
Lactic Acid Bacterial Fermentation of Esterified Agave Fructans in Simulated Physicochemical Colon Conditions for Local Delivery of Encapsulated Drugs

 ,
,  , and
, and
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
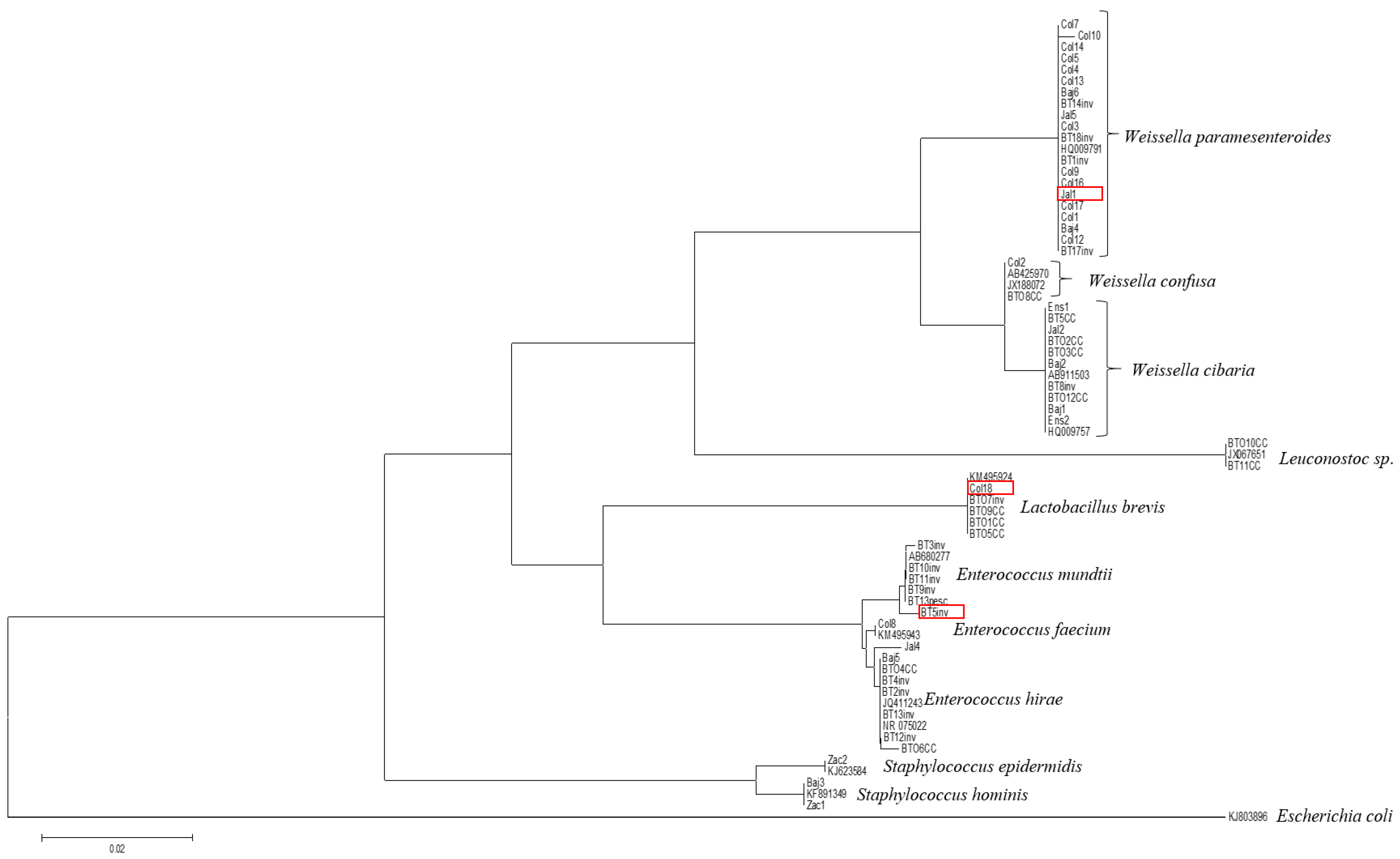
2.2.1. Identification of Lactic Acid Bacteria Strains
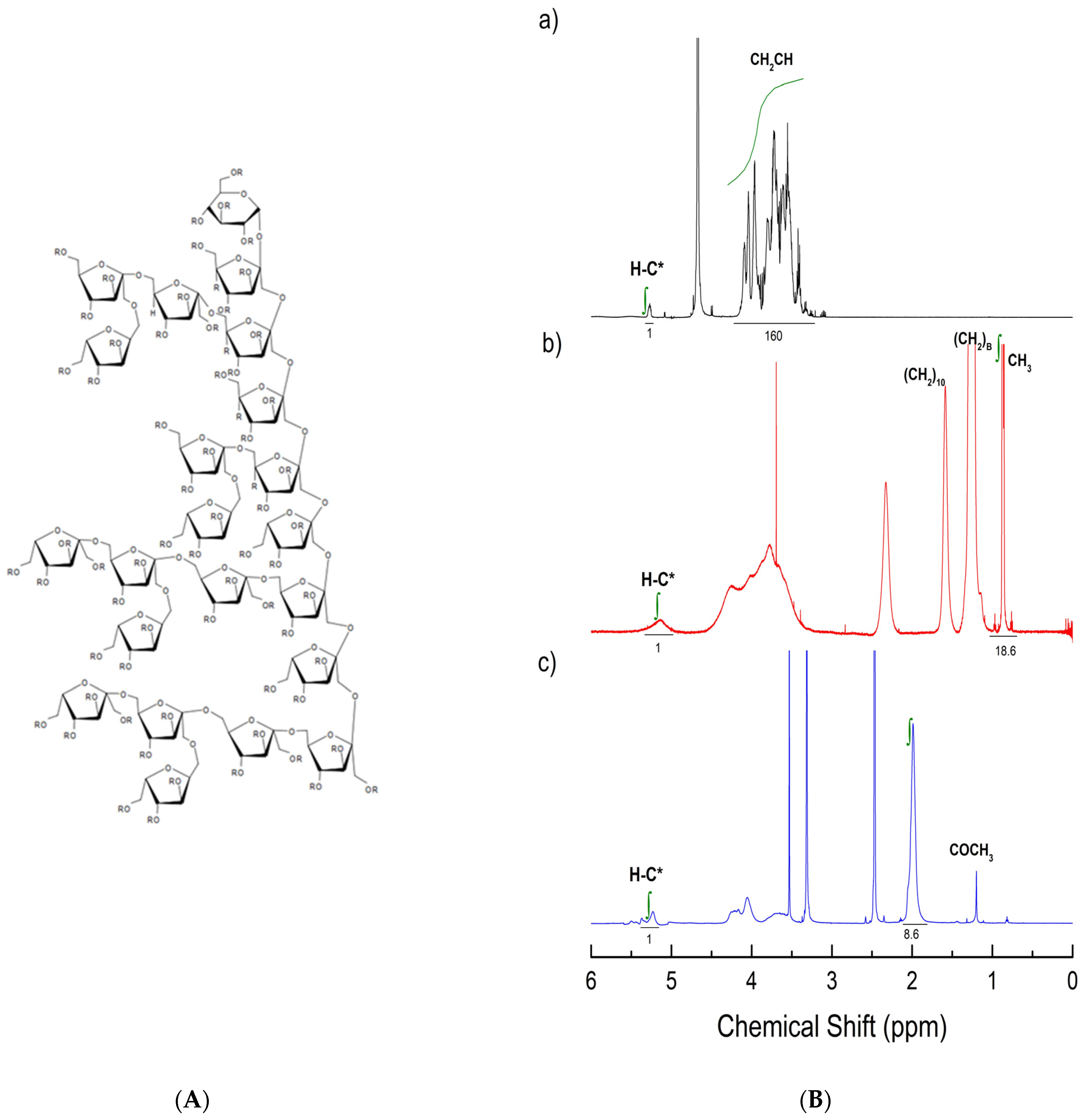
2.2.2. Esterification and NMR Characterization of Agave Fructans
2.2.3. Microencapsulation of Ibuprofen
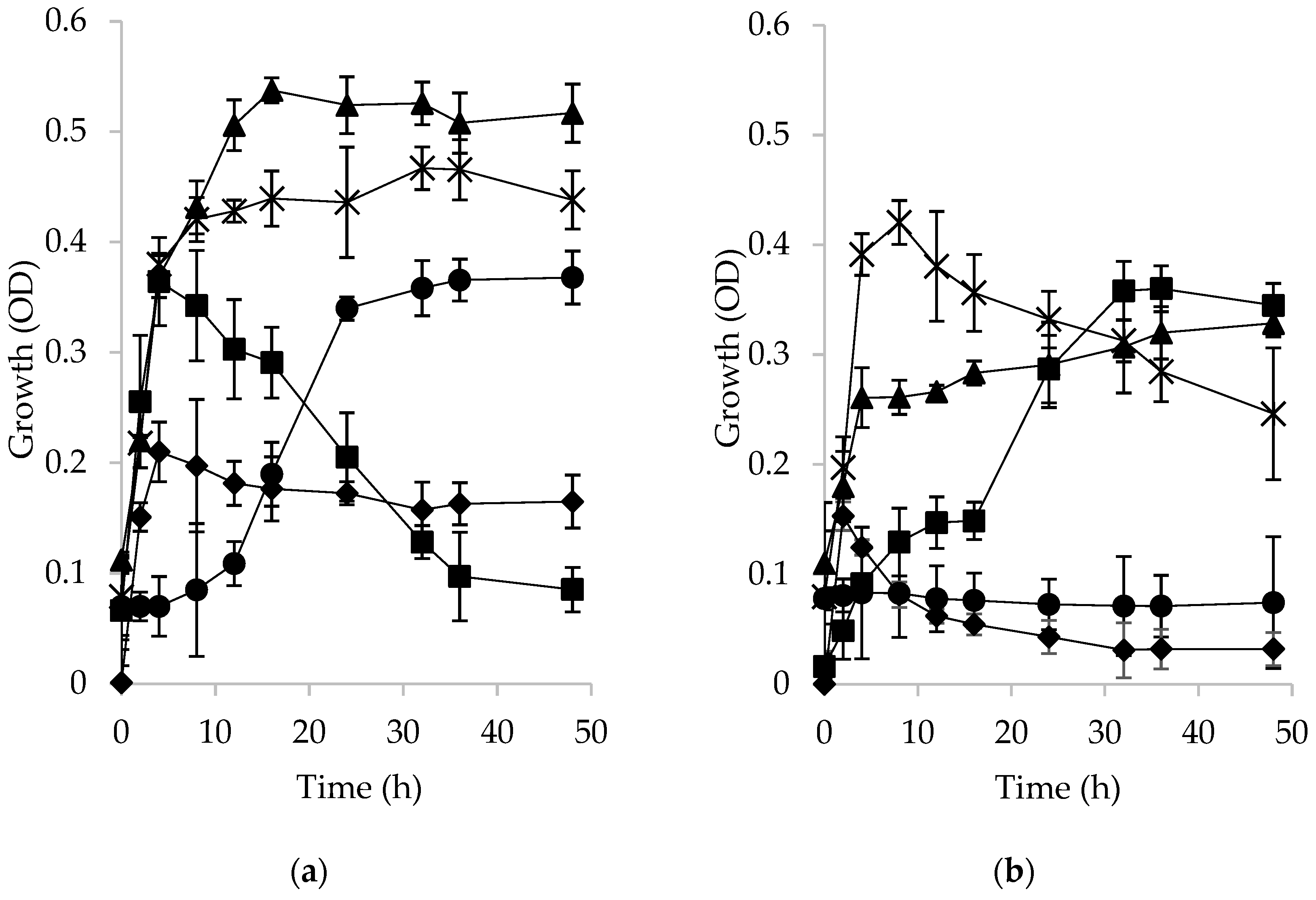
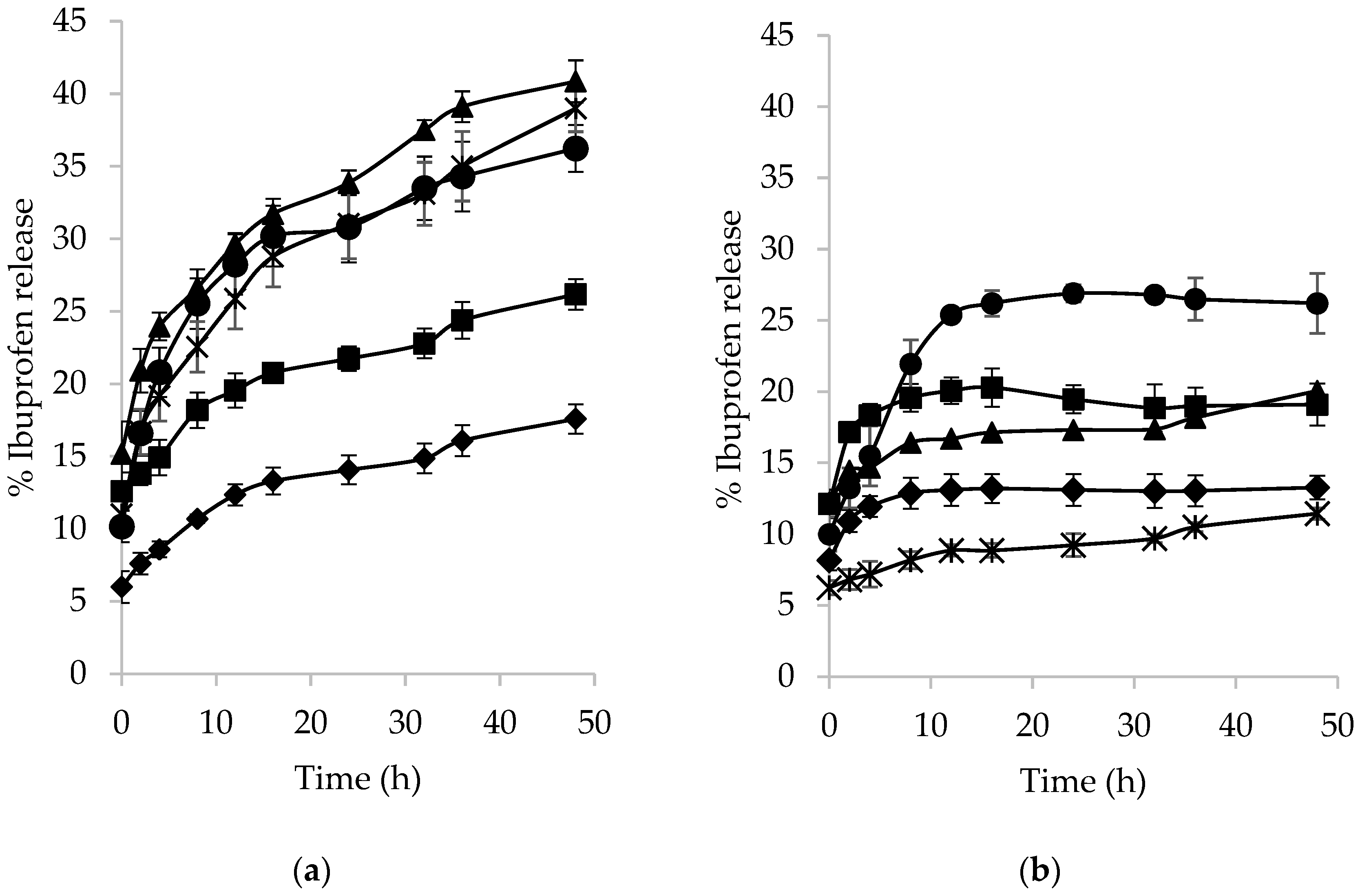
2.2.4. Growth of Lactic Acid Bacteria on Esterified Fructan Microspheres and Release of Ibuprofen
3. Results
3.1. Identification of Lactic Acid Bacteria Strains
3.2. NMR Characterization
3.3. Morphological Characterization of Microspheres
3.4. Bacterial Growth in Acetylated or Palmitoylated Fructan Microspheres
3.5. Release of Ibuprofen from Derivatized-Fructan Microspheres
3.6. Production of Short-Chain Fatty Acids (SCFAs)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nadeem, M.S.; Kumar, V.; Al-Abbasi, F.A.; Kamal, M.A.; Anwar, F. Risk of colorectal cancer in inflammatory bowel diseases. Semin. Cancer Biol. 2020, 64, 51–60. [Google Scholar] [CrossRef]
- Amidon, S.; Brown, J.E.; Dave, V.S. Colon-targeted oral drug delivery systems: Design trends and approaches. AAPS Pharmscitech 2015, 16, 731–741. [Google Scholar] [CrossRef] [PubMed]
- Hua, S. Advances in Oral Drug Delivery for Regional Targeting in the Gastrointestinal Tract-Influence of Physiological, Pathophysiological and Pharmaceutical Factors. Front. Pharmacol. 2020, 11, 524. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.I.; Željko Prebeg, Ž.; Kurjaković, N. A pH-dependent colon targeted oral drug delivery system using methacrylic acid copolymers: I. Manipulation of drug release using Eudragit® L100-55 and Eudragit® S100 combinations. J. Control. Release 1999, 58, 215–222. [Google Scholar] [CrossRef]
- Kotla, N.G.; Rana, S.; Sivaraman, G.; Sunnapu, O.; Vemula, P.K.; Pandit, A.; Rochev, Y. Bioresponsive drug delivery systems in intestinal inflammation: State-of-the-art and future perspectives. Adv. Drug Deliv. Rev. 2019, 146, 248–266. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Bajracharya, R.; Min, J.Y.; Han, J.W.; Park, B.J.; Han, H.K. Strategic Approaches for Colon Targeted Drug Delivery: An Overview of Recent Advancements. Pharmaceutics 2020, 12, 68. [Google Scholar] [CrossRef]
- Arévalo-Pérez, R.; Maderuelo, C.; Lanao, J.M. Recent advances in colon drug delivery systems. J. Control. Release 2020, 327, 703–724. [Google Scholar] [CrossRef]
- Barclay, T.G.; Day, C.M.; Petrovsky, N.; Garg, S. Review of polysaccharide particle-based functional drug delivery. Carbohydr. Polym. 2019, 221, 94–112. [Google Scholar] [CrossRef]
- Miramontes-Corona, C.; Escalante, A.; Delgado, E.; Corona-González, R.I.; Vázquez-Torres, H.; Toriz, G. Hydrophobic agave fructans for sustained drug delivery to the human colon. React. Funct. Polym. 2020, 146, 104396. [Google Scholar] [CrossRef]
- Amaretti, A.; Bernardi, T.; Tamburini, E.; Zanoni, S.; Lomma, M.; Matteuzzi, D.; Rossi, M. Kinetics and Metabolism of Bifidobacterium adolescentis MB 239 Growing on Glucose, Galactose, Lactose, and Galactooligosaccharides. Appl. Environ. Microbiol. 2007, 73, 3637–3644. [Google Scholar] [CrossRef]
- Trevisi, P.; De Filippi, S.; Modesto, M.; Mazzoni, M.; Casini, L.; Biavati, B.; Bosi, P. Investigation on the ability of different strains and doses of exogenous Bifidobacteria, to translocate in the liver of weaning pigs. Livest. Sci. 2007, 108, 109–112. [Google Scholar] [CrossRef]
- Holscher, H.D. Dietary fiber and prebiotics and the gastrointestinal microbiota. Gut Microbes 2017, 8, 172–184. [Google Scholar] [CrossRef] [PubMed]
- Ashaolu, T.; Ashaolu, J.; Adeyeye, S. Fermentation of prebiotics by human colonic microbiota in vitro and short-chain fatty acids production: A critical review. J. Appl. Microbiol. 2021, 130, 677–687. [Google Scholar] [CrossRef] [PubMed]
- Delzenne, N.M.; Williams, C.M. Prebiotics and lipid metabolism. Curr. Opin. Lipidol. 2002, 13, 61–67. [Google Scholar] [CrossRef]
- Urías-Silvas, J.E.; López, M.G. Efecto Prebiótico de los Fructanos de Agave. 1er Encuentro Participación de La Mujer En La Ciencia. 2004. Available online: https://congresos.cio.mx/1_enc_mujer/files/Extensos/Posters/B-03.pdf (accessed on 22 July 2024).
- Macfarlane, S. Chapter 10—Prebiotics in the Gastrointestinal Tract. In Bioactive Foods in Promoting Health; Watson, R.R., Preedy, V.R., Eds.; Academic Press: Boston, MA, USA, 2010; pp. 145–156. [Google Scholar] [CrossRef]
- Catry, E.; Bindels, L.B.; Tailleux, A.; Lestavel, S.; Neyrinck, A.M.; Goossens, J.F.; Lobysheva, I.; Plovier, H.; Essaghir, A.; Demoulin, J.B.; et al. Targeting the gut microbiota with inulin-type fructans: Preclinical demonstration of a novel approach in the management of endothelial dysfunction. Gut 2018, 67, 271–283. [Google Scholar] [CrossRef]
- Yang, Q.; Liang, Q.; Balakrishnan, B.; Belobrajdic, D.P.; Feng, Q.J.; Zhang, W. Role of Dietary Nutrients in the Modulation of Gut Microbiota: A Narrative Review. Nutrients 2020, 12, 381. [Google Scholar] [CrossRef]
- Hua, S.; Marks, E.; Schneider, J.J.; Keely, S. Advances in oral nano-delivery systems for colon targeted drug delivery in inflammatory bowel disease: Selective targeting to diseased versus healthy tissue. Nanomed. Nanotechnol. Biol. Med. 2015, 11, 1117–1132. [Google Scholar] [CrossRef]
- Rafter, J.; Bennett, M.; Caderni, G.; Clune, Y.; Hughes, R.; Karlsson, P.C.; Klinder, A.; O’Riordan, M.; O’Sullivan, G.C.; Pool-Zobel, B.; et al. Dietary synbiotics reduce cancer risk factors in polypectomized and colon cancer patients. Am. J. Clin. Nutr. 2007, 85, 488–496. [Google Scholar] [CrossRef]
- Drago, L. Probiotics and Colon Cancer. Microorganisms 2019, 7, 66. [Google Scholar] [CrossRef]
- Le, B.; Ngoc, A.; Yang, S. Synbiotic Fermented Soymilk with Weissella cibaria FB069 and Xylooligosaccharides Prevents Proliferation in Human Colon Cancer Cells. J. Appl. Microbiol. 2020, 128, 1486–1496. [Google Scholar] [CrossRef]
- Davani-Davari, D.; Negahdaripour, M.; Karimzadeh, I.; Seifan, M.; Mohkam, M.; Masoumi, S.J.; Berenjian, A.; Ghasemi, Y. Prebiotics: Definition, Types, Sources, Mechanisms, and Clinical Applications. Foods 2019, 8, 92. [Google Scholar] [CrossRef] [PubMed]
- Partida, V.Z.; Lopez, A.C.; Gomez, A.d.J.M. Method of Producing Fructose Syrup from Agave Plants. U.S. Patent 5,846,333, 8 December 1998. [Google Scholar]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular Evolutionary Genetics Analysis Using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. Phylogenies and the Comparative Method. Am. Nat. 1985, 125, 1–15. [Google Scholar] [CrossRef]
- Rogge, T.M.; Stevens, C.V.; Colpaert, A.; Levecke, B.; Booten, K. Use of Acyl Phosphonates for the Synthesis of Inulin Esters and Their Use as Emulsion Stabilizing Agents. Biomacromolecules 2007, 8, 485–489. [Google Scholar] [CrossRef] [PubMed]
- Poulain, N.; Dez, I.; Perrio, C.; Lasne, M.C.; Prud’homme, M.P.; Nakache, E. Microspheres based on inulin for the controlled release of serine protease inhibitors: Preparation, characterization and in vitro release. J. Control. Release 2003, 92, 27–38. [Google Scholar] [CrossRef]
- Starbird, R.; Zuñiga, V.; Delgado, E.; Saake, B.; Toriz, G. Design of Microspheres from Blue Agave Fructans for Drug Delivery to the Colon. Part 1. Esterification of Agave Fructans. J. Biobased Mater. Bioenergy 2007, 1, 238–244. [Google Scholar] [CrossRef]
- Arellano-Ayala, K.; Ascencio-Valle, F.; Gutiérrez-González, P.; Estrada-Girón, Y.; Torres-Vitela, M.; Macías-Rodríguez, M. Hydrophobic and adhesive patterns of lactic acid bacteria and their antagonism against foodborne pathogens on tomato surface (Solanum lycopersicum L.). J. Appl. Microbiol. 2020, 129, 876–891. [Google Scholar] [CrossRef]
- Delgadillo, E.; Corona, R.I.; Toriz, G.; Contreras, H.J.; Sadeghifar, H.; Baobing, W.; Yang, G.; Lucia, L.A.; Delgado, E. Coacervated liposoluble fructan-based host–guest microspheres as unique drug delivery materials. RSC Adv. 2015, 5, 67759–67766. [Google Scholar] [CrossRef]
- Toriz, G.; Delgado, E.; Zúñiga, V. A proposed chemical structure for fructans from blue agave plant (Tequilana weber var. azul). e-Gnosis 2007, 5, 1. [Google Scholar]
- Kesharwani, S.S.; Dachineni, R.; Bhat, G.J.; Tummala, H. Hydrophobically modified inulin-based micelles: Transport mechanisms and drug delivery applications for breast cancer. J. Drug Deliv. Sci. Technol. 2019, 54, 101254. [Google Scholar] [CrossRef]
- Walz, M.; Hirth, T.; Weber, A. Investigation of chemically modified inulin as encapsulation material for pharmaceutical substances by spray-drying. Colloids Surf. A Physicochem. Eng. Asp. 2018, 536, 47–52. [Google Scholar] [CrossRef]
- Shivhare, K.; Garg, C.; Priyam, A.; Gupta, A.; Sharma, A.K.; Kumar, P. Enzyme sensitive smart inulin-dehydropeptide conjugate self-assembles into nanostructures useful for targeted delivery of ornidazole. Int. J. Biol. Macromol. 2018, 106, 775–783. [Google Scholar] [CrossRef] [PubMed]
- Pereira, A.K.S.; Reis, D.T.; Barbosa, K.M.; Scheidt, G.N.; da Costa, L.S.; Santos, L.S.S. Antibacterial effects and ibuprofen release potential using chitosan microspheres loaded with silver nanoparticles. Carbohydr. Res. 2020, 488, 107891. [Google Scholar] [CrossRef]
- Sauer, M.; Russmayer, H.; Grabherr, R.; Peterbauer, C.K.; Marx, H. The efficient clade: Lactic acid bacteria for industrial chemical production. Trends Biotechnol. 2017, 35, 756–769. [Google Scholar] [CrossRef]
- Nelson David, L.; Cox Michael, M.; Nelson David, L. Lehninger Principles of Biochemistry; WH Freeman: New York, NY, USA, 2005. [Google Scholar]
- Buitrago-Arias, C.; Londoño-Moreno, A.; Avila-Reyes, S.; Arenas-Ocampo, M.; Alamilla-Beltran, L.; Jimenez-Aparicio, A.; Camacho-Diaz, B. Evaluation of the fermentation of acetylated agave fructans (agavins), with Saccharomyces boulardii as a probiotic. Rev. Mex. Ing. Química 2021, 20. [Google Scholar] [CrossRef]
- Gänzle, M.G.; Vermeulen, N.; Vogel, R.F. Carbohydrate, peptide and lipid metabolism of lactic acid bacteria in sourdough. Food Microbiol. 2007, 24, 128–138. [Google Scholar] [CrossRef]
- Hu, Y.; Zhang, L.; Wen, R.; Chen, Q.; Kong, B. Role of lactic acid bacteria in flavor development in traditional Chinese fermented foods: A review. Crit. Rev. Food Sci. Nutr. 2022, 62, 2741–2755. [Google Scholar] [CrossRef]
- Toshimitsu, T. Development of a lactic acid bacteria strain that suppresses chronic inflammation and improves glucose and lipid metabolism. Biosci. Microbiota Food Health 2023, 42, 3–7. [Google Scholar] [CrossRef]
- Bintsis, T. Lactic acid bacteria as starter cultures: An update in their metabolism and genetics. AIMS Microbiol. 2018, 4, 665. [Google Scholar] [CrossRef]
- Kaushik, D.; Sardana, S.; Mishra, D. 5-fluorouracil loaded guar gum microspheres for colon delivery: Preparation, characterization and in vitro release. Yao Xue Xue Bao = Acta Pharm. Sin. 2009, 44, 1278–1284. [Google Scholar]
- Liu, Y.; Zhou, H. Budesonide-Loaded Guar Gum Microspheres for Colon Delivery: Preparation, Characterization and in Vitro/in Vivo Evaluation. Int. J. Mol. Sci. 2015, 16, 2693–2704. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, I.M. Advances in polysaccharide-based oral colon-targeted delivery systems: The journey so far and the road ahead. Cureus 2023, 15, e33636. [Google Scholar] [CrossRef] [PubMed]
- Kotla, N.; Shivapooja, A.; Muthyala, J.; Pinakin, P. Effect of guar gum and xanthan gum compression coating on release studies of metronidazole in human faecal media for colon targeted drug delivery system. Asian J. Pharm. Clin. Res. 2013, 6, 310–313. [Google Scholar]
- Usman, M.; Zhang, C.; Patil, P.J.; Mehmood, A.; Li, X.; Bilal, M.; Haider, J.; Ahmad, S. Potential applications of hydrophobically modified inulin as an active ingredient in functional foods and drugs—A review. Carbohydr. Polym. 2021, 252, 117176. [Google Scholar] [CrossRef]
- Sharon, G.; Garg, N.; Debelius, J.; Knight, R.; Dorrestein, P.C.; Mazmanian, S.K. Specialized metabolites from the microbiome in health and disease. Cell Metab. 2014, 20, 719–730. [Google Scholar] [CrossRef]

 ) propionic acid. BA: B. adolescentis; WP: Weisella paramesenteroides; EL: Enterococcus mundtii; LB: Lactobacillus brevis; C: consortium.
) propionic acid. BA: B. adolescentis; WP: Weisella paramesenteroides; EL: Enterococcus mundtii; LB: Lactobacillus brevis; C: consortium.
) propionic acid. BA: B. adolescentis; WP: Weisella paramesenteroides; EL: Enterococcus mundtii; LB: Lactobacillus brevis; C: consortium.
) propionic acid. BA: B. adolescentis; WP: Weisella paramesenteroides; EL: Enterococcus mundtii; LB: Lactobacillus brevis; C: consortium.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miramontes-Corona, C.; Cetina-Corona, A.; Macías-Rodríguez, M.E.; Escalante, A.; Corona-González, R.I.; Toriz, G. Lactic Acid Bacterial Fermentation of Esterified Agave Fructans in Simulated Physicochemical Colon Conditions for Local Delivery of Encapsulated Drugs. Fermentation 2024, 10, 478. https://doi.org/10.3390/fermentation10090478
Miramontes-Corona C, Cetina-Corona A, Macías-Rodríguez ME, Escalante A, Corona-González RI, Toriz G. Lactic Acid Bacterial Fermentation of Esterified Agave Fructans in Simulated Physicochemical Colon Conditions for Local Delivery of Encapsulated Drugs. Fermentation. 2024; 10(9):478. https://doi.org/10.3390/fermentation10090478
Chicago/Turabian StyleMiramontes-Corona, Carmen, Abraham Cetina-Corona, María Esther Macías-Rodríguez, Alfredo Escalante, Rosa Isela Corona-González, and Guillermo Toriz. 2024. "Lactic Acid Bacterial Fermentation of Esterified Agave Fructans in Simulated Physicochemical Colon Conditions for Local Delivery of Encapsulated Drugs" Fermentation 10, no. 9: 478. https://doi.org/10.3390/fermentation10090478
APA StyleMiramontes-Corona, C., Cetina-Corona, A., Macías-Rodríguez, M. E., Escalante, A., Corona-González, R. I., & Toriz, G. (2024). Lactic Acid Bacterial Fermentation of Esterified Agave Fructans in Simulated Physicochemical Colon Conditions for Local Delivery of Encapsulated Drugs. Fermentation, 10(9), 478. https://doi.org/10.3390/fermentation10090478