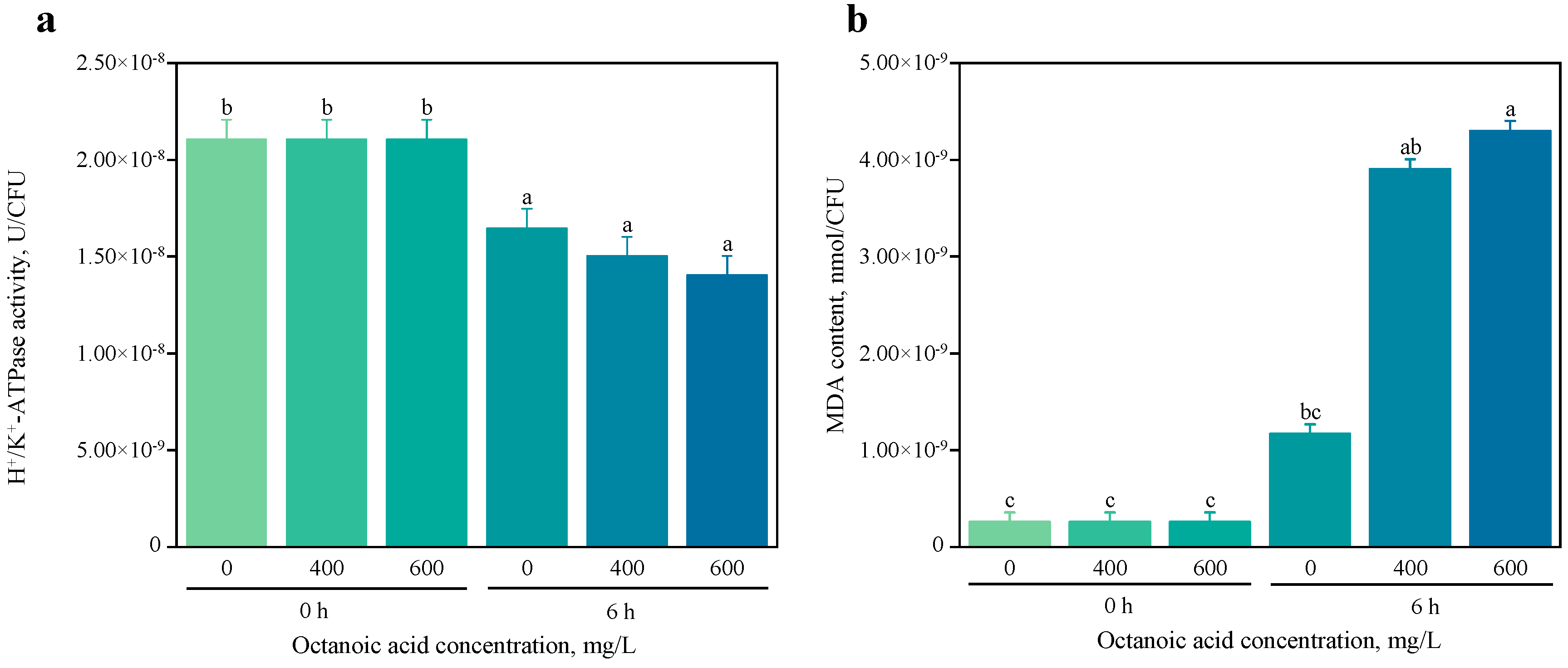


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Raw Reads | Raw Bases | Clean Reads | Useful Reads % |
|---|---|---|---|---|
| 0 mg/L | 37,637,308 | 5,683,233,508 | 35,536,034 | 94.41 ± 0.03 |
| 400 mg/L | 42,983,217 | 6,490,465,817 | 40,496,293 | 94.21 ± 0.08 |
| 600 mg/L | 38,825,746 | 5,862,687,747 | 36,624,445 | 94.33 ± 0.08 |
| Treatment | Useful bases % | Clean bases | Q20% | Q30% |
| 0 mg/L | 94.41 ± 0.03 | 5,365,941,134 | 98.17 ± 0.26 | 94.41 ± 0.68 |
| 400 mg/L | 94.21 ± 0.08 | 6,114,940,293 | 98.04 ± 0.21 | 94.09 ± 0.60 |
| 600 mg/L | 94.33 ± 0.08 | 5,530,291,245 | 98.04 ± 0.27 | 94.02 ± 0.72 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, Z.-H.; Shi, M.-Z.; Dong, W.-X.; Liu, X.-Z.; Tang, W.-Y.; Huang, M.-Z. Transcriptional and Physiological Responses of Saccharomyces cerevisiae CZ to Octanoic Acid Stress. Fermentation 2025, 11, 180. https://doi.org/10.3390/fermentation11040180
Yu Z-H, Shi M-Z, Dong W-X, Liu X-Z, Tang W-Y, Huang M-Z. Transcriptional and Physiological Responses of Saccharomyces cerevisiae CZ to Octanoic Acid Stress. Fermentation. 2025; 11(4):180. https://doi.org/10.3390/fermentation11040180
Chicago/Turabian StyleYu, Zhi-Hai, Ming-Zhi Shi, Wen-Xuan Dong, Xiao-Zhu Liu, Wei-Yuan Tang, and Ming-Zheng Huang. 2025. "Transcriptional and Physiological Responses of Saccharomyces cerevisiae CZ to Octanoic Acid Stress" Fermentation 11, no. 4: 180. https://doi.org/10.3390/fermentation11040180
APA StyleYu, Z.-H., Shi, M.-Z., Dong, W.-X., Liu, X.-Z., Tang, W.-Y., & Huang, M.-Z. (2025). Transcriptional and Physiological Responses of Saccharomyces cerevisiae CZ to Octanoic Acid Stress. Fermentation, 11(4), 180. https://doi.org/10.3390/fermentation11040180