
A Physiogenomic Study of the Tolerance of Saccharomyces cerevisiae to Isoamyl Alcohol
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strains
2.2. Media
2.3. Isoamyl Alcohol Tolerance Experiment
2.4. RNA-seq
2.5. Genetic Targets Verification
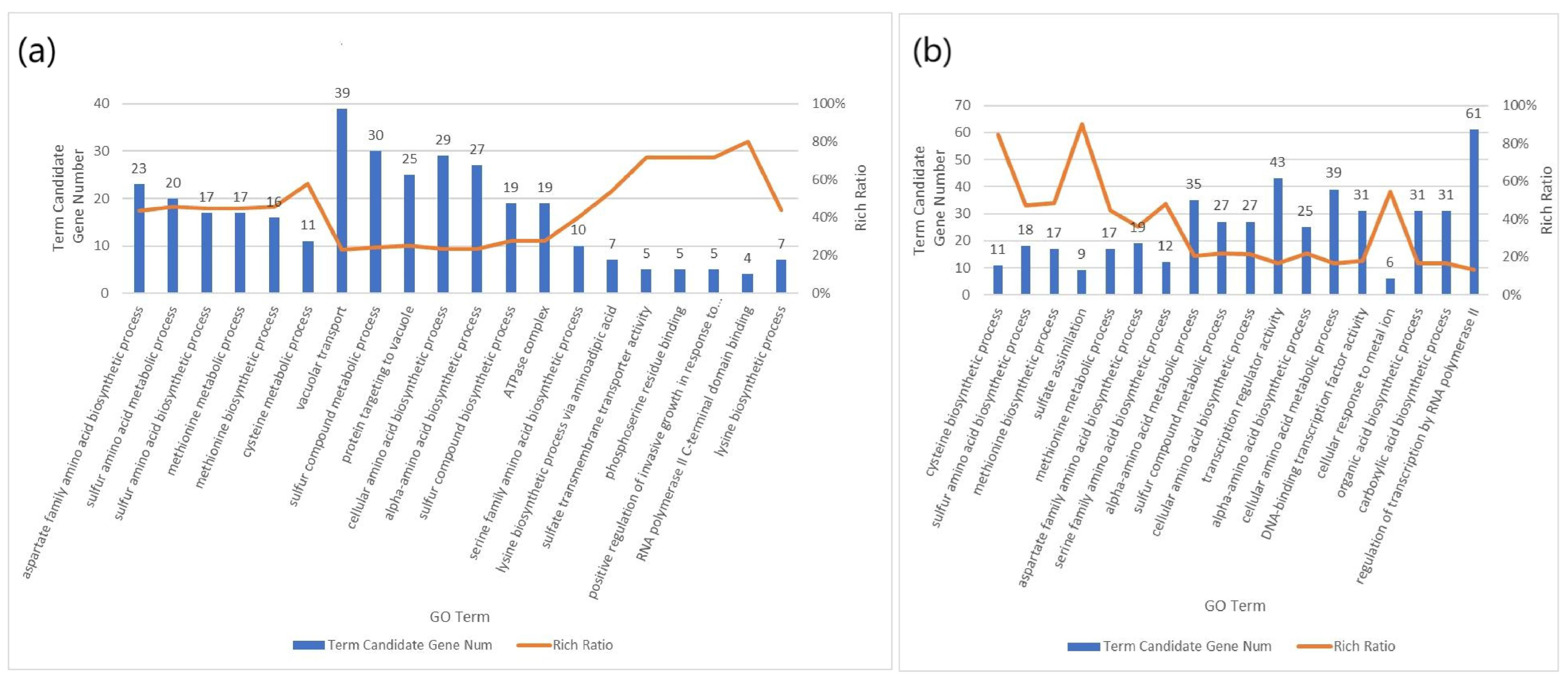
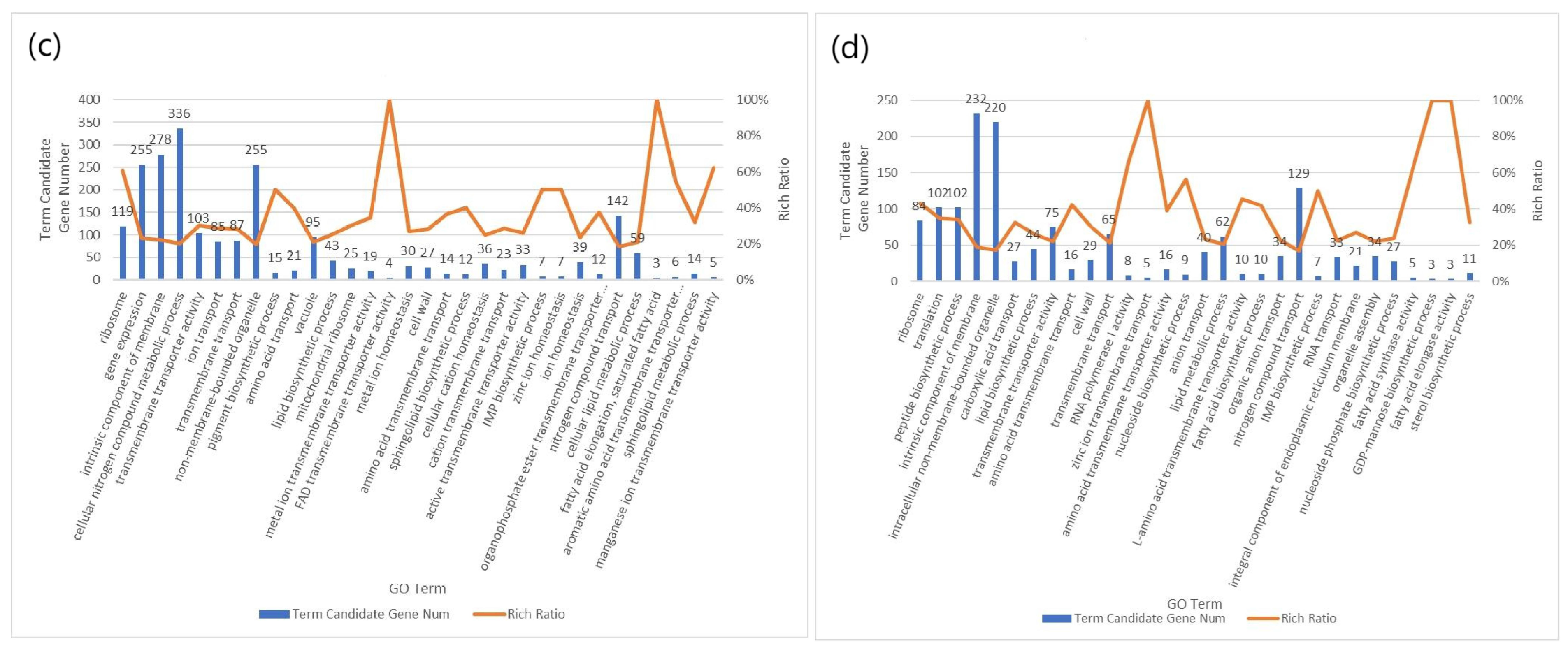
2.6. GO
3. Results and Discussion
3.1. Tolerance Test
3.2. Transcriptome Analysis
3.3. Cell Wall
3.4. Cell Membrane
3.5. Metal Ion Transporter
3.6. Mitochondrial Activity
3.7. Pentose Phosphate Pathway
3.8. Further Study on Potential Functional Genes
4. Conclusions and Perspectives
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McGinty, D.; Lapczynski, A.; Scognamiglio, J.; Letizia, C.S.; Api, A.M. Fragrance materials review on isoamyl alcohol. Food Chem. Toxicol. 2010, 48 (Suppl. S4), S102–S109. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Chen, X.; Mishra, P.; Ching, C.B. Metabolically engineered Saccharomyces cerevisiae for enhanced isoamyl alcohol production. Appl. Microbiol. Biotechnol. 2017, 101, 465–474. [Google Scholar] [CrossRef] [PubMed]
- Hazelwood, L.A.; Daran, J.M.; Maris, A.; Pronk, J.T.; Dickinson, J.R. The Ehrlich Pathway for Fusel Alcohol Production: A Century of Research on Saccharomyces cerevisiae Metabolism. Appl. Environ. Microbiol. 2008, 74, 2259–2266. [Google Scholar] [CrossRef]
- Atsumi, S.; Hanai, T.; Liao, J.C. Non-fermentative pathways for synthesis of branched-chain higher alcohols as biofuels. Nature 2008, 451, 86–89. [Google Scholar] [CrossRef] [PubMed]
- Dunlop, M.J. Engineering microbes for tolerance to next-generation biofuels. Biotechnol. Biofuels 2011, 4, 32. [Google Scholar] [CrossRef] [PubMed]
- Peralta-Yahya, P.P.; Keasling, J.D. Advanced biofuel production in microbes. Biotechnol. J. 2010, 5, 147–162. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Baidoo, E.E.K.; Amer, B.; Mukhopadhyay, A.; Adams, P.D.; Simmons, B.A.; Lee, T.S. Engineering Saccharomyces cerevisiae for isoprenol production. Metab. Eng. 2021, 64, 154–166. [Google Scholar] [CrossRef]
- Chen, X.; Nielsen, K.F.; Borodina, I.; Kielland-Brandt, M.C.; Karhumaa, K. Increased isobutanol production in Saccharomyces cerevisiae by overexpression of genes in valine metabolism. Biotechnol. Biofuels 2011, 4, 21. [Google Scholar] [CrossRef]
- Matsuda, F.; Ishii, J.; Kondo, T.; Ida, K.; Tezuka, H.; Kondo, A. Increased isobutanol production in Saccharomyces cerevisiae by eliminating competing pathways and resolving cofactor imbalance. Microb. Cell Factories 2013, 12, 119. [Google Scholar] [CrossRef]
- Yuan, J.; Mishra, P.; Ching, C.B. Engineering the leucine biosynthetic pathway for isoamyl alcohol overproduction in Saccharomyces cerevisiae. J. Ind. Microbiol. Biotechnol. 2017, 44, 107–117. [Google Scholar] [CrossRef]
- Kuroda, K.; Hammer, S.K.; Watanabe, Y.; Montaño López, J.; Fink, G.R.; Stephanopoulos, G.; Ueda, M.; Avalos, J.L. Critical Roles of the Pentose Phosphate Pathway and GLN3 in Isobutanol-Specific Tolerance in Yeast. Cell Syst. 2019, 9, 534–547.e5. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, M.; Kato, S.; Fukuda, S.; Izawa, S. Acquired Resistance to Severe Ethanol Stress in Saccharomyces cerevisiae Protein Quality Control. Appl. Environ. Microbiol. 2021, 87, e02353-20. [Google Scholar] [CrossRef] [PubMed]
- Jin, D.; Gu, B.; Xiong, D.; Huang, G.; Huang, X.; Liu, L.; Xiao, J. A Transcriptomic Analysis of Saccharomyces cerevisiae under the Stress of 2-Phenylethanol. Curr. Microbiol. 2018, 75, 1068–1076. [Google Scholar] [CrossRef] [PubMed]
- Gordon, M.R.; Zhu, J.; Qu, V.; Li, R. A case of convergent-gene interference in the budding yeast knockout library causing chromosome instability. G3 Genes Genomes Genet. 2021, 11, jkab084. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, H.C.; Rossi, S.A.; García-Barbazán, I.; Zaragoza, Ó.; Trevijano-Contador, N. Cell wall integrity pathway involved in morphogenesis, virulence and antifungal susceptibility in cryptococcus neoformans. J. Fungi 2021, 7, 831. [Google Scholar] [CrossRef]
- Taylor, E.J.; Campbell, S.G.; Griffiths, C.D.; Reid, P.J.; Slaven, J.W.; Harrison, R.J.; Sims, P.F.G.; Pavitt, G.D.; Delneri, D.; Ashe, M.P. Fusel alcohols regulate translation initiation by inhibiting eIF2B to reduce ternary complex in a mechanism that may involve altering the integrity and dynamics of the eIF2B body. Mol. Biol. Cell 2010, 21, 2202–2216. [Google Scholar] [CrossRef]
- Orlean, P. Architecture and biosynthesis of the Saccharomyces cerevisiae cell wall. Genetics 2012, 192, 775–818. [Google Scholar] [CrossRef]
- Elsztein, C.; de Lima, R.d.C.P.; de Barros Pita, W.; de Morais, M.A. NCW2, a Gene Involved in the Tolerance to Polyhexamethylene Biguanide (PHMB), May Help in the Organisation of β-1,3-Glucan Structure of Saccharomyces cerevisiae Cell Wall. Curr. Microbiol. 2016, 73, 341–345. [Google Scholar] [CrossRef]
- Elsztein, C.; de Menezes, J.A.S.; de Morais, M.A. Polyhexamethyl biguanide can eliminate contaminant yeasts from fuel-ethanol fermentation process. J. Ind. Microbiol. Biotechnol. 2008, 35, 967–973. [Google Scholar] [CrossRef]
- Qi, Y.; Xu, N.; Li, Z.; Wang, J.; Meng, X.; Gao, C.; Chen, J.; Chen, W.; Chen, X.; Liu, L. Mediator Engineering of Saccharomyces cerevisiae to Improve Multidimensional Stress Tolerance. Appl. Environ. Microbiol. 2022, 88, e01627-21. [Google Scholar] [CrossRef]
- Li, X.H.; Peng, Y.J.; Ding, J.L.; Feng, M.G.; Ying, S.H. A homologue of yeast acyl-coa synthetase faa1 contributes to cytomembrane functionality involved in development and virulence in the insect pathogenic fungus beauveria bassiana. Microb. Pathog. 2022, 164, 105419. [Google Scholar] [CrossRef] [PubMed]
- Ishmayana, S.; Kennedy, U.J.; Learmonth, R.P. Further investigation of relationships between membrane fluidity and ethanol tolerance in Saccharomyces cerevisiae. World J. Microbiol. Biotechnol. 2017, 33, 218. [Google Scholar] [CrossRef]
- Kwiatek, J.M.; Carman, G.M. Yeast phosphatidic acid phosphatase Pah1 hops and scoots along the membrane phospholipid bilayer. J. Lipid Res. 2020, 61, 1232–1243. [Google Scholar] [CrossRef] [PubMed]
- Ozturk, M.; Metin, M.; Altay, V.; de Filippis, L.; Ünal, B.T.; Khursheed, A.; Gul, A.; Hasanuzzaman, M.; Nahar, K.; Kawano, T.; et al. Molecular Biology of Cadmium Toxicity in Saccharomyces cerevisiae. Biol. Trace Elem. Res. 2021, 199, 4832–4846. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Fang, T.; Yan, H.; Jiang, L. The protein kinase Cmk2 negatively regulates the calcium/calcineurin signalling pathway and expression of calcium pump genes PMR1 and PMC1 in budding yeast. Cell Commun. Signal. 2019, 17, 7. [Google Scholar] [CrossRef] [PubMed]
- Capps, D.; Hunter, A.; Chiang, M.; Pracheil, T.; Liu, Z. Ubiquitin-Conjugating Enzymes Ubc1 and Ubc4 Mediate the Turnover of Hap4, a Master Regulator of Mitochondrial Biogenesis in Saccharomyces cerevisiae. Microorganisms 2022, 10, 2370. [Google Scholar] [CrossRef] [PubMed]
- Carrillo-Garmendia, A.; Martinez-Ortiz, C.; Canizal-Garcia, M.; González-Hernández, J.C.; Arvizu-Medrano, S.M.; Gracida, J.; Madrigal-Perez, L.A.; Regalado-Gonzalez, C. Cytotoxicity of quercetin is related to mitochondrial respiration impairment in Saccharomyces cerevisiae. Yeast 2022, 39, 617–628. [Google Scholar] [CrossRef]
- Matsumoto, T.; Osawa, T.; Taniguchi, H.; Saito, A.; Yamada, R.; Ogino, H. Mitochondrial expression of metabolic enzymes for improving carotenoid production in Saccharomyces cerevisiae. Biochem. Eng. J. 2022, 189, 108720. [Google Scholar] [CrossRef]
- Pandey, A.K.; Pain J, J.B.; Dancis, A.; Pain, D. Essential mitochondrial role in iron-sulfur cluster assembly of the cytoplasmic isopropylmalate isomerase Leu1 in Saccharomyces cerevisiae. Mitochondrion 2023, 69, 104–115. [Google Scholar] [CrossRef]
- Gaignard, P.; Menezes, M.; Schiff, M.; Bayot, A.; Rak, M.; Ogier De Baulny, H.; Su, C.H.; Gilleron, M.; Lombes, A.; Abida, H.; et al. Mutations in CYC1, Encoding Cytochrome c1 Subunit of Respiratory Chain Complex III, Cause Insulin-Responsive Hyperglycemia. Am. J. Hum. Genet. 2013, 93, 384–389. [Google Scholar] [CrossRef]
- Lin, Y.-Y.; Kee, P.E.; Ng, H.-S.; Lan, J.C.-W. Recovery efficiency of a hydrophilic ionic-liquid aqueous biphasic system for the primary purification of cytochrome c from simulated Saccharomyces cerevisiae fermentation broth. Process Biochem. 2020, 94, 110–115. [Google Scholar] [CrossRef]
- Partow, S.; Hyl, P.B.; Mahadevan, R. Synthetic rescue couples NADPH generation to metabolite overproduction in Saccharomyces cerevisiae. Metab. Eng. 2017, 43, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Cadière, A.; Ortiz-Julien, A.; Camarasa, C.; Dequin, S. Evolutionary engineered Saccharomyces cerevisiae wine yeast strains with increased in vivo flux through the pentose phosphate pathway. Metab. Eng. 2011, 13, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Gorsich, S.W.; Dien, B.S.; Nichols, N.N.; Slininger, P.J.; Liu, Z.L.; Skory, C.D. Tolerance to furfural-induced stress is associated with pentose phosphate pathway genes ZWF1, GND1, RPE1, and TKL1 in Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2006, 71, 339–349. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| YKO Lib | Strains | Background |
|---|---|---|
| BY4741 | BY4741, ccw12Δ, bgl2Δ, sun4Δ, elo1Δ, erg2Δ, opi3Δ, ncw2Δ, faa1Δ, ftr1Δ, fet3Δ, pmc1Δ, zwf1Δ, ino1Δ, lys12Δ, ade4Δ, ald6Δ, trm2Δ, fcy2Δ, eft1Δ, sul2Δ, izh2Δ, izh1Δ, btn2Δ, grx8Δ, pdc1Δ, met5Δ, pdc6Δ, gas1Δ, mae1Δ, jlp1Δ, bna2Δ, hmt1Δ, rps12Δ, spi1Δ, hsp150Δ, hsp26Δ, leu4Δ, leu9Δ, oac1Δ, zps1Δ, zno1Δ, utr2Δ, exg1Δ, scs7Δ, lac1Δ, zrt1Δ, are1Δ, vcx1Δ, ctr2Δ, nhx1Δ, mrs2Δ, gdt1Δ, tat1Δ, atx2Δ, hxt1Δ, gsf2Δ, tdh1Δ, alg8Δ, lpd1Δ, sdh1Δ, atg8Δ, mdh1Δ, sol3Δ, gnd1Δ. | BY family |
| Go-Terms | Difference Multiple Value Range | In Range from Cluster * |
|---|---|---|
| Mannose | CCW12, SRL1, TIP1, MNN2, PMT1, MNN11, PMT4, KTR1 | |
| CCW12, SRL1, TIP1, CIS3, MNN2, MNN11, PMT4, KTR1 | ||
| CCW12, SRL1, KTR1 | ||
| Chitin | UTR2, BGL2, CHS3, PCM1 | |
| UTR2, BGL2 | ||
| CRZ1, CHS1 | ||
| Glucan | BGL2, SUN4, KNH1, EXG1, FKS1, SCW10, NCW2 | |
| BGL2, SUN4, KNH1, EXG1, FKS1, SCW10 |
| Go-Terms | Difference Multiple Value Range | In Range from Cluster * |
|---|---|---|
| Fatty acid | ELO1, YAT1, ELO2, TSC13, ACP1, ELO3, IZH2, ERG10, AIM45, OLE1, FAT1, OAR1, FAA4, SCS7 | |
| RTG1,FAA1, IDP3 | ||
| YAT1, ELO2, TSC13, OLE1, ELO1, ACP1, ELO3, IZH2, ERG10, AIM45 | ||
| RTG1, FAA1 | ||
| Ergosterol | ERG4, ERG25, NCP1, ERG3, HMG1, ERG13, ERG2, ERG10, CYB5, ARE1, ERG11 | |
| GRE2, ARE2 | ||
| ERG4, ERG25, NCP1, ERG3, HMG1, ERG13, ERG2, ERG10, CYB5 | ||
| GRE2 | ||
| Phospholipid | ALG7, PER1, SLC1, GPI11, SPF1, SCS2, SCS3, GPI10, EGD2, OPI3, AUR1, ARV1, GAB1, HMG1, ERG13, PLB2, ANY1, CDC33, ARF3, PIS1, IPT1, AGE1, ATG27, SEC59, SCS7, OPT2 | |
| SWH1, ATG8, ATG20, IVY1, VAM7, BBC1, PDR16, MET22, VPS30, INO1 | ||
| ALG7, PER1, SLC1, GPI11, SPF1, SCS2, SCS3, GPI10, EGD2, OPI3, AUR1, ARV1, GAB1, HMG1, ERG13, PLB2, ANY1, CDC33, ARF3, PIS1 | ||
| SWH1, ATG8, ATG20, IVY1, VAM7, BBC1, PDR16, MET22, VPS30 |
| Go-Terms | Difference Multiple Value Range | In Range from Cluster * |
|---|---|---|
| Calcium ion transport | FTR1, FET3, SIT1, FLC2 | |
| PMC1 | ||
| FLC2, GDT1, VCX1, SPF1 | ||
| PMC1 | ||
| Vacuolar homeostasis | GDT1, PER1, VCX1, SIT1, VMA16, ZRT3, EMP70, PFK2, ZRC1, CTR2, TMN2, NHX1, VMA3, TMN3, FET5, SMF2, YHC3, VNX1, ENB1 | |
| PMC1, VPS52, VMA22, GEX1, VPS3 | ||
| GDT1, PER1, VCX1, SIT1, VMA16, CTR2, ZRT3, EMP70, PFK2, ZRC1 | ||
| PMC1, VPS52, VMA22, GEX1, MNR2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, J.; Wang, Y.; Xu, H.; Liu, J.; Wang, J.; Zhang, H.; Nie, C. A Physiogenomic Study of the Tolerance of Saccharomyces cerevisiae to Isoamyl Alcohol. Fermentation 2024, 10, 4. https://doi.org/10.3390/fermentation10010004
Song J, Wang Y, Xu H, Liu J, Wang J, Zhang H, Nie C. A Physiogenomic Study of the Tolerance of Saccharomyces cerevisiae to Isoamyl Alcohol. Fermentation. 2024; 10(1):4. https://doi.org/10.3390/fermentation10010004
Chicago/Turabian StyleSong, Jialin, Yu Wang, Hengyuan Xu, Jinshang Liu, Jianping Wang, Haojun Zhang, and Cong Nie. 2024. "A Physiogenomic Study of the Tolerance of Saccharomyces cerevisiae to Isoamyl Alcohol" Fermentation 10, no. 1: 4. https://doi.org/10.3390/fermentation10010004
APA StyleSong, J., Wang, Y., Xu, H., Liu, J., Wang, J., Zhang, H., & Nie, C. (2024). A Physiogenomic Study of the Tolerance of Saccharomyces cerevisiae to Isoamyl Alcohol. Fermentation, 10(1), 4. https://doi.org/10.3390/fermentation10010004