
Assessment of Fibrinogen Thermal Conductivity and Interaction Energy with Polyarylethersulfone (PAES) Clinical Hemodialysis Membranes at Normal and Elevated Patient Body Temperatures



Abstract
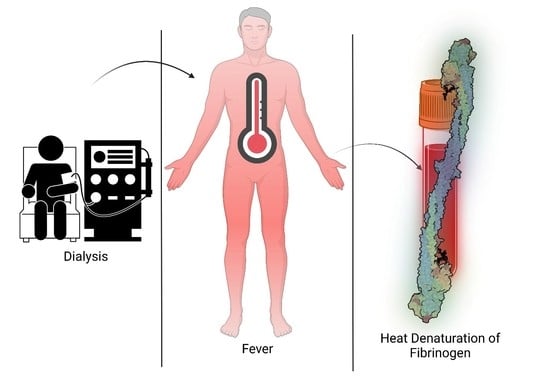
1. Introduction
2. Computational Methods
3. Results and Discussion
3.1. Thermal Conductivity
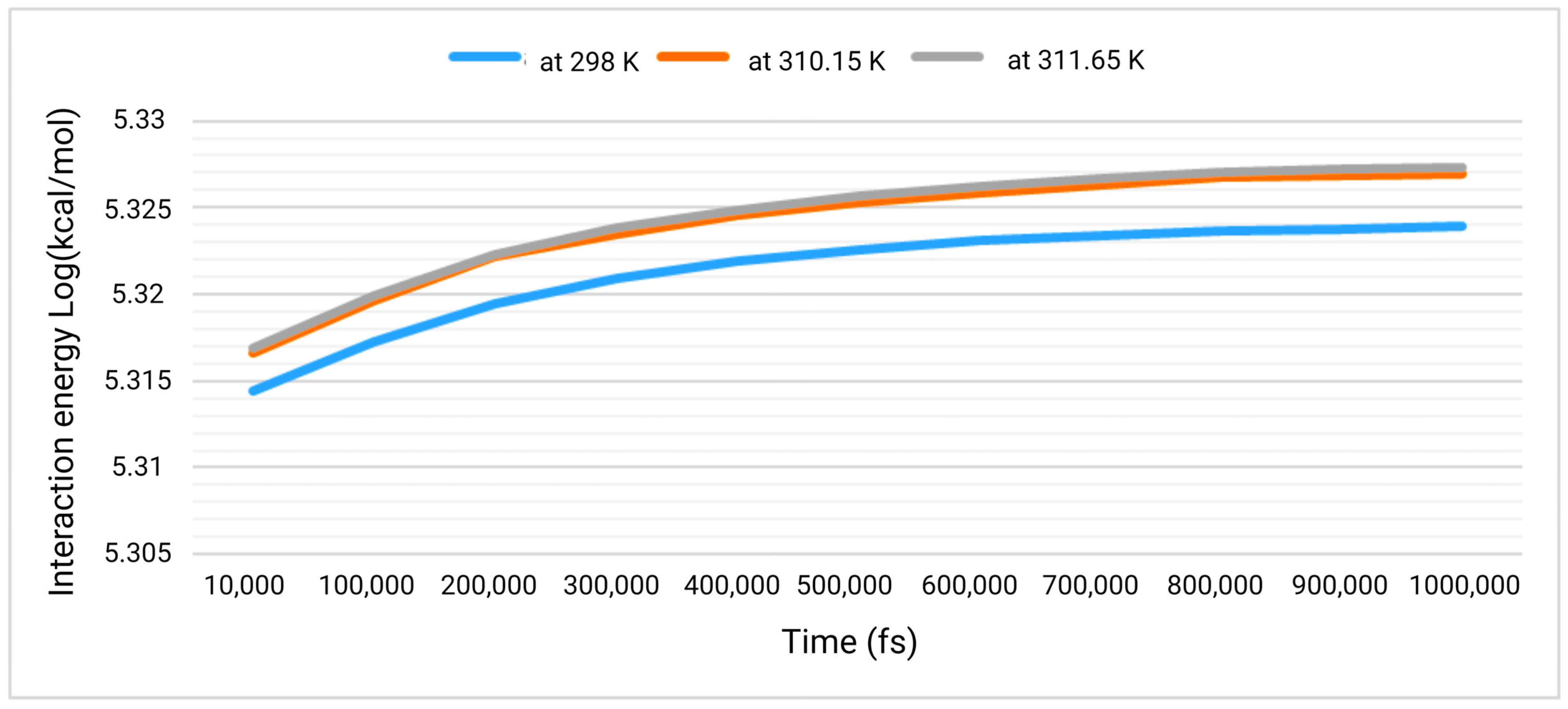
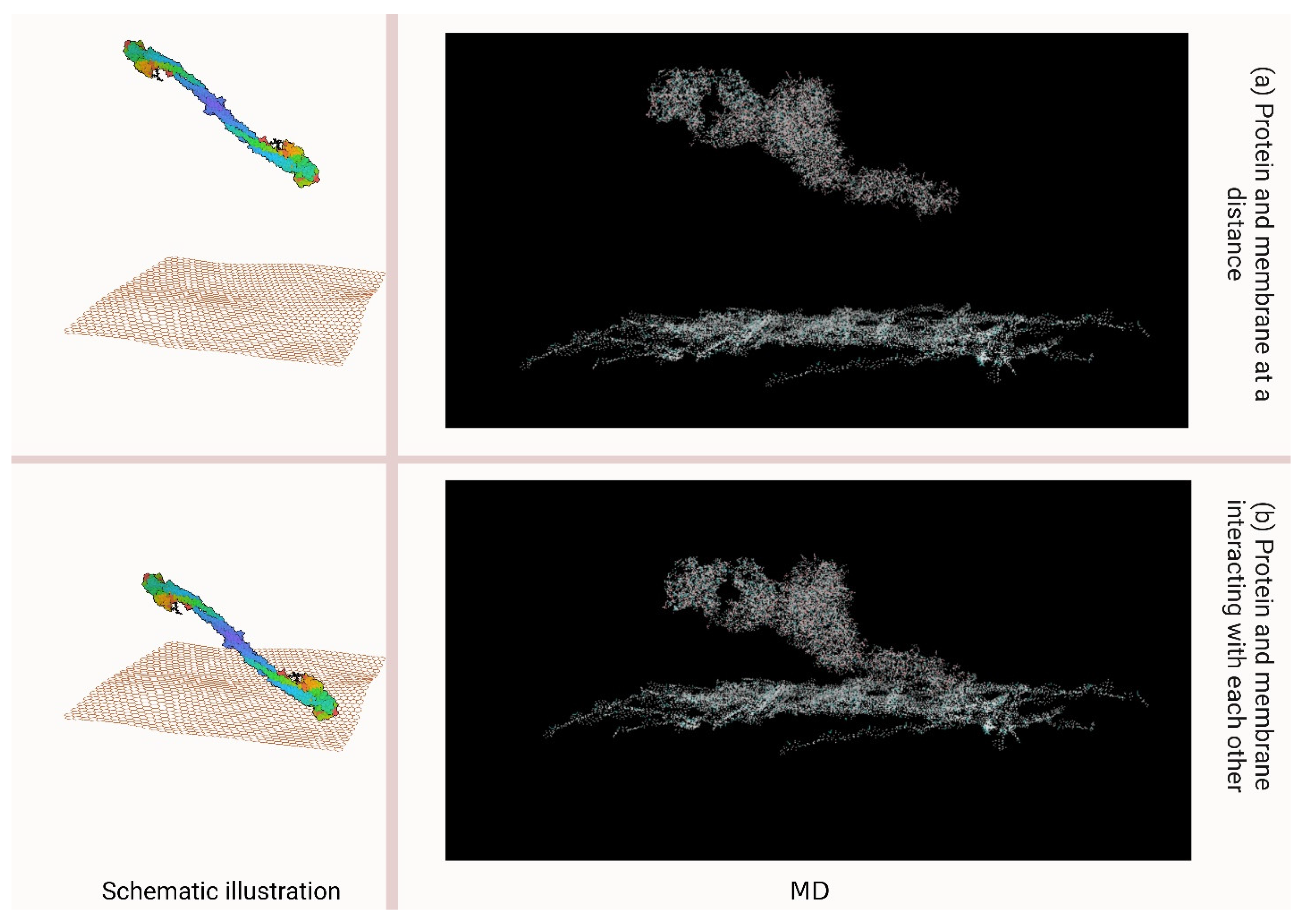
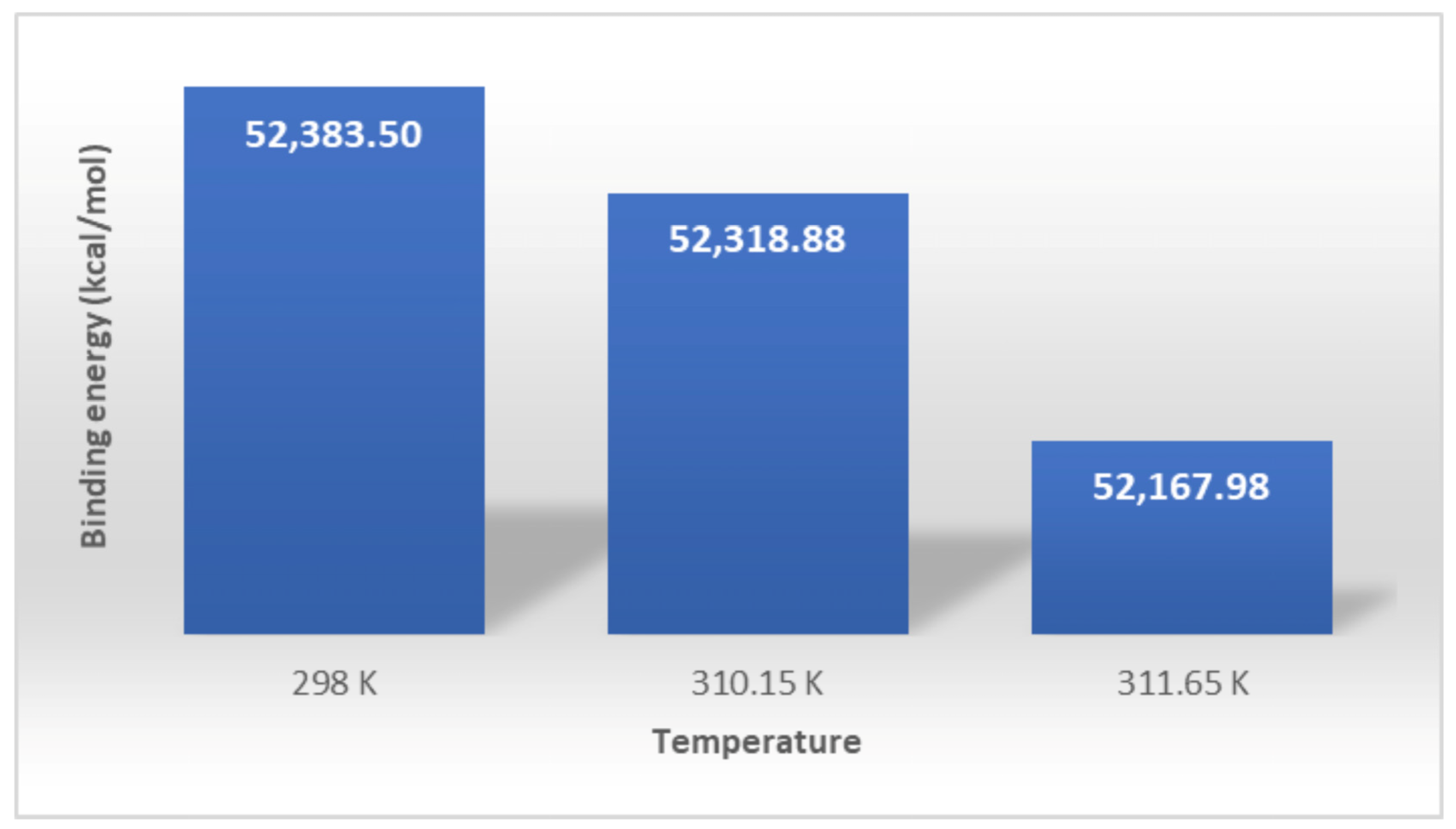
3.2. Interaction Studies
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Code Availability
Conflicts of Interest
Abbreviations
| Name | Acronyms |
| Chronic kidney disease | CKD |
| End-stage renal disease | ESRD |
| Fibrinogen | FB |
| Hemodialysis | HD |
| Human serum albumin | HSA |
| Human serum protein | HSP |
| Human serum transferrin | TRF |
| Molecular dynamics simulation | MD |
| Polyarylethersulfone | PAES |
| Polysulfone | PSF |
| Thermal conductivity | TC |
| van der Waals | VDW |
References
- Choi, S.R.; Lee, Y.-K.; Cho, A.J.; Park, H.C.; Han, C.H.; Choi, M.-J.; Koo, J.-R.; Yoon, J.-W.; Noh, J.W. Malnutrition, inflammation, progression of vascular calcification and survival: Inter-relationships in hemodialysis patients. PLoS ONE 2019, 14, e0216415. [Google Scholar] [CrossRef] [PubMed]
- Tong, J.; Liu, M.; Li, H.; Luo, Z.; Zhong, X.; Huang, J.; Liu, R.; He, F.; Fu, J. Mortality and associated risk factors in dialysis patients with cardiovascular disease. Kidney Blood Press. Res. 2016, 41, 479–487. [Google Scholar] [CrossRef]
- Sarnak, M.J. Cardiovascular complications in chronic kidney disease. Am. J. Kidney Dis. 2003, 41, 11–17. [Google Scholar] [CrossRef]
- Inrig, J.K. Intradialytic hypertension: A less-recognized cardiovascular complication of hemodialysis. Am. J. Kidney Dis. 2010, 55, 580–589. [Google Scholar] [CrossRef]
- Thomas, R.; Kanso, A.; Sedor, J. Chronic kidney disease and its complications. Prim. Care Clin. Off. Pract. 2008, 35, 329–344. [Google Scholar] [CrossRef] [PubMed]
- Mollahosseini, A.; Abdelrasoul, A.; Shoker, A. A critical review of recent advances in hemodialysis membranes hemocompatibility and guidelines for future development. Mater. Chem. Phys. 2020, 248, 122911. [Google Scholar] [CrossRef]
- Mollahosseini, A.; Abdelrasoul, A.; Shoker, A. Challenges and advances in hemodialysis membranes. Adv. Membr. Technol. 2020, 151. [Google Scholar]
- Mollahosseini, A.; Abdelrasoul, A.; Shoker, A. Latest advances in zwitterionic structures modified dialysis membranes. Mater. Today Chem. 2020, 15, 100227. [Google Scholar] [CrossRef]
- Khoshhal Salestan, S.; Seyedpour, S.F.; Rahimpour, A.; Shamsabadi, A.A.; Tiraferri, A.; Soroush, M. Molecular Dynamics Insights into the Structural and Water Transport Properties of a Forward Osmosis Polyamide Thin Film Nanocomposite Membrane Modified with Graphene Quantum Dots. Ind. Eng. Chem. Res. 2020, 59, 14447–14457. [Google Scholar] [CrossRef]
- Karimipour, A.; Karimipour, A.; Jolfaei, N.A.; Hekmatifar, M.; Toghraie, D.; Sabetvand, R.; Rostami, S. Prediction of the interaction between HIV viruses and human serum albumin (HSA) molecules using an equilibrium dynamics simulation program for application in bio medical science. J. Mol. Liq. 2020, 318, 113989. [Google Scholar] [CrossRef]
- Durrant, J.D.; McCammon, J.A. Molecular dynamics simulations and drug discovery. BMC Biol. 2011, 9, 71. [Google Scholar] [CrossRef] [PubMed]
- Lemkul, J.A.; Bevan, D.R. Assessing the stability of Alzheimer’s amyloid protofibrils using molecular dynamics. J. Phys. Chem. B 2010, 114, 1652–1660. [Google Scholar] [CrossRef] [PubMed]
- Noorian, H.; Toghraie, D.; Azimian, A. The effects of surface roughness geometry of flow undergoing Poiseuille flow by molecular dynamics simulation. Heat Mass Transf. 2014, 50, 95–104. [Google Scholar] [CrossRef]
- Toghraie, D.; Mokhtari, M.; Afrand, M. Molecular dynamic simulation of copper and platinum nanoparticles Poiseuille flow in a nanochannels. Phys. E Low-Dimens. Syst. Nanostructures 2016, 84, 152–161. [Google Scholar] [CrossRef]
- Afrouzi, H.H.; Ahmadian, M.; Moshfegh, A.; Toghraie, D.; Javadzadegan, A. Statistical analysis of pulsating non-Newtonian flow in a corrugated channel using Lattice-Boltzmann method. Phys. A Stat. Mech. Its Appl. 2019, 535, 122486. [Google Scholar] [CrossRef]
- Alrashed, A.A.; Akbari, O.A.; Heydari, A.; Toghraie, D.; Zarringhalam, M.; Shabani, G.A.S.; Seifi, A.R.; Goodarzi, M. The numerical modeling of water/FMWCNT nanofluid flow and heat transfer in a backward-facing contracting channel. Phys. B Condens. Matter 2018, 537, 176–183. [Google Scholar] [CrossRef]
- Sasongko, N.; Siahaan, P.; Lusiana, R.A.; Prasasty, V. Understanding the interaction of polysulfone with urea and creatinine at the molecular level and its application for hemodialysis membrane. In Journal of Physics: Conference Series; IOP Publishing: Bristol, UK, 2020. [Google Scholar]
- Mollahosseini, A.; Argumeedi, S.; Abdelrasoul, A.; Shoker, A. A case study of poly (aryl ether sulfone) hemodialysis membrane interactions with human blood: Molecular dynamics simulation and experimental analyses. Comput. Methods Programs Biomed. 2020, 197, 105742. [Google Scholar] [CrossRef]
- Mollahosseini, A.; Saadati, S.; Abdelrasoul, A. Effects of mussel-inspired co-deposition of 2-hydroxymethyl methacrylate and poly (2-methoxyethyl acrylate) on the hydrophilicity and binding tendency of common hemodialysis membranes: Molecular dynamics simulations and molecular docking studies. J. Comput. Chem. 2022, 43, 57–73. [Google Scholar] [CrossRef]
- Mollahosseini, A.; Lee, K.M.; Abdelrasoul, A.; Doan, H.; Zhu, N. Innovative in situ investigations using synchrotron-based micro tomography and molecular dynamics simulation for fouling assessment in ceramic membranes for dairy and food industry. Int. J. Appl. Ceram. Technol. 2021, 18, 2143–2157. [Google Scholar] [CrossRef]
- Mollahosseini, A.; Saadati, S.; Abdelrasoul, A. A Comparative Assessment of Human Serum Proteins Interactions with Hemodialysis Clinical Membranes using Molecular Dynamics Simulation. Macromol. Theory Simul. 2022, 31, 2200016. [Google Scholar] [CrossRef]
- Mollahosseini, A.; Abdelrasoul, A. Molecular dynamics simulation for membrane separation and porous materials: A current state of art review. J. Mol. Graph. Model. 2021, 107, 107947. [Google Scholar] [CrossRef]
- Mollahosseini, A.; Abdelrasoul, A. Novel insights in hemodialysis: Most Recent theories on the membrane hemocompatibility improvement. Biomed. Eng. Adv. 2022, 3, 100034. [Google Scholar] [CrossRef]
- Dobson, C.M. Protein folding and disease: A view from the first Horizon Symposium. Nat. Rev. Drug Discov. 2003, 2, 154–160. [Google Scholar] [CrossRef]
- Goto, Y.; Adachi, M.; Muta, H.; So, M. Salt-induced formations of partially folded intermediates and amyloid fibrils suggests a common underlying mechanism. Biophys. Rev. 2018, 10, 493–502. [Google Scholar] [CrossRef]
- Ashkezari, A.Z.; Jolfaei, N.A.; Hekmatifar, M.; Toghraie, D.; Sabetvand, R.; Rostami, S. Calculation of the thermal conductivity of human serum albumin (HSA) with equilibrium/non-equilibrium molecular dynamics approaches. Comput. Methods Programs Biomed. 2020, 188, 105256. [Google Scholar] [CrossRef] [PubMed]
- Goodship, T.H. Fibrinogen in hemodialysis: The worst of both worlds? Kidney Int. 2003, 63, 379–380. [Google Scholar] [CrossRef]
- Cacciafesta, P.; Humphris, A.D.L.; Jandt, K.D.; Miles, M.J. Human plasma fibrinogen adsorption on ultraflat titanium oxide surfaces studied with atomic force microscopy. Langmuir 2000, 16, 8167–8175. [Google Scholar] [CrossRef]
- Werner, C.; Maitz, M.; Sperling, C. Current strategies towards hemocompatible coatings. J. Mater. Chem. 2007, 17, 3376–3384. [Google Scholar] [CrossRef]
- Bergström, J.; Lindholm, B. What are the causes and consequences of the chronic inflammatory state in chronic dialysis patients? In Seminars in Dialysis; Blackwell Science Inc.: Boston, MA, USA, 2000. [Google Scholar]
- Goicoechea, M.; De Vinuesa, S.G.; Gomez-Campderá, F.; Aragoncillo, I.; Verdalles, U.; Mosse, A.; Luño, J. Serum fibrinogen levels are an independent predictor of mortality in patients with chronic kidney disease (CKD) stages 3 and 4: New strategies to prevent cardiovascular risk in chronic kidney disease. Kidney Int. 2008, 74, S67–S70. [Google Scholar] [CrossRef]
- Yu, J.; Lin, T.; Huang, N.; Xia, X.; Li, J.; Qiu, Y.; Yang, X.; Mao, H.; Huang, F. Plasma fibrinogen and mortality in patients undergoing peritoneal dialysis: A prospective cohort study. BMC Nephrol. 2020, 21, 349. [Google Scholar] [CrossRef] [PubMed]
- Botan, V.; Backus, E.H.; Pfister, R.; Moretto, A.; Crisma, M.; Toniolo, C.; Nguyen, P.H.; Stock, G.; Hamm, P. Energy transport in peptide helices. Proc. Natl. Acad. Sci. USA 2007, 104, 12749–12754. [Google Scholar] [CrossRef]
- Yamato, T.; Wang, T.; Sugiura, W.; Laprévote, O.; Katagiri, T. Computational Study on the Thermal Conductivity of a Protein. J. Phys. Chem. B 2022, 126, 3029–3036. [Google Scholar] [CrossRef]
- Peng, H.; Dang, L.; Toghraie, D. Molecular dynamics simulation of thermal characteristics of globulin protein dissolved in dilute salt solutions using equilibrium and non-equilibrium methods. J. Therm. Biol. 2023, 113, 103505. [Google Scholar] [CrossRef]
- Fang, Y.; Bokov, D.O.; Hachem, K.; Sabetvand, R.; Alsultany, F.H.; Suksatan, W.; Hekmatifar, M.; Toghraie, D. The computational investigation of thermal conductivity of 11S globulin protein for biological applications: Molecular dynamics simulation. J. Mol. Liq. 2022, 346, 118267. [Google Scholar] [CrossRef]
- Zhang, L.; Bai, Z.; Ban, H.; Liu, L. Effects of the amino acid sequence on thermal conduction through β-sheet crystals of natural silk protein. Phys. Chem. Chem. Phys. 2015, 17, 29007–29013. [Google Scholar] [CrossRef] [PubMed]
- Mayo, S.L.; Olafson, B.; Goddard, W. DREIDING: A generic force field for molecular simulations. J. Phys. Chem. 1990, 94, 8897–8909. [Google Scholar] [CrossRef]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Hanwell, M.D.; Hanwell, M.D.; Curtis, D.E.; Lonie, D.C.; Vandermeersch, T.; Zurek, E.; Hutchison, G.R. Avogadro: An advanced semantic chemical editor, visualization, and analysis platform. J. Cheminform. 2012, 4, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Martínez, L.; Andrade, R.; Birgin, E.G.; Martínez, J.M. PACKMOL: A package for building initial configurations for molecular dynamics simulations. J. Comput. Chem. 2009, 30, 2157–2164. [Google Scholar] [CrossRef]
- Wypych, G. Handbook of Polymers; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar]
- Mollahosseini, A.; Abdelrasoul, A. Zwitterionization of common hemodialysis membranes: Assessment of different immobilized structure impact on hydrophilicity and biocompatibility of poly aryl ether sulfone (PAES) and cellulose triacetate (CTA) hemodialysis membranes. Struct. Chem. 2022, 33, 1965–1982. [Google Scholar] [CrossRef]
- Jolfaei, N.A.; Hekmatifar, M.; Piranfar, A.; Toghraie, D.; Sabetvand, R.; Rostami, S. Investigation of thermal properties of DNA structure with precise atomic arrangement via equilibrium and non-equilibrium molecular dynamics approaches. Comput. Methods Programs Biomed. 2020, 185, 105169. [Google Scholar] [CrossRef]
- Ponder, E. The coefficient of thermal conductivity of blood and of various tissues. J. Gen. Physiol. 1962, 45, 545–551. [Google Scholar] [CrossRef]
- Tekin, I.O.; Pocan, B.; Borazan, A.; Ucar, E.; Kuvandik, G.; Ilikhan, S.; Demircan, N.; Ozer, C.; Kadayifci, S. Positive correlation of CRP and fibrinogen levels as cardiovascular risk factors in early stage of continuous ambulatory peritoneal dialysis patients. Ren. Fail. 2008, 30, 219–225. [Google Scholar] [CrossRef]
- Prinsen, B.H.; Rabelink, T.J.; Beutler, J.J.; Kaysen, G.A.; De Boer, J.; Boer, W.H.; Hagen, E.C.; Berger, R.; Velden, M.G.D.S.-V.D. Increased albumin and fibrinogen synthesis rate in patients with chronic renal failure. Kidney Int. 2003, 64, 1495–1504. [Google Scholar] [CrossRef] [PubMed]
- Giordano, M.; DE Feo, P.; Lucidi, P.; Depascale, E.; Giordano, G.; Infantone, L.; Zoccolo, A.M.; Castellino, P. Increased albumin and fibrinogen synthesis in hemodialysis patients with normal nutritional status. J. Am. Soc. Nephrol. 2001, 12, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Zoccali, C.; Mallamaci, F.; Tripepi, G.; Cutrupi, S.; Parlongo, S.; Malatino, L.S.; Bonanno, G.; Rapisarda, F.A.; Fatuzzo, P.M.; Seminara, G.; et al. Fibrinogen, mortality and incident cardiovascular complications in end-stage renal failure. J. Intern. Med. 2003, 254, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Fumagall, G.; Panichi, V. Biocompatibility of the Dialysis System. In Critical Care Nephrology; Elsevier: Amsterdam, The Netherlands, 2019; pp. 918–922. e2. [Google Scholar]
- Chidambaram, T.; Oren, Y.; Noel, M. Fouling of nanofiltration membranes by dyes during brine recovery from textile dye bath wastewater. Chem. Eng. J. 2015, 262, 156–168. [Google Scholar] [CrossRef]
- Ma, B.; Wu, G.; Li, W.; Miao, R.; Li, X.; Wang, P. Roles of membrane–foulant and inter/intrafoulant species interaction forces in combined fouling of an ultrafiltration membrane. Sci. Total Environ. 2019, 652, 19–26. [Google Scholar] [CrossRef]
- Khan, H.M.; He, T.; Fuglebakk, E.; Grauffel, C.; Yang, B.; Roberts, M.F.; Gershenson, A.; Reuter, N. A role for weak electrostatic interactions in peripheral membrane protein binding. Biophys. J. 2016, 110, 1367–1378. [Google Scholar] [CrossRef]
- Zhao, J.; He, G.; Liu, G.; Pan, F.; Wu, H.; Jin, W.; Jiang, Z. Manipulation of interactions at membrane interfaces for energy and environmental applications. Prog. Polym. Sci. 2018, 80, 125–152. [Google Scholar] [CrossRef]
- Harano, Y.; Yoshidome, T.; Kinoshita, M. Molecular mechanism of pressure denaturation of proteins. J. Chem. Phys. 2008, 129, 10B607. [Google Scholar] [CrossRef] [PubMed]
- Durand, D.; Gimel, J.; Nicolai, T. Aggregation, gelation and phase separation of heat denatured globular proteins. Phys. A Stat. Mech. Its Appl. 2002, 304, 253–265. [Google Scholar] [CrossRef]
- Rabe, M.; Verdes, D.; Seeger, S. Understanding protein adsorption phenomena at solid surfaces. Adv. Colloid Interface Sci. 2011, 162, 87–106. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Temperature (K) | EMD (W/m·K) | NEMD (W/m·K) |
|---|---|---|
| 310.15 | 0.044 | 0.084 |
| 311.65 | 0.577 | 0.114 |
| Polymer–Protein Interaction | Total Energy (kcal/mol) | van der Waals Interaction (kcal/mol) | Electrostatic Interaction (kcal/mol) | Hydrogen Bonding (kcal/mol) | Number of Hydrogen Bonds | Binding Energy (kcal/mol) |
|---|---|---|---|---|---|---|
| T1-PAES-FB * | 231,664.31 | 16,233.86 | 36,347.13 | −192.24 | 1306 | 52,383.50 |
| T2-PAES-FB * | 233,075.42 | 16,392.48 | 36,269.15 | −189.33 | 1305 | 52,318.88 |
| T3-PAES-FB * | 233,107.8 | 16,370.63 | 36,218.66 | −188.28 | 1306 | 52,167.98 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mollahosseini, A.; Abdelrasoul, A. Assessment of Fibrinogen Thermal Conductivity and Interaction Energy with Polyarylethersulfone (PAES) Clinical Hemodialysis Membranes at Normal and Elevated Patient Body Temperatures. C 2023, 9, 33. https://doi.org/10.3390/c9010033
Mollahosseini A, Abdelrasoul A. Assessment of Fibrinogen Thermal Conductivity and Interaction Energy with Polyarylethersulfone (PAES) Clinical Hemodialysis Membranes at Normal and Elevated Patient Body Temperatures. C. 2023; 9(1):33. https://doi.org/10.3390/c9010033
Chicago/Turabian StyleMollahosseini, Arash, and Amira Abdelrasoul. 2023. "Assessment of Fibrinogen Thermal Conductivity and Interaction Energy with Polyarylethersulfone (PAES) Clinical Hemodialysis Membranes at Normal and Elevated Patient Body Temperatures" C 9, no. 1: 33. https://doi.org/10.3390/c9010033
APA StyleMollahosseini, A., & Abdelrasoul, A. (2023). Assessment of Fibrinogen Thermal Conductivity and Interaction Energy with Polyarylethersulfone (PAES) Clinical Hemodialysis Membranes at Normal and Elevated Patient Body Temperatures. C, 9(1), 33. https://doi.org/10.3390/c9010033