
Ethanol- and PARP-Mediated Regulation of Ribosome-Associated Long Non-Coding RNA (lncRNA) in Pyramidal Neurons

 ,
,
Abstract
:1. Introduction
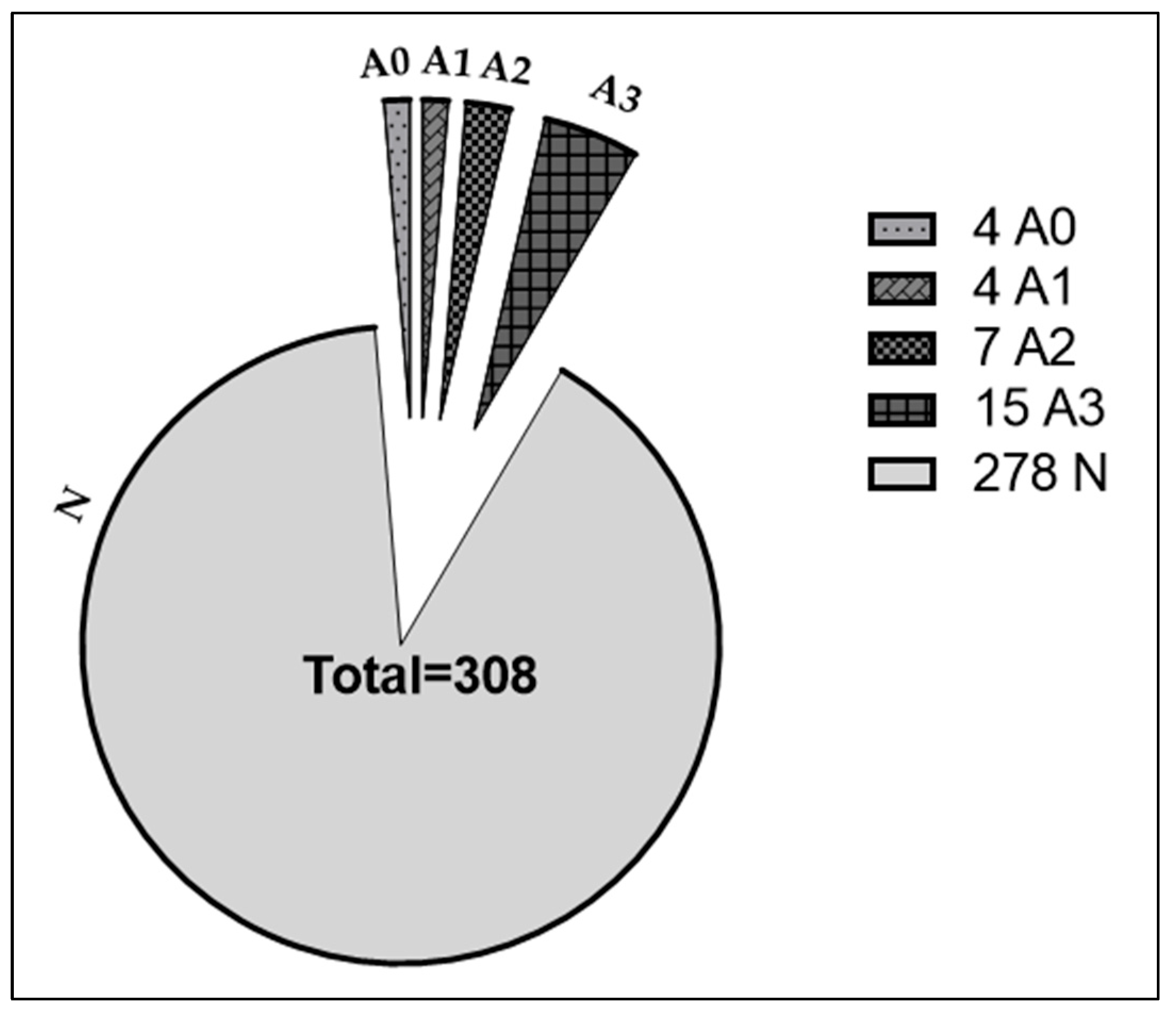
2. Results
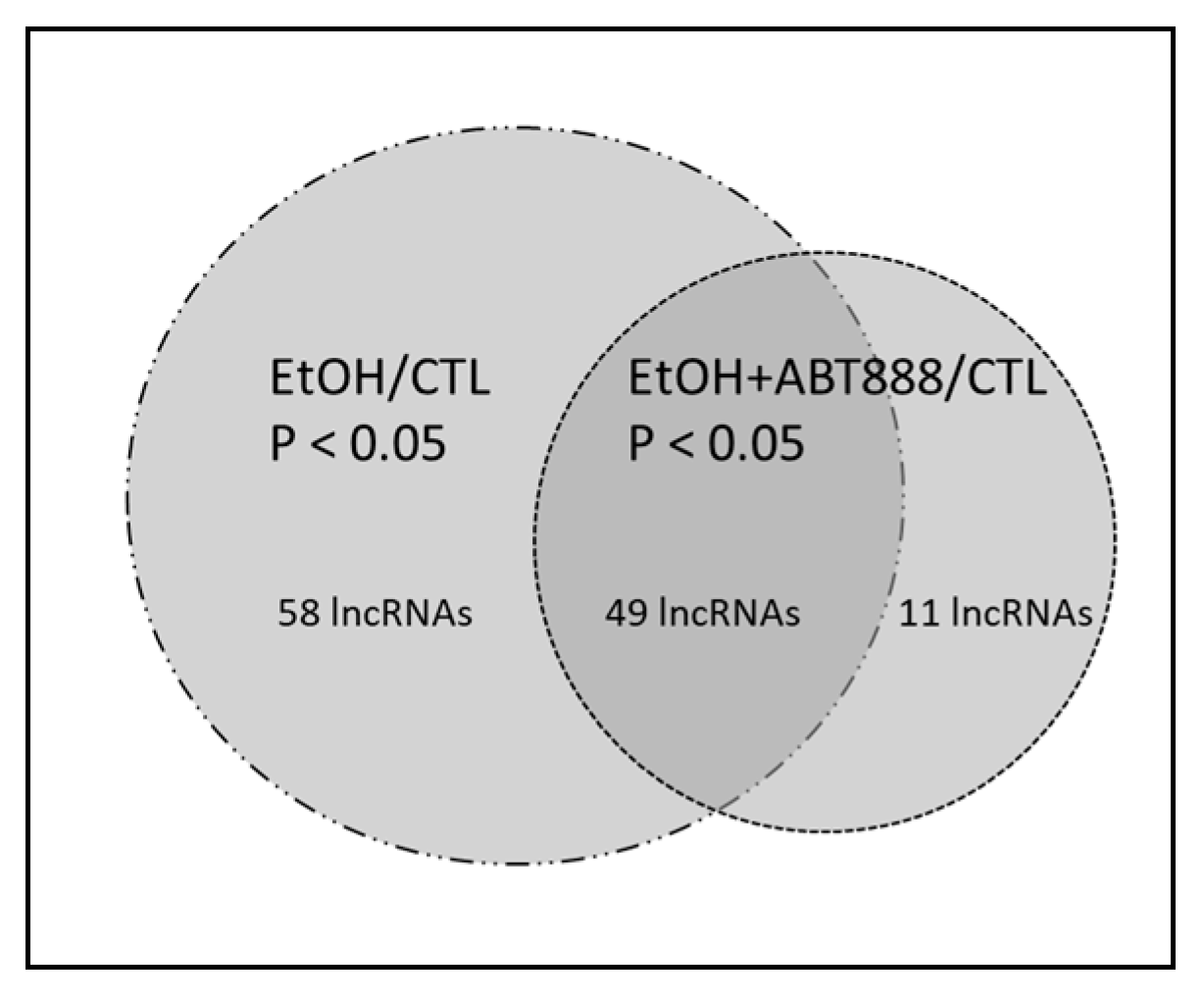
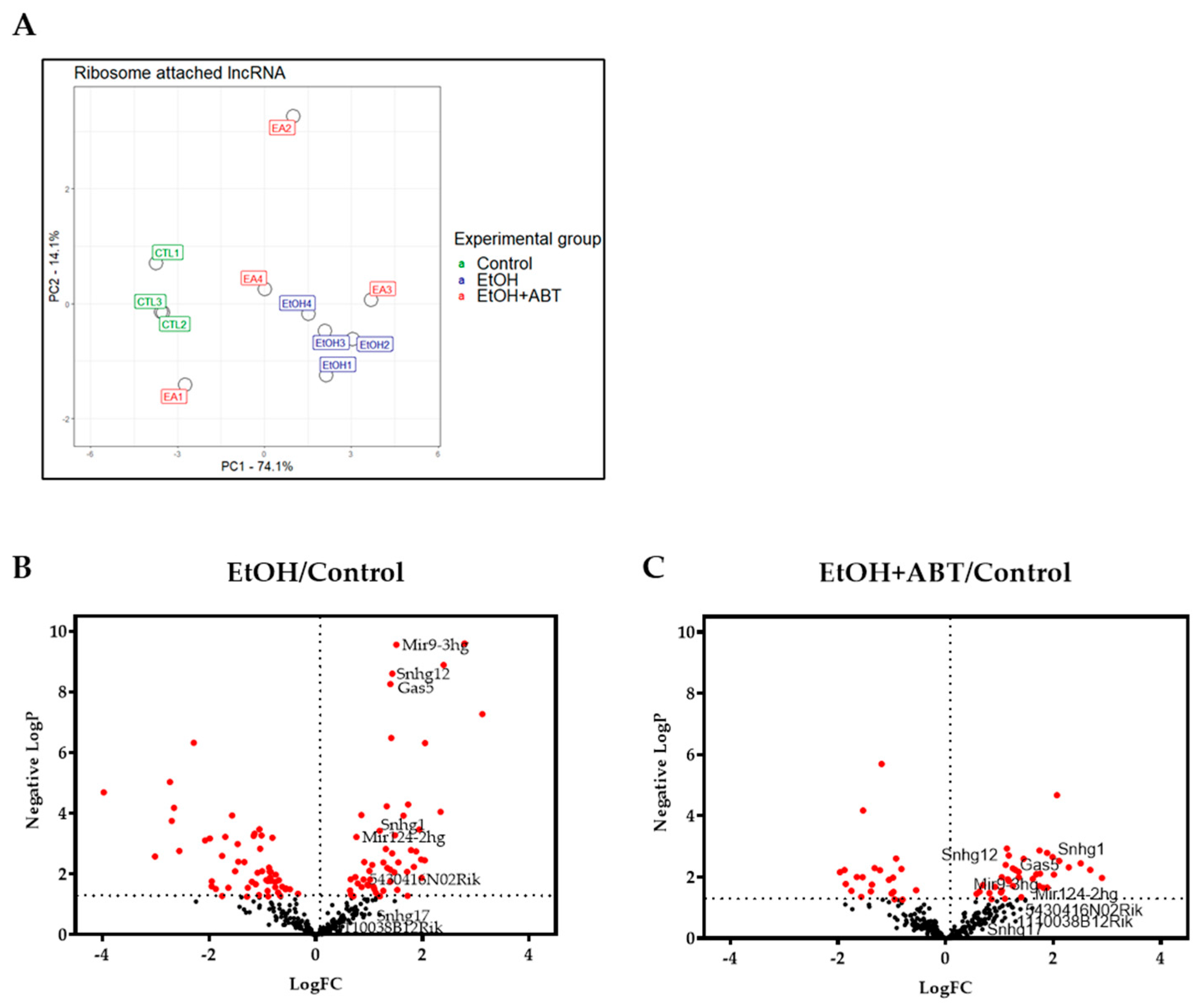
2.1. Effects of EtOH and EtOH+ABT-888 over CTL on Ribosome-Attached lncRNAs Measured by RNA-Seq
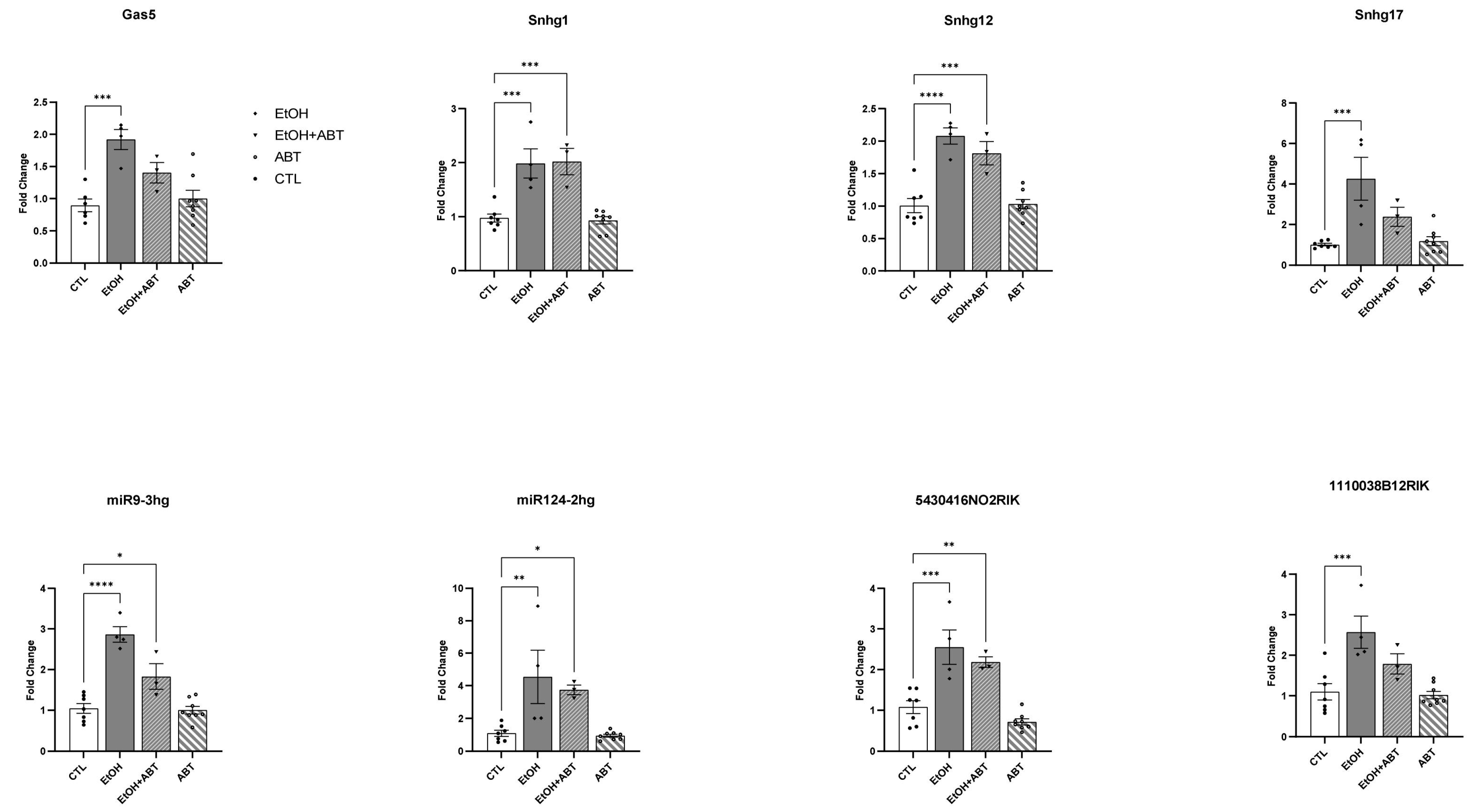
2.2. qRT-PCR Validation of Putative Protein-Coding lncRNA

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LncRNA | Protein-Coding Potential | Known Function |
|---|---|---|
| MIR124-2hg | A0 | Chr 3: MIR124-2 host gene is the most abundant brain-specific miR, promotes microglial quiescence, inhibits microglial and astrocyte activation, and suppresses experimental autoimmune encephalomyelitis by deactivating macrophages via the CEBPA/C/EBP-a-SPI1/PU.1 pathway. |
| 5430416N02Rik | A0 | Chr 5: lncRNA 5430416N02Rik locus regulates Mid1 expression through 3D chromatin organization. Klf4 and CTCF bind to 5430416N02Rik RNA [31]. Knockout of the 5430416N02Rik locus reduces the proliferation rate of embryonic stem cells (ESCs). |
| Snhg17 | A2 | Chr 2: Small nucleolar RNA host gene 17 is expressed in nervous system, acts as a sponge for multiple miRNAs, such as miR-23a-3p, miR-485-5p, and miR-361-3p, promoting tumor progression in various malignancies. |
| Snhg12 | A3 | Chr 4: Small nucleolar RNA host gene 12 is an oncogene that promotes tumor formation through cell cycle, invasion, metastasis, and apoptosis. It also affects the Wnt-β-catenin and MLK3/IκB/NF-κB pathway, Notch-1 signaling pathway, and PI3K/AKT pathway, and serves as a competing endogenous RNA (ceRNA), similar to a miRNA sponge, wherein it attenuates the effects of miRNAs, often leading to cancer progression. |
| Snhg1 | A3 | Chr 19: Small nucleolar RNA host gene 1 has a role in synapse formation. It promotes cell proliferation via upregulating β-catenin, c-Myc, and cyclin D1 pathways. It negatively regulates tumor suppressor genes such as tumor protein p53. SNHG1 could directly interact with Polycomb Repressive Complex 2 (PRC2) and modulate the histone methylation of promoter of Kruppel-like factor 2 (KLF2) and Cyclin-dependent kinase inhibitor 2B (CDKN2B) in the nucleus. |
| MIR9-3hg | A3 | Chr 7: LncRNA with a role in immune function. It can maintain neural cells in an undifferentiated state by regulating NR2E1. |
| Gas5 | A3 | Chr 5: Growth arrest-specific 5 has anti-tumor effects by inhibiting a number of miRNAs and lncRNAs, in part by functioning as a molecular sponge for some miRNAs. In cancer, it promotes apoptosis, inhibits proliferation, and invasion. GAS5 binds to the promoter of the insulin receptor gene and increases its expression. It directly interacts with EZH2 and also acts as a decoy for the glucocorticoid receptor (GR), blocking the upregulation of gene expression by activated GR. |
| 1110038B12Rik | A3 | Chr 17: RIKEN cDNA 1110038B12 aka Atrolnc-1 is abundantly expressed in skeletal muscles, and its expression is markedly increased in atrophying muscles by interacting with ABIN-1, an endogenous NF-κB inhibitor, leading to increased MuRF-1 expression and muscle proteolysis. |
3. Discussion
4. Materials and Methods
4.1. Transgenic Mice
4.2. Animal Treatment and Sample Collection
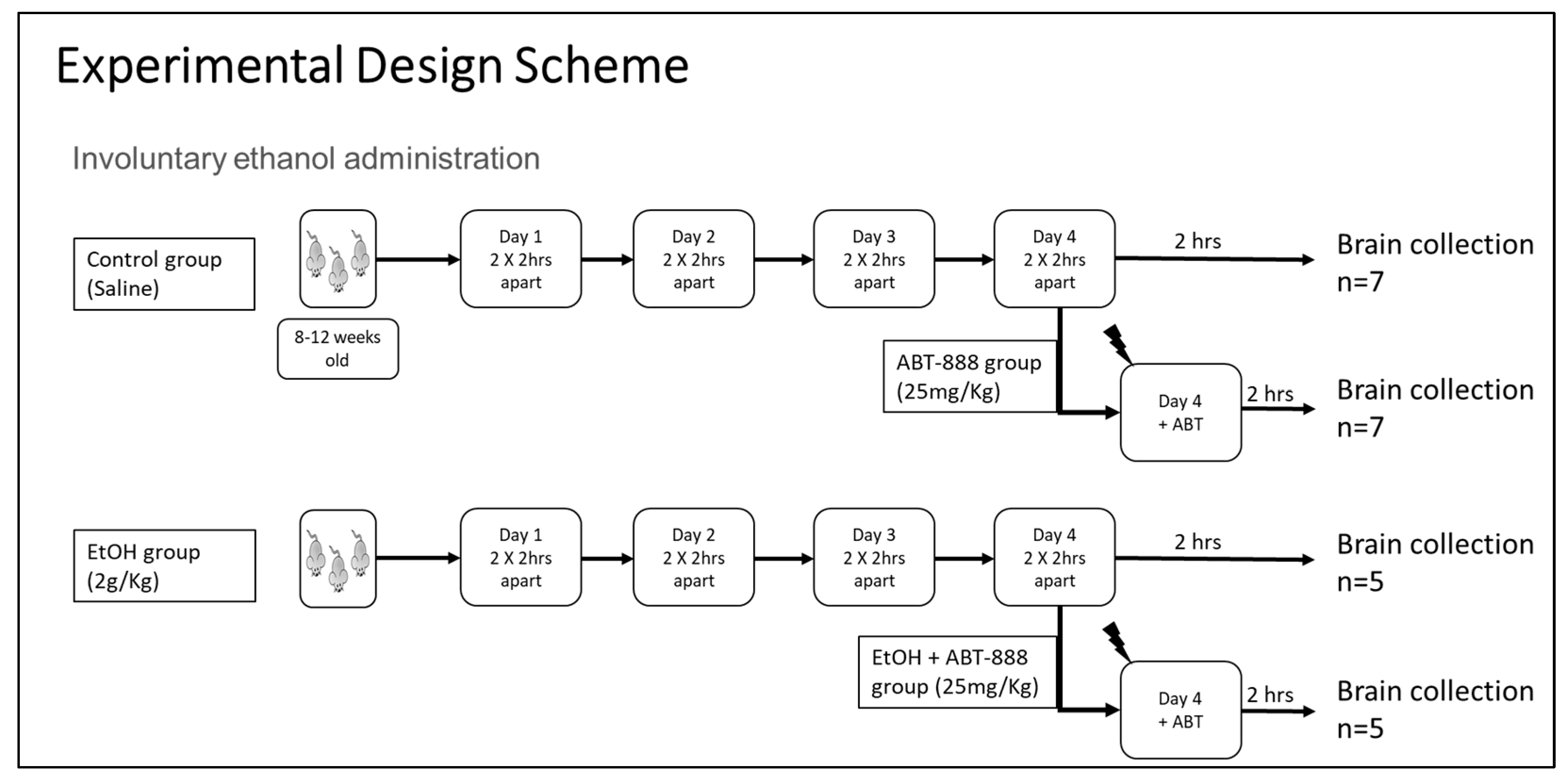
4.3. Blood Ethanol Concentration (BEC)
4.4. Immunoprecipitation and RNA Extraction (TRAP Protocol)
4.5. RNA-Seq Sample Preparation and Analysis
4.6. In Silico Comparison of Protein-Coding Potential Identities of Ribosome-Attached lncRNAs
4.7. Quantitative Real-Time Reverse Transcription Polymerase Chain Reaction-Seq Sample Preparation and Analysis
4.8. Validation of Translating RNA from CaMKIIα-Expressing Cell Isolation Using Gene-Specific Expression
5. Limitations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brown, R.M.; Kim, A.K.; Khoo, S.Y.S.; Kim, J.H.; Jupp, B.; Lawrence, A.J. Orexin-1 receptor signalling in the prelimbic cortex and ventral tegmental area regulates cue-induced reinstatement of ethanol-seeking in iP rats. Addict. Biol. 2016, 21, 603–612. [Google Scholar] [CrossRef]
- Cannady, R.; McGonigal, J.T.; Newsom, R.J.; Woodward, J.J.; Mulholland, P.J.; Gass, J.T. Prefrontal cortex KCa2 channels regulate mGlu5-dependent plasticity and extinction of alcohol-seeking behavior. J. Neurosci. 2017, 37, 4359–4369. [Google Scholar] [CrossRef]
- Gass, J.T.; Trantham-Davidson, H.; Kassab, A.S.; Glen, W.B.; Olive, M.F.; Chandler, L.J. Enhancement of extinction learning attenuates ethanol-seeking behavior and alters plasticity in the prefrontal cortex. J. Neurosci. 2014, 34, 7562–7574. [Google Scholar] [CrossRef]
- Kapranov, P.; Cheng, J.; Dike, S.; Nix, D.A.; Duttagupta, R.; Willingham, A.T.; Stadler, P.F.; Hertel, J.; Hackermüller, J.; Hofacker, I.L. RNA maps reveal new RNA classes and a possible function for pervasive transcription. Science 2007, 316, 1484–1488. [Google Scholar] [CrossRef]
- Gavin, D.P.; Kusumo, H.; Zhang, H.; Guidotti, A.; Pandey, S.C. Role of growth arrest and DNA damage-inducible, beta in alcohol-drinking behaviors. Alcohol. Clin. Exp. Res. 2016, 40, 263–272. [Google Scholar] [CrossRef]
- Vallerini, G.P.; Cheng, Y.-H.; Chase, K.A.; Sharma, R.P.; Kusumo, H.; Khakhkhar, S.; Feinstein, D.L.; Guizzetti, M.; Gavin, D.P. Modulation of Poly ADP Ribose Polymerase (PARP) Levels and Activity by Alcohol Binge-Like Drinking in Male Mice. Neuroscience 2020, 448, 1–13. [Google Scholar] [CrossRef]
- Krishnan, H.R.; Vallerini, G.P.; Gavin, H.E.; Guizzetti, M.; Rizavi, H.S.; Gavin, D.P.; Sharma, R.P. Effects of alcohol and PARP inhibition on RNA ribosomal engagement in cortical excitatory neurons. Front. Mol. Neurosci. 2023, 16, 1125160. [Google Scholar] [CrossRef]
- Huang, X.; Luo, Y.-L.; Mao, Y.-S.; Ji, J.-L. The link between long noncoding RNAs and depression. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2017, 73, 73–78. [Google Scholar] [CrossRef]
- Sartor, G.C.; St. Laurent, G., III; Wahlestedt, C. The emerging role of non-coding RNAs in drug addiction. Front. Genet. 2012, 3, 106. [Google Scholar] [CrossRef]
- Sudhalkar, N.; Rosen, C.; Melbourne, J.K.; Park, M.R.; Chase, K.A.; Sharma, R.P. Long non-coding RNAs associated with heterochromatin function in immune cells in psychosis. Non Coding RNA 2018, 4, 43. [Google Scholar] [CrossRef]
- Tian, T.; Wei, Z.; Chang, X.; Liu, Y.; Gur, R.E.; Sleiman, P.M.; Hakonarson, H. The long noncoding RNA landscape in amygdala tissues from schizophrenia patients. EBioMedicine 2018, 34, 171–181. [Google Scholar] [CrossRef]
- Kopp, F.; Mendell, J.T. Functional classification and experimental dissection of long noncoding RNAs. Cell 2018, 172, 393–407. [Google Scholar] [CrossRef]
- Cabili, M.N.; Trapnell, C.; Goff, L.; Koziol, M.; Tazon-Vega, B.; Regev, A.; Rinn, J.L. Integrative annotation of human large intergenic noncoding RNAs reveals global properties and specific subclasses. Genes Dev. 2011, 25, 1915–1927. [Google Scholar] [CrossRef]
- Derrien, T.; Johnson, R.; Bussotti, G.; Tanzer, A.; Djebali, S.; Tilgner, H.; Guernec, G.; Martin, D.; Merkel, A.; Knowles, D.G. The GENCODE v7 catalog of human long noncoding RNAs: Analysis of their gene structure, evolution, and expression. Genome Res. 2012, 22, 1775–1789. [Google Scholar] [CrossRef]
- Long, Y.; Wang, X.; Youmans, D.T.; Cech, T.R. How do lncRNAs regulate transcription? Sci. Adv. 2017, 3, eaao2110. [Google Scholar] [CrossRef]
- Marchese, F.P.; Raimondi, I.; Huarte, M. The multidimensional mechanisms of long noncoding RNA function. Genome Biol. 2017, 18, 1–13. [Google Scholar] [CrossRef]
- Statello, L.; Guo, C.-J.; Chen, L.-L.; Huarte, M. Gene regulation by long non-coding RNAs and its biological functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef]
- Zeng, C.; Fukunaga, T.; Hamada, M. Identification and analysis of ribosome-associated lncRNAs using ribosome profiling data. BMC Genom. 2018, 19, 1–14. [Google Scholar] [CrossRef]
- Bazin, J.; Baerenfaller, K.; Gosai, S.J.; Gregory, B.D.; Crespi, M.; Bailey-Serres, J. Global analysis of ribosome-associated noncoding RNAs unveils new modes of translational regulation. Proc. Natl. Acad. Sci. USA 2017, 114, E10018–E10027. [Google Scholar] [CrossRef]
- Carrieri, C.; Cimatti, L.; Biagioli, M.; Beugnet, A.; Zucchelli, S.; Fedele, S.; Pesce, E.; Ferrer, I.; Collavin, L.; Santoro, C. Long non-coding antisense RNA controls Uchl1 translation through an embedded SINEB2 repeat. Nature 2012, 491, 454–457. [Google Scholar] [CrossRef]
- Ingolia, N.T.; Brar, G.A.; Stern-Ginossar, N.; Harris, M.S.; Talhouarne, G.J.; Jackson, S.E.; Wills, M.R.; Weissman, J.S. Ribosome profiling reveals pervasive translation outside of annotated protein-coding genes. Cell Rep. 2014, 8, 1365–1379. [Google Scholar] [CrossRef]
- Jabnoune, M.; Secco, D.; Lecampion, C.; Robaglia, C.; Shu, Q.; Poirier, Y. A rice cis-natural antisense RNA acts as a translational enhancer for its cognate mRNA and contributes to phosphate homeostasis and plant fitness. Plant Cell 2013, 25, 4166–4182. [Google Scholar] [CrossRef]
- Yoon, J.-H.; Abdelmohsen, K.; Gorospe, M. Posttranscriptional gene regulation by long noncoding RNA. J. Mol. Biol. 2013, 425, 3723–3730. [Google Scholar] [CrossRef]
- Cai, X.; Cullen, B.R. The imprinted H19 noncoding RNA is a primary microRNA precursor. RNA 2007, 13, 313–316. [Google Scholar] [CrossRef]
- Wilson, B.A.; Masel, J. Putatively noncoding transcripts show extensive association with ribosomes. Genome Biol. Evol. 2011, 3, 1245–1252. [Google Scholar] [CrossRef]
- Van Heesch, S.; van Iterson, M.; Jacobi, J.; Boymans, S.; Essers, P.B.; de Bruijn, E.; Hao, W.; MacInnes, A.W.; Cuppen, E.; Simonis, M. Extensive localization of long noncoding RNAs to the cytosol and mono-and polyribosomal complexes. Genome Biol. 2014, 15, R6. [Google Scholar] [CrossRef]
- Heiman, M.; Schaefer, A.; Gong, S.; Peterson, J.D.; Day, M.; Ramsey, K.E.; Suarez-Farinas, M.; Schwarz, C.; Stephan, D.A.; Surmeier, D.J. A translational profiling approach for the molecular characterization of CNS cell types. Cell 2008, 135, 738–748. [Google Scholar] [CrossRef]
- Lyons, L.C.; Chatterjee, S.; Vanrobaeys, Y.; Gaine, M.E.; Abel, T. Translational changes induced by acute sleep deprivation uncovered by TRAP-Seq. Mol. Brain 2020, 13, 165. [Google Scholar] [CrossRef]
- Wu, H.; Lai, C.-F.; Chang-Panesso, M.; Humphreys, B.D. Proximal tubule translational profiling during kidney fibrosis reveals proinflammatory and long noncoding RNA expression patterns with sexual dimorphism. J. Am. Soc. Nephrol. 2020, 31, 23–38. [Google Scholar] [CrossRef]
- Zhou, P.; Zhang, Y.; Ma, Q.; Gu, F.; Day, D.S.; He, A.; Zhou, B.; Li, J.; Stevens, S.M.; Romo, D. Interrogating translational efficiency and lineage-specific transcriptomes using ribosome affinity purification. Proc. Natl. Acad. Sci. USA 2013, 110, 15395–15400. [Google Scholar] [CrossRef]
- Kung, J.T.; Kesner, B.; An, J.Y.; Ahn, J.Y.; Cifuentes-Rojas, C.; Colognori, D.; Jeon, Y.; Szanto, A.; del Rosario, B.C.; Pinter, S.F.; et al. Locus-specific targeting to the X chromosome revealed by the RNA interactome of CTCF. Mol. Cell 2015, 57, 361–375. [Google Scholar] [CrossRef]
- Haft, R.J.; Keating, D.H.; Schwaegler, T.; Schwalbach, M.S.; Vinokur, J.; Tremaine, M.; Peters, J.M.; Kotlajich, M.V.; Pohlmann, E.L.; Ong, I.M. Correcting direct effects of ethanol on translation and transcription machinery confers ethanol tolerance in bacteria. Proc. Natl. Acad. Sci. USA 2014, 111, E2576–E2585. [Google Scholar] [CrossRef]
- Noble, E.P.; Tewari, S. Ethanol and Brain Ribosomes. In Federation Proceedings; Federation of American Societies for Experimental Biology: Bethesda, MD, USA, 1975. [Google Scholar]
- Kim, D.-S.; Challa, S.; Jones, A.; Kraus, W.L. PARPs and ADP-ribosylation in RNA biology: From RNA expression and processing to protein translation and proteostasis. Genes Dev. 2020, 34, 302–320. [Google Scholar] [CrossRef]
- Huang, D.; Kraus, W.L. The expanding universe of PARP1-mediated molecular and therapeutic mechanisms. Mol. Cell 2022, 82, 2315–2334. [Google Scholar] [CrossRef]
- Munnur, D.; Bartlett, E.; Mikolčević, P.; Kirby, I.T.; Rack, J.G.M.; Mikoč, A.; Cohen, M.S.; Ahel, I. Reversible ADP-ribosylation of RNA. Nucleic Acids Res. 2019, 47, 5658–5669. [Google Scholar] [CrossRef]
- Weixler, L.; Schäringer, K.; Momoh, J.; Lüscher, B.; Feijs, K.L.; Žaja, R. ADP-ribosylation of RNA and DNA: From in vitro characterization to in vivo function. Nucleic Acids Res. 2021, 49, 3634–3650. [Google Scholar] [CrossRef]
- Kino, T.; Hurt, D.E.; Ichijo, T.; Nader, N.; Chrousos, G.P. Noncoding RNA gas5 is a growth arrest–and starvation-associated repressor of the glucocorticoid receptor. Sci. Signal. 2010, 3, ra8. [Google Scholar] [CrossRef]
- He, X.; Chen, X.; Zhang, X.; Duan, X.; Pan, T.; Hu, Q.; Zhang, Y.; Zhong, F.; Liu, J.; Zhang, H. An Lnc RNA (GAS5)/SnoRNA-derived piRNA induces activation of TRAIL gene by site-specifically recruiting MLL/COMPASS-like complexes. Nucleic Acids Res. 2015, 43, 3712–3725. [Google Scholar] [CrossRef]
- Zhao, T.; Cai, M.; Liu, M.; Su, G.; An, D.; Moon, B.; Lyu, G.; Si, Y.; Chen, L.; Lu, W. lncRNA 5430416N02Rik promotes the proliferation of mouse embryonic stem cells by activating Mid1 expression through 3D chromatin architecture. Stem Cell Rep. 2020, 14, 493–505. [Google Scholar] [CrossRef]
- Hitachi, K.; Honda, M.; Tsuchida, K. The Functional Role of Long Non-Coding RNA in Myogenesis and Skeletal Muscle Atrophy. Cells 2022, 11, 2291. [Google Scholar] [CrossRef]
- Haemmig, S.; Yang, D.; Sun, X.; Das, D.; Ghaffari, S.; Molinaro, R.; Chen, L.; Deng, Y.; Freeman, D.; Moullan, N. Long noncoding RNA SNHG12 integrates a DNA-PK–mediated DNA damage response and vascular senescence. Sci. Transl. Med. 2020, 12, eaaw1868. [Google Scholar] [CrossRef]
- Du, Q.; Xiao, R.-D.; Luo, R.-G.; Xie, J.-B.; Su, Z.-D.; Wang, Y. Construction of long non-coding RNA-and microRNA-mediated competing endogenous RNA networks in alcohol-related esophageal cancer. PLoS ONE 2022, 17, e0269742. [Google Scholar] [CrossRef]
- Nowak, J.S.; Michlewski, G. miRNAs in development and pathogenesis of the nervous system. Biochem. Soc. Trans. 2013, 41, 815–820. [Google Scholar] [CrossRef]
- Nowak, J.S.; Choudhury, N.R.; de Lima Alves, F.; Rappsilber, J.; Michlewski, G. Lin28a regulates neuronal differentiation and controls miR-9 production. Nat. Commun. 2014, 5, 3687. [Google Scholar] [CrossRef]
- Sonntag, K.C.; Woo, T.-U.W.; Krichevsky, A.M. Converging miRNA functions in diverse brain disorders: A case for miR-124 and miR-126. Exp. Neurol. 2012, 235, 427–435. [Google Scholar] [CrossRef]
- Truong, T.T.; Bortolasci, C.C.; Spolding, B.; Panizzutti, B.; Liu, Z.S.; Kidnapillai, S.; Richardson, M.; Gray, L.; Smith, C.M.; Dean, O.M. Co-expression networks unveiled long non-coding RNAs as molecular targets of drugs used to treat bipolar disorder. Front. Pharmacol. 2022, 1255, 873271. [Google Scholar] [CrossRef]
- Fang, M.; Wang, J.; Zhang, X.; Geng, Y.; Hu, Z.; Rudd, J.A.; Ling, S.; Chen, W.; Han, S. The miR-124 regulates the expression of BACE1/β-secretase correlated with cell death in Alzheimer’s disease. Toxicol. Lett. 2012, 209, 94–105. [Google Scholar] [CrossRef]
- Politz, J.C.R.; Tuft, R.A.; Prasanth, K.V.; Baudendistel, N.; Fogarty, K.E.; Lifshitz, L.M.; Langowski, J.; Spector, D.L.; Pederson, T. Rapid, diffusional shuttling of poly (A) RNA between nuclear speckles and the nucleoplasm. Mol. Biol. Cell 2006, 17, 1239–1249. [Google Scholar] [CrossRef]
- Jones, E.; Huntley, G.; Benson, D. Alpha calcium/calmodulin-dependent protein kinase II selectively expressed in a subpopulation of excitatory neurons in monkey sensory-motor cortex: Comparison with GAD-67 expression. J. Neurosci. 1994, 14, 611–629. [Google Scholar] [CrossRef]
- Tighilet, B.; Hashikawa, T.; Jones, E.G. Cell-and lamina-specific expression and activity-dependent regulation of type II calcium/calmodulin-dependent protein kinase isoforms in monkey visual cortex. J. Neurosci. 1998, 18, 2129–2146. [Google Scholar] [CrossRef]
- Walter, T.J.; Crews, F.T. Microglial depletion alters the brain neuroimmune response to acute binge ethanol withdrawal. J. Neuroinflammation 2017, 14, 86. [Google Scholar] [CrossRef]
- Jung, G.; Ferard, G. Enzyme-coupled measurement of ethanol in whole blood and plasma with a centrifugal analyzer. Clin. Chem. 1978, 24, 873–876. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef]
- McCarthy, D.J.; Chen, Y.; Smyth, G.K. Differential expression analysis of multifactor RNA-Seq experiments with respect to biological variation. Nucleic Acids Res. 2012, 40, 4288–4297. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rizavi, H.S.; Gavin, H.E.; Krishnan, H.R.; Gavin, D.P.; Sharma, R.P. Ethanol- and PARP-Mediated Regulation of Ribosome-Associated Long Non-Coding RNA (lncRNA) in Pyramidal Neurons. Non-Coding RNA 2023, 9, 72. https://doi.org/10.3390/ncrna9060072
Rizavi HS, Gavin HE, Krishnan HR, Gavin DP, Sharma RP. Ethanol- and PARP-Mediated Regulation of Ribosome-Associated Long Non-Coding RNA (lncRNA) in Pyramidal Neurons. Non-Coding RNA. 2023; 9(6):72. https://doi.org/10.3390/ncrna9060072
Chicago/Turabian StyleRizavi, Hooriyah S., Hannah E. Gavin, Harish R. Krishnan, David P. Gavin, and Rajiv P. Sharma. 2023. "Ethanol- and PARP-Mediated Regulation of Ribosome-Associated Long Non-Coding RNA (lncRNA) in Pyramidal Neurons" Non-Coding RNA 9, no. 6: 72. https://doi.org/10.3390/ncrna9060072
APA StyleRizavi, H. S., Gavin, H. E., Krishnan, H. R., Gavin, D. P., & Sharma, R. P. (2023). Ethanol- and PARP-Mediated Regulation of Ribosome-Associated Long Non-Coding RNA (lncRNA) in Pyramidal Neurons. Non-Coding RNA, 9(6), 72. https://doi.org/10.3390/ncrna9060072