
Continuous Secretion of Human Epidermal Growth Factor Based on Escherichia coli Biofilm


 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Strains and Plasmids
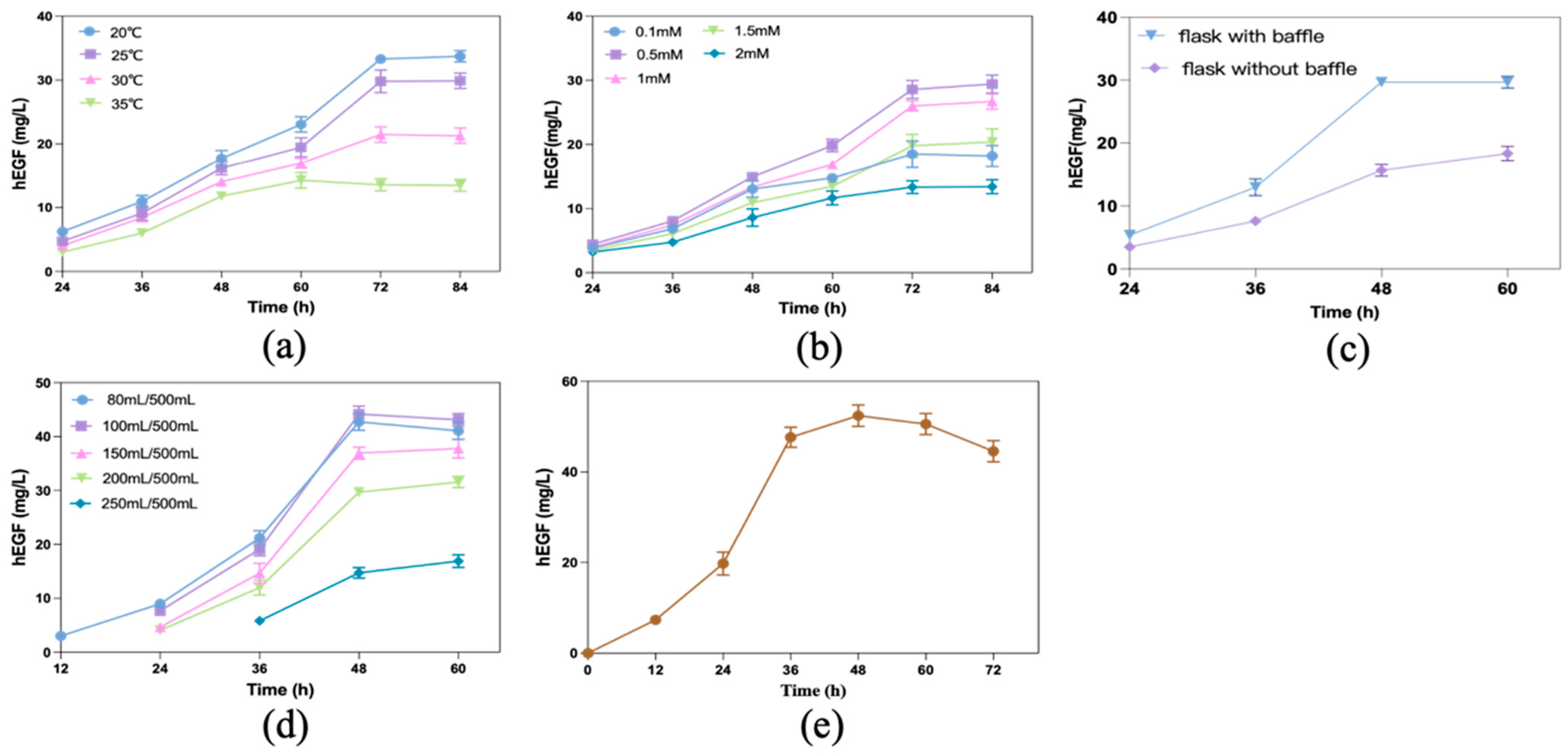
2.2. Media and Growth Conditions
2.3. Fermentation and Analytical Methods
2.4. Characterization of Biofilm Formation
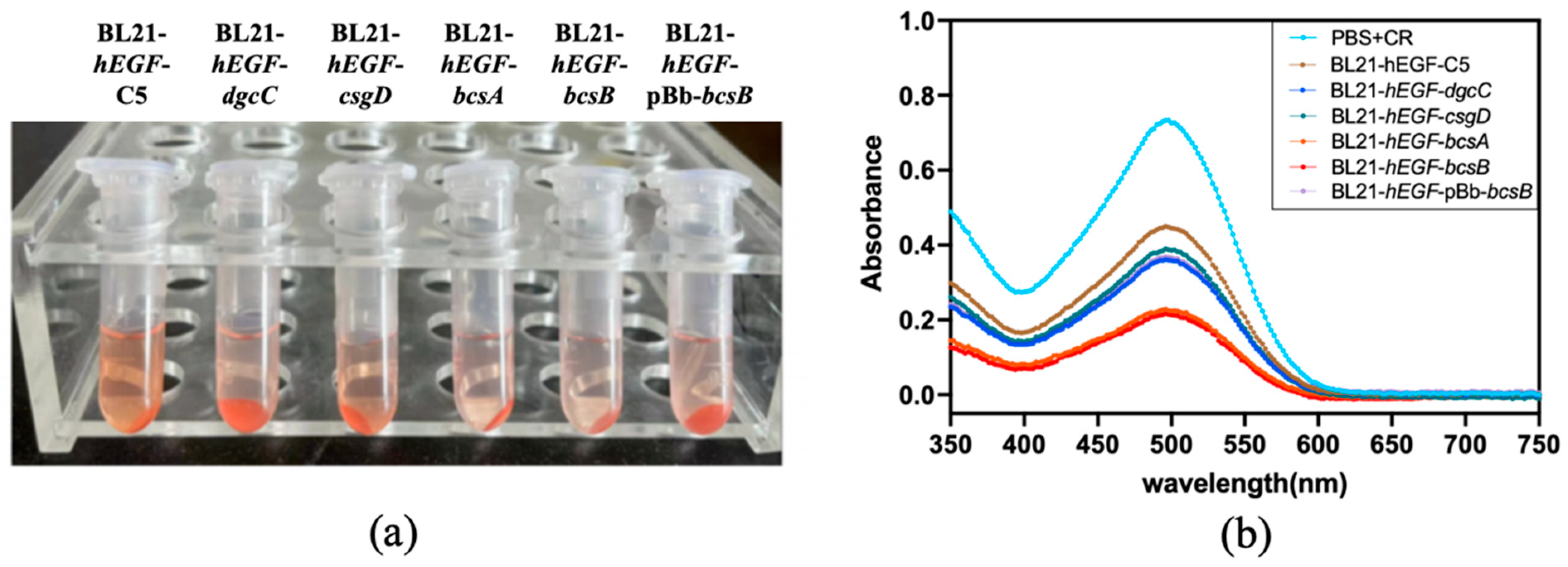
2.5. Congo Red Assay
2.6. Confocal Laser Scanning Microscope
3. Results
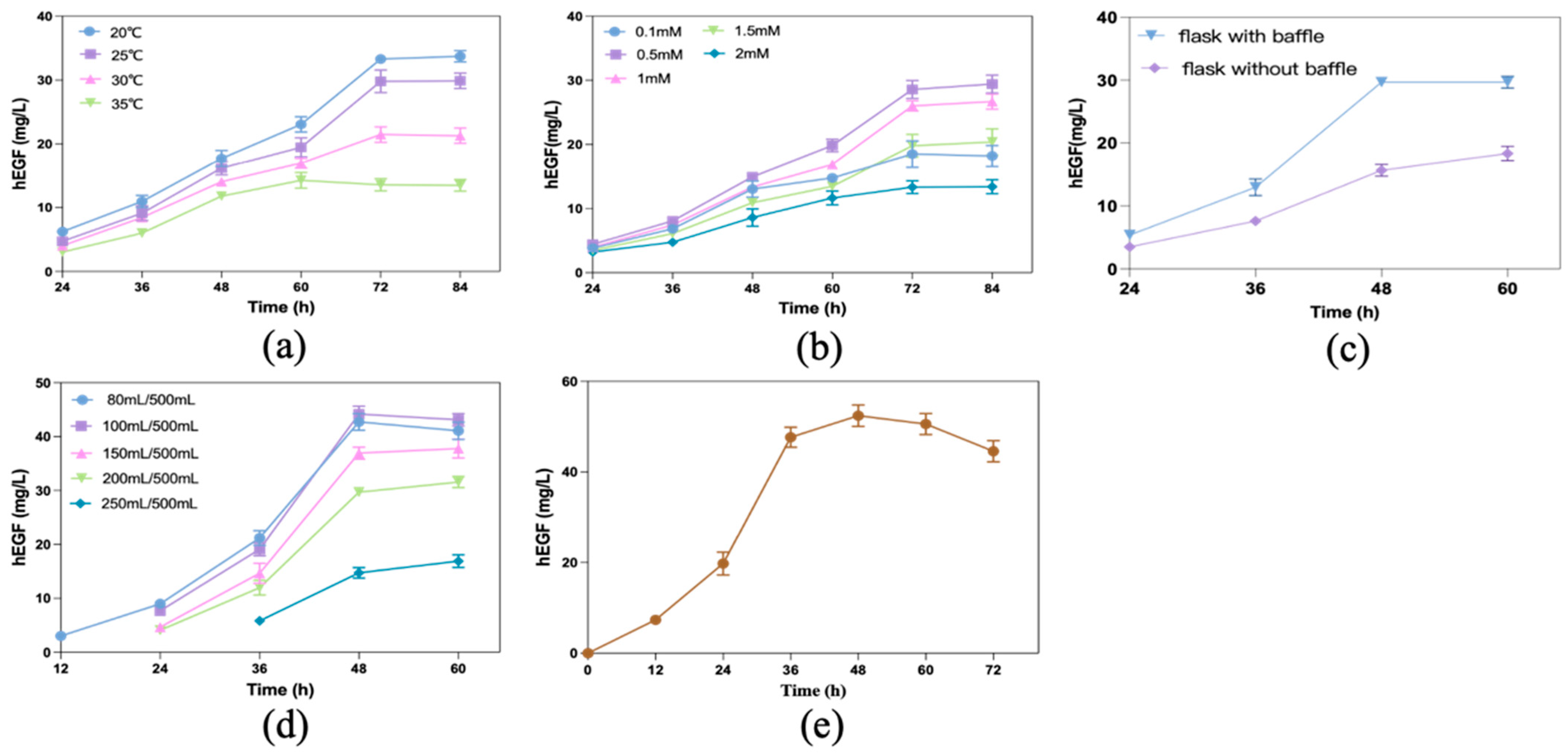
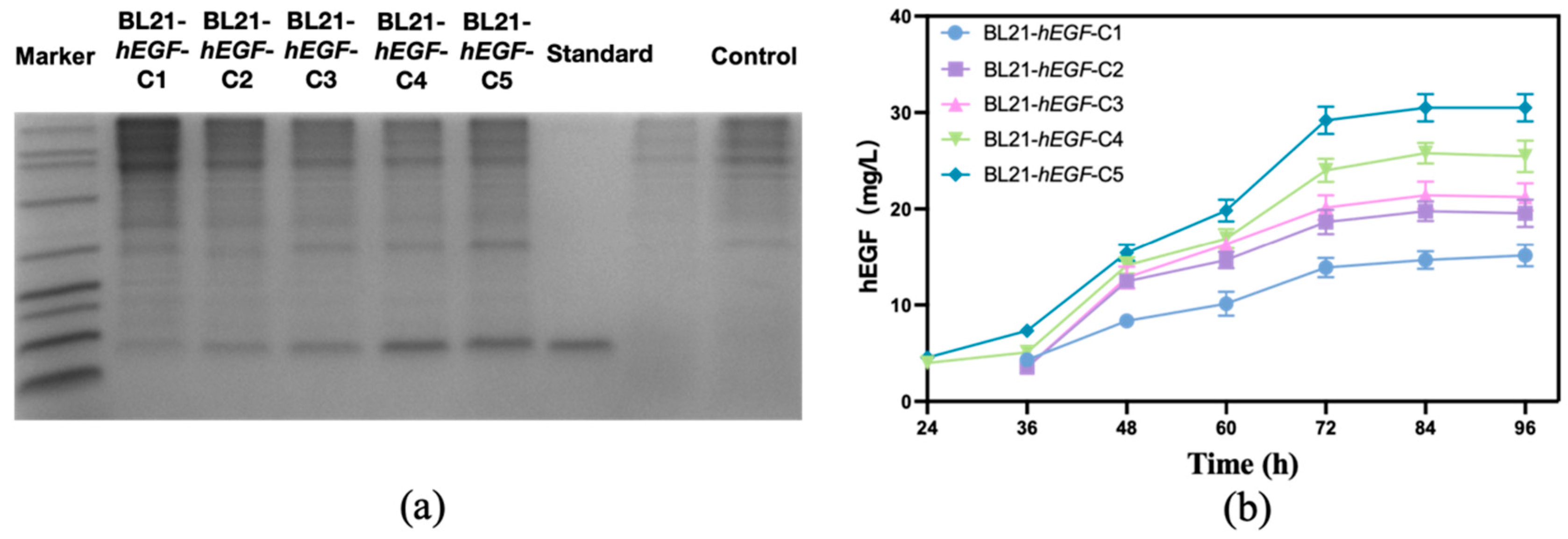
3.1. Screening Results of High Production Strain
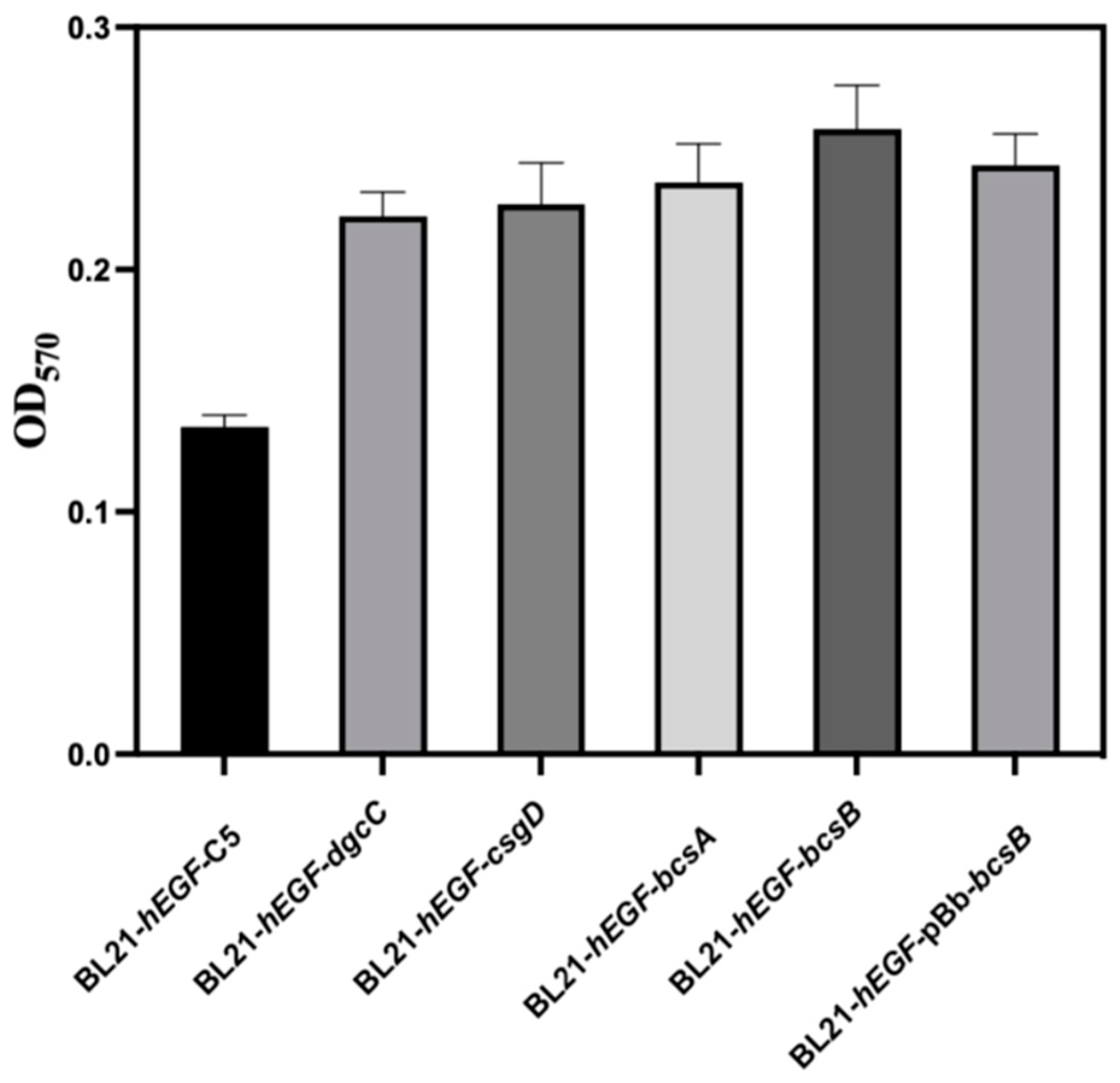
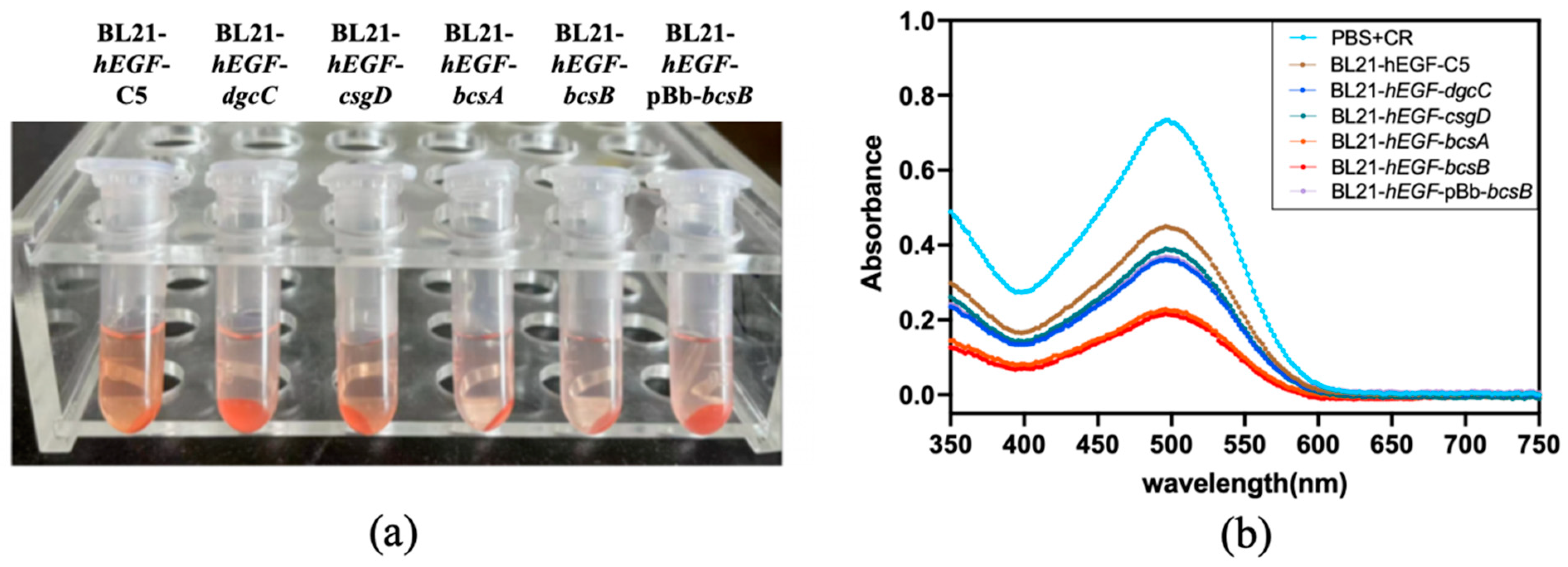
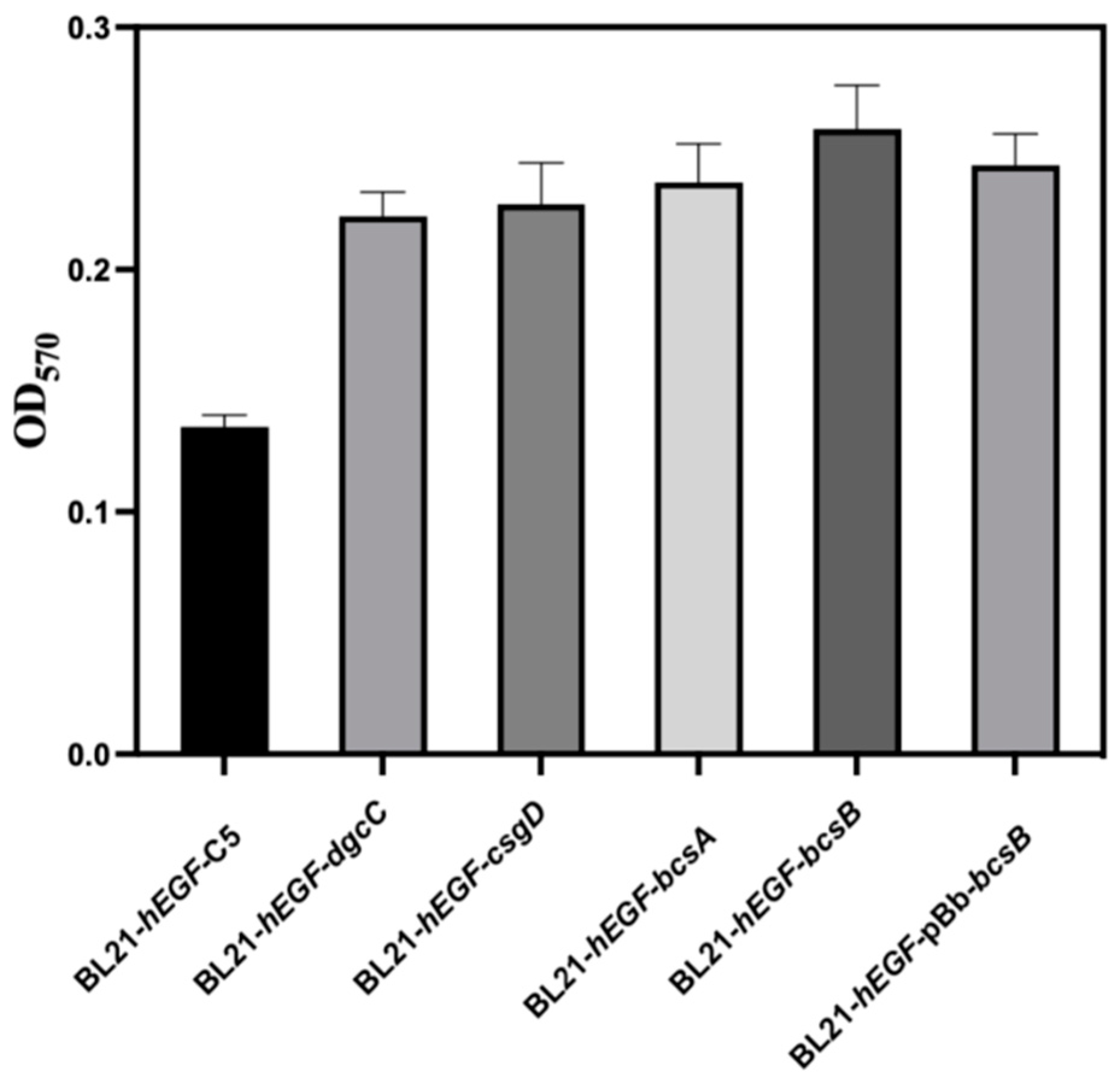
3.2. Effect of Gene Modification on Biofilm Formation
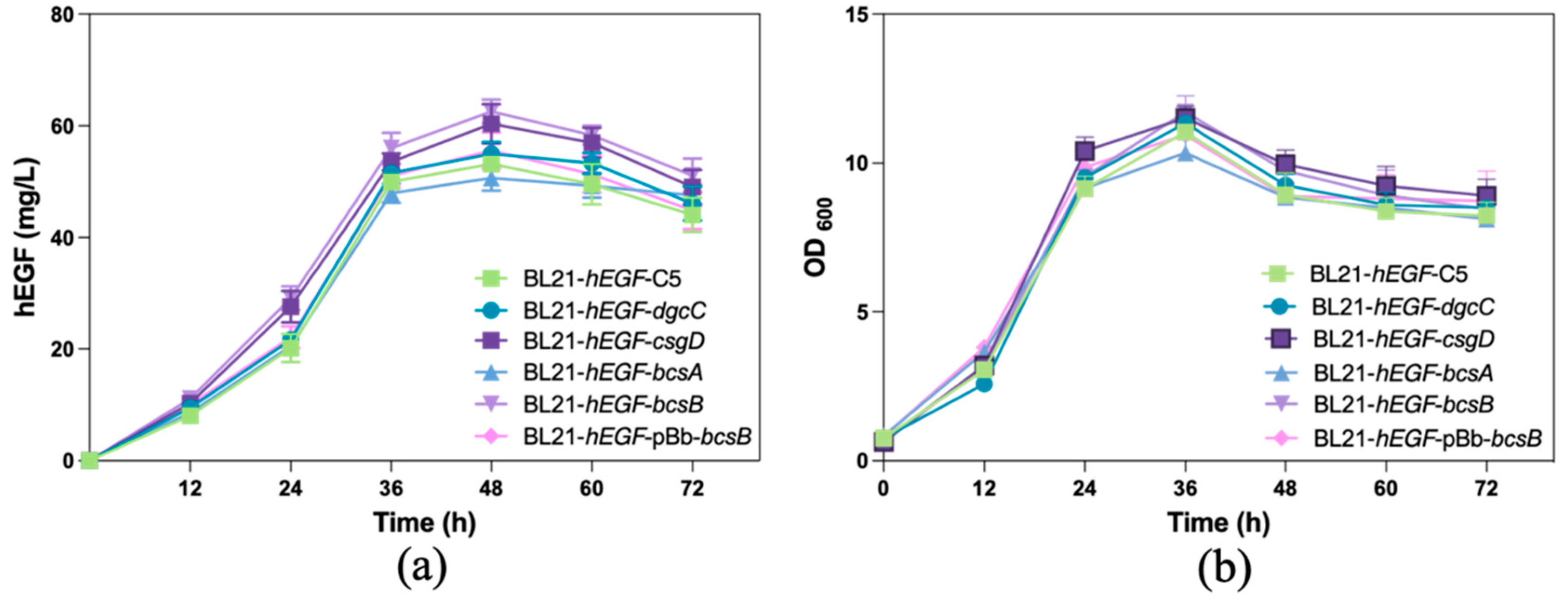
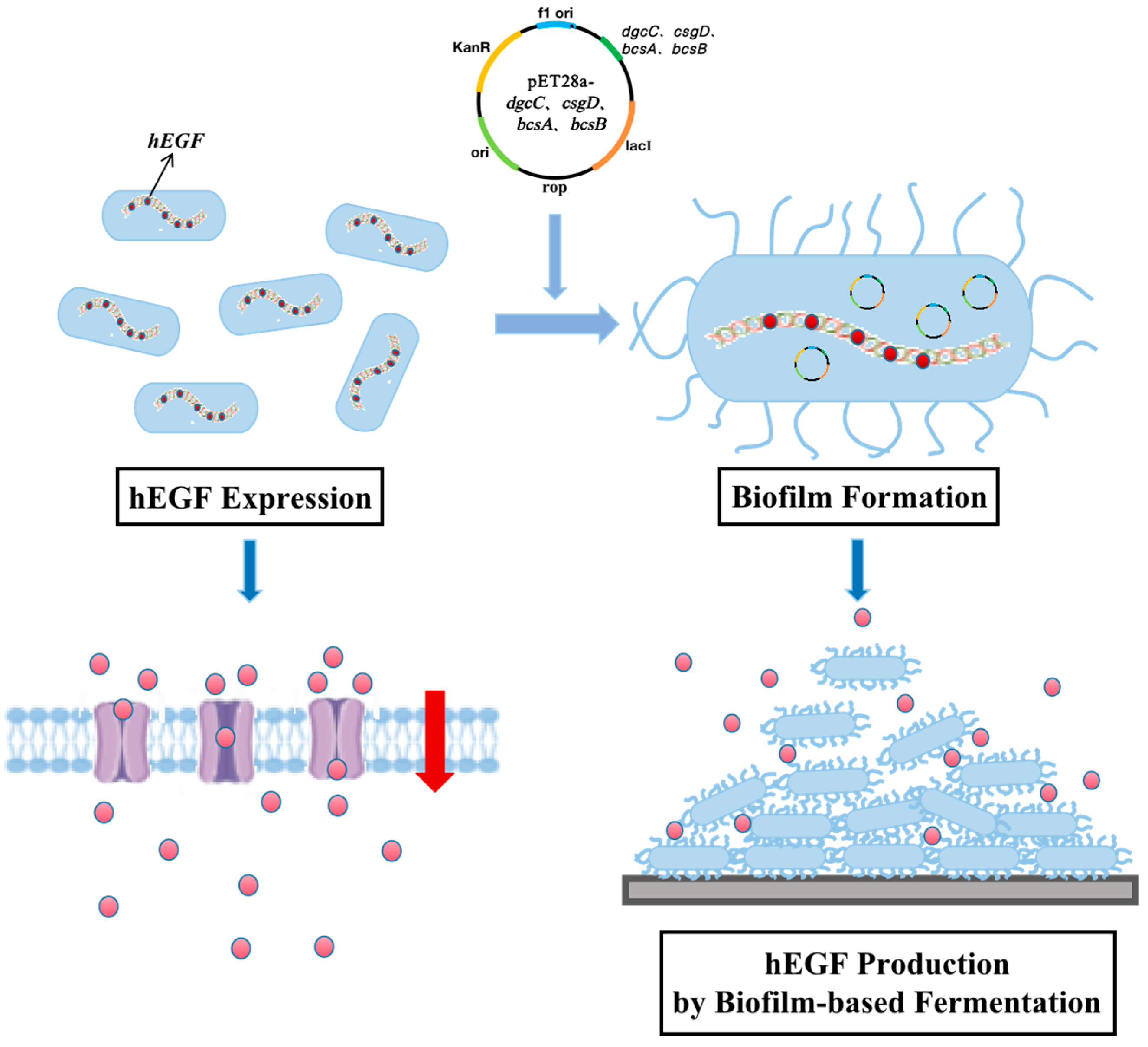
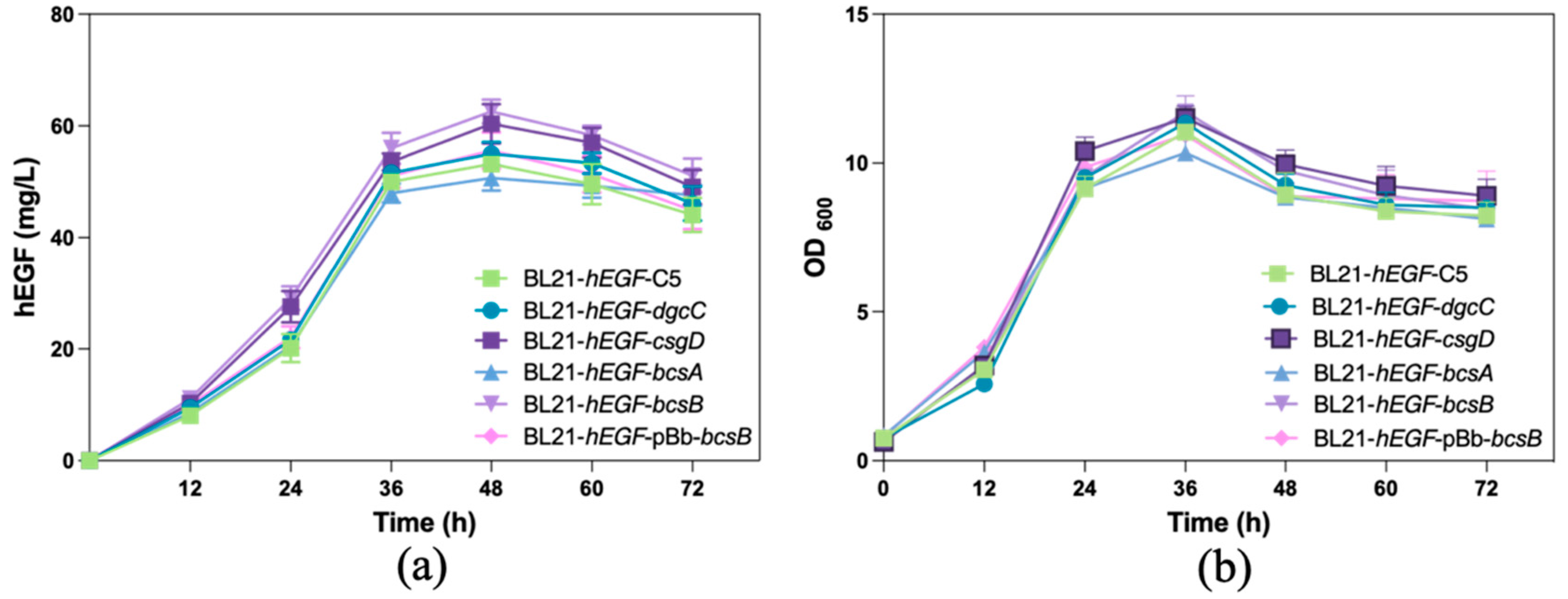
3.3. Effects of Overexpression of Biofilm-Related Genes on hEGF Secretion
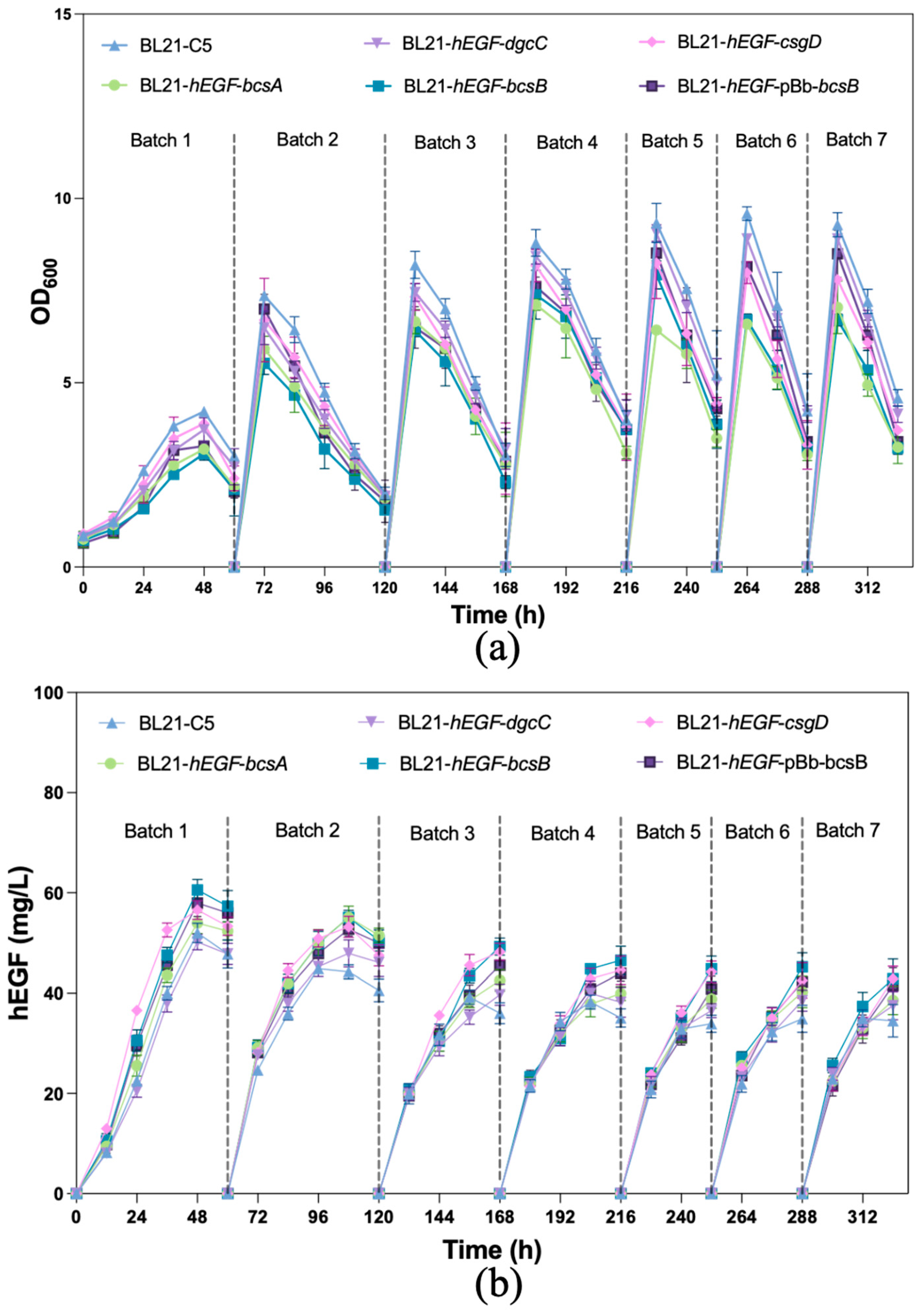
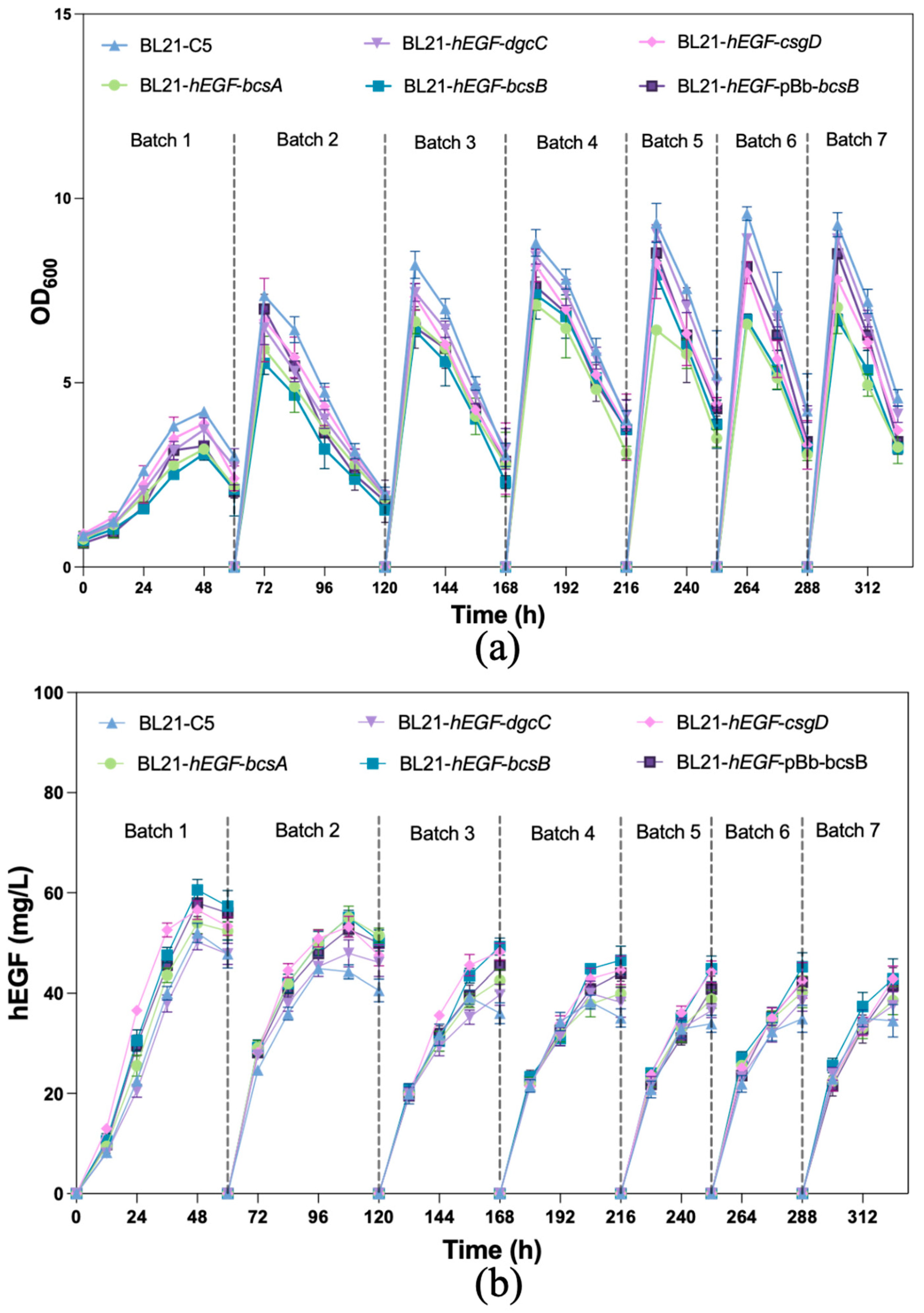
3.4. Production of hEGF by Biofilm-Immobilized Continuous Fermentation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Qiang, W.; Zhou, T.; Lan, X.; Zhang, X.; Guo, Y.; Noman, M.; Du, L.; Zheng, J.; Li, W.; Li, H.; et al. A New Nanoscale Transdermal Drug Delivery System: Oil Body-Linked Oleosin-hEGF Improves Skin Regeneration to Accelerate Wound Healing. J. Nanobiotechnology 2018, 16, 62. [Google Scholar] [CrossRef] [PubMed]
- Kwong, K.W.Y.; Ng, A.K.L.; Wong, W.K.R. Engineering Versatile Protein Expression Systems Mediated by Inteins in Escherichia coli. Appl. Microbiol. Biotechnol. 2016, 100, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Park, S.A.; Bhatia, S.K.; Park, H.A.; Kim, S.Y.; Sudheer, P.D.V.N.; Yang, Y.-H.; Choi, K.-Y. Bacillus subtilis as a Robust Host for Biochemical Production Utilizing Biomass. Crit. Rev. Biotechnol. 2021, 41, 827–848. [Google Scholar] [CrossRef]
- Wu, M.; Ruan, J.; Ye, X.; Zhao, S.; Tang, X.; Wang, X.; Li, H.; Zhong, B. P25 Gene Knockout Contributes to Human Epidermal Growth Factor Production in Transgenic Silkworms. Int. J. Mol. Sci. 2021, 22, 2709. [Google Scholar] [CrossRef] [PubMed]
- Pontrelli, S.; Chiu, T.-Y.; Lan, E.I.; Chen, F.Y.-H.; Chang, P.; Liao, J.C. Escherichia coli as a Host for Metabolic Engineering. Metab. Eng. 2018, 50, 16–46. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.H.; Lee, S.Y. Secretory and Extracellular Production of Recombinant Proteins Using Escherichia coli. Appl. Microbiol. Biotechnol. 2004, 64, 625–635. [Google Scholar] [CrossRef] [PubMed]
- Indriyani, A.; Anggraeni, N.I.; Sriwidodo, S.; Maksum, I.P. Optimization Extracellular Secretion of Recombinant Human Epidermal Growth Factor (hEGF) in Escherichia coli BL21 (DE3) pD881-OmpA-hEGF by Using Response Surface Method (RSM). Int. J. Res. Pharm. Sci. 2019, 10, 1824–1831. [Google Scholar] [CrossRef]
- Sriwidodo, S.; Maksum, I.P.; Riswanto, N.; Rostinawati, T.; Subroto, T. Extracellular Secretion Recombinant of Human Epidermal Growth Factor (hEGF) Using Pectate Lyase B (PelB) Signal Peptide in Escherichia coli BL21 (DE3). Int. J. Res. Pharm. Sci. 2017, 8, 33–40. [Google Scholar]
- Yang, Z.; Zhang, Z. Engineering Strategies for Enhanced Production of Protein and Bio-Products in Pichia pastoris: A Review. Biotechnol. Adv. 2018, 36, 182–195. [Google Scholar] [CrossRef]
- Karygianni, L.; Ren, Z.; Koo, H.; Thurnheer, T. Biofilm Matrixome: Extracellular Components in Structured Microbial Communities. Trends Microbiol. 2020, 28, 668–681. [Google Scholar] [CrossRef]
- Ferrando, D.; Toubiana, D.; Kandiyote, N.S.; Nguyen, T.H.; Nejidat, A.; Herzberg, M. Ambivalent Role of Calcium in the Viscoelastic Properties of Extracellular Polymeric Substances and the Consequent Fouling of Reverse Osmosis Membranes. Desalination 2018, 429, 12–19. [Google Scholar] [CrossRef]
- Lan, T.-Q.; Wei, D.; Yang, S.-T.; Liu, X. Enhanced Cellulase Production by Trichoderma viride in a Rotating Fibrous Bed Bioreactor. Bioresour. Technol. 2013, 133, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Morikawa, M. Beneficial Biofilm Formation by Industrial Bacteria Bacillus subtilis and Related Species. J. Biosci. Bioeng. 2006, 101, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Mohsin, M.Z.; Omer, R.; Huang, J.; Mohsin, A.; Guo, M.; Qian, J.; Zhuang, Y. Advances in Engineered Bacillus subtilis Biofilms and Spores, and Their Applications in Bioremediation, Biocatalysis, and Biomaterials. Synth. Syst. Biotechnol. 2021, 6, 180–191. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Wang, Z.; Zhou, M.; Zhang, C.; Zhi, K.; Liu, S.; Sun, X.; Wang, Z.; Liu, J.; Liu, D. Continuous Production of Human Epidermal Growth Factor Using Escherichia coli Biofilm. Front. Microbiol. 2022, 13, 855059. [Google Scholar] [CrossRef]
- Li, Q.; Sun, B.; Chen, J.; Zhang, Y.; Jiang, Y.; Yang, S. A Modified pCas/pTargetF System for CRISPR-Cas9-Assisted Genome Editing in Escherichia coli. Acta Biochim. Biophys. Sin. 2021, 53, 620–627. [Google Scholar] [CrossRef] [PubMed]
- Gasser, B.; Saloheimo, M.; Rinas, U.; Dragosits, M.; Rodríguez-Carmona, E.; Baumann, K.; Giuliani, M.; Parrilli, E.; Branduardi, P.; Lang, C.; et al. Protein Folding and Conformational Stress in Microbial Cells Producing Recombinant Proteins: A Host Comparative Overview. Microb. Cell Factories 2008, 7, 11. [Google Scholar] [CrossRef] [PubMed]
- Dvorak, P.; Chrast, L.; Nikel, P.I.; Fedr, R.; Soucek, K.; Sedlackova, M.; Chaloupkova, R.; de Lorenzo, V.; Prokop, Z.; Damborsky, J. Exacerbation of Substrate Toxicity by IPTG in Escherichia coli BL21(DE3) Carrying a Synthetic Metabolic Pathway. Microb. Cell Factories 2015, 14, 201. [Google Scholar] [CrossRef] [PubMed]
- Yakupova, E.I.; Bobyleva, L.G.; Vikhlyantsev, I.M.; Bobylev, A.G. Congo Red and Amyloids: History and Relationship. Biosci. Rep. 2019, 39, BSR20181415. [Google Scholar] [CrossRef]
- Mahto, K.U.; Kumari, S.; Das, S. Unraveling the Complex Regulatory Networks in Biofilm Formation in Bacteria and Relevance of Biofilms in Environmental Remediation. Crit. Rev. Biochem. Mol. Biol. 2022, 57, 305–332. [Google Scholar] [CrossRef]
- Yin, W.; Wang, Y.; Liu, L.; He, J. Biofilms: The Microbial “Protective Clothing” in Extreme Environments. Int. J. Mol. Sci. 2019, 20, 3423. [Google Scholar] [CrossRef] [PubMed]
- Castiblanco, L.F.; Sundin, G.W. Cellulose Production, Activated by Cyclic Di-GMP through BcsA and BcsZ, is a Virulence Factor and an Essential Determinant of the Three-Dimensional Architectures of Biofilms Formed by Erwinia amylovora Ea1189. Mol. Plant Pathol. 2018, 19, 90–103. [Google Scholar] [CrossRef] [PubMed]
- Tsai, M.-H.; Liang, Y.-H.; Chen, C.-L.; Chiu, C.-H. Characterization of Salmonella Resistance to Bile during Biofilm Formation. J. Microbiol. Immunol. Infect. 2020, 53, 518–524. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains or Plasmids | Relevant Characteristics | Sources | |
|---|---|---|---|
| Strains | E. coli BL21(DE3) | Host strain | Invitrogen |
| BL21-hEGF-C1 | Genome-integrated expression of hEGF in one locus | This study | |
| BL21-hEGF-C2 | Genome-integrated expression of hEGF in two loci | This study | |
| BL21-hEGF-C3 | Genome-integrated expression of hEGF in three loci | This study | |
| BL21-hEGF-C4 | Genome-integrated expression of hEGF in four loci | This study | |
| BL21-hEGF-C5 | Genome-integrated expression of hEGF in five loci | This study | |
| BL21-hEGF-dgcC | Plasmid-based expression of dgcC | This study | |
| BL21-hEGF-csgD | Plasmid-based expression of csgD | This study | |
| BL21-hEGF-bcsA | Plasmid-based expression of bcsA | This study | |
| BL21-hEGF-bcsB | Plasmid-based expression of bcsB | This study | |
| BL21-hEGF-pBb-bcsB * | Plasmid-based expression of bcsB | This study | |
| Plasmids | pET30a-PelB-hEGF | hEGF expression | [15] |
| pCas | CRISPR editing | [16] | |
| pTarget | CRISPR editing | [16] | |
| pET28a | Expression vector | Invitrogen | |
| pBbE1a | Expression vector | [15] | |
| pET28a-dgcC | Plasmid-based expression of dgcC | This study | |
| pET28a-csgD | Plasmid-based expression of csgD | This study | |
| pET28a-bcsA | Plasmid-based expression of bcsA | This study | |
| pET28a-bcsB | Plasmid-based expression of bcsB | This study | |
| pBbE1a-bcsB | Plasmid-based expression of bcsB | [15] | |
| Strains | Congo Red Binding Ratio |
|---|---|
| BL21-hEGF-C5 | 0.37 |
| BL21-hEGF-dgcC | 0.51 |
| BL21-hEGF-csgD | 0.47 |
| BL21-hEGF-bcsA | 0.69 |
| BL21-hEGF-bcsB | 0.72 |
| BL21-hEGF-pBb-bcsB | 0.51 |
| Strains | hEGF Secretion of the Modified Strains Compared to the Control Strain (%) | |||||||
|---|---|---|---|---|---|---|---|---|
| 1st | 2nd | 3rd | 4th | 5th | 6th | 7th | Average | |
| BL21-hEGF-dgcC | −3.5 | 8.4 | 10.2 | 8.9 | 7.7 | 10.3 | 8.7 | 7.2 |
| BL21-hEGF-csgD | 8.8 | 20.3 | 33.9 | 27.0 | 30.4 | 21.8 | 24.1 | 23.8 |
| BL21-hEGF-bcsA | 3.8 | 24.5 | 18.1 | 13.9 | 14.5 | 15.3 | 11.7 | 14.5 |
| BL21-hEGF-bcsB | 16.5 | 24.5 | 36.7 | 32.7 | 32.2 | 29.6 | 24.1 | 28.0 |
| BL21-hEGF-pBb- bcsB | 11.5 | 19.2 | 26.4 | 25.3 | 20.4 | 21.0 | 20.4 | 20.6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, C.; Liao, J.; Li, Y.; Liu, S.; Li, M.; Zhang, D.; Wang, Z.; Liu, D.; Ying, H. Continuous Secretion of Human Epidermal Growth Factor Based on Escherichia coli Biofilm. Fermentation 2024, 10, 202. https://doi.org/10.3390/fermentation10040202
Zhang C, Liao J, Li Y, Liu S, Li M, Zhang D, Wang Z, Liu D, Ying H. Continuous Secretion of Human Epidermal Growth Factor Based on Escherichia coli Biofilm. Fermentation. 2024; 10(4):202. https://doi.org/10.3390/fermentation10040202
Chicago/Turabian StyleZhang, Chong, Jinglin Liao, Yuancong Li, Shuli Liu, Mengting Li, Di Zhang, Zhenyu Wang, Dong Liu, and Hanjie Ying. 2024. "Continuous Secretion of Human Epidermal Growth Factor Based on Escherichia coli Biofilm" Fermentation 10, no. 4: 202. https://doi.org/10.3390/fermentation10040202
APA StyleZhang, C., Liao, J., Li, Y., Liu, S., Li, M., Zhang, D., Wang, Z., Liu, D., & Ying, H. (2024). Continuous Secretion of Human Epidermal Growth Factor Based on Escherichia coli Biofilm. Fermentation, 10(4), 202. https://doi.org/10.3390/fermentation10040202