Abstract
Seaweed is a fast-growing biomass source that is currently studied as feedstock for sustainable industrial production in a wide variety of markets. Being composed mostly of polysaccharides, macroalgae can be integrated in biorefineries for obtaining bioproducts via fermentation. Kappaphycus alvarezii has been introduced experimentally to Brazil’s south coastline in 1995 and is now cultivated on a large scale to keep up with the high carrageenan demand in various industrial sectors. In this review article, an introduction is given on renewable biomass and environmental issues, focusing especially on third-generation biomass and its promising features and use advantages. Later on, the processing of K. alvarezii for the use of its saccharide portion for fermentative processes is approached. The current state of research conducted alongside challenges and hurdles in K. alvarezii hydrolysate fermentation processes provides insight into future studies needed to make new fermentation processes viable. Next, some fermentation products are discussed, and the metabolism of galactose in microorganisms is also presented to bring to light other possible fermentation products that are not yet, but can be, obtained from K. alvarezii. Finally, a simple and comprehensive scheme for K. alvarezii fermentation biorefinery is presented to demonstrate a generic example for a possible configuration for obtaining valuable bio-products. In the literature, production of ethanol and lactic acid were already reported from K. alvarezii. This review aims to help envision new industrial processes that can be developed for this most valuable macroalga.
1. Introduction
Over the last decades, finding new biomass sources for chemical compound and commodity production is being researched to eventually substitute similar fossil-based products. This intense search emerged along with the rise in public awareness of global warming, which, in turn, could be directly related to large carbon dioxide emissions caused by the need to meet mankind civilization’s energy requirements.
Large-scale chemical industries usually have high energy requirements, leading to a relatively high carbon footprint. The carbon footprint theme is relatively new, being studied as an index for the contribution of individuals, companies, and government entities to global warming [1,2]. Sustainability is the measure of human production in harmony with nature and maintenance of planet Earth’s climate stability [3].
Carbon dioxide emissions have increased from 2600 Mt in 1960 to 3500 Mt in 2018 in the European Union alone [4]. Globally, there is still an increase in CO2 emissions. Liu et al. [5] reported that the emissions in 2022 were 1.5% higher than the prior year. The use of alternative energy sources can mitigate the increase in carbon emissions in order to reach emission goals set by international agreements.
Renewable raw materials are natural biological resources, continuously available for human consumption with no risk of running out if not used excessively [6]. The environmental impact of a specific bio-based industrial process can be evaluated by a life cycle assessment (LCA), a quantifying tool that includes the raw materials, production process, product distribution, use, and waste treatment as factors for measuring negative environmental effects [7]. Carbon emission mass balances can be used as an indicator of the carbon footprint of specific bioprocesses. As shown by Terlouw et al. [8], mass balance for carbon dioxide emissions should include farming, plantation, land use, logistics, industrial process, and distribution of products to accurately calculate the carbon input or output of a given bio-based process.
Algal biomass is estimated to be the most probable focus on biorefinery study investment for the next decades in both the US and European Union [9,10]. One of the reasons algal biomass is an attractive feedstock is its fast growth rates and carbon dioxide sequestration, which lead to a lower carbon footprint. Moreover, seaweed requires less land use for cultivation compared to terrestrial crops and can even be grown in artificial bodies of water when given a circulation of seawater to provide nutrients [11].
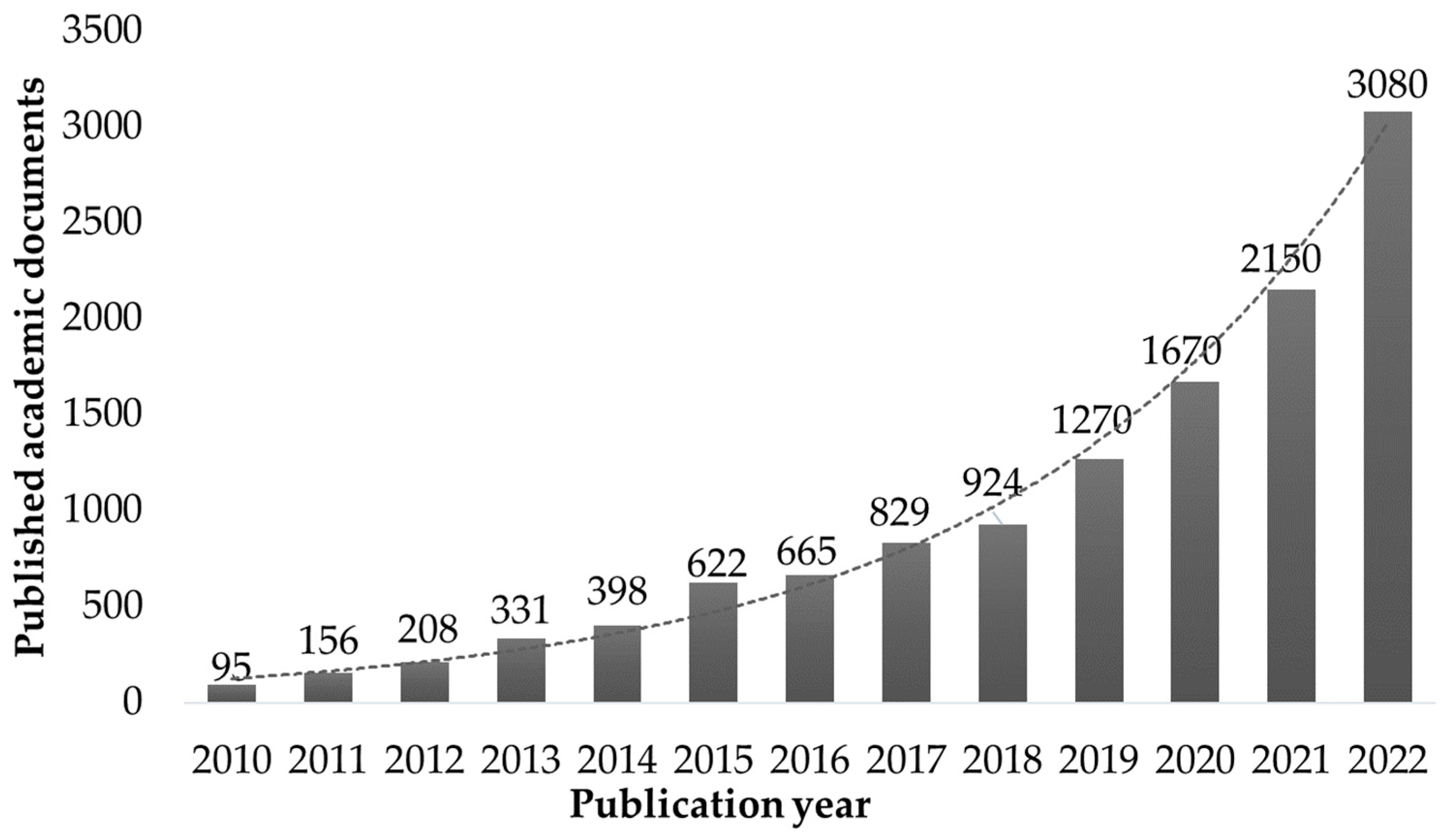
Macroalgae are composed mainly of polysaccharides, making them adequate as feedstock for producing fermentation products [12]. Most species are rich in valuable minerals and salts absorbed from seawater, which can be extracted alongside proteins and antioxidants with minimal environmental impact [13,14,15]. In the last decade, interest in seaweed refining processes has increased, with a current number of academic publications using the terms “biorefinery” and “macroalgae” in the same document larger in a two-order magnitude than the quantity published in the beginning of the second decade of the 21st century (Figure 1).
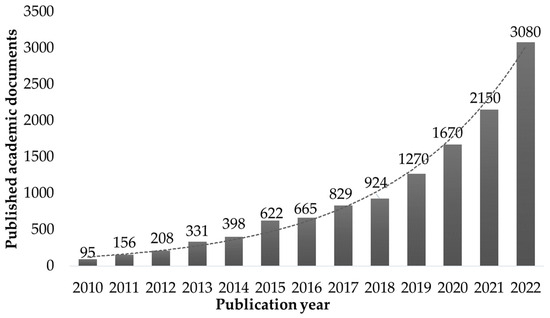
Figure 1.
Number of yearly publications of academic papers which use both the term “biorefinery” and “macroalgae” (data mined from Google Scholar search engine).
Kappaphycus alvarezii is a red seaweed which consists of carrageenan, a galactan polysaccharide, and linear cellulose. These natural polymers can be degraded mainly into two fermentable monomers, glucose and galactose. Carrageenan content can reach up to 49% of the algal biomass [16], and the exploitation of the species for its refining products has more than doubled over the last two decades [17].
The implementation of a K. alvarezii fermentation biorefinery process presents some critical hurdles, first of which is economic viability due to the high raw material price which is about four times higher than the sugarcane bagasse acquisition cost [18,19]. Government policies that provide incentives for farming, the development of new products, and scientific research could make using K. alvarezii more profitable, the same way as first-generation ethanol production was made more efficient in Brazil [20].
Fermentable saccharides from algal biomass can be obtained via dilute-acid and enzymatic hydrolysis, both of which can be used in sequence [21,22]. High-temperature dilute-acid hydrolysis of carrageenan leads to solutions containing high concentrations of 5-hydroxymethylfurfural (HMF), a compound that has inhibitory effects on fermentation processes [23]. In some cases, HMF should be removed from fermentation media prior to the microorganism inoculation. The removal of HMF could be carried out using activated charcoal [22,24], and overliming processes [24,25,26,27]. Alternatively, HMF can be removed directly from the hydrolysates, applying reactive extraction via zeolites as catalysts or using absorption in an organic phase in slug flow reactors or stripping columns [28]. It should be noted that HMF is a valuable molecule for some chemical industries, and it could be used as a precursor of levulinic and formic acids [13], as well as a building block for more complex added-value products [29].
Enzymatic hydrolysis can be applied to the cellulose content of K. alvarezii biomass using commercial enzyme cocktails [21], and the carrageenan itself can be reduced to monomers enzymatically with enzymes obtained from marine microflora [30]. Enzymatic hydrolysis provides specific chemical manipulation and bond cleavage, ensuring the preservation of other valuable compounds present in algal biomass, such as pigments, proteins, and phenolic compounds [31]. Application of marine-sourced enzymes is at the forefront of algae-polysaccharide degradation processes. In the case of K. alvarezii, the carrageenan and linear cellulose content can be reduced to oligomers with antioxidant properties and fermentable monomers.
Some K. alvarezii biorefinery schemes are available in the literature. Álvarez-Viñas et al. [13] show four different configurations for K. alvarezii processing, three with bio stimulants, ethanol, fertilizers, and biogas products in addition to carrageenan extraction. The other biorefinery presented is for a thermochemical process that produces HMF, levulinic and formic acids. This last configuration uses the biomass residue for energy generation via combustion. Torres et al. [12] present a K. alvarezii biorefining process where ethanol is produced from algal biomass residue after carrageenan extraction.
K. alvarezii is one of the most widely cultivated seaweeds on the planet, a fact that makes it readily available as raw material for obtaining bioproducts. This paper aims to relay information about the processing of K. alvarezii biomass for fermentation processes, including acid hydrolysis, detoxification and enzymatic hydrolysis. A short review of galactose metabolic pathways brings to light potential products that can be obtained from the algal biomass via fermentation.
2. Classification of Renewable Raw Materials
Biofuels are popularly classified in generations according to the raw materials used as feedstock to produce them. This “generation” term is also extended in the literature to biorefineries dividing them into generation groups by the same criteria [32,33]. Biomasses, like biofuels, can be divided in the same manner. First-generation (1G) biomass is essentially food products, extracted from agricultural crops such as sugarcane, wheat, corn, etc. Second-generation (2G) biomass is lignocellulosic material obtained from agricultural product waste, or other biomass not used for human consumption. Biofuels derived from algal biomass, being it microalgae or macroalgae, can be defined as third-generation (3G) biofuels [3,34]; thus, algal biomass can be defined as third-generation biomass. The literature mentions also a fourth-generation biomass, which consists of microorganisms, such as cyanobacteria, with genetically engineered metabolic pathways, that can convert sunlight directly into biofuels and other desired products [34,35,36].
Alternatively, raw materials can be classified according to their composition as suggested by McKendry in 2002 [37]. The humidity, caloric value, and the proportion of carbon, ash, alkaline metals, as well as the cellulose-to-lignin ratio can determine the potential products that could be obtained from any given biomass. This analysis helps to evaluate if a certain biomass can be used as a drop-in material in already-functioning industries. In any case, the choice of an appropriate raw material is a cumbersome task that requires considering logistics of transportation, plantation, and harvest. Moreover, all process technological aspects should be taken into account for the creation of a successful industrial endeavor.
It is worth mentioning that all renewable sources, especially for the energy sector, should be explored in roadmap plans for a sustainable future. This idea has been stressed in the literature for a long time [3,34,38]. Improvement in technology for 1st-, 2nd-, and 3rd-generation biofuels, making them more economically viable, will lead to the gradual substitution of fossil-based fuels. Eventually, promising technology such as cultivation of microalgal biomass would be made viable, enabling the conversion of sunlight directly into valuable bioproducts in photoreactors [39,40].
2.1. Raw Material of the 1st and 2nd Generations
1G bioprocess technologies are well consolidated, and their products are commercialized worldwide. Although renewable, this type of biomass cannot, on its own, guarantee sustainability in the long run, for being food products further converted to valuable products, the increase in world population could strain the food and energy market. Various authors have alerted about possible direct food market and bioproduct conflicts, which can cause price oscillations and consumer goods shortages [41,42,43].
In contrast to 1G biomass, 2G biomass is inedible, unaffecting the food market, and not creating a socioeconomic debate about nutritional resource division and competition. The cultivation of raw material for 2G industries creates opportunities for companies, generating jobs that boost economic development. Major players in the chemical industry are already investing in 2G biorefineries [42,44,45].
The use of renewable raw material promotes sustainability better than non-renewable fossil-based resources; however, obtaining products with no negative environmental impact is extremely hard to achieve. Most 1G and 2G biomasses require in-land cultivation, which can cause a delay in carbon gas sequestration from the atmosphere [46].
Replacement of wild flora by cropland could potentially create a long deficit in carbon fixation. The lack of carbon sequestration could take long periods of time to compensate, from 40 to 120 years with high-yield crops, and reach up to 1500 years in the case of low-yielding crops such as maize and soybeans [47]. Harper et al. [46] stress that forestation mitigation plans can be used to achieve environmental global warming goals.
Even given that the CO2 fixation of crop replantation and growth can account for some of the industrial process emissions, the destruction of local flora when preparing land for farming exploration can force wildlife from its habitat and disrupt the process of evapotranspiration, responsible for the planet’s healthy water cycle [48]. Indirect land use by industries is also underestimated and marked as a potential, unaccounted-for, greenhouse gas emission cause [49]. Without using the appropriate boundaries for carbon life cycle calculations, processes could mistakably present negative emissions [50].
2.2. Macroalgae: Third-Generation Raw Materials
Technologies for 3G biomass processing in the bioindustry are at an initial stage in comparison to 1G and 2G biomass processing, which puts it in a disadvantage, mainly regarding the efficiency of farming, production, and transportation [51]. Nevertheless, macroalgae present a potential to have a role in future mitigation of climate change, either by the maintenance or reforestation of seaweed natural forests [52]. Seaweed partake in the global fixation and embedment of CO2 in ocean floors by farming it either on or offshore [52,53].
Cradle-to-grave LCA estimates for macroalgae have been conducted in the last few years, mainly for the production processes of biofuels such as methane and ethanol, with promising results. In 2012, Langlois et al. [54] concluded that even though, at the time, natural gas showed less impact than macroalgae biomethane, technological advancement and high production rates could make methane produced by seaweed biomass emit less CO2 than natural gas. Brockmann et al. [53] stressed in 2015 that although producing bioethanol from green macroalgae presents higher emissions than sugarcane ethanol, this is due mainly to logistics, infrastructure, and electricity consumption. The need to pump nutrients from the sea makes for high energy requirements. Even in the face of the present data, the authors believe that seaweed bioethanol presents long-lasting environmental benefits and could reach higher standards with time.
Seghetta et al. [55] showed how seaweed cultivation could lead to water restoration and reach negative emissions for the production of ethanol jointly with extraction of proteins and fertilizers. In a review by Duarte et al. [52], it was claimed that for every ton of dry seaweed used for biofuel production, 961 kg of carbon dioxide can be removed from the atmosphere. LCA tools were also used to assess the impact of production of polymeric packaging materials [56]. Rose [57] compiled LCA studies for various biofuels, ranking them for the quantity of carbon dioxide capture and permanence of its storage out of the atmosphere. In another compilation amassed by Pangestuti et al. [51] in 2021, 50% of biofuels LCA showed negative emissions, and some showed less emissions than 1G similar processes.
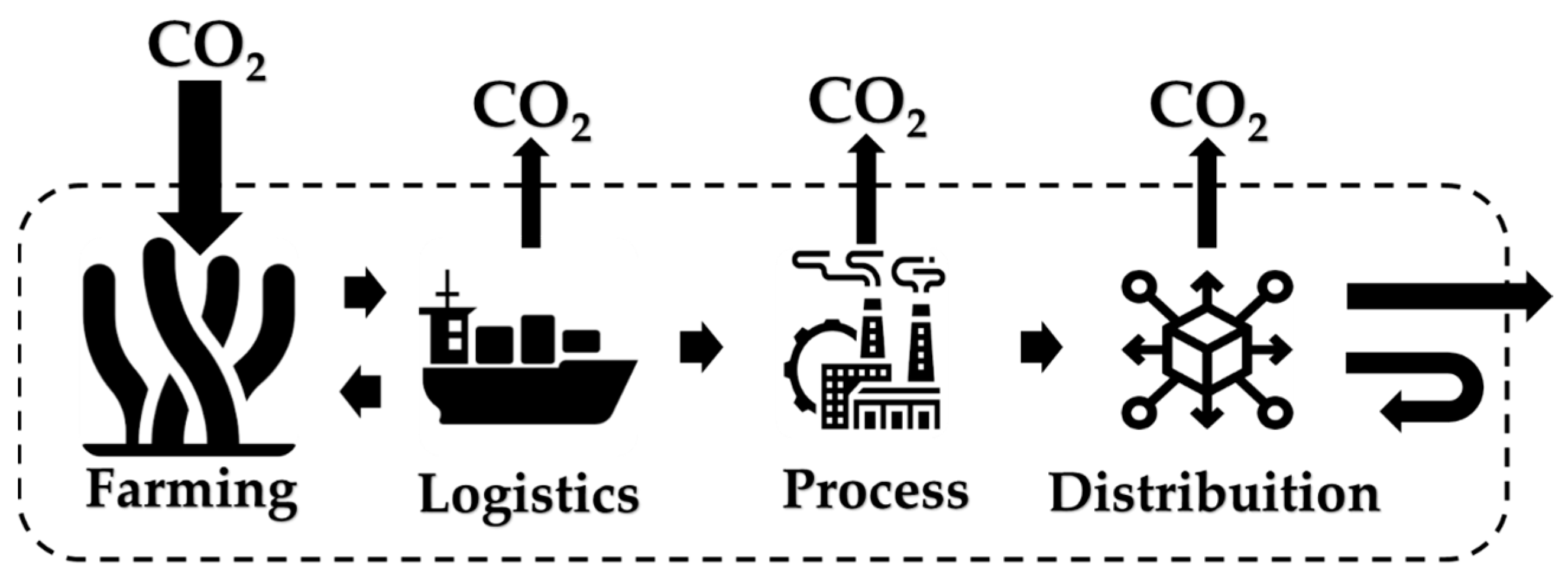
A comprehensive illustration for system boundaries in LCA for products obtained from macroalgae is presented in Figure 2, considering land use as negligible, but it should be advisable to take indirect land use into account for farming purposes.
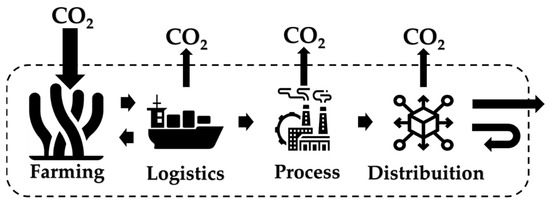
Figure 2.
Illustration of a life cycle assessment (LCA) of carbon dioxide for bioprocesses using algal biomass as raw material.
Macroalgae are composed mainly of gel-forming polysaccharides and cellulose from which a variety of fermentable monomers are obtainable. Green macroalgae monosaccharides include d-glucose, l-rhamnose, d-psicose, d-tagatose, and l-idronic acid. Brown seaweed contain l-fucose, d-mannitol, mannuronic acid, and glucuronic acid. Red seaweed contain galactose, 3,6-anydro-d-galactose, and agarobiose. Some pentoses such as l-arabinose, d-xylose, and d-ribose can also be found in macroalgae [58]. This high saccharide content makes fermentation processes with seaweed as raw material yield-effective. Fasahati el al. [59] demonstrated that using fermentation processes for ethanol production is the best alternative for energy production from seaweed, with physical processing (i.e., grinding and chopping) and transportation (i.e., wet or dry transportation and pumping) as main contributors to negative environmental impacts.
The carbohydrates of seaweeds make up, on average, 45% of their total dry mass. Due to their high mineral content, macroalgae normally contain a high concentration of calcium, magnesium, iron, and potassium in comparison to most terrestrial biomass, making them attractive for human consumption. The protein content is species-dependent, with significant quantities of amino acids and peptides, ranging from 3 to 15% in brown macroalgae (Phaeophyta), about 25% in green macroalgae (Chlorophyta), and reaching 40% in red macroalgae (Rhodophyta). Seaweeds are also known to contain valuable extractable pigments [12].
Traditionally farmed for consumption as part of the human diet, many seaweeds are a nutritive and healthy substance source that can be consumed in natura, or in the form of extracts, added to food products. In more recent times, seaweed extracts are used as gels and hydrocolloids in the cosmetic and health-care industries [14,60]. From the beginning of the 21st century, seaweed extract has been used as a bio-stimulant and fertilizer in terrestrial plantations, and as feed and compliment nutrition for poultry, cattle, and swine [12].
For their high sugar content, macroalgae are extensively studied as raw material for biofuels, mainly ethanol and butanol via fermentation processes. In addition to liquid fuels, which also include biodiesel from the low lipid content in macroalgae [61], gaseous fuels, such as methane and hydrogen, can be obtained from anaerobic digestion and dark fermentation. Biochar and bio-oil can be obtained through thermochemical transformation as well [12].
2.3. Kappaphycus alvarezii
Named by the biologist Vincente Alvarez, K. alvarezii, a red (Rhodophyta) seaweed, is considered as a dominant source of carrageenan [62]. K. alvarezii is fast-growing, able to duplicate its mass in less than a 30-day period [63]. It is a tropical species, viable strictly within 20° S and 20° N latitudes [64].
Like most seaweeds, K. alvarezii is composed mostly of saccharides (57.2%), in the form of carrageenan that contains galactose, and cellulose containing glucose. K. alvarezii also has a high quantity of minerals (15.8%), proteins (2.6%), and lipids (5.2%) [13]. The elkhorn-shaped sea moss can be divided into two essential parts: an exterior hydro-soluble gelatinous wall composed of carrageenan and a crystalline cellulosic interior. A macromolecular composition of K. alvarezii cultivated in Brazil as reported by Masarin et al. [65] is given in Table 1.

Table 1.
Macromolecular composition of Kappaphycus alvarezii cultivated in Brazil (adapted from Masarin et al. [65]).
The hydro-soluble fraction is made mostly of galactose, these concentrations can reach more than 60% of the plants’ exterior mass when adding up the quantities of galactose, 3,6-anydro-galactose, and 6-O-methylgalactose obtained from carrageenan which consists of a repetition of the two former monomers. The hydro-insoluble residue of K. alvarezii, made mainly of linear cellulose, has a 70% glucose content, 14% galactose, about 4% of mannose, and 1% of xylose [66].
In addition to the sugar content, carrageenan can have a high calcium cation content (140 mmol/100 g) [67] and proteins [68]. K. alvarezii has a complex and adaptive pigment content. A detailed pigment composition can be found in the review of Indriatmoko et al. [69].
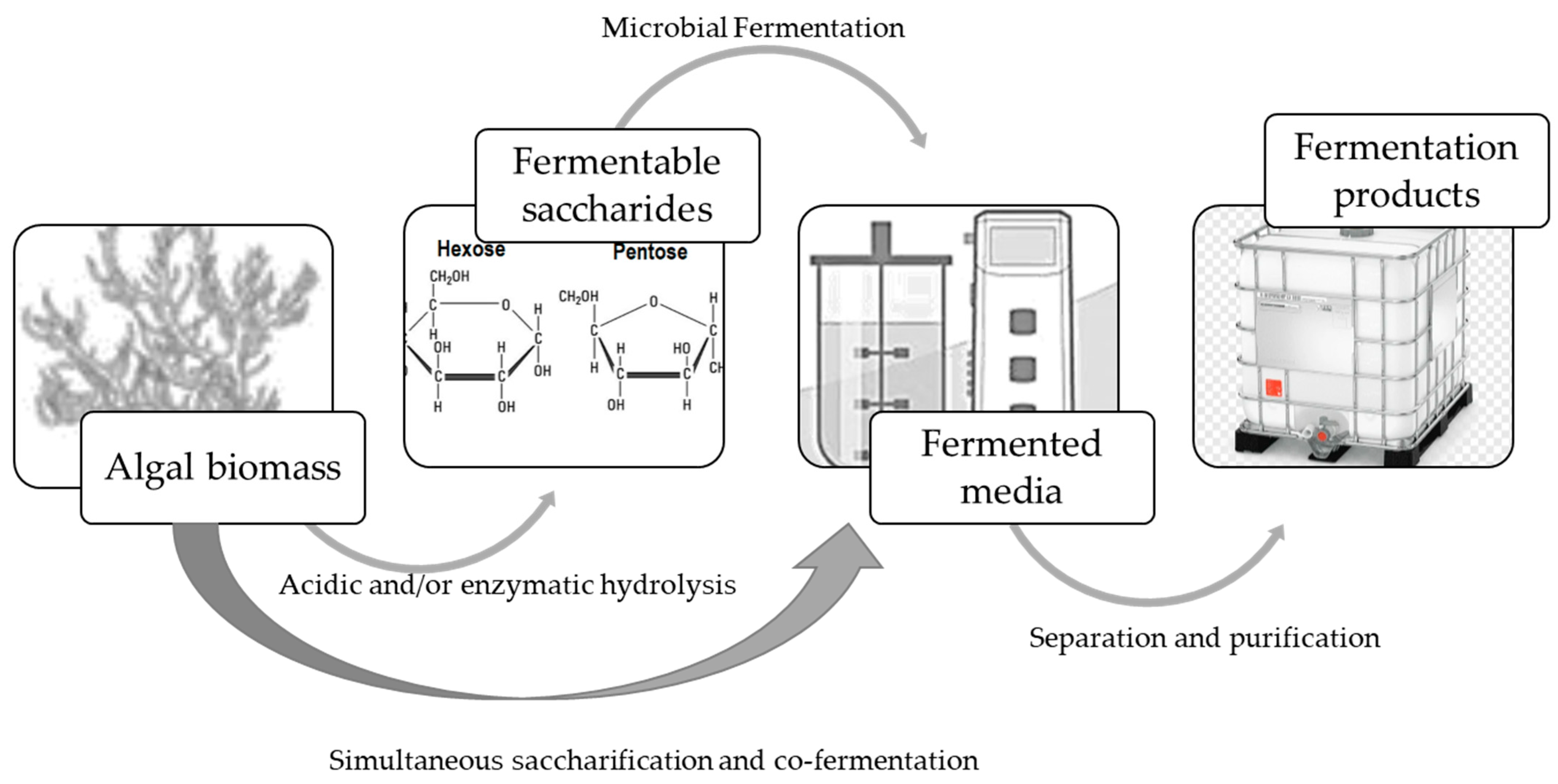
Fermentation processes, using seaweed biomass as raw material (Figure 3), involve three main steps. First, the polysaccharides are hydrolyzed to obtain the fermentable monomers, which are then fermented. Fermentation can also be carried out jointly with saccharification, e.g., simultaneous saccharification and co-fermentation (SSCF). The products are then separated from the fermented media, prepared, and packed for commercialization.
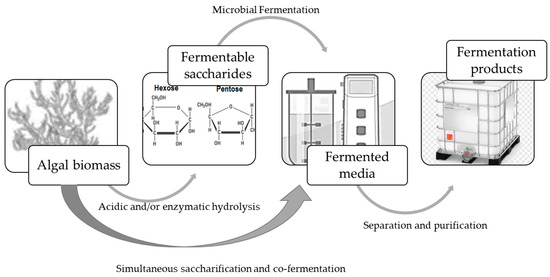
Figure 3.
Fermentation processes using algal biomass as raw material.
3. Processing K. alvarezii Biomass for Fermentation Processes
Sugars from K. alvarezii biomass can be obtained either by dilute-acid hydrolysis or enzymatic hydrolysis (Table 2). In most cases in the literature, carrageenan content is turned into galactose in a low concentration solution of either sulfuric or hydrochloric acid under temperatures ranging from 100 to 140 °C. Meinita et al. [70] and Ra et al. [71] reached concentrations of about 38.5 g/L of fermentable sugars from a 10–12% (w/v) biomass concentrated mix with a 0.2 molar concentration of sulfuric acid at 130–140 °C. These hydrolysis procedures in relatively higher temperatures and lower biomass concentration resulted in the formation of about 5 g/L of HMF. When using higher biomass concentrations in more prolonged hydrolysis procedures, higher galactose concentrations can be obtained. As shown by Khambhaty et al. [72] and Tabacof et al. [24], more than 50 g/L of galactose can be reached in a mixture with about a 30% (w/v) biomass concentration. The hydrolysis conditions in these cases were 100–110 °C and 1% (v/v) sulfuric acid concentration, and the hydrolysis time was 45 to 60 min long. Using similar conditions with an even higher biomass concentration of 50% (w/v) mix, Hargreaves et al. [22] reached a galactose concentration of 82 g/L. The work carried out by Hargreaves et al. [22] and Tabacof et al. [24,73] showed that exposing K. alvarezii biomass to prolonged acid hydrolysis results in the formation of 1 g/L of HMF for every 4 g/L of galactose.
Table 2.
K. alvarezii dilute-acid and enzymatic hydrolysis conditions and sugar products.
As can also be seen in Table 2, most enzymatic hydrolysis research has been carried out to investigate the exploitation of the crystalline linear cellulose content of K. alvarezii biomass using commercial enzyme cocktails such as Cellic CTec2 [21,22,65] and Celluclast® 1.5 L [74], obtaining more fermentable sugars, mostly glucose. Attempting to make most of the total K. alvarezii biomass, Abd-Rahim et al. [74] reported an increment of 15 g/L of reducing sugars to the 35 g/L obtained from dilute-acid hydrolysis reaching approximately 63% of hydrolysis efficiency from the combination of both methods in comparison to the 45% obtained from the acid hydrolysis alone. Hargreaves et al. [22] were able to simultaneously saccharify and ferment the cellulosic residue, as well as doing so while co-fermenting the galactose from a dilute-acid hydrolysate for ethanol production using Saccharomyces cerevisiae as the fermentation agent. The fermentation of the galactose from dilute-acid hydrolysis from 1000 g of dry seaweed resulted in 48.1 g of ethanol and the glucose from the 150 g of cellulosic residue being converted to an additional 43.7 g of ethanol, showing an almost double amount of product yield when combining chemical and enzymatic hydrolysis methods. Puspawati et al. [75] also successfully produced ethanol from the cellulosic residue.
3.1. Enabling Dilute-Acid Hydrolysate Fermentation
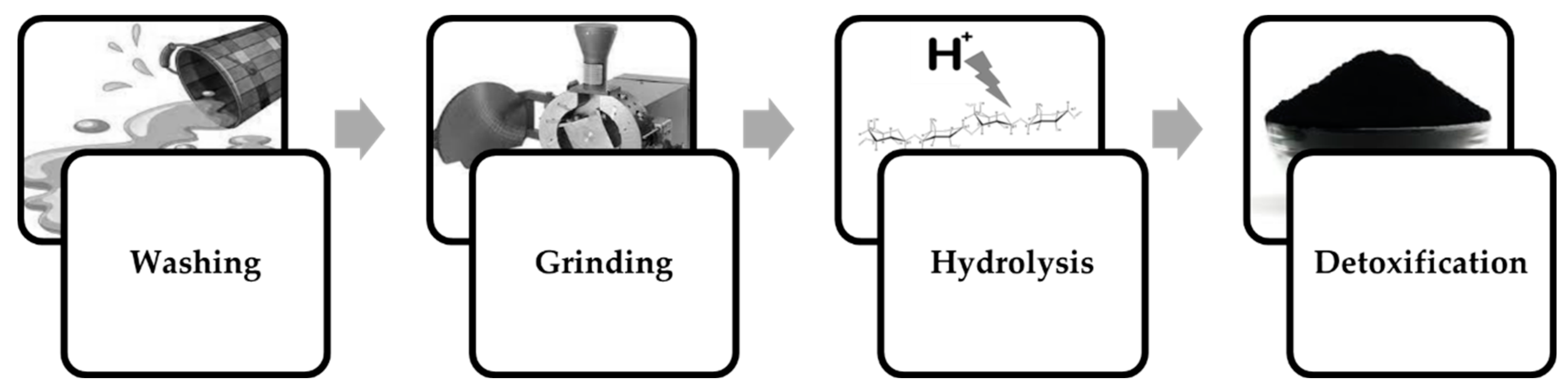
Processing seaweed for fermentation processes requires removal of elements that can potentially hinder cellular growth or inhibit the formation of the desired fermentation product. The macroalgae biomass must first be washed to reduce its salinity and remove debris that could have accumulated during cultivation and dehydration. The desalinated biomass is then grinded or finely chopped either at its wet state or, as more often reported, after being dehydrated once more [76]. Since furanic and furfural compounds can form as products of acid hydrolysis [26], in some cases the hydrolysate will need to be detoxified (Figure 4).
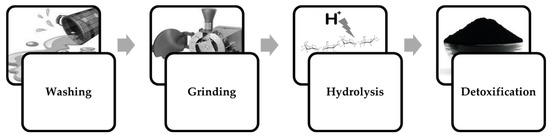
Figure 4.
Algal biomass pre-treatment for fermentation processes.
During dilute-acid hydrolysis in high temperatures, HMF is formed from the acid interaction with the carrageenan monomers, galactose and 3,6-anydro-α-l-galactose. As explained in the work of Oh et al. [77], high HMF quantities are formed mainly from the dehydration of the 3,6-anydro-α-l-galactose molecule via acid catalysis, and the HMF, in turn, can be rehydrated to levulinic and formic acid.
3.1.1. HMF Removal from K. alvarezii Hydrolysates
Removal of HMF from K. alvarezii hydrolysates can be performed via activated charcoal filtration, where HMF is captured within the charcoal’s pores [22,24,78]. Hargreaves et al. [22] determined that fine powdered charcoal is preferable to charcoal granules, and that approximately 1 g/L of charcoal should be used for each 1 g/L of HMF removed from the hydrolysate. Activated charcoal can also be regenerated via oxidation processes [79], desorption and decomposition methods [80], and thermal treatment [73,81]. Since HMF is a valuable chemical compound [29], its recuperation from the activated charcoal should be considered. An interesting technique is the removal of HMF from the charcoal pores using an organic solvent. Slak et al. [28] showed a similar process using carbon black that could be replicated for activated charcoal.
Another reported method for HMF removal is that of overliming, which consists of the addition of calcium oxide (CaO) [25] or calcium hydroxide (Ca(OH)2) [24,26,27]. A byproduct of HMF formation under acid-hydrothermal treatment are humins. Humin is a general term used for describing the bulk of insoluble organic compounds in soils [82]. Although the natural formation of humin in soil is not well defined in the literature, its formation mechanisms from HMF and carbohydrates have been proposed by various authors [83,84,85,86]. Humins are known to agglomerate and become insoluble under alkaline conditions [82,87]. Furthermore, aldol reactions, which are part of the mechanisms for humin polymerization, can be base catalyzed [88]. This way, the overliming alkaline conditions facilitate the condensation of HMF into the humin macromolecules, generating even larger humic substances, rendering them more insoluble.
Overliming methods vary. Xia et al. [89] reported that 50% of furans and HMF were removed when adjusting the pH of corn stalk hydrolysates to 10 at 60 °C for 2 h. Chi et al. [26] studied overliming processes for sugar mixtures obtained by pyrolysis. Preparing the solution for ethanol production by E. coli, 18.5 g/L of CA(OH)2 was added, and 8–16 h of treatment were carried out at 20 °C. The process was accelerated to a 1–4 h treatment period when temperatures were maintained at 60 °C. Mohgheghi et al. [27] used a shorter treatment period (30 min), showing that pH and temperature are key factors for efficient removal of HMF. In a recent study, K. alvarezii hydrolysates containing 24 g/L were reduced to 9 g/L using the addition of Ca(OH)2, elevating pH values to over 11 for 30 min [24].
Combining both methods, first applying overliming conditions and subsequently treating hydrolysates with powdered activated charcoal, can remove nearly all traces of HMF in K. alvarezii hydrolysates [24]. The charcoal pores absorb the soluble humins left over from the overliming treatment alongside the HMF that was not condensed into insoluble organic material.
The downside to using charcoal and calcium hydroxide is the material cost at large-scale production. Other efficient and cost-effective methods can also be proposed for HMF removal. such as nanofiltration and reverse osmosis [90]. It is worth mentioning that activated charcoal can be regenerated by chemical and thermal methods [73], and the calcium sulphate (gypsum) produced after neutralization of the overlimed solution can be put to use for construction purposes.
3.1.2. Cellular Acclimation to Hydrolysate
The HMF tolerance varies depending on the species of the microorganisms. Some microorganisms have extremely low HMF tolerance, and its presence can lead to a halt in cell growth, substrate consumption, and product formation [91,92]. Microorganisms are hardy and fast-evolving and can acquire tolerance to new, and potentially harmful, environments. Microbial cells can adapt to stress caused by harsh medium conditions or by differing carbon sources and nutrient availability. Short-term adaptation is more complex, including enhanced expression of certain genes. For instance, in yeast cells adapting to a galactose-rich environment, the GAL proteins were overexpressed, identifying change in expression of their transcription factors [93]. Long-term adaptation requires exposure to new mediums in which cells go through genetic mutations and obtain favorable phenotypes that help them better cope with their growth medium. A detailed explanation of adaptation can be found in a review by Tan et al. [94].
Yeast species, such as S. cerevisiae, can have a high HMF tolerance and, in some cases, convert up to 100% of furfural and HMF elements to less inhibitory components [95]. Adaptation to certain substrate can help to make more efficient use of certain metabolic pathways. Khambhaty et al. [72] and Hargreaves et al. [22] both used S. cerevisiae to produce bioethanol using K. alvarezii dilute-acid hydrolysates. The former did not report an adaptation process to a galactose-rich medium, while the latter acclimated the cells used for inoculum to galactose. After acclimation, the total consumption of galactose took place in less than 20 h compared to the 48 h fermentation period when the same species was not acclimated to galactose.
The presence of HMF can lead to metabolic shifts and different undesired fermentation products. Monlau et al. [96] reported that hydrogen-producing pathways during dark fermentation processes are turned to lactate, acetate, and ethanol pathways in the presence of HMF. An alternative treatment solution is biotransformation. HMF can be bio-transformed into 2,5-furandicarboxylic acid that can be used as a chemical building block [97].
Some bacteria, such as E. coli, are unviable in non-treated medium [26]. The Lactobacillus pentosus strain used in a study by Tabacof et al. [24] showed a threefold faster lag phase, two times the substrate consumption rate, and a higher product formation rate when exposed gradually to K. alvarezii hydrolysates. Due to the vast variety of fermentation agents, it is imperative to study the effect of fermentation media on the chosen microorganisms.
3.2. Enzymatic Hydrolysis Using Carrageenases-Type Enzymes
Enzymatic hydrolysis is on the forefront of algal biomass refining. Saccharification and extraction of intact compounds requires the use of enzymes capable of breaking down seaweed polysaccharides without degrading valuable organic material. Enzymes used for algal biomass are obtained mainly from marine microorganisms. By applying enzymes specific to the polymer chains of the soluble gelatinous polymers or the cellulose content of the algal biomass, valuable compounds such as pigments and proteins can be safely extracted from the biomass for commercial or industrial use [98]. Furthermore, biopolymers can be degraded to oligosaccharides and polyphenols with antioxidant, anticoagulant, anti-inflammatory, antiviral, and antitumoral properties [99].
The formation of oligomers via enzymatic hydrolysis has a high degree of homogeneity. Since enzymes function in relatively mild conditions, the degradation or undesired change in molecular structure of valuable compounds can be avoided. Moreover, obtaining oligomers from carrageenan through other methods has some disadvantages. Microwave degradation requires special equipment and operates in high pressures. Ozone depolymerization promotes an undesired chemical reaction, leading to carboxylic compounds. Ultrasound degradation is less efficient, resulting in non-homogenic oligomers. The use of gamma radiation to form oligomers is efficient, albeit the products could be toxic and need to be further investigated [100].
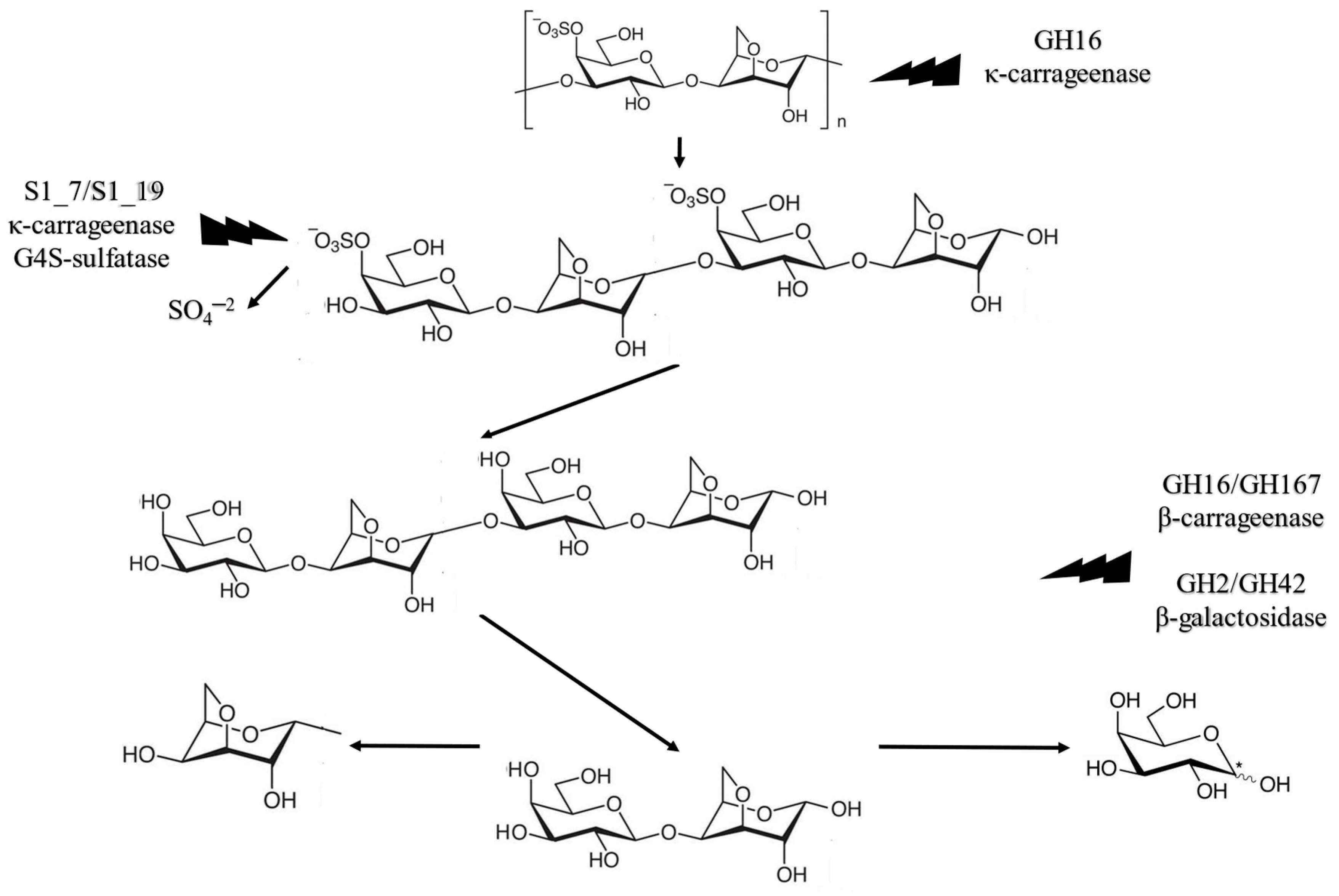
As observed in Table 2, up-to-date enzyme hydrolysis of K. alvarezii biomass focused on exploiting the seaweed’s cellulose for fermentation processes. Carrageenan contains a large amount of galactose that can be extracted via enzymes specific to its structure, the carrageenases. The study of marine vegetal biomass is at an early stage. Bäugman et al. [30] described in their review the state of the art for seaweed enzyme degradation of carrageenan, ulvan, agar, porphyran, and laminarian polysaccharide complexes. In K. alvarezii, µ-carrageenan turns into κ-carrageenan upon alkaline extraction. The enzymatic saccharification of κ-carrageenan includes four steps. First, the main chain is cleaved into four-monomer oligomers called neo-carrageenan by GH16 κ-carrageenase. These are then desulphurized to β-carrageenan by S1_7 and S1_19 κ-carrageenan-G4S-sulfatase. After desulphurization, β-carrageenan is further cleaved by GH16 and GH167 β-carrageenases into the basic carrageenan dimer, made of galactose and 3,6-anydro-d-galactose. Finally, d-galactose and 3,6-anydro-d-galactose are formed via the GH127 and GH129 3,6-anydro-d-galactosidase enzymes. Table 3 summarizes the steps for κ-carrageenan enzymatic saccharification, and Figure 5 illustrates the metabolic pathway for the enzymatic activity.
Table 3.
κ-carrageenan saccharification steps (summarized from Bäugman et al. [30]).
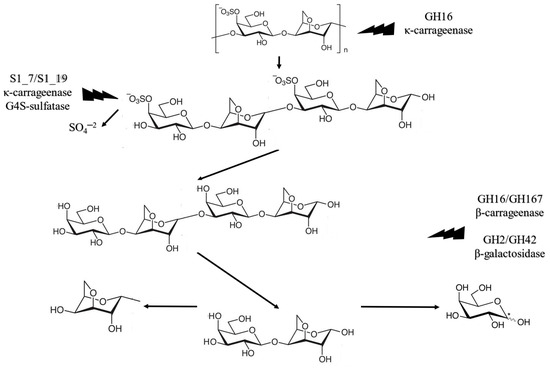
Figure 5.
Metabolic κ-carrageenan degradation pathway by carrageenases (Bäugman et al. [30]).
Carrageenan-degrading enzyme mixtures can be obtained from marine bacteria, the most cited being Pseudoalteromonas atlantica, Zobellia galactanivorans, and Pseudoalteromonas carrageenovora [30]. The marine microbiome is diverse and adapted to environments with extreme conditions. Extremophiles can produce enzymes that can function under high salinity, in alkaline or acidic media, and in extreme pressures and temperatures [101]. Such enzymes can favor parallel saccharification and fermentation processes. Pseudoalteromonas sp. and its variants are bacteria well adapted to extreme conditions as well as severely low temperatures. They are also cited as efficient degraders of galactose-rich polysaccharides other than carrageenan, e.g., agar, pectin, and alginate [102].
Aside from monomer formation for fermentation processes and oligomers for pharmaceutical and food additives, carrageenases can have other applications. Carrageenases can be added to detergents to degrade gums during cloth washing, or used in the textile industry, for their interaction with alginate and starch. Carrageenases can also be used to expose protoplast in algae cell walls, aiding genetic studies [103].
It stands clear that the efficient use of algal biomass as raw material within the biorefinery concept requires a combination of various extraction methods. The study of enzyme hydrolysis is key for reaching efficient extraction technologies that will lead to obtaining a large range of products alongside reducing sugars for fermentation processes.
4. The Fermentation of Rhodophyta-Type Saccharides
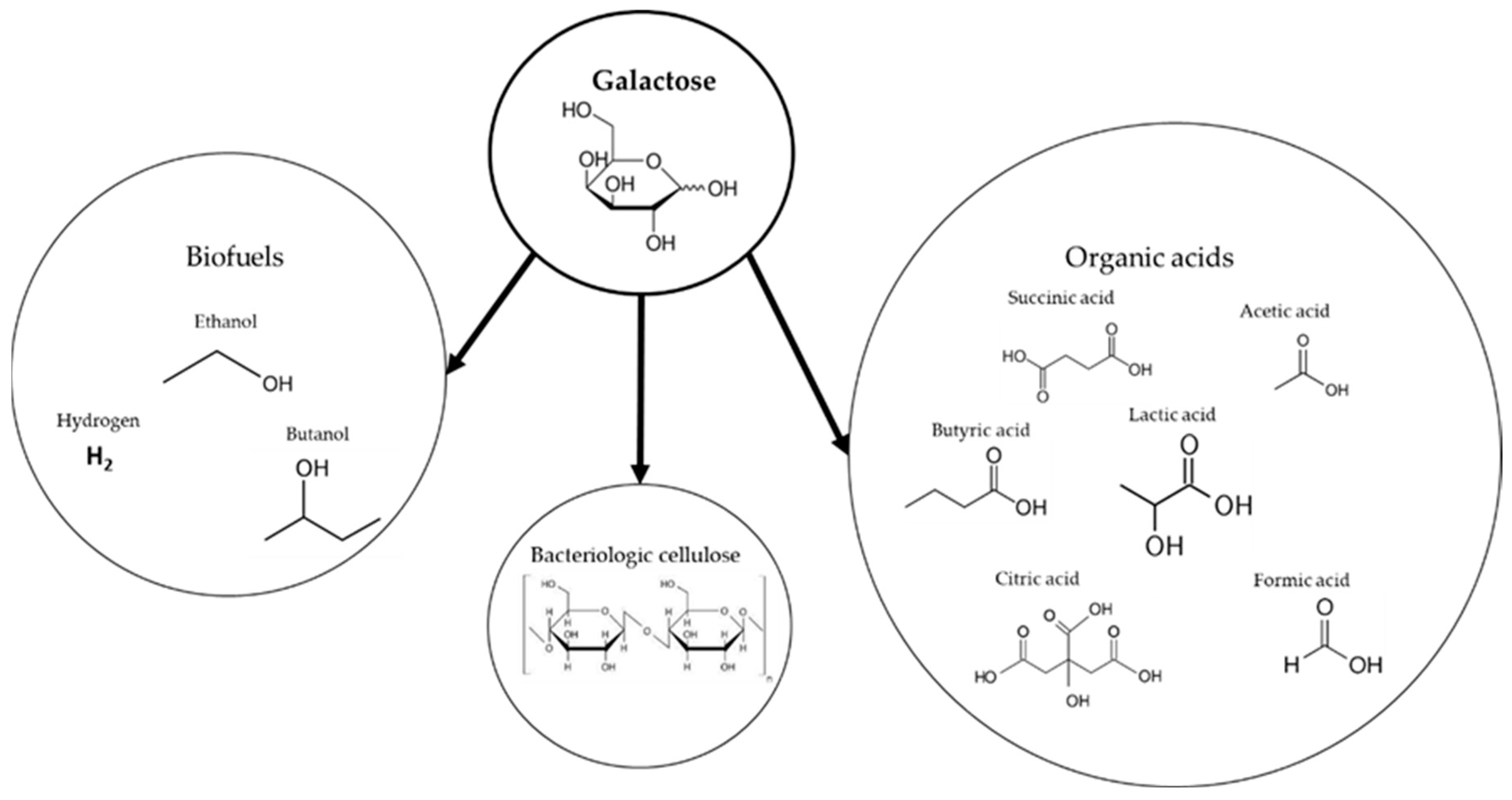
Rhodophyta algae are rich in galactose, a naturally occurring sugar which is processed by a wide range of microorganisms. Due to its abundance and potential industrial use, it has attracted academic interest [104] and is tested for various fermentation products (Figure 6).
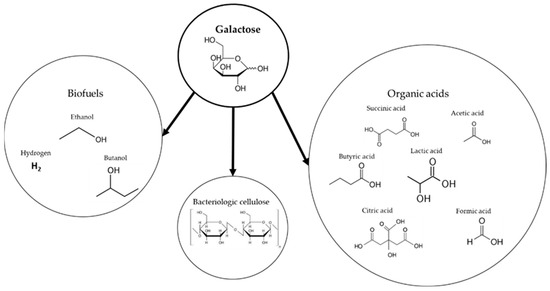
Figure 6.
Possible fermentation products from galactose metabolism reported in the literature.
4.1. Metabolic Pathways for Galactose Fermentation
d-galactose is metabolized through two types of pathways, the Tagatose-5-phosphate (TP) and the Leloir pathways (Table 4 and Table 5) [58]. The TP pathway involves galactose-specific, active transport by a phosphotransferase (Gal-PTS), the resulting galactose-6′phosphate is then turned into tagatose-6phosphate by lacAB, and further phosphorylated by lacC into tagatose-1,6-diphosphate. The enzyme lacD breaks tagatose-1,6-diphosphate into dihydroxyacetone phosphate and glyceraldyde-3-phosphate, which go through glycolysis.
Table 4.
Tagatose-6-phosphate pathway (summarized from Nagarajan et al. [58]).
Table 5.
Leloir pathway (summarized from Nagarajan et al. [58]).
In the Leloir pathway, d-galactose is transported passively through the cell membrane with a galactose permease galP. The intracellular molecule, β-galactose, gets phosphorylated by galM into galactose-1-phosphate. Next, the joint action of the enzymes galT, galE, and galU morph the galatose-1-phosphate into glucose-1-phosphate, which is turned into tagatose-1,6-diphosphate by pgmB and thereafter goes through glycolysis. A detailed explanation of both the TP and Leloir pathways can be found in the work of Nagarajan et al. [58].
4.2. Products from Galactose Fermentation
As can be seen in Figure 6, galactose has been shown to be fermented into a wide variety of products, including biofuels, organic acids, and bacterial cellulose. In order to assess the viability of each product, its specific production rate and yield from algal biomass sugars should be compared to those obtained with 1st- and 2nd-generation biomass. A summary of bioproducts from K. alvarezii or another galactose source is shown in Table 6.
Table 6.
Metabolites produced via fermentation of K. alvarezii biomass and other potential metabolites that can be obtained from galactose-rich media.
4.2.1. Biofuels
Among biofuels, ethanol has already been shown to be producible directly from galactose-rich macroalgae. Aside from K. alvarezii, Rhodophytae such as Gelidium amansii, Gracilaria sp., Chondrus crispus, and Palmaria palmata were also proved to be promising raw material for bioethanol production via fermentation [76]. For K. alvarezii hydrolysates, S. cerevisiae was used by a number of research groups [22,72]. The highest production was recorded by Hargreaves et al. [22], yielding 48.1 g of ethanol produced from galactose obtained via dilute-acid hydrolysates.
Hydrogen is a zero-carbon emission energy source producing only water when reacting with oxygen, which makes it one of the cleanest fuels of the future [116]. Recent research has also been conducted to evaluate the fermentation of galactose into hydrogen, envisioning the use of galactose-rich algal biomass. Heat treatments are used to separate heat-resistant spore-forming bacteria such as Clostridium butyricum [105] and Clostridium beijerinckii [106] that can successfully convert galactose into hydrogen via acetic and butyric acid pathways. In 2014, Park et al. [107] showed the predominance of the Clostridium cluster for hydrogen fermentation from galactose, reaching hydrogen from galactose yield of 1 mol/mol. Xia et al. [105] and Fonseca et al. [106] more than doubled this yield in 2016, with production rates of 33.6 mL/(g galactose·h) and 117.5 mL H2/L·h, respectively. One of the main hurdles in efficiency in hydrogen fermentation is preventing it from being consumed. This can be achieved by controlling the carbon dioxide concentration in fermentation media. Moreover, genetic and metabolic methods can be used to enhance hydrogen production and inactivate its intake by microorganisms [105].
Butanol is another molecule that can be used as a fuel, although its main use is as a solvent and chemical intermediate. Butanol-producing bacterium, such as C. beijerinckii, are able to intake galactose and produce butanol [108]. Better expression of galactose metabolic pathways is needed to accelerate galactose catabolism, making it equivalent to the processing speeds of cellobiose and glucose in microorganisms.
4.2.2. Organic Acids
Organic acids have extensive use as additives in food, cosmetic products, and solvents. Some organic acids are used as chemical building block platforms for the production of added-value products and biodegradable polymers.
The growth of the poly-lactic acid market has led to the search of new raw materials for lactic acid fermentation. Different seaweed species are already being studied as raw material for lactic acid fermentation, obtaining fermentable sugars principally through dilute-acid hydrolysis. Lactic acid has already been successfully obtained from galactose-rich algae such as Gelidium amansii, Euchema denticulatum, and Gracilaria sp., with final concentrations reaching 30 g/L and yields of lactic acid from reducing sugars around 0.9 g/g [58]. Microorganisms used for lactic acid production from galactose include Lactobacillus strains, L. rhamnosus, L. acidophilis, and L. plantarum, as well as other strains such as Weissella paramesenteroides and Bacillus coagulans. In 2022, Sudhakar and Dharani [110] demonstrated that K. alvarezii biomass can be processed into lactic acid by L. plantarum. In the work of Tabacof et al. [73] in 2023, it was shown that 115 kg of lactic acid can be produced by 1 ton of dry weight K. alvarezii using L. pentosus.
Succinic acid is also a valuable organic acid that can be used to produce poly-butyl-succinate among other added-value products. Actinobacillus succinogenes is a popular choice for studying succinic acid production, and this bacterium can utilize galactose as a substrate [117,118]. Anaerobiospirillum succinoproducenes was also proven to be able to utilize galactose alongside glucose and therefore be an interesting bacterium for red algal biomass hydrolysate fermentation [119].
Algal biomass is a potential feedstock for succinic acid. Laminaria digitata [120] and Saccharina latissimi [121] hydrolysates were fermented by A. succinogenes. As for galactose-producing macroalgae, Palmaria palmata hydrolysates containing glucose and galactose were fermented by a genetically modified strain of E. coli by Olajuyin et al. [109], reaching a 22.4 g/L volume of succinic acid solution with a 1.13 mol/mol yield.
Other widely commercialized organic acids have been produced from galactose fermentation. The vinegar acid, acetic acid, has a wide range of applications and is also a precursor of valuable products such as vinyl acetate. Ehsanipour et al. [113] used the acetogenic bacteria, Moorella thermoacetica, to ferment lignocellulosic sugars, which include galactose. In this case, the galactose was consumed in a slower rate than other reducing sugars and was detected in the final fermented media in low quantities. The authors stressed that recombinant strains should be developed to make galactose intake more efficient.
Citric acid is used as a food additive for preservation and flavoring. The fungi Aspergillus niger can produce citric acid from galactose in whey protein [111]. Citric acid-producing yeast can also be used to convert galactose to citric acid. Lazar et al. [112] demonstrated that the commonly used yeast Yarrowia lipolytica can express all the Leloir pathway genes and efficiently uptake galactose for citric acid production. The yeast, Candida guillermondii, was shown to produce citric acid from galactose as efficiently as from glucose when each carbohydrate was used as a sole substrate [122]. Although in both cases Y. lipolytica and C. gillermondii showed a strong preference to glucose, uptake of galactose shows the potential of applying these microorganisms to ferment galactose-rich media like K. alvarezii hydrolysates.
Spent coffee hydrolysates contain galactose that can be used as substrate for bioproduction via fermentation. He et al. [114] utilized the fermenting agent Clostridium tyrobutyricum to demonstrate production of butyric acid in galactose-rich media, indicating the potential of the production of this valuable organic acid that is used to produce butyrate esters and cellulose acetate butyrate.
4.2.3. Bacterial Cellulose
Cellulose is an abundant natural resource from plant tissue, and it is traditionally used for the manufacture of paper and fibers. Bio-cellulose derived from cellulose-excreting bacteria has a higher purity and crystallinity, which leads to an elevated degree of polymerization, an increase in water-holding capacity, and a superior permeability to oxygen [123].
Gluconacetobacter xylinus has been shown to be able to process galactose for bio-cellulose production [115,123,124]. Al-abdalla and Dahman [115] used wheat straw hydrolysates for bio-cellulose production, showing close to 72% of galactose consumption. Dahman et al. [123] reported that consumption of galactose reached 90% when it was used as substrate for bio-cellulose production with a 6.79 g/g yield. Although galactose in some cases has been shown to be consumed less efficiently than other substrate [124], a resolution could be found in genetic engineering and acclimation methods for activating and overexpressing the Leloir or Tagatose-6-Phosphate pathways.
5. K. alvarezii Fermentation Biorefinery Scheme
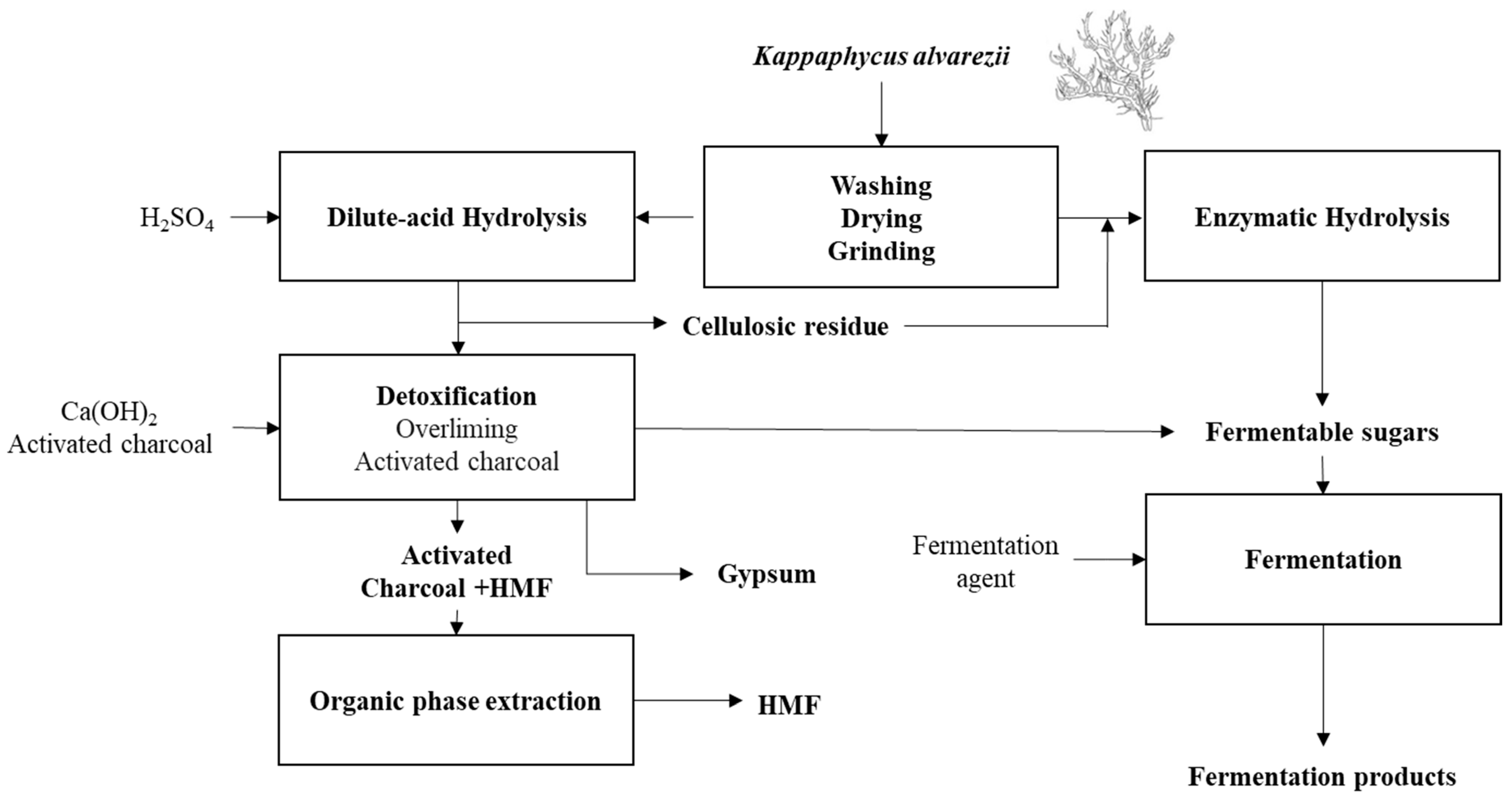
A generic process flow diagram for a K. alvarezii fermentation biorefinery is shown in Figure 7. Given the current state of K. alvarezii processing, it is found that the reducing sugars used for fermentation processes can be obtained either by dilute-acid hydrolysis, preferably with sulfuric acid or by enzymatic hydrolysis with the use of carrageenases for processing the carrageenan, and cellulases for obtaining glucose from the algal cellulose component. The cellulosic residue from the dilute-acid hydrolysis can also be enzymatically processed to obtain more fermentable sugars. As discussed earlier, fermentation products can include bio-cellulose, biofuels such as hydrogen, ethanol, and butanol, and organic acids, i.e., lactate, succinate, citrate, butyrate, and acetate.
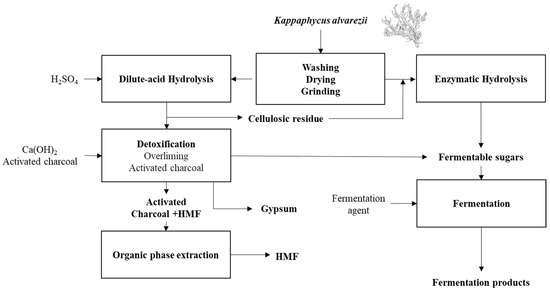
Figure 7.
Kappaphycus alvarezii fermentation biorefinery scheme with HMF and gypsum as subproducts integrating dilute-acid and enzymatic hydrolysis.
For dilute-acid hydrolysis, the overliming and activated charcoal methods were shown to be efficient in HMF removal for enabling fermentation. Gypsum can be obtained after the neutralization of the sulfuric acid with calcium hydroxide, and HMF can be extracted from charcoal via organic phase extraction, rendering another valuable refinery product.
Red seaweed is more cultivated than other types of seaweed, representing more than half of the macroalgae cultivated worldwide [13]. Furthermore, these macroalgae do not compete as much with food industries, as they are mainly used for their extraction of their industrial gums (agar and carrageenan). Some species are cultivated more than others in different regions or countries. K. alvarezii is mostly cultivated in Indonesia, the Philippines, and Malaysa [17]. In Brazil, production of K. alvarezii is on the rise as the market for carrageenan amplifies [31]. Development of fermentation processes for K. alvarezii in countries where it is efficiently cultivated on a large scale are relevant because they can create market opportunities never before explored.
In general, fermentation processes broaden the array of products a biorefinery can manufacture. A feasible economic strategy is to integrate fermentation processes for K. alvarezii in existing refineries for other products, using biomass surpluses (if available) for fermentation processes. As enzymatic saccharification technology matures, the extraction of high-value compounds such as natural pigments from K. alvarezii, in addition to fermentable sugars, will be possible. These product combinations with valuable merchandise can make fermentation processes more attractive for industrial investment.
6. Concluding Remarks
Fermentation processes are used to obtain a wide array of products and their use is highly consolidated, and most products are industrially manufactured. In this review, a current state of the art of processing the macroalgae K. alvarezii was discussed. Ethanol and lactic acid have already been produced from this widely cultivated seaweed, and more products that can be potentially derived from K. alvarezii were presented. The main challenges for efficient processing of the biomass and making the sugar-rich medium more fermentable, detoxification and microorganism acclimation were also detailed.
K. alvarezii is one of the most widely cultivated types of seaweed worldwide; its sugar content makes it a promising feedstock for the production of biofuels, organic compounds, and highly valuable chemical building blocks via fermentation. The use of third-generation biomass can become an important milestone in the roadmap for sustainable industrial processes. Thus, it is of great importance to bring to light all the possibilities for further research and elaboration of processes for obtaining bioproducts from this biomass.
Author Contributions
Conceptualization, A.T. and N.P.J.; Data curation, A.T.; Formal analysis, A.T. and N.P.J.; Funding acquisition, N.P.J.; Investigation, A.T.; Methodology, A.T. and N.P.J.; Project administration, N.P.J.; Resources, N.P.J.; Supervision, V.C. and N.P.J.; Writing—original draft, A.T.; Writing—review and editing, N.P.J. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Shi, S.; Yin, J. Global Research on Carbon Footprint: A Scientometric Review. Environ. Impact Assess. Rev. 2021, 89, 106571. [Google Scholar] [CrossRef]
- Yaacob, N.F.F.; Mat Yazid, M.R.; Abdul Maulud, K.N.; Ahmad Basri, N.E. A Review of the Measurement Method, Analysis and Implementation Policy of Carbon Dioxide Emission from Transportation. Sustainability 2020, 12, 5873. [Google Scholar] [CrossRef]
- Lackner, M. 3rd-Generation Biofuels: Bacteria and Algae as Sustainable Producers and Converters. In Handbook of Climate Change Mitigation and Adaptation; Springer: New York, NY, USA, 2015; pp. 1–32. [Google Scholar]
- Paraschiv, S.; Paraschiv, L.S. Trends of Carbon Dioxide (CO2) Emissions from Fossil Fuels Combustion (Coal, Gas and Oil) in the EU Member States from 1960 to 2018. Energy Rep. 2020, 6, 237–242. [Google Scholar] [CrossRef]
- Liu, Z.; Deng, Z.; Davis, S.; Ciais, P. Monitoring Global Carbon Emissions in 2022. Nat. Rev. Earth Environ. 2023, 4, 205–206. [Google Scholar] [CrossRef] [PubMed]
- ElHady, S.; Amin, O.; Elhussieny, A.; Fahim, I.S. Bioplastics, Biodegradable Plastics, and Degradation in Natural Environments. In Biodegradability of Conventional Plastics; Elsevier: Amsterdam, The Netherlands, 2023; pp. 47–67. [Google Scholar]
- Tukker, A. Life Cycle Assessment as a Tool in Environmental Impact Assessment. Environ. Impact Assess. Rev. 2000, 20, 435–456. [Google Scholar] [CrossRef]
- Terlouw, T.; Bauer, C.; Rosa, L.; Mazzotti, M. Life Cycle Assessment of Carbon Dioxide Removal Technologies: A Critical Review. Energy Environ. Sci. 2021, 14, 1701–1721. [Google Scholar] [CrossRef]
- Fiorese, G.; Catenacci, M.; Verdolini, E.; Bosetti, V. Advanced Biofuels: Future Perspectives from an Expert Elicitation Survey. Energy Policy 2013, 56, 293–311. [Google Scholar] [CrossRef]
- Ziolkowska, J.; Simon, L.; Zilberman, D. Capturing Uncertainties in Evaluation of Biofuels Feedstocks: A Multi-Criteria Approach for the US; ETH Zurich: Zurich, Switzerland, 2011. [Google Scholar]
- Buschmann, A.H.; Camus, C.; Infante, J.; Neori, A.; Israel, Á.; Hernández-González, M.C.; Pereda, S.V.; Gomez-Pinchetti, J.L.; Golberg, A.; Tadmor-Shalev, N.; et al. Seaweed Production: Overview of the Global State of Exploitation, Farming and Emerging Research Activity. Eur. J. Phycol. 2017, 52, 391–406. [Google Scholar] [CrossRef]
- Torres, M.D.; Kraan, S.; Domínguez, H. Seaweed Biorefinery. Rev. Environ. Sci. Biotechnol. 2019, 18, 335–388. [Google Scholar] [CrossRef]
- Álvarez-Viñas, M.; Flórez-Fernández, N.; Torres, M.D.; Domínguez, H. Successful Approaches for a Red Seaweed Biorefinery. Mar. Drugs 2019, 17, 620. [Google Scholar] [CrossRef]
- Lange, L.; Bak, U.G.; Hansen, S.C.B.; Gregersen, O.; Harmsen, P.; Karlsson, E.N.; Meyer, A.; Mikkelsen, M.D.; Van Den Broek, L.; Hreggviðsson, G.Ó. Opportunities for Seaweed Biorefinery. In Sustainable Seaweed Technologies: Cultivation, Biorefinery, and Applications; Elsevier: Amsterdam, The Netherlands, 2020; pp. 3–31. ISBN 9780128179437. [Google Scholar]
- Sudhakar, K.; Mamat, R.; Samykano, M.; Azmi, W.H.; Ishak, W.F.W.; Yusaf, T. An Overview of Marine Macroalgae as Bioresource. Renew. Sustain. Energy Rev. 2018, 91, 165–179. [Google Scholar] [CrossRef]
- Hung, L.D.; Hori, K.; Nang, H.Q.; Kha, T.; Hoa, L.T. Seasonal Changes in Growth Rate, Carrageenan Yield and Lectin Content in the Red Alga Kappaphycus alvarezii Cultivated in Camranh Bay, Vietnam. J. Appl. Phycol. 2009, 21, 265–272. [Google Scholar] [CrossRef]
- Campbell, R.; Hotchkiss, S. Carrageenan Industry Market Overview. In Tropical Seaweed Farming Trends, Problems and Opportunities; Springer International Publishing: Cham, Switzerland, 2017; pp. 193–205. [Google Scholar]
- Chandel, A.K.; Albarelli, J.Q.; Santos, D.T.; Chundawat, S.P.; Puri, M.; Meireles, M.A.A. Comparative Analysis of Key Technologies for Cellulosic Ethanol Production from Brazilian Sugarcane Bagasse at a Commercial Scale. Biofuels Bioprod. Biorefining 2019, 13, 994–1014. [Google Scholar] [CrossRef]
- Nogueira, M.C.F.; Henriques, M.B. Large-Scale versus Family-Sized System Production: Economic Feasibility of Cultivating Kappaphycus alvarezii along the Southeastern Coast of Brazil. J. Appl. Phycol. 2020, 32, 1893–1905. [Google Scholar] [CrossRef]
- Nass, L.L.; Pereira, P.A.A.; Ellis, D. Biofuels in Brazil: An Overview. Crop Sci. 2007, 47, 2228–2237. [Google Scholar] [CrossRef]
- Paz-Cedeno, F.R.; Solórzano-Chávez, E.G.; de Oliveira, L.E.; Gelli, V.C.; Monti, R.; de Oliveira, S.C.; Masarin, F. Sequential Enzymatic and Mild-Acid Hydrolysis of By-Product of Carrageenan Process from Kappaphycus alvarezii. Bioenergy Res. 2019, 12, 419–432. [Google Scholar] [CrossRef]
- Hargreaves, P.I.; Barcelos, C.A.; da Costa, A.C.A.; Pereira, N. Production of Ethanol 3G from Kappaphycus alvarezii: Evaluation of Different Process Strategies. Bioresour. Technol. 2013, 134, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Wang, K.; Chen, Y.; Tan, T.; Nielsen, J. Third-Generation Biorefineries as the Means to Produce Fuels and Chemicals from CO2. Nat. Catal. 2020, 3, 274–288. [Google Scholar] [CrossRef]
- Tabacof, A.; Calado, V.; Pereira, N. Third Generation Lactic Acid Production by Lactobacillus pentosus from the Macroalgae Kappaphycus alvarezii Hydrolysates. Fermentation 2023, 9, 319. [Google Scholar] [CrossRef]
- Baig, M.Z.; Dharmadhikari, S.M. Optimization of Detoxification with Over Liming and Charcoal Treatment for Increasing the Fermentability of Cotton Stalk Hydrolyzate. Indian. J. Appl. Res. 2014, 5, 453–455. [Google Scholar] [CrossRef]
- Chi, Z.; Rover, M.; Jun, E.; Deaton, M.; Johnston, P.; Brown, R.C.; Wen, Z.; Jarboe, L.R. Overliming Detoxification of Pyrolytic Sugar Syrup for Direct Fermentation of Levoglucosan to Ethanol. Bioresour. Technol. 2013, 150, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Mohagheghi, A.; Ruth, M.; Schell, D.J. Conditioning Hemicellulose Hydrolysates for Fermentation: Effects of Overliming PH on Sugar and Ethanol Yields. Process Biochem. 2006, 41, 1806–1811. [Google Scholar] [CrossRef]
- Slak, J.; Pomeroy, B.; Kostyniuk, A.; Grilc, M.; Likozar, B. A Review of Bio-Refining Process Intensification in Catalytic Conversion Reactions, Separations and Purifications of Hydroxymethylfurfural (HMF) and Furfural. Chem. Eng. J. 2022, 429, 132325. [Google Scholar] [CrossRef]
- Rosatella, A.A.; Simeonov, S.P.; Frade, R.F.M.; Afonso, C.A.M. 5-Hydroxymethylfurfural (HMF) as a Building Block Platform: Biological Properties, Synthesis and Synthetic Applications. Green Chem. 2011, 13, 754–793. [Google Scholar] [CrossRef]
- Bäumgen, M.; Dutschei, T.; Bornscheuer, U.T. Marine Polysaccharides: Occurrence, Enzymatic Degradation and Utilization. ChemBioChem 2021, 22, 2247–2256. [Google Scholar] [CrossRef] [PubMed]
- Rudke, A.R.; de Andrade, C.J.; Ferreira, S.R.S. Kappaphycus Alvarezii Macroalgae: An Unexplored and Valuable Biomass for Green Biorefinery Conversion. Trends Food Sci. Technol. 2020, 103, 214–224. [Google Scholar] [CrossRef]
- Louw, J.; Dogbe, E.S.; Yang, B.; Görgens, J.F. Prioritisation of Biomass-Derived Products for Biorefineries Based on Economic Feasibility: A Review on the Comparability of Techno-Economic Assessment Results. Renew. Sustain. Energy Rev. 2023, 188, 113840. [Google Scholar] [CrossRef]
- Bhatia, L.; Bachheti, R.K.; Garlapati, V.K.; Chandel, A.K. Third-Generation Biorefineries: A Sustainable Platform for Food, Clean Energy, and Nutraceuticals Production. Biomass Convers. Biorefinery 2022, 12, 4215–4230. [Google Scholar] [CrossRef]
- Ahorsu, R.; Medina, F.; Constantí, M. Significance and Challenges of Biomass as a Suitable Feedstock for Bioenergy and Biochemical Production: A Review. Energies 2018, 11, 3366. [Google Scholar] [CrossRef]
- Dutta, K.; Daverey, A.; Lin, J.-G. Evolution Retrospective for Alternative Fuels: First to Fourth Generation. Renew. Energy 2014, 69, 114–122. [Google Scholar] [CrossRef]
- Lü, J.; Sheahan, C.; Fu, P. Metabolic Engineering of Algae for Fourth Generation Biofuels Production. Energy Environ. Sci. 2011, 4, 2451. [Google Scholar] [CrossRef]
- McKendry, P. Energy Production from Biomass (Part 1): Overview of Biomass. Bioresour. Technol. 2002, 83, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Maurya, P.K.; Mondal, S.; Kumar, V.; Singh, S.P. Roadmap to Sustainable Carbon-Neutral Energy and Environment: Can We Cross the Barrier of Biomass Productivity? Environ. Sci. Pollut. Res. 2021, 28, 49327–49342. [Google Scholar] [CrossRef] [PubMed]
- Barboza-Rodríguez, R.; Rodríguez-Jasso, R.M.; Rosero-Chasoy, G.; Rosales Aguado, M.L.; Ruiz, H.A. Photobioreactor Configurations in Cultivating Microalgae Biomass for Biorefinery. Bioresour. Technol. 2024, 394, 130208. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Mukhambet, Y.; Esakkimuthu, S.; Abomohra, A.E.-F. Integrated Microalgal Biorefinery—Routes, Energy, Economic and Environmental Perspectives. J. Clean. Prod. 2022, 348, 131245. [Google Scholar] [CrossRef]
- Dahman, Y.; Syed, K.; Begum, S.; Roy, P.; Mohtasebi, B. Biofuels. In Biomass, Biopolymer-Based Materials, and Bioenergy; Elsevier: Amsterdam, The Netherlands, 2019; pp. 277–325. [Google Scholar]
- Ho, D.P.; Ngo, H.H.; Guo, W. A Mini Review on Renewable Sources for Biofuel. Bioresour. Technol. 2014, 169, 742–749. [Google Scholar] [CrossRef] [PubMed]
- Bentivoglio, D.; Finco, A.; Bacchi, M. Interdependencies between Biofuel, Fuel and Food Prices: The Case of the Brazilian Ethanol Market. Energies 2016, 9, 464. [Google Scholar] [CrossRef]
- Bonomi, A.; Cavalett, O.; da Cunha, M.P.; Lima, M.A.P. The Virtual Sugarcane Biorefinery Concept. In Virtual Biorefinery: An Optimization Strategy for Renewable Carbon Valorization; Springer: Cham, Switzerland, 2016; pp. 5–11. [Google Scholar]
- Soares, G.A. O Avanço Das Tecnologias de Segunda Geração e Seus Impactos Na Indústria Do Etanol. Master’s Thesis, Universidade Federal do Rio de Janeiro, Rio de Janeiro, Brazil, 2016. [Google Scholar]
- Harper, A.B.; Powell, T.; Cox, P.M.; House, J.; Huntingford, C.; Lenton, T.M.; Sitch, S.; Burke, E.; Chadburn, S.E.; Collins, W.J.; et al. Land-Use Emissions Play a Critical Role in Land-Based Mitigation for Paris Climate Targets. Nat. Commun. 2018, 9, 2938. [Google Scholar] [CrossRef]
- Gibbs, H.K.; Johnston, M.; Foley, J.A.; Holloway, T.; Monfreda, C.; Ramankutty, N.; Zaks, D. Carbon Payback Times for Crop-Based Biofuel Expansion in the Tropics: The Effects of Changing Yield and Technology. Environ. Res. Lett. 2008, 3, 034001. [Google Scholar] [CrossRef]
- Spera, S.A.; Galford, G.L.; Coe, M.T.; Macedo, M.N.; Mustard, J.F. Land-use Change Affects Water Recycling in Brazil’s Last Agricultural Frontier. Glob. Chang. Biol. 2016, 22, 3405–3413. [Google Scholar] [CrossRef]
- Plevin, R.J.; Jones, A.D.; Torn, M.S.; Gibbs, H.K. Greenhouse Gas Emissions from Biofuels’ Indirect Land Use Change Are Uncertain but May Be Much Greater than Previously Estimated. Environ. Sci. Technol. 2010, 44, 8015–8021. [Google Scholar] [CrossRef] [PubMed]
- Tanzer, S.E.; Ramírez, A. When Are Negative Emissions Negative Emissions? Energy Environ. Sci. 2019, 12, 1210–1218. [Google Scholar] [CrossRef]
- Pangestuti, M.B.; Suhartini, S.; Hidayat, N. Life Cycle Assessment of Bioenergy Production from Macroalgae: A Review. IOP Conf. Ser. Earth Environ. Sci. 2021, 924, 012070. [Google Scholar] [CrossRef]
- Duarte, C.M.; Wu, J.; Xiao, X.; Bruhn, A.; Krause-Jensen, D. Can Seaweed Farming Play a Role in Climate Change Mitigation and Adaptation? Front. Mar. Sci. 2017, 4, 100. [Google Scholar] [CrossRef]
- Brockmann, D.; Pradinaud, C.; Champenois, J.; Benoit, M.; Hélias, A. Environmental Assessment of Bioethanol from Onshore Grown Green Seaweed. Biofuels Bioprod. Biorefining 2015, 9, 696–708. [Google Scholar] [CrossRef]
- Langlois, J.; Sassi, J.; Jard, G.; Steyer, J.; Delgenes, J.; Hélias, A. Life Cycle Assessment of Biomethane from Offshore-cultivated Seaweed. Biofuels Bioprod. Biorefining 2012, 6, 387–404. [Google Scholar] [CrossRef]
- Seghetta, M.; Hou, X.; Bastianoni, S.; Bjerre, A.-B.; Thomsen, M. Life Cycle Assessment of Macroalgal Biorefinery for the Production of Ethanol, Proteins and Fertilizers—A Step towards a Regenerative Bioeconomy. J. Clean. Prod. 2016, 137, 1158–1169. [Google Scholar] [CrossRef]
- Amponsah, L.; Chuck, C.; Parsons, S. Life Cycle Assessment of a Marine Biorefinery Producing Protein, Bioactives and Polymeric Packaging Material. Int. J. Life Cycle Assess. 2023, 29, 174–191. [Google Scholar] [CrossRef]
- Rose, D. Life Cycle of Carbon in Macroalgae for Various Products; U.S. Department of Energy: Richland, WA, USA, 2021.
- Nagarajan, D.; Chen, C.-Y.; Ariyadasa, T.U.; Lee, D.-J.; Chang, J.-S. Macroalgal Biomass as a Potential Resource for Lactic Acid Fermentation. Chemosphere 2022, 309, 136694. [Google Scholar] [CrossRef]
- Fasahati, P.; Dickson, R.; Saffron, C.M.; Woo, H.C.; Liu, J.J. Seaweeds as a Sustainable Source of Bioenergy: Techno-Economic and Life Cycle Analyses of Its Biochemical Conversion Pathways. Renew. Sustain. Energy Rev. 2022, 157, 112011. [Google Scholar] [CrossRef]
- Leandro, A.; Pereira, L.; Gonçalves, A.M.M. Diverse Applications of Marine Macroalgae. Mar. Drugs 2019, 18, 17. [Google Scholar] [CrossRef] [PubMed]
- El-Sheek, M.; Abomhohra, A.E.-F. Handbook of Algal Biofuels; Elsevier: Amsterdam, The Netherlands, 2022. [Google Scholar]
- Rupert, R.; Rodrigues, K.F.; Thien, V.Y.; Yong, W.T.L. Carrageenan From Kappaphycus Alvarezii (Rhodophyta, Solieriaceae): Metabolism, Structure, Production, and Application. Front. Plant Sci. 2022, 13, 859635. [Google Scholar] [CrossRef] [PubMed]
- Cabrera, R.; Umanzor, S.; Díaz-Larrea, J.; Araújo, P.G. Kappaphycus alvarezii (Rhodophyta): New Record of an Exotic Species for the Caribbean Coast of Costa Rica. Am. J. Plant Sci. 2019, 10, 1888–1902. [Google Scholar] [CrossRef]
- Castelar, B.; de Siqueira, M.F.; Sánchez-Tapia, A.; Reis, R.P. Risk Analysis Using Species Distribution Modeling to Support Public Policies for the Alien Alga Kappaphycus alvarezii Aquaculture in Brazil. Aquaculture 2015, 446, 217–226. [Google Scholar] [CrossRef]
- Masarin, F.; Cedeno, F.R.P.; Chavez, E.G.S.; de Oliveira, L.E.; Gelli, V.C.; Monti, R. Chemical Analysis and Biorefinery of Red Algae Kappaphycus Alvarezii for Efficient Production of Glucose from Residue of Carrageenan Extraction Process. Biotechnol. Biofuels 2016, 9, 122. [Google Scholar] [CrossRef] [PubMed]
- Lechat, H.; Amat, M.; Mazoyer, J.; Gallant2, D.J.; Buléon, A.; Lahaye, M. Cell Wall Composition of the Carrageenophyte Kappaphycus alvarezii (Gigartinales, Rhodophyta) Partitioned by Wet Sieving. Hydrobiologia 1997, 9, 565–572. [Google Scholar] [CrossRef]
- Estevez, J.M.; Ciancia, M.; Cerezo, A.S. The System of Galactans of the Red Seaweed, Kappaphycus alvarezii, with Emphasis on Its Minor Constituents. Carbohydr. Res. 2004, 339, 2575–2592. [Google Scholar] [CrossRef]
- Suresh Kumar, K.; Ganesan, K.; Selvaraj, K.; Subba Rao, P.V. Studies on the Functional Properties of Protein Concentrate of Kappaphycus alvarezii (Doty) Doty—An Edible Seaweed. Food Chem. 2014, 153, 353–360. [Google Scholar] [CrossRef]
- Indriatmoko; Heriyanto; Limantara, L.; Brotosudarmo, T.H.P. Composition of Photosynthetic Pigments in a Red Alga Kappaphycus alvarezi Cultivated in Different Depths. Procedia Chem. 2015, 14, 193–201. [Google Scholar] [CrossRef]
- Meinita, M.D.N.; Hong, Y.-K.; Jeong, G.-T. Comparison of Sulfuric and Hydrochloric Acids as Catalysts in Hydrolysis of Kappaphycus alvarezii (Cottonii). Bioprocess. Biosyst. Eng. 2012, 35, 123–128. [Google Scholar] [CrossRef]
- Ra, C.H.; Nguyen, T.H.; Jeong, G.T.; Kim, S.K. Evaluation of Hyper Thermal Acid Hydrolysis of Kappaphycus alvarezii for Enhanced Bioethanol Production. Bioresour. Technol. 2016, 209, 66–72. [Google Scholar] [CrossRef]
- Khambhaty, Y.; Mody, K.; Gandhi, M.R.; Thampy, S.; Maiti, P.; Brahmbhatt, H.; Eswaran, K.; Ghosh, P.K. Kappaphycus alvarezii as a Source of Bioethanol. Bioresour. Technol. 2012, 103, 180–185. [Google Scholar] [CrossRef]
- Tabacof, A.; Calado, V.; Pereira, N. Lactic Acid Fermentation of Carrageenan Hydrolysates from the Macroalga Kappaphycus alvarezii: Evaluating Different Bioreactor Operation Modes. Polysaccharides 2023, 4, 256–270. [Google Scholar] [CrossRef]
- Abd-Rahim, F.; Wasoh, H.; Rafein, M.; Ariff, A. Food Hydrocolloids Production of High Yield Sugars from Kappaphycus alvarezii Using Combined Methods of Chemical and Enzymatic Hydrolysis. Food Hydrocoll. 2014, 42, 309–315. [Google Scholar] [CrossRef]
- Puspawati, S.; Wagiman; Ainuri, M.; Nugraha, D.A. Haslianti The Production of Bioethanol Fermentation Substrate from Eucheuma Cottonii Seaweed through Hydrolysis by Cellulose Enzyme. Agric. Agric. Sci. Procedia 2015, 3, 200–205. [Google Scholar] [CrossRef]
- Dave, N.; Selvaraj, R.; Varadavenkatesan, T.; Vinayagam, R. A Critical Review on Production of Bioethanol from Macroalgal Biomass. Algal Res. 2019, 42, 101606. [Google Scholar] [CrossRef]
- Oh, S.J.; Park, J.; Na, J.G.; Oh, Y.K.; Chang, Y.K. Production of 5-Hydroxymethylfurfural from Agarose by Using a Solid Acid Catalyst in Dimethyl Sulfoxide. RSC Adv. 2015, 5, 47983–47989. [Google Scholar] [CrossRef]
- Meinita, M.D.N.; Hong, Y.-K.; Jeong, G.-T. Detoxification of Acidic Catalyzed Hydrolysate of Kappaphycus alvarezii (Cottonii). Bioprocess. Biosyst. Eng. 2012, 35, 93–98. [Google Scholar] [CrossRef]
- Santos, D.H.d.S.; Xiao, Y.; Chaukura, N.; Hill, J.M.; Selvasembian, R.; Zanta, C.L.P.S.; Meili, L. Regeneration of Dye-Saturated Activated Carbon through Advanced Oxidative Processes: A Review. Heliyon 2022, 8, e10205. [Google Scholar] [CrossRef]
- Zanella, O.; Tessaro, I.C.; Féris, L.A. Desorption- and Decomposition-Based Techniques for the Regeneration of Activated Carbon. Chem. Eng. Technol. 2014, 37, 1447–1459. [Google Scholar] [CrossRef]
- Carratalá-Abril, J.; Lillo-Ródenas, M.A.; Linares-Solano, A.; Cazorla-Amorós, D. Regeneration of Activated Carbons Saturated with Benzene or Toluene Using an Oxygen-Containing Atmosphere. Chem. Eng. Sci. 2010, 65, 2190–2198. [Google Scholar] [CrossRef]
- Hayes, M.H.B.; Mylotte, R.; Swift, R.S. Humin: Its Composition and Importance in Soil Organic Matter. In Advances in Agronomy; Sparks, D.L., Ed.; Academic Press: Cambridge, MA, USA, 2017; Volume 143, pp. 47–138. [Google Scholar]
- Xu, Z.; Yang, Y.; Yan, P.; Xia, Z.; Liu, X.; Zhang, Z.C. Mechanistic Understanding of Humin Formation in the Conversion of Glucose and Fructose to 5-Hydroxymethylfurfural in [BMIM]Cl Ionic Liquid. RSC Adv. 2020, 10, 34732–34737. [Google Scholar] [CrossRef] [PubMed]
- Calderón, J.C.V.; Arora, J.S.; Mushrif, S.H. Mechanistic Investigation into the Formation of Humins in Acid-Catalyzed Biomass Reactions. ACS Omega 2022, 7, 44786–44795. [Google Scholar] [CrossRef]
- Liu, S.; Zhu, Y.; Liao, Y.; Wang, H.; Liu, Q.; Ma, L.; Wang, C. Advances in Understanding the Humins: Formation, Prevention and Application. Appl. Energy Combust. Sci. 2022, 10, 100062. [Google Scholar] [CrossRef]
- Sumerskii, I.V.; Krutov, S.M.; Zarubin, M.Y. Humin-like Substances Formed under the Conditions of Industrial Hydrolysis of Wood. Russ. J. Appl. Chem. 2010, 83, 320–327. [Google Scholar] [CrossRef]
- Li, C.; Gao, S.; Gao, Q.; Wang, L.; Zhang, J. Characterization of Bulk Soil Humin and Its Alkaline-Soluble and alkaline-Insoluble Fractions. Rev. Bras. Cienc. Solo 2015, 39, 120–126. [Google Scholar] [CrossRef]
- Körner, P.; Jung, D.; Kruse, A. Influence of the PH Value on the Hydrothermal Degradation of Fructose. ChemistryOpen 2019, 8, 1121–1132. [Google Scholar] [CrossRef]
- Xia, J.; Qiu, Z.; Ma, S.; Liu, Q.; Han, R.; Liu, X.; Xu, J. Efficient Polymalic Acid Production from Corn Straw Hydrolysate by Detoxification of Phenolic Inhibitors. Front. Bioeng. Biotechnol. 2023, 11, 1339982. [Google Scholar] [CrossRef]
- Wang, T.; Meng, Y.; Qin, Y.; Feng, W.; Wang, C. Removal of Furfural and HMF from Monosaccharides by Nanofiltration and Reverse Osmosis Membranes. J. Energy Inst. 2018, 91, 473–480. [Google Scholar] [CrossRef]
- Anburajan, P.; Pugazhendhi, A.; Park, J.-H.; Kumar, G.; Choi, C.-S.; Kim, S.-H. Inhibitory Effect of 5-Hydroxymethylfurfural on Continuous Hydrogen Fermentation by Mixed Culture in a Fixed Bed Reactor. Int. J. Hydrogen Energy 2017, 42, 27570–27576. [Google Scholar] [CrossRef]
- Dessie, W.; Xin, F.; Zhang, W.; Zhou, J.; Wu, H.; Ma, J.; Jiang, M. Inhibitory Effects of Lignocellulose Pretreatment Degradation Products (Hydroxymethylfurfural and Furfural) on Succinic Acid Producing Actinobacillus succinogenes. Biochem. Eng. J. 2019, 150, 107263. [Google Scholar] [CrossRef]
- Lohr, D.; Venkov, P.; Zlatanova, J. Transcriptional Regulation in the Yeast GAL Gene Family: A Complex Genetic Network. FASEB J. 1995, 9, 777–787. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.S.; Zhang, R.K.; Liu, Z.H.; Li, B.Z.; Yuan, Y.J. Microbial Adaptation to Enhance Stress Tolerance. Front. Microbiol. 2022, 13. [Google Scholar] [CrossRef]
- Liu, Z.L.; Slininger, P.J.; Gorsich, S.W. Enhanced Biotransformation of Furfural and Hydroxymethylfurfural by Newly Developed Ethanologenic Yeast Strains. Appl. Biochem. Biotechnol. 2005, 121, 0451–0460. [Google Scholar] [CrossRef]
- Monlau, F.; Sambusiti, C.; Barakat, A.; Quéméneur, M.; Trably, E.; Steyer, J.-P.; Carrère, H. Do Furanic and Phenolic Compounds of Lignocellulosic and Algae Biomass Hydrolyzate Inhibit Anaerobic Mixed Cultures? A Comprehensive Review. Biotechnol. Adv. 2014, 32, 934–951. [Google Scholar] [CrossRef]
- Yang, C.-F.; Huang, C.-R. Biotransformation of 5-Hydroxy-Methylfurfural into 2,5-Furan-Dicarboxylic Acid by Bacterial Isolate Using Thermal Acid Algal Hydrolysate. Bioresour. Technol. 2016, 214, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Sana, B. Marine Microbial Enzymes: Current Status and Future Prospects. In Hb25_Springer Handbook of Marine Biotechnology; Springer: Berlin/Heidelberg, Germany, 2015; pp. 905–917. [Google Scholar]
- Gurpilhares, D.d.B.; Cinelli, L.P.; Simas, N.K.; Pessoa, A., Jr.; Sette, L.D. Marine Prebiotics: Polysaccharides and Oligosaccharides Obtained by Using Microbial Enzymes. Food Chem. 2019, 280, 175–186. [Google Scholar] [CrossRef]
- Torres, M.D.; Flórez-Fernández, N.; Domínguez, H. Integral Utilization of Red Seaweed for Bioactive Production. Mar. Drugs 2019, 17, 314. [Google Scholar] [CrossRef]
- Suriya, J.; Bharathiraja, S.; Krishnan, M.; Manivasagan, P.; Kim, S.-K. Extremozymes from Marine Actinobacteria. In Advances in Food and Nutrition Research; Elsevier: Amsterdam, The Netherlands, 2016; pp. 43–66. [Google Scholar]
- Parrilli, E.; Tedesco, P.; Fondi, M.; Tutino, M.L.; Lo Giudice, A.; de Pascale, D.; Fani, R. The Art of Adapting to Extreme Environments: The Model System Pseudoalteromonas. Phys. Life Rev. 2021, 36, 137–161. [Google Scholar] [CrossRef]
- Ghanbarzadeh, M.; Golmoradizadeh, A.; Homaei, A. Carrageenans and Carrageenases: Versatile Polysaccharides and Promising Marine Enzymes. Phytochem. Rev. 2018, 17, 535–571. [Google Scholar] [CrossRef]
- Chen, Q.; Xu, W.; Wu, H.; Guang, C.; Zhang, W.; Mu, W. An Overview of D-Galactose Utilization through Microbial Fermentation and Enzyme-Catalyzed Conversion. Appl. Microbiol. Biotechnol. 2021, 105, 7161–7170. [Google Scholar] [CrossRef] [PubMed]
- Xia, A.; Jacob, A.; Herrmann, C.; Murphy, J.D. Fermentative Bio-Hydrogen Production from Galactose. Energy 2016, 96, 346–354. [Google Scholar] [CrossRef]
- Fonseca, B.C.; Guazzaroni, M.-E.; Reginatto, V. Fermentative Production of H2 from Different Concentrations of Galactose by the New Isolate Clostridium beijerinckii Br21. Int. J. Hydrogen Energy 2016, 41, 21109–21120. [Google Scholar] [CrossRef]
- Park, J.-H.; Lee, S.-H.; Yoon, J.-J.; Kim, S.-H.; Park, H.-D. Predominance of Cluster I Clostridium in Hydrogen Fermentation of Galactose Seeded with Various Heat-Treated Anaerobic Sludges. Bioresour. Technol. 2014, 157, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Ezeji, T.C.; Qureshi, N.; Blaschek, H.P. Bioproduction of Butanol from Biomass: From Genes to Bioreactors. Curr. Opin. Biotechnol. 2007, 18, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Olajuyin, A.M.; Yang, M.; Liu, Y.; Mu, T.; Tian, J.; Adaramoye, O.A.; Xing, J. Efficient Production of Succinic Acid from Palmaria Palmata Hydrolysate by Metabolically Engineered Escherichia Coli. Bioresour. Technol. 2016, 214, 653–659. [Google Scholar] [CrossRef] [PubMed]
- Sudhakar, M.P.; Dharani, G. Evaluation of Seaweed for the Production of Lactic Acid by Fermentation Using Lactobacillus plantarum. Bioresour. Technol. Rep. 2022, 17, 100890. [Google Scholar] [CrossRef]
- Murad, A.E.-H.; Khalaf, S.A.-D. Citric Acid Production from Whey with Sugars and Additives by Aspergillus niger. Afr. J. Biotechnol. 2003, 2, 356–359. [Google Scholar] [CrossRef]
- Lazar, Z.; Gamboa-Meléndez, H.; Le Coq, A.-M.C.-; Neuvéglise, C.; Nicaud, J.-M. Awakening the Endogenous Leloir Pathway for Efficient Galactose Utilization by Yarrowia lipolytica. Biotechnol. Biofuels 2015, 8, 185. [Google Scholar] [CrossRef]
- Ehsanipour, M.; Suko, A.V.; Bura, R. Fermentation of Lignocellulosic Sugars to Acetic Acid by Moorella thermoacetica. J. Ind. Microbiol. Biotechnol. 2016, 43, 807–816. [Google Scholar] [CrossRef]
- He, F.; Qin, S.; Yang, Z.; Bai, X.; Suo, Y.; Wang, J. Butyric Acid Production from Spent Coffee Grounds by Engineered Clostridium tyrobutyricum Overexpressing Galactose Catabolism Genes. Bioresour. Technol. 2020, 304, 122977. [Google Scholar] [CrossRef] [PubMed]
- Al-Abdallah, W.; Dahman, Y. Production of Green Biocellulose Nanofibers by Gluconacetobacter xylinus through Utilizing the Renewable Resources of Agriculture Residues. Bioprocess. Biosyst. Eng. 2013, 36, 1735–1743. [Google Scholar] [CrossRef] [PubMed]
- Hassan, Q.; Azzawi, I.D.J.; Sameen, A.Z.; Salman, H.M. Hydrogen Fuel Cell Vehicles: Opportunities and Challenges. Sustainability 2023, 15, 11501. [Google Scholar] [CrossRef]
- Cheng, K.; Zhao, X.; Zeng, J.; Zhang, J. Biotechnological Production of Succinic Acid: Current State and Perspectives. Biofuels Bioprod. Biorefining 2012, 6, 302–318. [Google Scholar] [CrossRef]
- Almqvist, H.; Pateraki, C.; Alexandri, M.; Koutinas, A.; Lidén, G. Succinic Acid Production by Actinobacillus succinogenes from Batch Fermentation of Mixed Sugars. J. Ind. Microbiol. Biotechnol. 2016, 43, 1117–1130. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.C.; Lee, S.Y.; Chang, H.N. Succinic Acid Production by Anaerobiospirillum Succiniciproducens ATCC 29305 Growing on Galactose, Galactose/Glucose, and Galactose/Lactose. J. Microbiol. Biotechnol. 2008, 18, 1792–1796. [Google Scholar] [CrossRef] [PubMed]
- Alvarado-Morales, M.; Gunnarsson, I.B.; Fotidis, I.A.; Vasilakou, E.; Lyberatos, G.; Angelidaki, I. Laminaria Digitata as a Potential Carbon Source for Succinic Acid and Bioenergy Production in a Biorefinery Perspective. Algal Res. 2015, 9, 126–132. [Google Scholar] [CrossRef]
- Marinho, G.S.; Alvarado-Morales, M.; Angelidaki, I. Valorization of Macroalga Saccharina latissima as Novel Feedstock for Fermentation-Based Succinic Acid Production in a Biorefinery Approach and Economic Aspects. Algal Res. 2016, 16, 102–109. [Google Scholar] [CrossRef]
- Gutierrez, N.A.; McKay, I.A.; French, C.E.; Brooks, J.D.; Maddox, I.S. Repression of Galactose Utilization by Glucose in the Citrate-Producing YeastCandida Guilliermondii. J. Ind. Microbiol. 1993, 11, 143–146. [Google Scholar] [CrossRef]
- Dahman, Y.; Jayasuriya, K.E.; Kalis, M. Potential of Biocellulose Nanofibers Production from Agricultural Renewable Resources: Preliminary Study. Appl. Biochem. Biotechnol. 2010, 162, 1647–1659. [Google Scholar] [CrossRef]
- Mikkelsen, D.; Flanagan, B.M.; Dykes, G.A.; Gidley, M.J. Influence of Different Carbon Sources on Bacterial Cellulose Production by Gluconacetobacter xylinus Strain ATCC 53524. J. Appl. Microbiol. 2009, 107, 576–583. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).