
Optogenetic Fine-Tuning of Sus scrofa Basic Fibroblast Growth Factor Expression in Escherichia coli

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Plasmids
2.2. Media and Culture Conditions
2.3. Construction of Expression Strains
2.4. Blue Light-Induced Expression System
2.5. Protein Expression, Purification and Analysis
2.6. RNA Extraction and Quantitative Real-Time PCR
2.7. Genetic Optimization of Engineered bFGF-Producing Strain
2.8. Optimization of Blue Light-Induced bFGF Fermentation
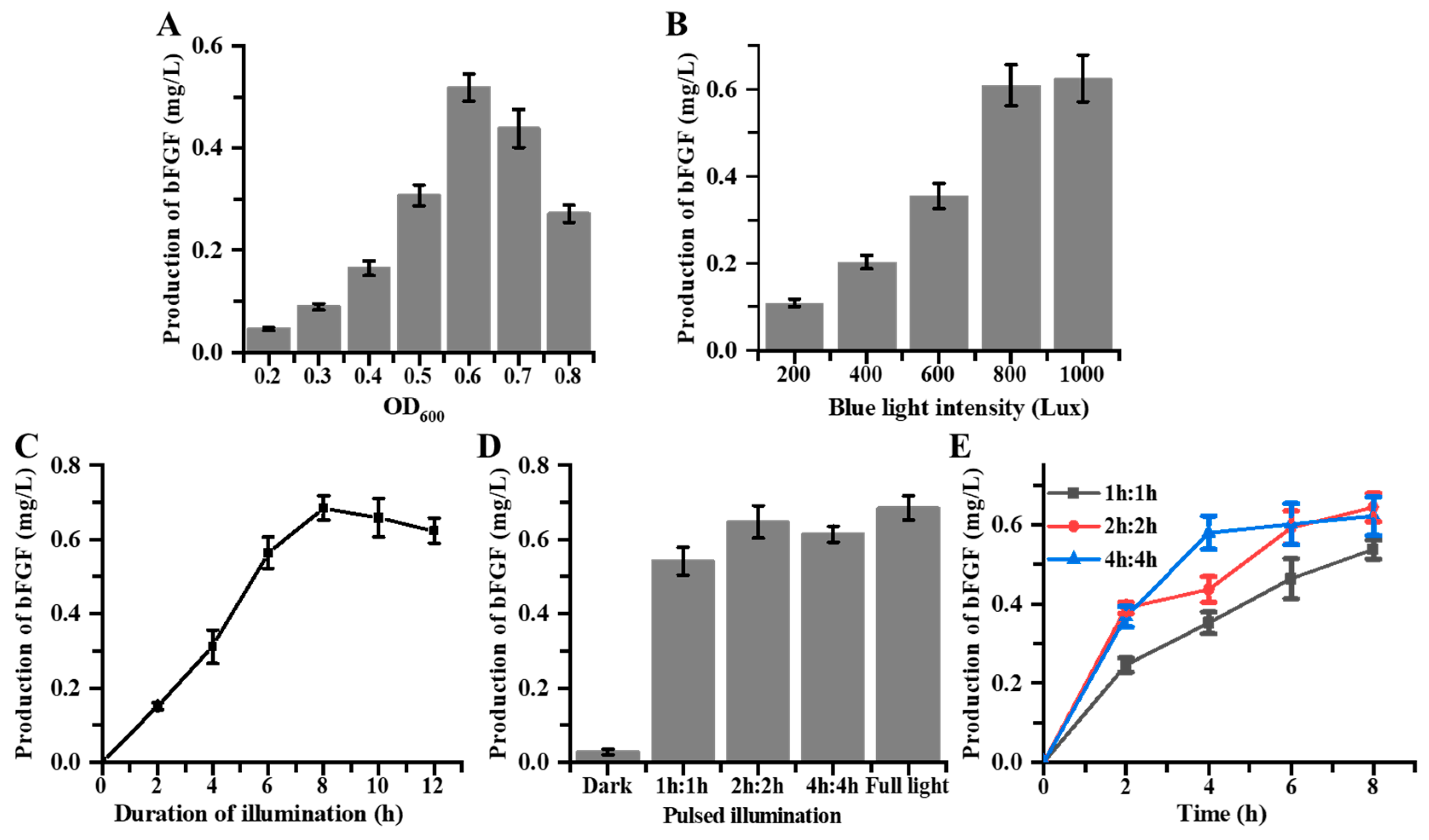
3. Results and Discussion
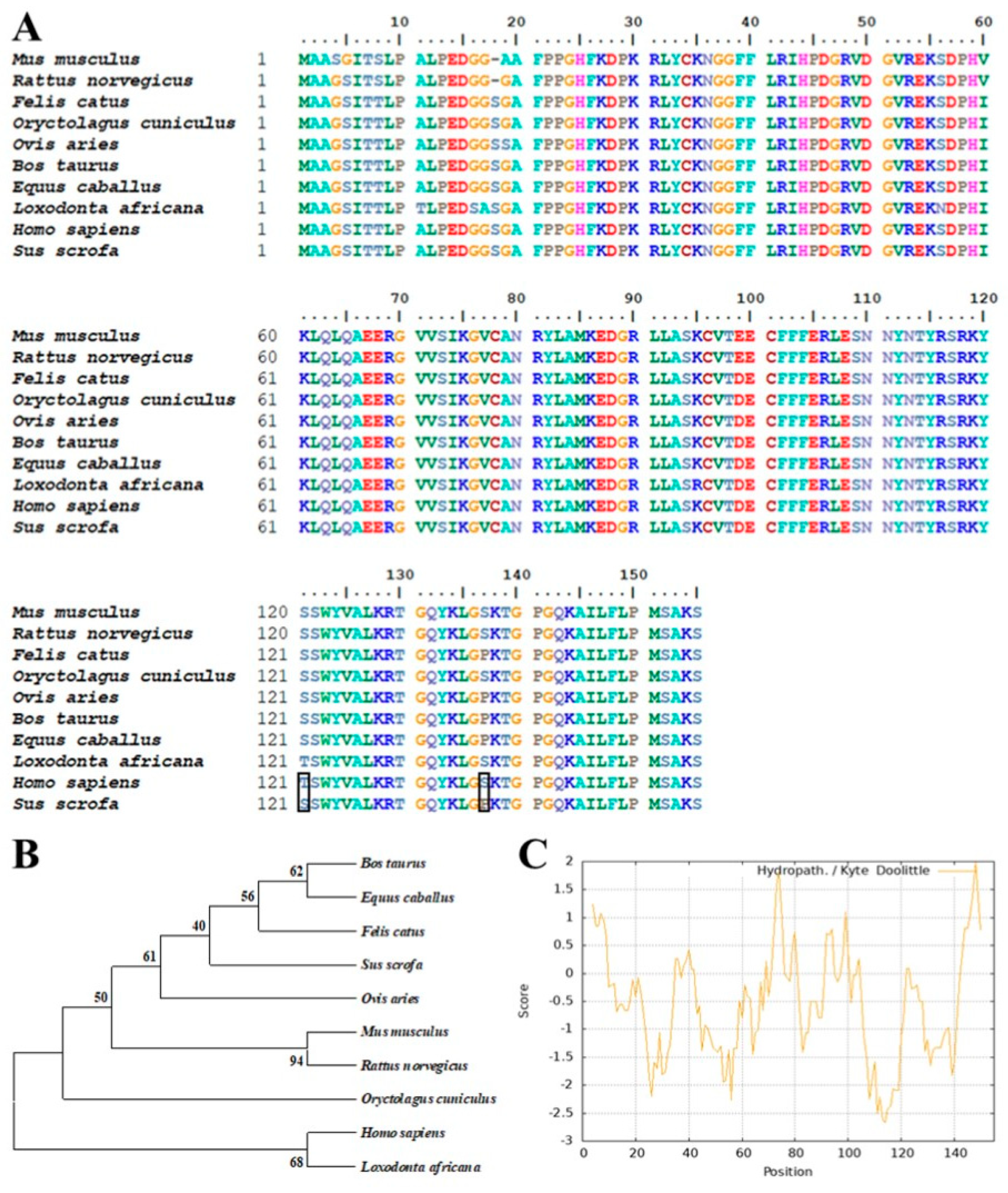
3.1. Bioinformatics Analysis of Porcine bFGF
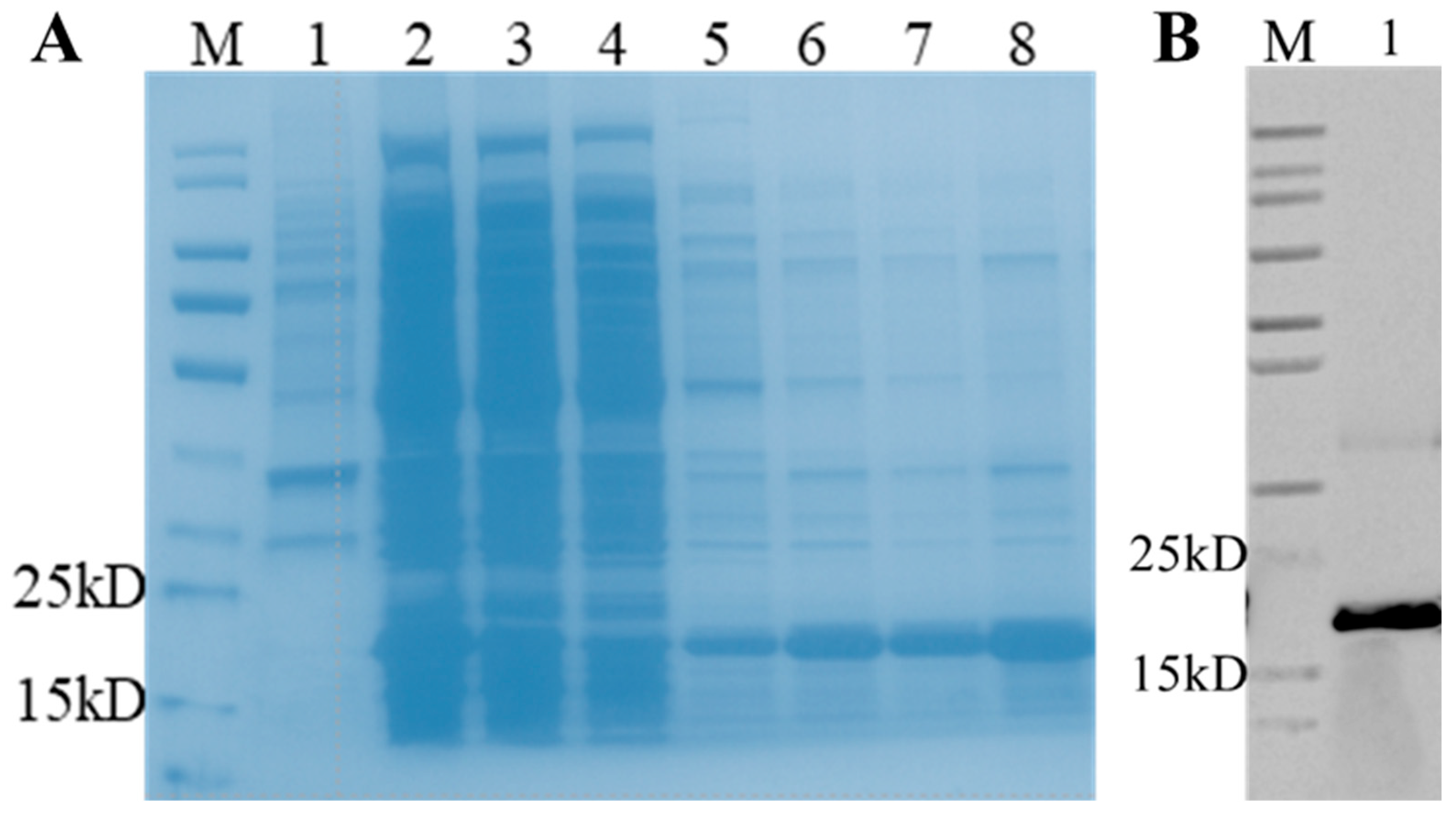
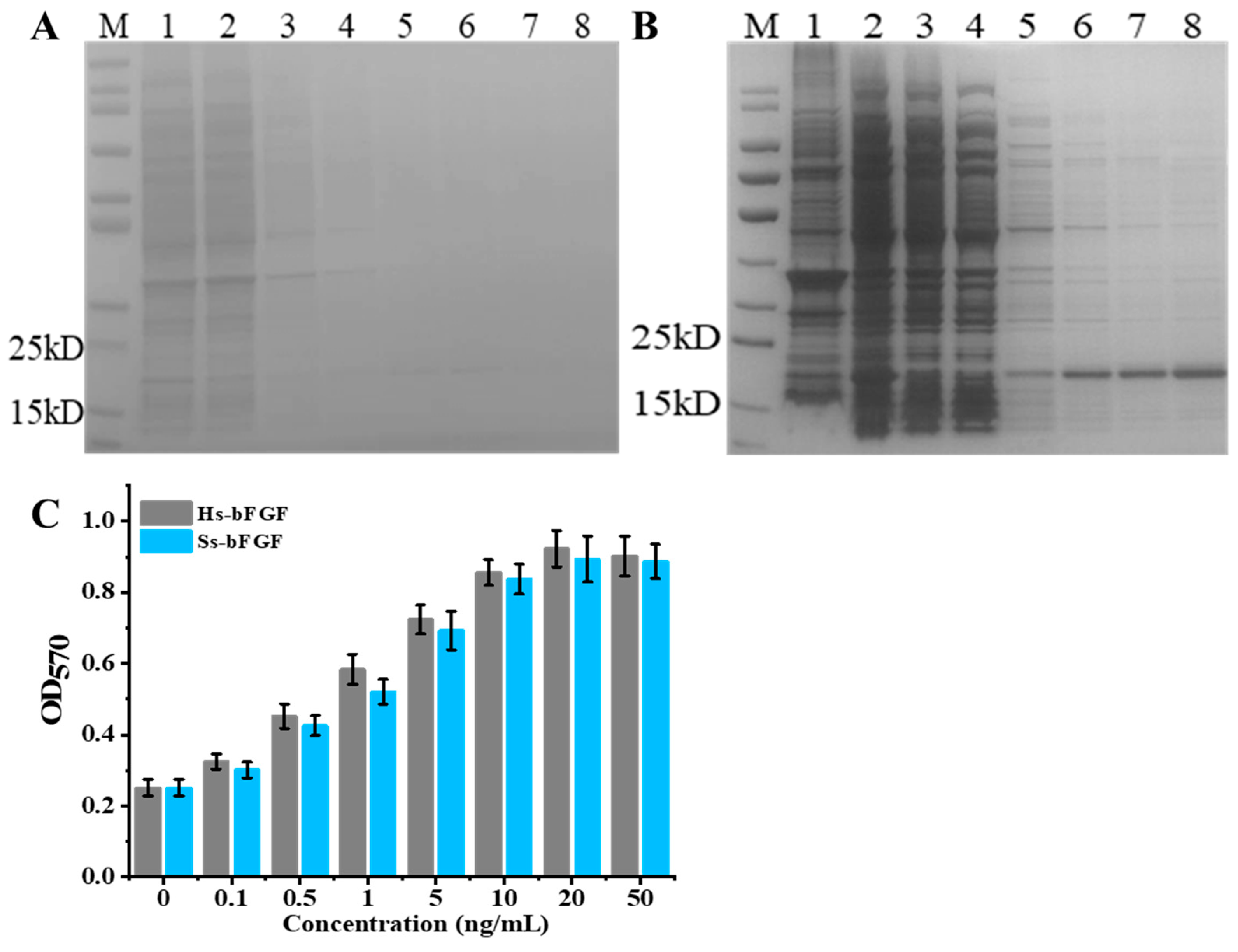
3.2. Expression and Purification of Recombinant Porcine bFGF
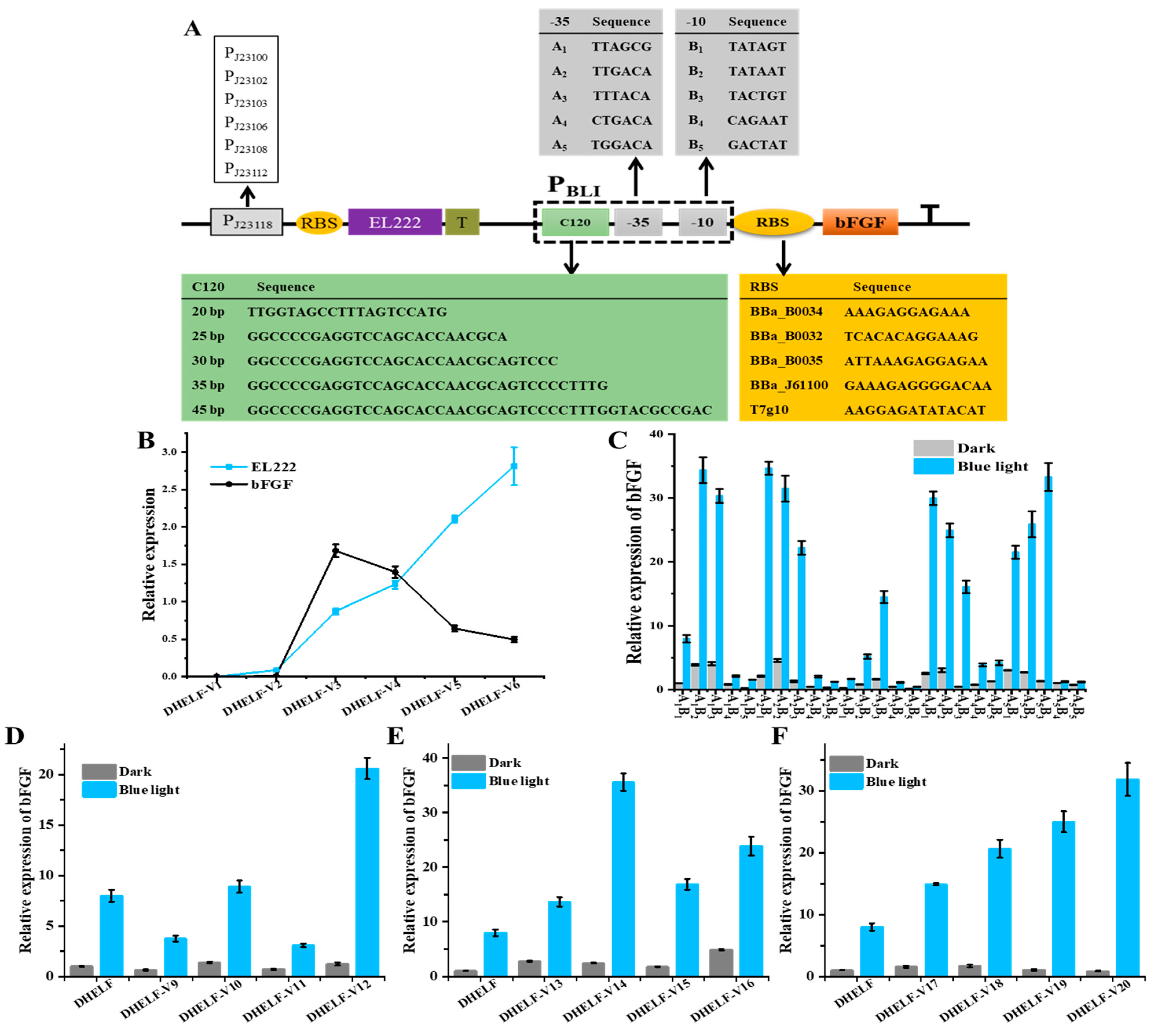
3.3. Optimization of EL222-Based Engineered Strain
3.4. Fermentation of Blue Light-Induced bFGF Expression in E. coli
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Stout, A.J.; Mirliani, A.B.; Rittenberg, M.L.; Shub, M.; White, E.C.; Yuen, J.S.K.; Kaplan, D.L. Simple and effective serum-free medium for sustained expansion of bovine satellite cells for cell cultured meat. Commun. Biol. 2022, 5, 466. [Google Scholar] [CrossRef] [PubMed]
- Chu, S.G.; Haraguchi, Y.; Asahi, T.; Kato, Y.; Kondo, A.; Hasunuma, T.; Shimizu, T. A serum-free culture medium production system by co-culture combining growth factor-secreting cells and L-lactate-assimilating cyanobacteria for sustainable cultured meat production. Sci. Rep. 2024, 14, 19578. [Google Scholar] [CrossRef] [PubMed]
- Yao, T.; Asayama, Y. Animal-cell culture media: History, characteristics, and current issues. Reprod. Med. Biol. 2017, 16, 99–117. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, E.N.; Cosenza, Z.A.; Baar, K.; Block, D.E. Considerations for the development of cost-effective cell culture media for cultivated meat production. Compr. Rev. Food Sci. Food Saf. 2021, 20, 686–709. [Google Scholar] [CrossRef] [PubMed]
- Hubalek, S.; Post, M.J.; Moutsatsou, P. Towards resource-efficient and cost-efficient cultured meat. Curr. Opin. Food Sci. 2022, 47, 100885. [Google Scholar] [CrossRef]
- Zhang, M.C.; Zhao, X.Y.; Li, Y.; Ye, Q.H.; Wu, Y.W.; Niu, Q.Y.; Zhang, Y.; Fan, G.H.; Chen, T.X.; Xia, J.R.; et al. Advances in serum-free media for CHO cells: From traditional serum substitutes to microbial-derived substances. Biotechnol. J. 2024, 19, e2400251. [Google Scholar] [CrossRef]
- Geng, S.L.; Zou, Y.; Bai, Z.Y.; Zhang, M.; Wang, C.; Wang, T.Y. Serum-free medium for recombinant protein expression in insect cells. Biotechnol. Appl. Biochem. 2024, 1–15. [Google Scholar] [CrossRef]
- Romero, S.G.; Boyle, N. Systems biology and metabolic modeling for cultivated meat: A promising approach for cell culture media optimization and cost reduction. Compr. Rev. Food Sci. Food Saf. 2023, 22, 3422–3443. [Google Scholar] [CrossRef]
- O’Flaherty, R.; Bergin, A.; Flampouri, E.; Mota, L.M.; Obaidi, I.; Quigley, A.; Xie, Y.J.; Butler, M. Mammalian cell culture for production of recombinant proteins: A review of the critical steps in their biomanufacturing. Biotechnol. Adv. 2020, 43, 107552. [Google Scholar] [CrossRef]
- Zhang, G.Q.; Zhao, X.R.; Li, X.L.; Du, G.C.; Zhou, J.W.; Chen, J. Challenges and possibilities for bio-manufacturing cultured meat. Trends Food Sci. Technol. 2020, 97, 443–450. [Google Scholar] [CrossRef]
- Kunert, R.; Reinhart, D. Advances in recombinant antibody manufacturing. Appl. Microbiol. Biotechnol. 2016, 100, 3451–3461. [Google Scholar] [CrossRef] [PubMed]
- He, B.; Lin, J.X.; Li, J.; Mi, Y.L.; Zeng, W.D.; Zhang, C.Q. Basic fibroblast growth factor suppresses meiosis and promotes mitosis of ovarian germ cells in embryonic chickens. Gen. Comp. Endocrinol. 2012, 176, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.H.; Zhang, G.W.; Gu, T.X.; Li-Ling, J.; Wen, T.; Zhao, Y.; Wang, C.; Fang, Q.; Yu, L.; Liu, B. Exogenous basic fibroblast growth factor promotes cardiac stem cell-mediated myocardial regeneration after miniswine acute myocardial infarction. Coron. Artery Dis. 2011, 22, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Nowwarote, N.; Sawangmake, C.; Pavasant, P.; Osathanon, T. Review of the role of basic fibroblast growth factor in dental tissue-derived mesenchymal stem cells. Asian Biomed. 2015, 9, 271–283. [Google Scholar]
- Ramasamy, R.; Tong, C.K.; Yip, W.K.; Vellasamy, S.; Tan, B.C.; Seow, H.F. Basic fibroblast growth factor modulates cell cycle of human umbilical cord-derived mesenchymal stem cells. Cell Prolif. 2012, 45, 132–139. [Google Scholar] [CrossRef]
- Garor, R.; Abir, R.; Erman, A.; Felz, C.; Nitke, S.; Fisch, B. Effects of basic fibroblast growth factor on in vitro development of human ovarian primordial follicles. Fertil. Steril. 2009, 91, 1967–1975. [Google Scholar] [CrossRef]
- de Araújo, R.; Lôbo, M.; Trindade, K.; Silva, D.F.; Pereira, N. Fibroblast Growth Factors: A Controlling Mechanism of Skin Aging. Skin Pharmacol. Physiol. 2019, 32, 275–282. [Google Scholar] [CrossRef]
- Kim, Y.S.; Lee, H.J.; Han, M.H.; Yoon, N.K.; Kim, Y.C.; Ahn, J. Effective production of human growth factors in Escherichia coli by fusing with small protein 6HFh8. Microb. Cell Fact. 2021, 20, 9. [Google Scholar] [CrossRef]
- Gasparian, M.E.; Elistratov, P.A.; Drize, N.I.; Nifontova, I.N.; Dolgikh, D.A.; Kirpichnikov, M.P. Overexpression in Escherichia coli and purification of human fibroblast growth factor (FGF-2). Biochemistry 2009, 74, 221–225. [Google Scholar] [CrossRef]
- Mu, X.P.; Kong, N.; Chen, W.L.; Zhang, T.; Shen, M.; Yan, W.Q. High-level expression, purification, and characterization of recombinant human basic fibroblast growth factor in Pichia pastoris. Protein Expr. Purif. 2008, 59, 282–288. [Google Scholar] [CrossRef]
- Imsoonthornruksa, S.; Pruksananonda, K.; Parnpai, R.; Rungsiwiwut, R.; Ketudat-Cairns, M. Expression and purification of recombinant human basic fibroblast growth factor fusion proteins and their uses in human stem cell culture. J. Mol. Microbiol. Biotechnol. 2015, 25, 372–380. [Google Scholar] [CrossRef] [PubMed]
- Soleyman, M.R.; Khalili, M.; Khansarinejad, B.; Baazm, M. High-level expression and purification of active human FGF-2 in Escherichia coli by codon and culture condition optimization. Iran. Red Crescent Med. J. 2016, 18, e21615. [Google Scholar] [CrossRef] [PubMed]
- Alibolandi, M.; Mirzahoseini, H.; Abad, M.A.K.; Movahed, M.A. High level expression of human basic fibroblast growth factor in Escherichia coli: Evaluating the effect of the GC content and rare codons within the first 13 codons. Afr. J. Biotechnol. 2010, 9, 2456–2462. [Google Scholar]
- Masuda, A.; Xu, J.; Minamihata, K.; Kagawa, G.; Hamada, Y.; Morifuji, Y.; Yano, T.; Hino, M.; Morokuma, D.; Karasaki, N.; et al. Production of a biologically active human basic fibroblast growth factor using silkworm-baculovirus expression vector system. J. Asia-Pacif. Entomol. 2018, 21, 716–720. [Google Scholar] [CrossRef]
- Kurokawa, T.; Sasada, R.; Iwane, M.; Igarashi, K. Cloning and expression of cDNA encoding human basic fibroblast growth factor. FEBS Lett. 1987, 213, 189–194. [Google Scholar] [CrossRef]
- Cheng, T.; Cao, W.; Wen, R.; Steinberg, R.H.; LaVail, M.M. Prostaglandin E2 induces vascular endothelial growth factor and basic fibroblast growth factor mRNA expression in cultured rat Müller cells. Investig. Ophthalmol. Vis. Sci. 1998, 39, 581–591. [Google Scholar]
- Tripathi, N.K.; Shrivastava, A. Recent developments in bioprocessing of recombinant proteins: Expression hosts and process development. Front. Bioeng. Biotechnol. 2019, 7, 420. [Google Scholar] [CrossRef]
- Karbalaei, M.; Rezaee, S.A.; Farsiani, H. Pichia pastoris: A highly successful expression system for optimal synthesis of heterologous proteins. J. Cell. Physiol. 2020, 235, 5867–5881. [Google Scholar] [CrossRef]
- Chia, N.; Lee, S.Y.; Tong, Y. Optogenetic tools for microbial synthetic biology. Biotechnol. Adv. 2022, 59, 107953. [Google Scholar] [CrossRef]
- Muller, K.; Naumann, S.; Weber, W.; Zurbriggen, M.D. Optogenetics for gene expression in mammalian cells. Biol. Chem. 2015, 396, 145–152. [Google Scholar] [CrossRef]
- Omelina, E.S.; Yushkova, A.A.; Motorina, D.M.; Volegov, G.A.; Kozhevnikova, E.N.; Pindyurin, A.V. Optogenetic and chemical induction systems for regulation of transgene expression in plants: Use in basic and applied research. Int. J. Mol. Sci. 2022, 23, 1737. [Google Scholar] [CrossRef] [PubMed]
- Lalwani, M.A.; Ip, S.S.; Carrasco-Lopez, C.; Day, C.; Zhao, E.M.; Kawabe, H.; Avalos, J.L. Optogenetic control of the lac operon for bacterial chemical and protein production. Nat. Chem. Biol. 2021, 17, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Pouzet, S.; Banderas, A.; Le Bec, M.; Lautier, T.; Truan, G.; Hersen, P. The promise of optogenetics for bioproduction: Dynamic control strategies and scale-up instruments. Bioengineering 2020, 7, 151. [Google Scholar] [CrossRef] [PubMed]
- Motta-Mena, L.B.; Reade, A.; Mallory, M.J.; Glantz, S.; Weiner, O.D.; Lynch, K.W.; Gardner, K.H. An optogenetic gene expression system with rapid activation and deactivation kinetics. Nat. Chem. Biol. 2014, 10, 196–202. [Google Scholar] [CrossRef]
- Fernandez-Rodriguez, J.; Moser, F.; Song, M.; Voigt, C.A. Engineering RGB color vision into Escherichia coli. Nat. Chem. Biol. 2017, 13, 706–708. [Google Scholar] [CrossRef]
- Hennemann, J.; Iwasaki, R.S.; Grund, T.N.; Diensthuber, R.P.; Richter, F.; Moglich, A. Optogenetic control by pulsed illumination. ChemBioChem 2018, 19, 1296–1304. [Google Scholar] [CrossRef]
- Emiliani, V.; Entcheva, E.; Hedrich, R.; Hegemann, P.; Konrad, K.R.; Luscher, C.; Mahn, M.; Pan, Z.H.; Sims, R.R.; Vierock, J.; et al. Optogenetics for light control of biological systems. Nat. Rev. Methods Primers 2022, 2, 55. [Google Scholar] [CrossRef]
- Soffer, G.; Perry, J.M.; Shih, S.C.C. Real-time optogenetics system for controlling gene expression using a model-based design. Anal. Chem. 2021, 93, 3181–3188. [Google Scholar] [CrossRef]
- Nash, A.I.; McNulty, R.; Shillito, M.E.; Swartz, T.E.; Bogomolni, R.A.; Luecke, H.; Gardner, K.H. Structural basis of photosensitivity in a bacterial light-oxygen-voltage/helix-turn-helix (LOV-HTH) DNA-binding protein. Proc. Natl. Acad. Sci. USA 2011, 108, 9449–9454. [Google Scholar] [CrossRef]
- Zoltowski, B.D.; Motta-Mena, L.B.; Gardner, K.H. Blue light-induced dimerization of a bacterial LOV-HTH DNA-binding protein. Biochemistry 2013, 52, 6653–6661. [Google Scholar] [CrossRef]
- Jayaraman, P.; Devarajan, K.; Chua, T.K.; Zhang, H.; Gunawan, E.; Poh, C.L. Blue light-mediated transcriptional activation and repression of gene expression in bacteria. Nucleic Acids Res. 2016, 44, 6994–7005. [Google Scholar] [CrossRef] [PubMed]
- Ohlendorf, R.; Möglich, A. Light-regulated gene expression in bacteria: Fundamentals, advances, and perspectives. Front. Bioeng. Biotechnol. 2022, 10, 1029403. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Geng, F.; Shen, J.; Zhu, P.; Lu, Z.; Lu, F.; Zhou, L. Blue light-mediated gene expression as a promising strategy to reduce antibiotic resistance in Escherichia coli. Biotechnol. J. 2024, 19, 2400023. [Google Scholar] [CrossRef] [PubMed]
- Lukaszewicz, A.; Savatier, P.; Cortay, V.; Kennedy, H.; Dehay, C. Contrasting effects of basic fibroblast growth factor and neurotrophin 3 on cell cycle kinetics of mouse cortical stem cells. J. Neurosci. 2002, 22, 6610–6622. [Google Scholar] [CrossRef]
- Hung, J.H.; Weng, Z. Sequence alignment and homology search with BLAST and ClustalW. Cold Spring Harb. Protoc. 2016, 2016, 1016–1021. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Hallgren, J.; Tsirigos, K.D.; Pedersen, M.D.; Almagro Armenteros, J.J.; Marcatili, P.; Nielsen, H.; Krogh, A.; Winther, O. DeepTMHMM predicts alpha and beta transmembrane proteins using deep neural networks. bioRxiv 2022, bioRxiv:2022.04.08.487609. [Google Scholar]
- Nielsen, H.; Teufel, F.; Brunak, S.; von Heijne, G. SignalP: The Evolution of a Web Server. Methods Mol. Biol. 2024, 2836, 331–367. [Google Scholar]
- Ding, Q.; Ma, D.; Liu, G.Q.; Li, Y.; Guo, L.; Gao, C.; Hu, G.; Ye, C.; Liu, J.; Liu, L.; et al. Light-powered Escherichia coli cell division for chemical production. Nat. Commun. 2020, 11, 2262. [Google Scholar] [CrossRef]
- Stohr, A.M.; Ma, D.R.; Chen, W.L.; Blenner, M. Engineering conditional protein-protein interactions for dynamic cellular control. Biotechnol. Adv. 2024, 77, 108457. [Google Scholar] [CrossRef]
- Li, M.; Chen, H.; Liu, C.; Guo, J.; Xu, X.; Zhang, H.; Nian, R.; Xian, M. Improvement of isoprene production in Escherichia coli by rational optimization of RBSs and key enzymes screening. Microb. Cell Fact. 2019, 18, 4. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meng, F.; Xu, Z.; Fan, X.; Wang, Z.; Zhou, L. Optogenetic Fine-Tuning of Sus scrofa Basic Fibroblast Growth Factor Expression in Escherichia coli. Fermentation 2024, 10, 612. https://doi.org/10.3390/fermentation10120612
Meng F, Xu Z, Fan X, Wang Z, Zhou L. Optogenetic Fine-Tuning of Sus scrofa Basic Fibroblast Growth Factor Expression in Escherichia coli. Fermentation. 2024; 10(12):612. https://doi.org/10.3390/fermentation10120612
Chicago/Turabian StyleMeng, Fanqiang, Zhimin Xu, Xia Fan, Zhisheng Wang, and Libang Zhou. 2024. "Optogenetic Fine-Tuning of Sus scrofa Basic Fibroblast Growth Factor Expression in Escherichia coli" Fermentation 10, no. 12: 612. https://doi.org/10.3390/fermentation10120612
APA StyleMeng, F., Xu, Z., Fan, X., Wang, Z., & Zhou, L. (2024). Optogenetic Fine-Tuning of Sus scrofa Basic Fibroblast Growth Factor Expression in Escherichia coli. Fermentation, 10(12), 612. https://doi.org/10.3390/fermentation10120612