
Optimizing the Production of High-Quality Silage from Jingkenuo 2000 Fresh Waxy Maize: The Synergistic Effects of Microbial Fertilizer and Fermentation Agents

 , , and
, , and
Abstract
1. Introduction
2. Materialsand Methods
2.1. Planting Experimental Design
- (1)
- Conventional fertilization treatment (CK)
- (2)
- Conventional fertilization + drip irrigation bacterial solution treatment (JJ)
- (3)
- Conventional fertilization + microbial organic fertilizer treatment (YJ)
2.2. Tissue Sample Collection
2.3. Fermentation Sample Collection
2.4. Chemical Composition
2.5. Analysis of Bacterial Communities in Fermented Feed
2.6. Statistical Analyses
3. Results
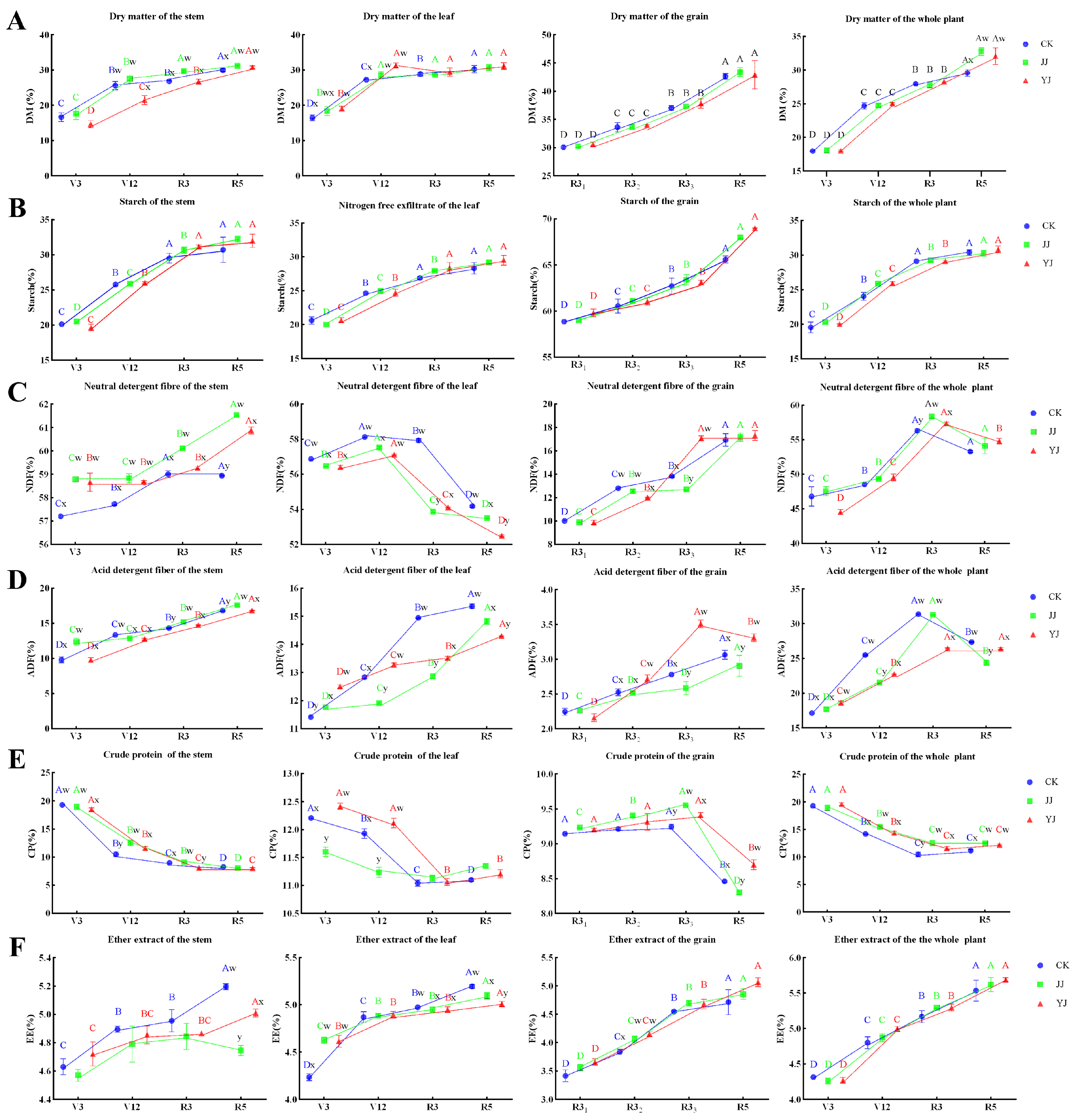
3.1. Nutrient Accumulation in the Tissues and Whole Plant
3.2. Nutrient Accumulation in the Tissues and Whole Plant
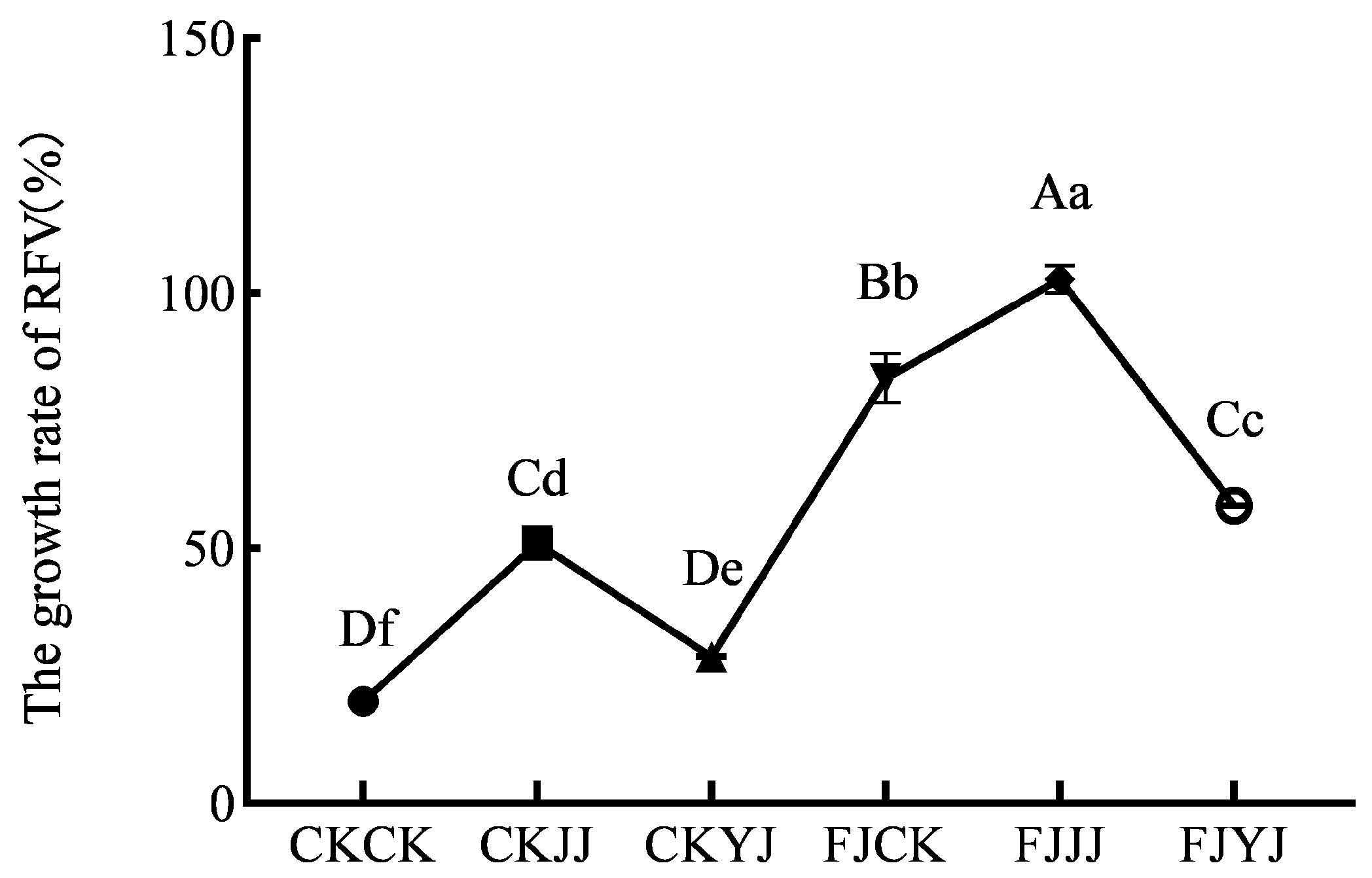
3.3. Fermentation Quality of the Silage
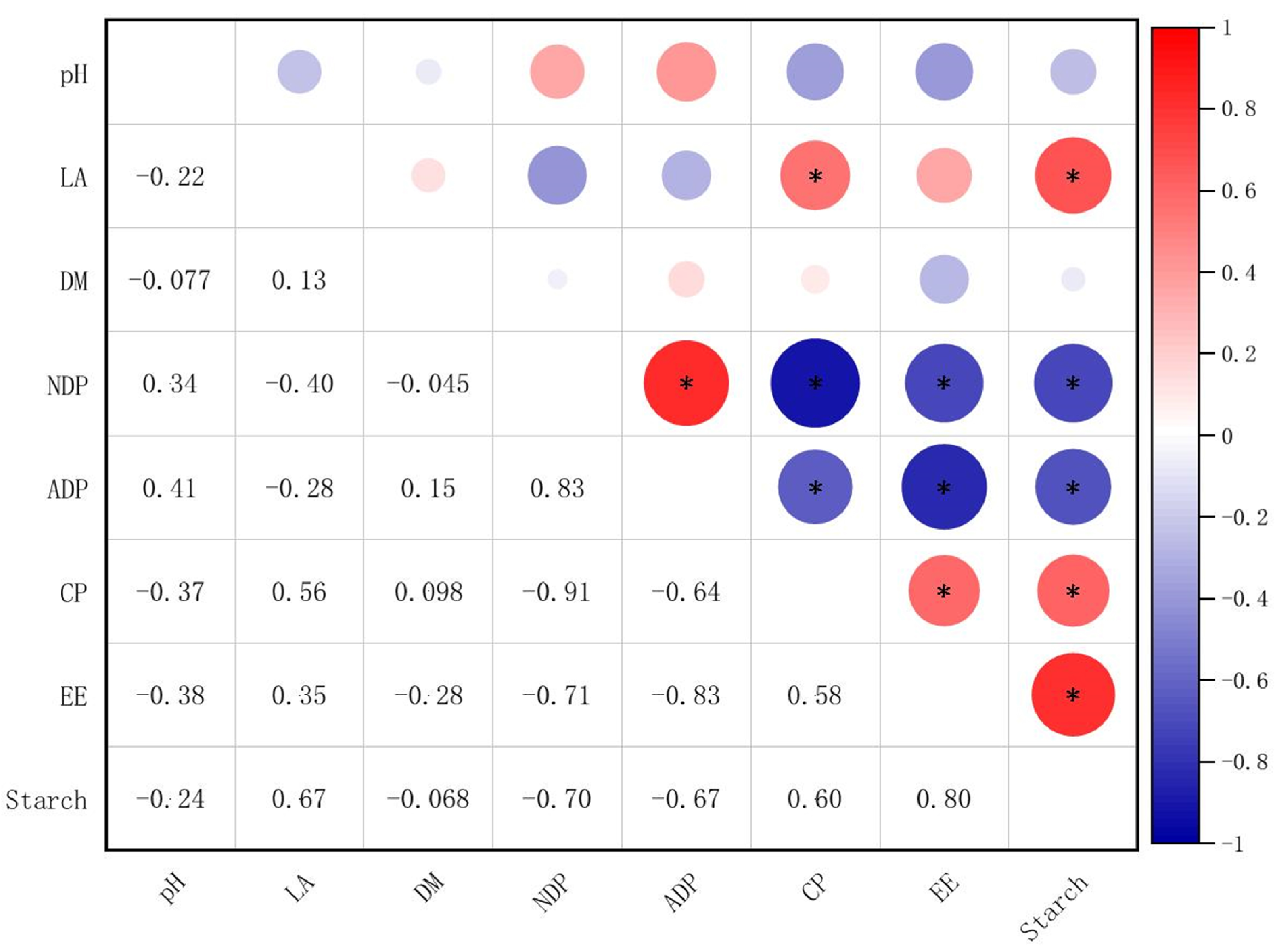
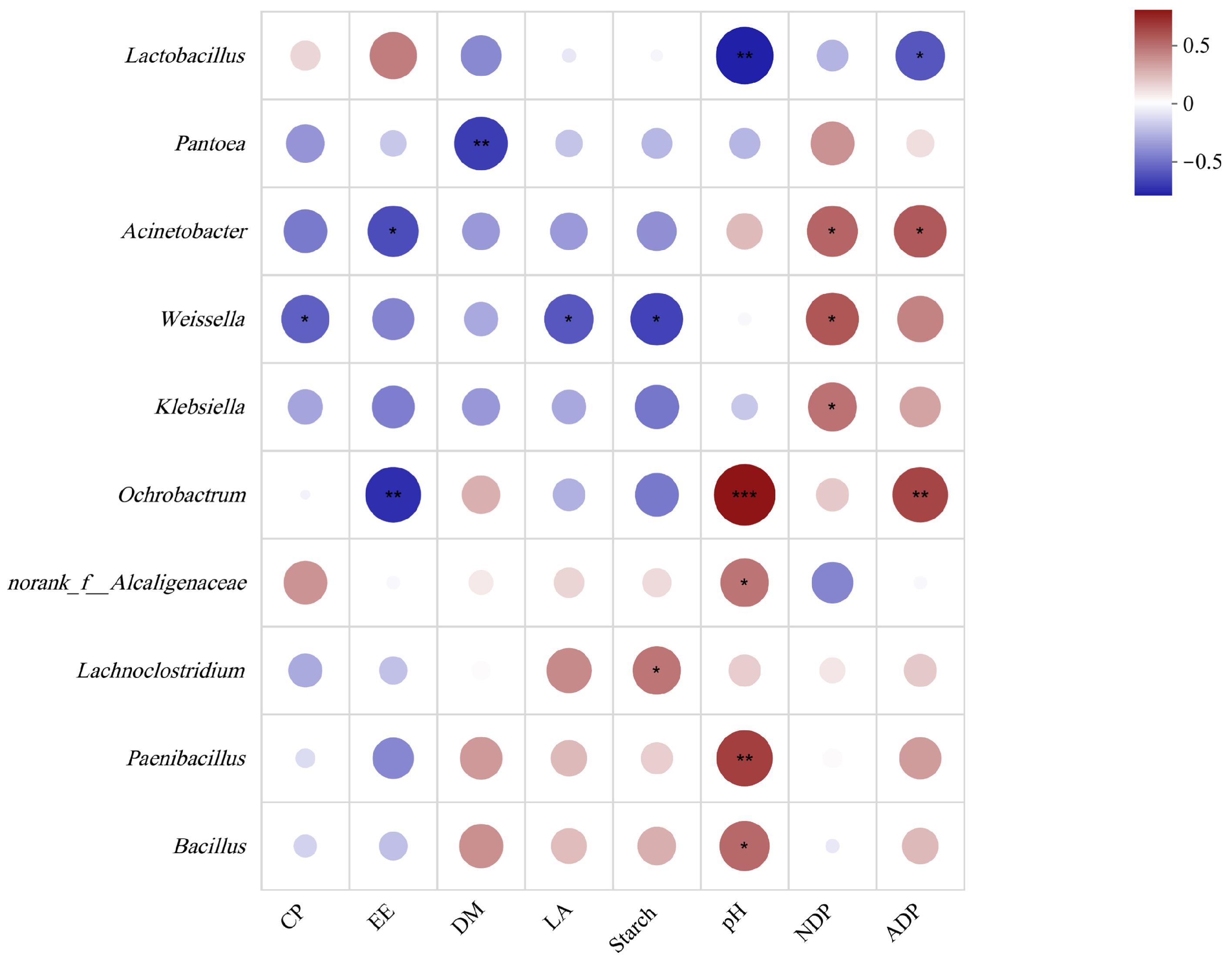
3.4. Correlation Analysis Between Silage Nutritional Quality and Fermentation Quality
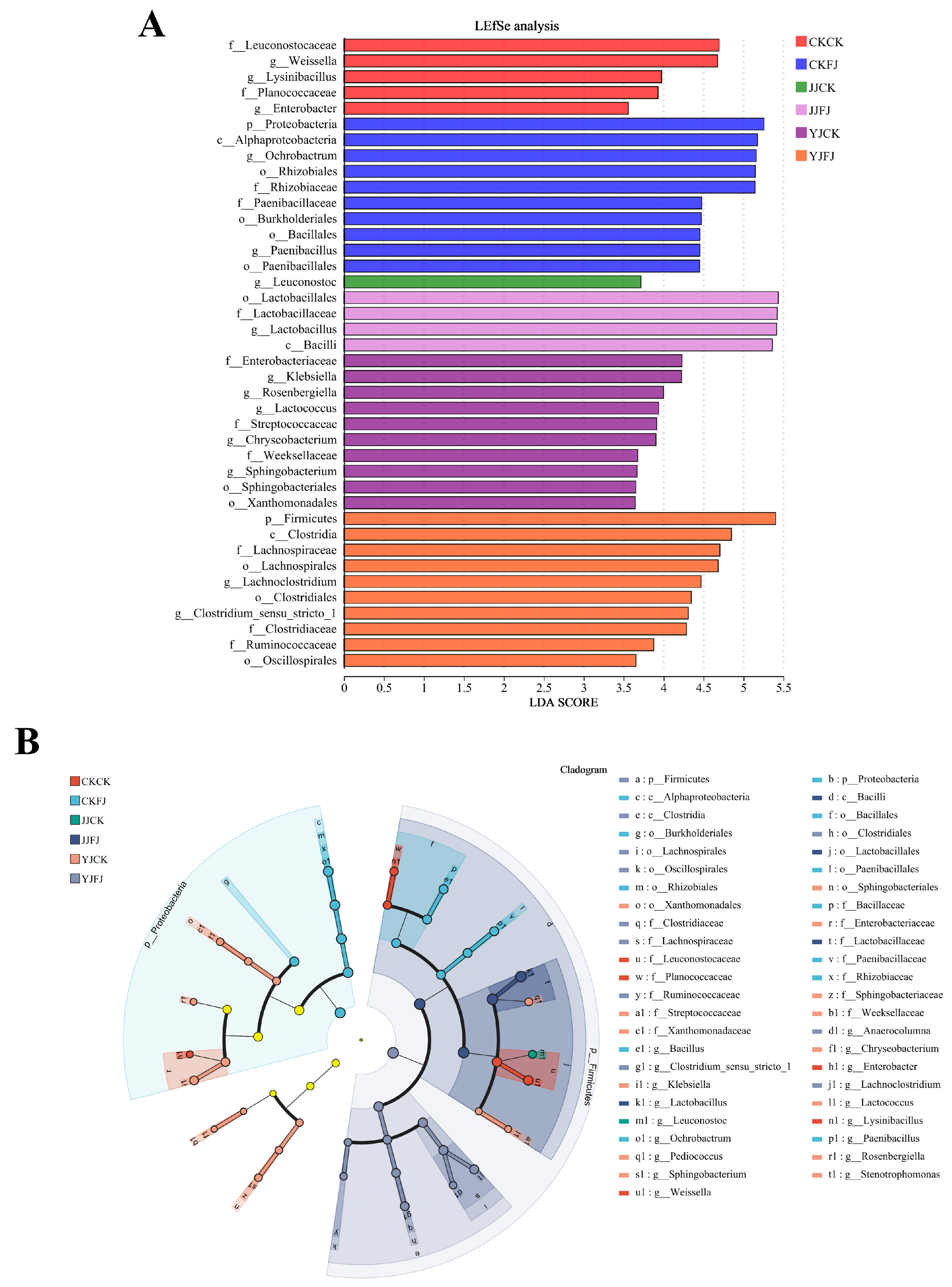
3.5. Bacterial Diversity and Community Composition of the Whole-Plant Corn Silage
3.6. Correlation Between Fermentation Parameters and Bacterial Communities
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ni, K.; Zhao, J.; Zhu, B.; Su, R.; Pan, Y.; Ma, J.; Zhou, G.; Tao, Y.; Liu, X.; Zhong, J. Assessing the fermentation quality and microbial community of the mixed silage of forage soybean with crop corn or sorghum. Bioresour. Technol. 2018, 265, 563–567. [Google Scholar] [CrossRef] [PubMed]
- Ni, K.; Wang, F.; Zhu, B.; Yang, J.; Zhou, G.; Pan, Y.I.; Tao, Y.; Zhong, J. Effects of lactic acid bacteria and molasses additives on the microbial community and fermentation quality of soybean silage. Bioresour. Technol. 2017, 238, 706–715. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Xu, H.; Sun, P.; Zhao, B.; Dong, S. Research Progress on Harmless Treatment of Dead Livestock and Poultry with Alkaline Hydrolysis in China. Anim. Husb. Feed. Sci. 2017, 9, 346–354. [Google Scholar]
- Guan, H.; Ran, Q.; Li, H.; Zhang, X. Succession of Microbial Communities of Corn Silage Inoculated with Heterofermentative Lactic Acid Bacteria from Ensiling to Aerobic Exposure. Fermentation 2021, 7, 258. [Google Scholar] [CrossRef]
- Lu, B.-S.; Xu, L.; Zhao, J.-R.; Shi, Y.-X.; Fan, Y.-L.; Xi, S.-L.; Zhang, C.-F.; Shi, Y.-M.; Zhang, X.-J. Breeding and Application of Jingkenuo2000 Series of Waxy Corn Varieties. J. Maize Sci. 2019, 27, 1. [Google Scholar] [CrossRef]
- Barlow, J.; Bernard, J.; Mullis, N. Production response to corn silage produced from normal, brown midrib, or waxy corn hybrids. J. Dairy Sci. 2012, 95, 4550–4555. [Google Scholar] [CrossRef]
- Gao, H.; Gadlage, M.J.; Lafitte, H.R.; Lenderts, B.; Yang, M.; Schroder, M.; Farrell, J.; Snopek, K.; Peterson, D.; Feigenbutz, L.; et al. Superior field performance of waxy corn engineered using CRISPR–Cas9. Nat. Biotechnol. 2020, 38, 579–581. [Google Scholar] [CrossRef]
- Yan, X.; Wu, Z.Z.; Zuo, Y.C.; Wang, H.L.; Wang, Q.F.; Li, Y.; Kou, J.; Du, Z.H. Silage Characteristics of Different Corn Plant Parts and Strategies for Improving Their Silage Quality. Acta Agrestia Sin. 2023, 31, 2275–2286. [Google Scholar] [CrossRef]
- Amanullah, A.; Khan, A. Phosphorus and Compost Management Influence Maize (Zea mays) Productivity Under Semiarid Condition with and without Phosphate Solubilizing Bacteria. Front. Plant Sci. 2015, 6, 1083. [Google Scholar] [CrossRef]
- Zafar-Ul-Hye, M.; Farooq, H.M.; Hussain, M. Bacteria in combination with fertilizers promote root and shoot growth of maize in saline-sodic soil. Braz. J. Microbiol. 2015, 46, 97–102. [Google Scholar] [CrossRef]
- Katsenios, N.; Andreou, V.; Sparangis, P.; Djordjevic, N.; Giannoglou, M.; Chanioti, S.; Kasimatis, C.N.; Kakabouki, I.; Leonidakis, D.; Danalatos, N.; et al. Assessment of plant growth promoting bacteria strains on growth, yield and quality of sweet corn. Sci. Rep. 2022, 12, 11598. [Google Scholar] [CrossRef] [PubMed]
- Meng, H.; Jiang, Y.; Wang, L.; Wang, S.; Zhang, Z.; Tong, X.; Wang, S. Effects of Different Soybean and Maize Mixed Proportions in a Strip Intercropping System on Silage Fermentation Quality. Fermentation 2022, 8, 696. [Google Scholar] [CrossRef]
- Okoye, C.O.; Wang, Y.; Gao, L.; Wu, Y.; Li, X.; Sun, J.; Jiang, J. The performance of lactic acid bacteria in silage production: A review of modern biotechnology for silage improvement. Microbiol. Res. 2022, 266, 127212. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Lee, K.D.; Choi, K.C. Role of lab in silage fermentation: Effect on nutritional quality and organic acid production—An overview. AIMS Agric. Food 2021, 6, 216–234. [Google Scholar] [CrossRef]
- Guo, X.; Xu, D.; Li, F.; Bai, J.; Su, R. Current approaches on the roles of lactic acid bacteria in crop silage. Microb. Biotechnol. 2023, 16, 67–87. [Google Scholar] [CrossRef]
- Chauhan, N.; Kumari, N.; Mani, V.; Pradhan, D.; Gowane, G.R.; Kumar, S.; Tyagi, N. Effects of Lactiplantibacillus plantarum, Limosilactobacillus fermentum, and Propionic Acid on the Fermentation Process Of Sugarcane Tops Silages Along with Variations in pH, Yeast and Mould Count After Aerobic Exposure. Waste Biomass Valorization 2024, 15, 2215–2230. [Google Scholar] [CrossRef]
- Momota Rani, D.; Brigitta, W.; Annette, L. Method development to reduce the fiber content of wheat bran and rice bran through anaerobic fermentation with rumen liquor for use in poultry feed. Asian-Australas. J. Anim. Sci. 2019, 32, 395–404. [Google Scholar] [CrossRef]
- Watanabe, M.; Kanaguri, Y.; Smith, R.L. Hydrothermal separation of lignin from bark of Japanese cedar. J. Supercrit. Fluids 2018, 133, 696–703. [Google Scholar] [CrossRef]
- Wang, Y. Study on the correlation of the different organs and the yield and quality in silage maize. Chin. Agric. Sci. Bull. 2015, 31, 22–27. [Google Scholar] [CrossRef]
- Cherdthong, A.; Khonkhaeng, B.; Seankamsorn, A.; Supapong, C.; Wanapat, M.; Gunun, N.; Gunun, P.; Chanjula, P.; Polyorach, S. Effects of feeding fresh cassava root with high-sulfur feed block on feed utilization, rumen fermentation, and blood metabolites in Thai native cattle. Trop. Anim. Health Prod. 2018, 50, 1365–1371. [Google Scholar] [CrossRef]
- Edson, C.; Takarwirwa, N.N.; Kuziwa, N.L.; Stella, N.; Maasdorp, B. Effect of mixed maize-legume silages on milk quality and quantity from lactating smallholder dairy cows. Trop. Anim. Health Prod. 2018, 50, 1255–1260. [Google Scholar] [CrossRef]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef] [PubMed]
- Tucak, M.; Ravlić, M.; Horvat, D.; Čupić, T. Improvement of Forage Nutritive Quality of Alfalfa and Red Clover through Plant Breeding. Agronomy 2021, 11, 2176. [Google Scholar] [CrossRef]
- Mohd, A.; Arvind, K.; Pourouchottamane, R.; Gupta, D.L.; Singh, M.K.; Rai, B. Effect of intercropping row ratios on yield and nutritive value of maize and cowpea fodder. Range Manag. Agrofor. 2022, 43, 292–298. [Google Scholar]
- Yan, Y.; Li, X.; Guan, H.; Huang, L.; Ma, X.; Peng, Y.; Li, Z.; Nie, G.; Zhou, J.; Yang, W.; et al. Microbial community and fermentation characteristic of Italian ryegrass silage prepared with corn stover and lactic acid bacteria. Bioresour. Technol. 2019, 279, 166–173. [Google Scholar] [CrossRef]
- Enes, P.; Panserat, S.; Kaushik, S.; Oliva-Teles, A. Effect of normal and waxy maize starch on growth, food utilization and hepatic glucose metabolism in European sea bass (Dicentrarchus labrax) juveniles. Comp. Biochem. Physiol. Part Mol. Integr. Physiol. 2006, 143, 89–96. [Google Scholar] [CrossRef]
- Hahnen, S.; Joeris, T.; Kreuzaler, F.; Peterhänsel, C. Quantification of photosynthetic gene expression in maize C(3) and C(4) tissues by real-time PCR. Photosynth. Res. 2003, 75, 183–192. [Google Scholar] [CrossRef]
- Kamran, M.; Ahmad, I.; Wang, H.; Wu, X.; Xu, J.; Liu, T.; Ding, R.; Han, Q. Mepiquat chloride application increases lodging resistance of maize by enhancing stem physical strength and lignin biosynthesis. Field Crop. Res. 2018, 224, 148–159. [Google Scholar] [CrossRef]
- Rehemujiang, H.; Yusuf, H.A.; Ma, T.; Diao, Q.; Kong, L.; Kang, L.; Tu, Y. Evaluating Fermentation Quality, Aerobic Stability, and Rumen-Degradation (In Situ) Characteristics of Various Protein-Based Total Mixed Rations. Animals 2023, 13, 2730. [Google Scholar] [CrossRef]
- Keshri, J.; Chen, Y.; Pinto, R.; Kroupitski, Y.; Weinberg, Z.G.; Sela Saldinger, S. Microbiome dynamics during ensiling of corn with and without Lactobacillus plantarum inoculant. Appl. Microbiol. Biotechnol. 2018, 102, 4025–4037. [Google Scholar] [CrossRef]
- Zhang, F.; Wang, X.; Lu, W.; Li, F.; Ma, C. Improved Quality of Corn Silage When Combining Cellulose-Decomposing Bacteria and Lactobacillus buchneri during Silage Fermentation. Biomed Res. Int. 2019, 2019, 4361358. [Google Scholar] [CrossRef]
- Nogoy, K.M.; Zhang, Y.; Lee, Y.H.; Li, X.Z.; Seong, H.A.; Choi, S.H. Nutrient composition and in vitro fermentability of corn grain and stover harvested at different periods in Goesan, a mountainous area. J. Anim. Sci. Technol. 2019, 61, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Moore, J.E.; Undersander, D. Relative forage quality: An alternative to relative feed value and quality index. In Proceedings of the 13th Annual Florida Ruminant Nutrition Symposium, Gainesville, FL, USA, 11–12 January 2002. [Google Scholar]
- Zhai, J.; Zhang, G.; Zhang, Y.; Xu, W.; Xie, R.; Ming, B.; Hou, P.; Wang, K.; Xue, J.; Li, S. Effect of the Rate of Nitrogen Application on Dry Matter Accumulation and Yield Formation of Densely Planted Maize. Sustainability 2022, 14, 14940. [Google Scholar] [CrossRef]
- Bolsen, K.K.; Ashbell, G.; Weinberg, Z.G. Silage fermentation and silage additives—Review. Asian Australas. J. Anim. Sci. 1996, 9, 483–494. [Google Scholar] [CrossRef]
- Wen-zhan, M. Effects of biological organic fertilizer on feeding value of Mexico corn. Prataculturalscience 2013, 30, 763r766. [Google Scholar]
- Jiang, H.; Wang, H.; Bao, B.; Qu, H.; Wang, J.; Sun, L.; Liu, B.; Gao, F. Effect of compound additives on nutritional composition, fermentation quality, and bacterial community of high-moisture alfalfa silage. Fermentation 2023, 9, 453. [Google Scholar] [CrossRef]
- Queiroz, O.C.; Arriola, K.G.; Daniel, J.L.; Adesogan, A.T. Effects of 8 chemical and bacterial additives on the quality of corn silage. J. Dairy Sci. 2013, 96, 5836–5843. [Google Scholar] [CrossRef]
- Jatkauskas, J.; Vrotniakiene, V.; Eisner, I.; Witt, K.L.; Copani, G. Effect of different conditions to forage before ensiling and use of a lactic acid bacteria-based additive on the fermentation and aerobic stability of maize silage. Zemdirb.-Agric. 2023, 110, 183–190. [Google Scholar] [CrossRef]
- Mu, L.; Xie, Z.; Hu, L.; Chen, G.; Zhang, Z. Cellulase interacts with Lactobacillus plantarum to affect chemical composition, bacterial communities, and aerobic stability in mixed silage of high-moisture amaranth and rice straw. Bioresour. Technol. 2020, 315, 123772. [Google Scholar] [CrossRef]
- Dunière, L.; Sindou, J.; Chaucheyras-Durand, F.; Chevallier, I.; Sergentet, D. Silage processing and strategies to prevent persistence of undesirable microorganisms. Anim. Feed. Sci. Technol. 2013, 182, 1–15. [Google Scholar] [CrossRef]
- Lv, J.; Fang, X.; Feng, G.; Zhang, G.; Zhao, C.; Zhang, Y.; Li, Y. Effects of Sodium Formate and Calcium Propionate Additives on the Fermentation Quality and Microbial Community of Wet Brewers Grains after Short-Term Storage. Animals 2020, 10, 1608. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Chang, J.; Yu, J.; Li, S.; Niu, H. Diversity of bacterial community during ensiling and subsequent exposure to air in whole-plant maize silage. Asian-Australas. J. Anim. Sci. 2018, 31, 1464–1473. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Benno, Y.; Ogawa, M.; Kumai, S. Effect of Applying Lactic Acid Bacteria Isolated from Forage Crops on Fermentation Characteristics and Aerobic Deterioration of Silage. J. Dairy Sci. 1999, 82, 520–526. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Si, Q.; Sun, L.; Wang, Z.; Liu, M.; Du, S.; Ge, G.; Jia, Y. Effects of Cellulase and Xylanase Addition on Fermentation Quality, Aerobic Stability, and Bacteria Composition of Low Water-Soluble Carbohydrates Oat Silage. Fermentation 2023, 9, 638. [Google Scholar] [CrossRef]
- Santos, E.M.; Da Silva, T.C.; Macedo, C.H.O.; Campos, F.S. Lactic Acid Bacteria in Tropical Grass Silages. In Lactic Acid Bacteria-R & D for Food, Health and Livestock Purposes; IntechOpen: London, UK, 2013. [Google Scholar]
- Ali, N.; Wang, S.; Zhao, J.; Dong, Z.; Li, J.; Nazar, M.; Shao, T. Microbial diversity and fermentation profile of red clover silage inoculated with reconstituted indigenous and exogenous epiphytic microbiota. Bioresour. Technol. 2020, 314, 123606. [Google Scholar] [CrossRef]
- Dewar, W.A.; McDonald, P.; Whittenbury, R. The hydrolysis of grass hemicelluloses during ensilage. J. Sci. Food Agric. 2010, 14, 411–417. [Google Scholar] [CrossRef]
- Yang, C.; Huang, B.; Lin, J.; Yang, Q.; Guo, Y.; Liu, D.; Sun, B. Isolation and screening of high biofilm producing lactic acid bacteria, and exploration of its effects on the microbial hazard in corn straw silage. J. Hazard. Mater. 2024, 480, 136009. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tissue | Treatment | V3 | V12 | R3 | R5 |
|---|---|---|---|---|---|
| stem | CK | 132.16 ± 0.58 Aa | 126.50 ± 0.10 a | 122.57 ± 0.19 Aa | 119.65 ± 0.14 Aa |
| JJ | 125.42 ± 0.40 Cc | 124.74 ± 0.55 b | 119.31 ± 0.07 Cc | 113.72 ± 0.07 Cc | |
| YJ | 128.88 ± 0.43 Bb | 125.20 ± 0.27 b | 121.49 ± 0.12 Bb | 115.83 ± 0.33 Bb | |
| leaf | CK | 130.86 ± 0.08 bc | 126.28 ± 0.04 Cc | 124.08 ± 0.21 Cc | 132.09 ± 0.02 Cc |
| JJ | 131.34 ± 0.18 a | 128.79 ± 0.09 Aa | 136.25 ± 0.19 Aa | 134.52 ± 0.16 Bb | |
| YJ | 130.54 ± 0.30 b | 127.94 ± 0.01 Bb | 134.73 ± 0.01 Bb | 137.77 ± 0.10 Aa | |
| grain | CK | - | - | 583.49 ± 3.71 Bb | 475.66 ± 15.46 |
| JJ | - | - | 636.65 ± 3.41 Aa | 469.44 ± 9.09 | |
| YJ | - | - | 469.45 ± 5.89 Cc | 463.90 ± 11.79 | |
| whole | CK | 150.21 ± 4.21 | 132.28 ± 0.79 b | 106.54 ± 0.63 Bb | 117.97 ± 0.41 |
| JJ | 146.95 ± 1.54 | 136.00 ± 0.40 a | 102.83 ± 0.59 Cc | 120.21 ± 2.89 | |
| YJ | 155.30 ± 0.89 | 133.63 ± 1.23 bc | 110.78 ± 0.32 Aa | 116.00 ± 0.71 |
| Treatment | Fertilization | DM | NDF | ADF | CP | EE | Starch | RFV |
|---|---|---|---|---|---|---|---|---|
| CK | CK | 28.17 ± 1.57 | 47.27 ± 0.06 Aa | 24.08 ± 0.11 Aa | 7.74 ± 0.01 Bc | 6.07 ± 0.08 Bb | 30.00 ± 0.08 | 138.05 ± 0.36 Cc |
| JJ | 26.85 ± 3.16 | 39.69 ± 0.03 Cc | 20.32 ± 0.04 Bc | 8.13 ± 0.02 Ab | 6.69 ± 0.01 Aa | 29.64 ± 0.35 | 171.27 ± 0.19 Aa | |
| YJ | 27.82 ± 1.95 | 45.32 ± 0.05 Bb | 23.57 ± 0.11 Ab | 8.2 ± 0.01 Aa | 6.05 ± 0.06 Bb | 29.98 ± 0.04 | 144.81 ± 0.33 Bb | |
| FJ | CK | 29.84 ± 1.30 a | 33.60 ± 0.74 Bb | 20.80 ± 0.04 Aa | 8.52 ± 0.02 Bb | 6.48 ± 0.09 Bb | 31.95 ± 0.10 Bb | 201.34 ± 4.36 Ab |
| JJ | 27.33 ± 2.08 ab | 30.62 ± 0.74 Bc | 19.94 ± 0.08 Bb | 8.82 ± 0.02 Aa | 7.76 ± 0.06 Aa | 32.32 ± 0.13 ABb | 222.99 ± 5.61 Aa | |
| YJ | 26.58 ± 1.01 b | 39.12 ± 0.04 Aa | 19.99 ± 0.11 Bb | 8.17 ± 0.01 Cc | 7.66 ± 0.03 Aa | 32.96 ± 0.19 Aa | 174.39 ± 0.37 Bc | |
| p-value | A | 0.316 | 0.208 | 0.355 | 0.594 | 0.281 | 0.459 | 0.163 |
| F | 0.751 | 0.470 | 0.142 | 0.205 | 0.097 | 0.014 | 0.040 | |
| A × F | 0.461 | <0.001 | <0.001 | <0.001 | <0.001 | 0.017 | <0.001 |
| Treament | Fertilization | pH | LA | AA | PA | BA |
|---|---|---|---|---|---|---|
| CK | CK | 3.96 ± 0.50 | 6.97 ± 0.00 Bb | ND | ND | ND |
| JJ | 3.72 ± 0.06 | 6.96 ± 0.01 Bb | ND | ND | ND | |
| YJ | 3.76 ± 0.02 | 7.83 ± 0.01 Aa | ND | ND | ND | |
| FJ | CK | 3.79 ± 0.01 Aa | 7.88 ± 0.03 | ND | ND | ND |
| JJ | 3.66 ± 0.02 Cc | 7.92 ± 0.00 | ND | ND | ND | |
| YJ | 3.72 ± 0.01 Bb | 7.94 ± 0.02 | ND | ND | ND | |
| p-value | A | 0.110 | 0.459 | - | - | - |
| F | 0.142 | 0.139 | - | - | - | |
| A × F | 0.851 | <0.001 | - | - | - |
| Treament | Fertilization | Ace | Chao1 | Shannon | Simpson | Coverage |
|---|---|---|---|---|---|---|
| CK | CK | 223.10 ± 16.86 c | 221.20 ± 16.22 ab | 2.19 ± 0.17 a | 0.23 ± 0.04 b | 0.999 ± 0.0004 a |
| JJ | 234.30 ± 19.67 b | 205.40 ± 15.94 b | 1.26 ± 0.33 b | 0.51 ± 0.11 Aa | 0.999 ± 0.00003 a | |
| YJ | 267.50 ± 47.23 a | 252.50 ± 45.71 a | 1.95 ± 0.44 a | 0.27 ± 0.08 b | 0.999 ± 0.0003 a | |
| FJ | CK | 126.20 ± 7.90 C | 117.70 ± 19.88 Cc | 2.05 ± 0.37 a | 0.22 ± 0.06 b | 1.000 ± 0.0002 a |
| JJ | 234.80 ± 16.21 B | 221.80 ± 33.37 ab | 1.09 ± 0.21 Bb | 0.54 ± 0.08 Aa | 0.999 ± 0.0002 a | |
| YJ | 246.30 ± 50.05 A | 228.60 ± 59.00 a | 1.95 ± 0.21 a | 0.24 ± 0.06 b | 0.999 ± 0.0003 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hou, M.; Li, Y.; Yang, F.; Cui, W.; Huang, X.; Dong, D.; Dong, L.; Zhang, B. Optimizing the Production of High-Quality Silage from Jingkenuo 2000 Fresh Waxy Maize: The Synergistic Effects of Microbial Fertilizer and Fermentation Agents. Fermentation 2025, 11, 151. https://doi.org/10.3390/fermentation11030151
Hou M, Li Y, Yang F, Cui W, Huang X, Dong D, Dong L, Zhang B. Optimizing the Production of High-Quality Silage from Jingkenuo 2000 Fresh Waxy Maize: The Synergistic Effects of Microbial Fertilizer and Fermentation Agents. Fermentation. 2025; 11(3):151. https://doi.org/10.3390/fermentation11030151
Chicago/Turabian StyleHou, Min, Yang Li, Fan Yang, Weidong Cui, Xinxin Huang, Deli Dong, Lifeng Dong, and Bo Zhang. 2025. "Optimizing the Production of High-Quality Silage from Jingkenuo 2000 Fresh Waxy Maize: The Synergistic Effects of Microbial Fertilizer and Fermentation Agents" Fermentation 11, no. 3: 151. https://doi.org/10.3390/fermentation11030151
APA StyleHou, M., Li, Y., Yang, F., Cui, W., Huang, X., Dong, D., Dong, L., & Zhang, B. (2025). Optimizing the Production of High-Quality Silage from Jingkenuo 2000 Fresh Waxy Maize: The Synergistic Effects of Microbial Fertilizer and Fermentation Agents. Fermentation, 11(3), 151. https://doi.org/10.3390/fermentation11030151