

Discovery and Comprehensive Characterization of Novel Circular RNAs of the Apoptosis-Related BOK Gene in Human Ovarian and Prostate Cancer Cells, Using Nanopore Sequencing

 ,
, 
 ,
,  , ,
, ,
Abstract
1. Introduction
2. Results
2.1. Identification and Characterization of Novel BOK circRNAs in Ovarian and Prostate Cancer Cell Lines
2.2. Expression Analysis of the Novel BOK circRNAs
2.3. Putative Interactions of BOK circRNAs with miRNAs
2.4. Prediction of the Translational Potential of BOK circRNAs
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Total RNA Extraction and Reverse Transcription
4.3. Primer Designing and Amplification of BOK circRNAs
4.4. Library Preparation and Third-Generation Sequencing with Nanopore Technology
4.5. Third-Generation Sequencing Data Analysis
4.6. In Silico Analysis for the Putative Functions of the Novel BOK circRNAs
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wu, Y.P.; Lin, X.D.; Chen, S.H.; Ke, Z.B.; Lin, F.; Chen, D.N.; Xue, X.Y.; Wei, Y.; Zheng, Q.S.; Wen, Y.A.; et al. Identification of Prostate Cancer-Related Circular RNA Through Bioinformatics Analysis. Front. Genet. 2020, 11, 892. [Google Scholar] [CrossRef] [PubMed]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Favaloro, B.; Allocati, N.; Graziano, V.; Di Ilio, C.; De Laurenzi, V. Role of apoptosis in disease. Aging 2012, 4, 330–349. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. The hallmarks of cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef]
- Singh, R.; Letai, A.; Sarosiek, K. Regulation of apoptosis in health and disease: The balancing act of BCL-2 family proteins. Nat. Rev. Mol. Cell Biol. 2019, 20, 175–193. [Google Scholar] [CrossRef]
- Gurumurthy, S.; Vasudevan, K.M.; Rangnekar, V.M. Regulation of apoptosis in prostate cancer. Cancer Metastasis Rev. 2001, 20, 225–243. [Google Scholar] [CrossRef]
- Al-Alem, L.F.; Baker, A.T.; Pandya, U.M.; Eisenhauer, E.L.; Rueda, B.R. Understanding and Targeting Apoptotic Pathways in Ovarian Cancer. Cancers 2019, 11, 1631. [Google Scholar] [CrossRef]
- Zhang, H.; Holzgreve, W.; De Geyter, C. Evolutionarily conserved Bok proteins in the Bcl-2 family. FEBS Lett. 2000, 480, 311–313. [Google Scholar] [CrossRef]
- Jaaskelainen, M.; Nieminen, A.; Pokkyla, R.M.; Kauppinen, M.; Liakka, A.; Heikinheimo, M.; Vaskivuo, T.E.; Klefstrom, J.; Tapanainen, J.S. Regulation of cell death in human fetal and adult ovaries—Role of Bok and Bcl-XL. Mol. Cell. Endocrinol. 2010, 330, 17–24. [Google Scholar] [CrossRef]
- Llambi, F.; Wang, Y.M.; Victor, B.; Yang, M.; Schneider, D.M.; Gingras, S.; Parsons, M.J.; Zheng, J.H.; Brown, S.A.; Pelletier, S.; et al. BOK Is a Non-canonical BCL-2 Family Effector of Apoptosis Regulated by ER-Associated Degradation. Cell 2016, 165, 421–433. [Google Scholar] [CrossRef] [PubMed]
- Soleymanlou, N.; Wu, Y.; Wang, J.X.; Todros, T.; Ietta, F.; Jurisicova, A.; Post, M.; Caniggia, I. A novel Mtd splice isoform is responsible for trophoblast cell death in pre-eclampsia. Cell Death Differ. 2005, 12, 441–452. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Einsele-Scholz, S.; Malmsheimer, S.; Bertram, K.; Stehle, D.; Johanning, J.; Manz, M.; Daniel, P.T.; Gillissen, B.F.; Schulze-Osthoff, K.; Essmann, F. Bok is a genuine multi-BH-domain protein that triggers apoptosis in the absence of Bax and Bak. J. Cell Sci. 2016, 129, 2213–2223. [Google Scholar] [CrossRef]
- Hsu, M.T.; Coca-Prados, M. Electron microscopic evidence for the circular form of RNA in the cytoplasm of eukaryotic cells. Nature 1979, 280, 339–340. [Google Scholar] [CrossRef]
- Memczak, S.; Jens, M.; Elefsinioti, A.; Torti, F.; Krueger, J.; Rybak, A.; Maier, L.; Mackowiak, S.D.; Gregersen, L.H.; Munschauer, M.; et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature 2013, 495, 333–338. [Google Scholar] [CrossRef]
- Chen, Y.; Li, C.; Tan, C.; Liu, X. Circular RNAs: A new frontier in the study of human diseases. J. Med. Genet. 2016, 53, 359–365. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Li, X.; Zhang, P.; Wang, J.; Zhou, Y.; Chen, M. Circular RNA: An emerging key player in RNA world. Brief. Bioinform. 2017, 18, 547–557. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Yang, Y.; Zhao, X.; Fan, Y.; Zhou, L.; Rong, J.; Yu, Y. Circular RNA circ0005276 promotes the proliferation and migration of prostate cancer cells by interacting with FUS to transcriptionally activate XIAP. Cell Death Dis. 2019, 10, 792. [Google Scholar] [CrossRef]
- Cao, C.; Sun, G.; Le, K.; Xu, Q.; Liu, C. The Circular RNA Circ_0085494 Regulates Prostate Cancer Progression Through NRBP1/miR-497-5p Axis. Biochem. Genet. 2023, 1–16. [Google Scholar] [CrossRef]
- Lv, D.; Cen, S.; Yang, S.; Zou, Z.; Zhong, J.; Pan, Z.; Deng, N.; Li, Y.; Wu, K.; Wang, J.; et al. Hsa_circ_0063329 inhibits prostate cancer growth and metastasis by modulating the miR-605-5p/tgif2 axis. Cell Cycle 2023, 22, 1101–1115. [Google Scholar] [CrossRef]
- Gan, X.; Zhu, H.; Jiang, X.; Obiegbusi, S.C.; Yong, M.; Long, X.; Hu, J. CircMUC16 promotes autophagy of epithelial ovarian cancer via interaction with ATG13 and miR-199a. Mol. Cancer 2020, 19, 45. [Google Scholar] [CrossRef]
- Zhao, Y.; Hu, Y.; Shen, Q.; Chen, Q.; Zhu, X.J.; Jiang, S.S.; Zhang, Q. CircRNA_MYLK promotes malignant progression of ovarian cancer through regulating microRNA-652. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 5281–5291. [Google Scholar] [CrossRef]
- Xu, Q.; Deng, B.; Li, M.; Chen, Y.; Zhuan, L. circRNA-UBAP2 promotes the proliferation and inhibits apoptosis of ovarian cancer though miR-382-5p/PRPF8 axis. J. Ovarian Res. 2020, 13, 81. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Wang, J.; Zheng, Y.; Zhang, J.; Chen, S.; Zhao, F. Comprehensive identification of internal structure and alternative splicing events in circular RNAs. Nat. Commun. 2016, 7, 12060. [Google Scholar] [CrossRef]
- Quick, J.; Loman, N.J.; Duraffour, S.; Simpson, J.T.; Severi, E.; Cowley, L.; Bore, J.A.; Koundouno, R.; Dudas, G.; Mikhail, A.; et al. Real-time, portable genome sequencing for Ebola surveillance. Nature 2016, 530, 228–232. [Google Scholar] [CrossRef] [PubMed]
- Sharon, D.; Tilgner, H.; Grubert, F.; Snyder, M. A single-molecule long-read survey of the human transcriptome. Nat. Biotechnol. 2013, 31, 1009–1014. [Google Scholar] [CrossRef]
- Athanasopoulou, K.; Boti, M.A.; Adamopoulos, P.G.; Skourou, P.C.; Scorilas, A. Third-Generation Sequencing: The Spearhead towards the Radical Transformation of Modern Genomics. Life 2021, 12, 30. [Google Scholar] [CrossRef]
- Xin, R.; Gao, Y.; Gao, Y.; Wang, R.; Kadash-Edmondson, K.E.; Liu, B.; Wang, Y.; Lin, L.; Xing, Y. isoCirc catalogs full-length circular RNA isoforms in human transcriptomes. Nat. Commun. 2021, 12, 266. [Google Scholar] [CrossRef]
- Zhang, J.; Hou, L.; Zuo, Z.; Ji, P.; Zhang, X.; Xue, Y.; Zhao, F. Comprehensive profiling of circular RNAs with nanopore sequencing and CIRI-long. Nat. Biotechnol. 2021, 39, 836–845. [Google Scholar] [CrossRef] [PubMed]
- Rahimi, K.; Veno, M.T.; Dupont, D.M.; Kjems, J. Nanopore sequencing of brain-derived full-length circRNAs reveals circRNA-specific exon usage, intron retention and microexons. Nat. Commun. 2021, 12, 4825. [Google Scholar] [CrossRef]
- Wu, W.; Ji, P.; Zhao, F. CircAtlas: An integrated resource of one million highly accurate circular RNAs from 1070 vertebrate transcriptomes. Genome Biol. 2020, 21, 101. [Google Scholar] [CrossRef] [PubMed]
- Creighton, C.J.; Benham, A.L.; Zhu, H.; Khan, M.F.; Reid, J.G.; Nagaraja, A.K.; Fountain, M.D.; Dziadek, O.; Han, D.; Ma, L.; et al. Discovery of novel microRNAs in female reproductive tract using next generation sequencing. PLoS ONE 2010, 5, e9637. [Google Scholar] [CrossRef]
- Li, G.; Zhang, Y.; Mao, J.; Hu, P.; Chen, Q.; Ding, W.; Pu, R. LncRNA TUC338 is overexpressed in prostate carcinoma and downregulates miR-466. Gene 2019, 707, 224–230. [Google Scholar] [CrossRef]
- Xu, G.; Meng, Y.; Wang, L.; Dong, B.; Peng, F.; Liu, S.; Li, S.; Liu, T. miRNA-214-5p inhibits prostate cancer cell proliferation by targeting SOX4. World J. Surg. Oncol. 2021, 19, 338. [Google Scholar] [CrossRef]
- Patop, I.L.; Wust, S.; Kadener, S. Past, present, and future of circRNAs. EMBO J. 2019, 38, e100836. [Google Scholar] [CrossRef]
- Deveson, I.W.; Brunck, M.E.; Blackburn, J.; Tseng, E.; Hon, T.; Clark, T.A.; Clark, M.B.; Crawford, J.; Dinger, M.E.; Nielsen, L.K.; et al. Universal Alternative Splicing of Noncoding Exons. Cell Syst. 2018, 6, 245–255.e5. [Google Scholar] [CrossRef] [PubMed]
- Papanota, A.M.; Karousi, P.; Kontos, C.K.; Artemaki, P.I.; Liacos, C.I.; Papadimitriou, M.A.; Bagratuni, T.; Eleutherakis-Papaiakovou, E.; Malandrakis, P.; Ntanasis-Stathopoulos, I.; et al. A Cancer-Related microRNA Signature Shows Biomarker Utility in Multiple Myeloma. Int. J. Mol. Sci. 2021, 22, 13144. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Yang, Z.; Yang, C.; Wang, Z.; Chen, D.; Xie, Y.; Wu, Y. Identification and genetic analysis of alternative splicing of long non-coding RNAs in tomato initial flowering stage. Genomics 2020, 112, 897–907. [Google Scholar] [CrossRef] [PubMed]
- Papatsirou, M.; Diamantopoulos, M.A.; Katsaraki, K.; Kletsas, D.; Kontos, C.K.; Scorilas, A. Identification of Novel Circular RNAs of the Human Protein Arginine Methyltransferase 1 (PRMT1) Gene, Expressed in Breast Cancer Cells. Genes 2022, 13, 1133. [Google Scholar] [CrossRef] [PubMed]
- Karousi, P.; Artemaki, P.I.; Sotiropoulou, C.D.; Christodoulou, S.; Scorilas, A.; Kontos, C.K. Identification of Two Novel Circular RNAs Deriving from BCL2L12 and Investigation of Their Potential Value as a Molecular Signature in Colorectal Cancer. Int. J. Mol. Sci. 2020, 21, 8867. [Google Scholar] [CrossRef]
- Sibley, C.R.; Blazquez, L.; Ule, J. Lessons from non-canonical splicing. Nat. Rev. Genet. 2016, 17, 407–421. [Google Scholar] [CrossRef] [PubMed]
- Robic, A.; Kuhn, C. Beyond Back Splicing, a Still Poorly Explored World: Non-Canonical Circular RNAs. Genes 2020, 11, 1111. [Google Scholar] [CrossRef]
- Papatsirou, M.; Scorilas, A.; Sideris, D.C.; Kontos, C.K. Targeted nanopore sequencing for the identification of novel PRMT1 circRNAs unveils a diverse transcriptional profile of this gene in breast cancer cells. Genes Dis. 2024, 11, 589–592. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, S.; Babin, L.; Andraos, E.; Bessiere, C.; Willier, S.; Schulte, J.H.; Gaspin, C.; Meggetto, F. Generation of full-length circular RNA libraries for Oxford Nanopore long-read sequencing. PLoS ONE 2022, 17, e0273253. [Google Scholar] [CrossRef]
- Rahimi, K.; Faerch Nielsen, A.; Veno, M.T.; Kjems, J. Nanopore long-read sequencing of circRNAs. Methods 2021, 196, 23–29. [Google Scholar] [CrossRef]
- Ragan, C.; Goodall, G.J.; Shirokikh, N.E.; Preiss, T. Insights into the biogenesis and potential functions of exonic circular RNA. Sci. Rep. 2019, 9, 2048. [Google Scholar] [CrossRef]
- Ashwal-Fluss, R.; Meyer, M.; Pamudurti, N.R.; Ivanov, A.; Bartok, O.; Hanan, M.; Evantal, N.; Memczak, S.; Rajewsky, N.; Kadener, S. circRNA biogenesis competes with pre-mRNA splicing. Mol. Cell 2014, 56, 55–66. [Google Scholar] [CrossRef]
- Dong, X.; Zhang, P.; Liu, L.; Li, H.; Cheng, S.; Li, S.; Wang, Y.; Zheng, C.; Dong, J.; Zhang, L. The Circ_0001367/miR-545-3p/LUZP1 Axis Regulates Cell Proliferation, Migration and Invasion in Glioma Cells. Front. Oncol. 2021, 11, 781471. [Google Scholar] [CrossRef]
- Wang, H.H.; Ma, J.N.; Zhan, X.R. Circular RNA Circ_0067934 Attenuates Ferroptosis of Thyroid Cancer Cells by miR-545-3p/SLC7A11 Signaling. Front. Endocrinol. 2021, 12, 670031. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Guo, Y.; Song, L.; Tong, T.; Fan, X. Circular RNA intraflagellar transport 80 facilitates endometrial cancer progression through modulating miR-545-3p/FAM98A signaling. J. Gynecol. Oncol. 2022, 33, e2. [Google Scholar] [CrossRef] [PubMed]
- Colden, M.; Dar, A.A.; Saini, S.; Dahiya, P.V.; Shahryari, V.; Yamamura, S.; Tanaka, Y.; Stein, G.; Dahiya, R.; Majid, S. MicroRNA-466 inhibits tumor growth and bone metastasis in prostate cancer by direct regulation of osteogenic transcription factor RUNX2. Cell Death Dis. 2017, 8, e2572. [Google Scholar] [CrossRef]
- Yang, Y.; Fan, X.; Mao, M.; Song, X.; Wu, P.; Zhang, Y.; Jin, Y.; Yang, Y.; Chen, L.L.; Wang, Y.; et al. Extensive translation of circular RNAs driven by N6-methyladenosine. Cell Res. 2017, 27, 626–641. [Google Scholar] [CrossRef]
- Li, H. Minimap2: Pairwise alignment for nucleotide sequences. Bioinformatics 2018, 34, 3094–3100. [Google Scholar] [CrossRef]
- Thorvaldsdottir, H.; Robinson, J.T.; Mesirov, J.P. Integrative Genomics Viewer (IGV): High-performance genomics data visualization and exploration. Brief. Bioinform. 2013, 14, 178–192. [Google Scholar] [CrossRef] [PubMed]
- Adamopoulos, P.G.; Theodoropoulou, M.C.; Scorilas, A. Alternative Splicing Detection Tool-a novel PERL algorithm for sensitive detection of splicing events, based on next-generation sequencing data analysis. Ann. Transl. Med. 2018, 6, 244. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, X. miRDB: An online database for prediction of functional microRNA targets. Nucleic Acids Res. 2020, 48, D127–D131. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zeng, P.; Li, Y.H.; Zhang, Z.; Cui, Q. SRAMP: Prediction of mammalian N6-methyladenosine (m6A) sites based on sequence-derived features. Nucleic Acids Res. 2016, 44, e91. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Gribskov, M. IRESpy: An XGBoost model for prediction of internal ribosome entry sites. BMC Bioinform. 2019, 20, 409. [Google Scholar] [CrossRef] [PubMed]




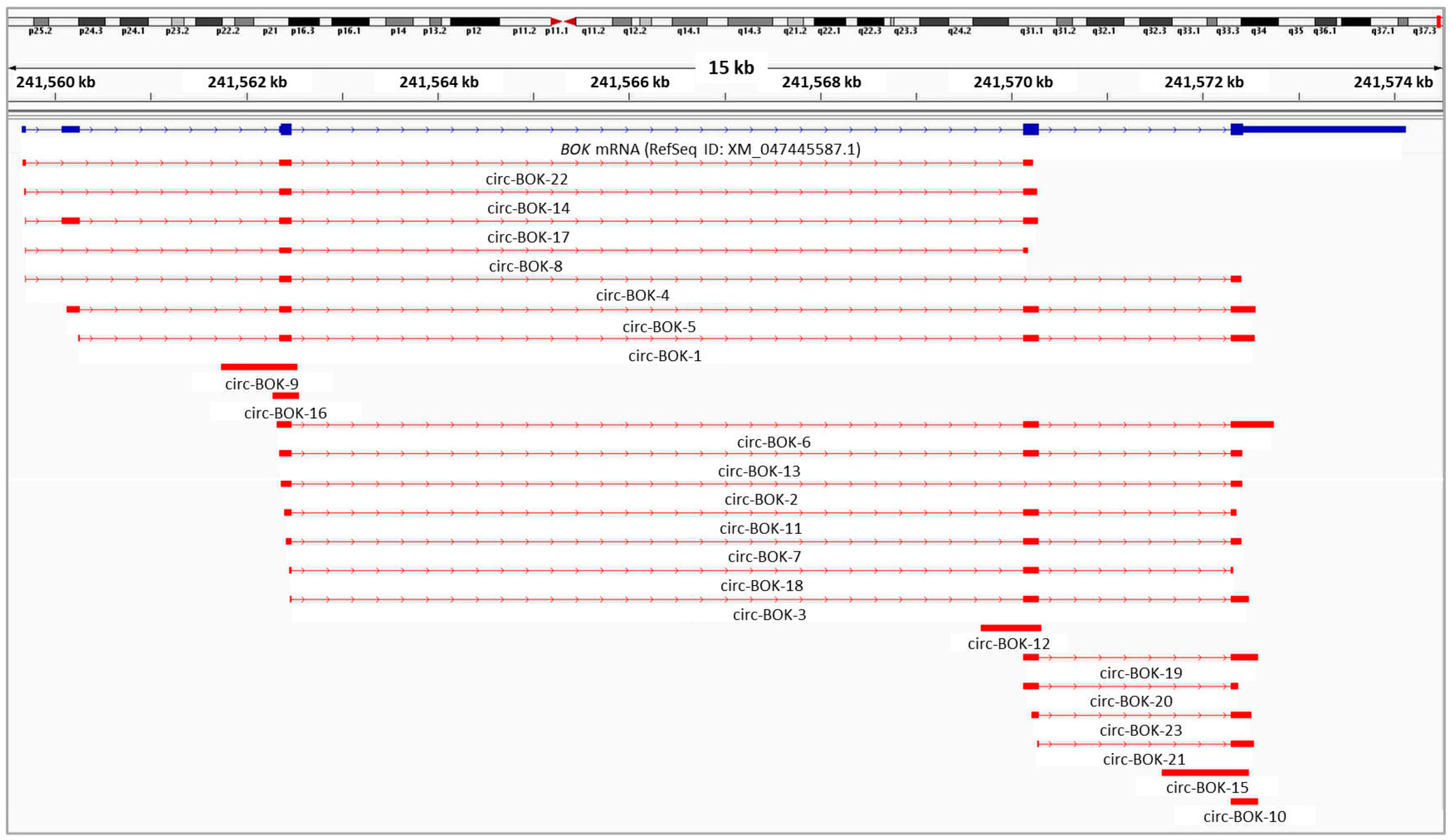{kind=link}
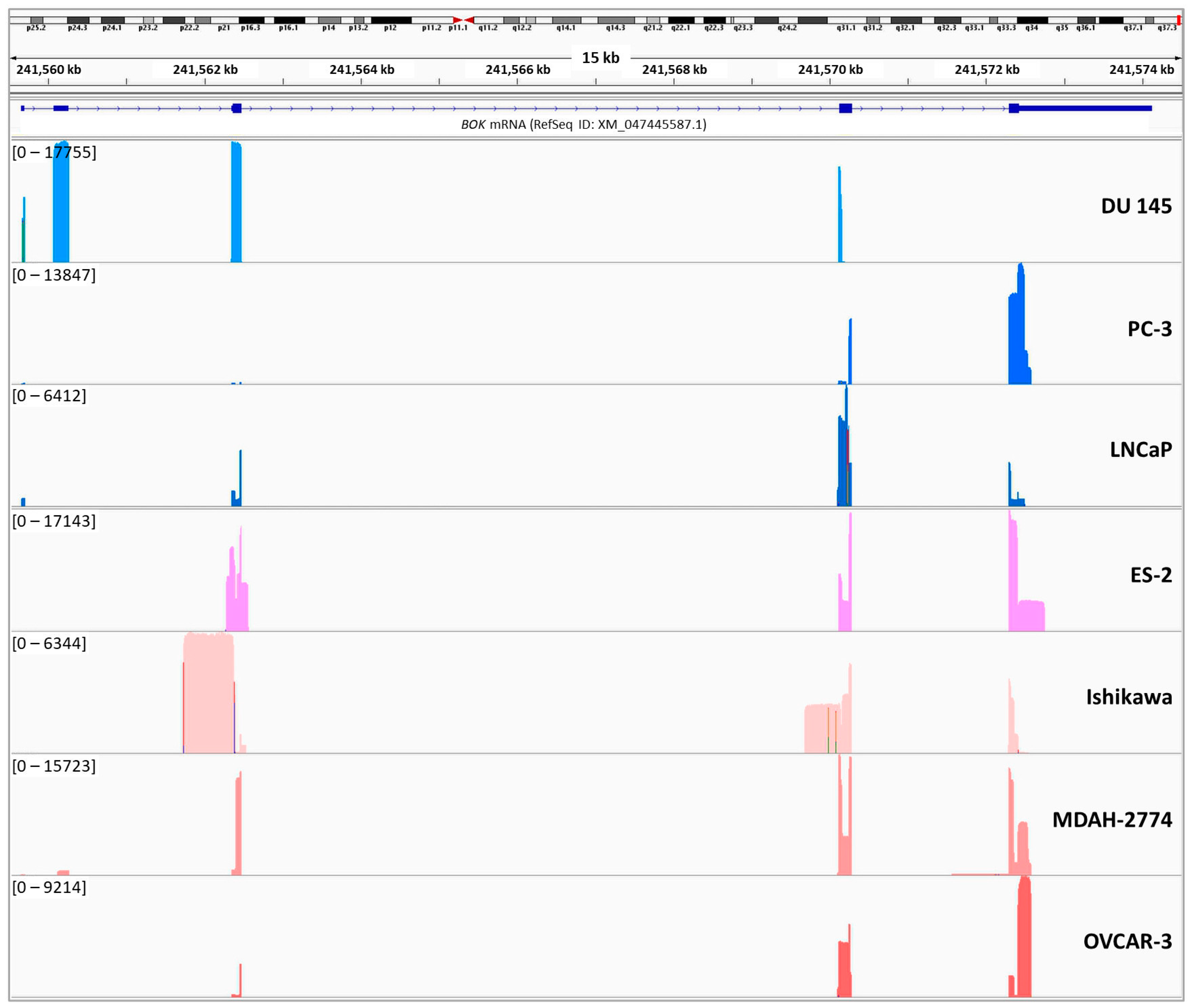{kind=link}
{kind=link}
| BOK circRNA | GenBank Accession # | Prostate Cancer Cell Lines | Ovarian Cancer Cell Lines | |||||
|---|---|---|---|---|---|---|---|---|
| DU 145 | LNCaP | PC-3 | ES-2 | Ishikawa | MDAH-2774 | OVCAR-3 | ||
| circ-BOK-1 | ON787895.1 | – | – | – | – | – | + | – |
| circ-BOK-2 | ON787896.1 | – | – | – | – | + | – | – |
| circ-BOK-3 | ON787897.1 | – | + | – | – | – | – | – |
| circ-BOK-4 | ON787898.1 | – | – | – | – | + | – | – |
| circ-BOK-5 | ON787899.1 | – | – | – | – | – | + | – |
| circ-BOK-6 | ON787900.1 | – | – | – | + | – | – | – |
| circ-BOK-7 | ON787901.1 | – | – | – | + | – | – | – |
| circ-BOK-8 | ON787902.1 | – | + | – | – | – | + | – |
| circ-BOK-9 | ON787903.1 | – | – | – | – | + | – | – |
| circ-BOK-10 | ON787904.1 | – | + | – | – | – | – | + |
| circ-BOK-11 | ON787905.1 | – | – | – | – | – | + | – |
| circ-BOK-12 | ON854799.1 | – | – | – | – | + | – | – |
| circ-BOK-13 | ON787906.1 | – | – | + | – | – | – | + |
| circ-BOK-14 | ON787907.1 | – | – | – | – | – | – | + |
| circ-BOK-15 | ON787908.1 | – | – | – | – | – | + | – |
| circ-BOK-16 | ON787909.1 | – | – | – | + | – | – | – |
| circ-BOK-17 | ON787910.1 | + | – | – | – | – | – | – |
| circ-BOK-18 | ON787911.1 | – | – | + | – | – | – | – |
| circ-BOK-19 | OR523823.1 | – | – | – | – | – | + | – |
| circ-BOK-20 | ON787912.1 | – | – | – | – | + | – | + |
| circ-BOK-21 | OR523824.1 | – | + | – | – | – | + | – |
| circ-BOK-22 | ON787913.1 | – | – | + | – | – | – | – |
| circ-BOK-23 | OQ291309.1 | – | – | + | – | – | – | – |
| BOK circRNA | miRNA Binding to circRNA | Prediction Score 1 | Binding Motif | Seed Location 2 |
|---|---|---|---|---|
| circ-BOK-2 | miR-3158-5p | 81 | UCUGCAGA | 110 |
| UCUGCAG | 184 | |||
| circ-BOK-4 | miR-3158-5p | 81 | UCUGCAGA | 133 |
| UCUGCAG | 207 | |||
| circ-BOK-6 | miR-4267 | 81 | GAGCUGGA | 29 |
| GAGCUGG | 635 | |||
| circ-BOK-9 | miR-6819-5p | 87 | CACCCCAA | 134 |
| CACCCCA | 187 | |||
| miR-6737-5p | 87 | CACCCCAA | 134 | |
| CACCCCA | 187 | |||
| miR-6812-5p | 82 | CACCCCAA | 134, 187 | |
| circ-BOK-12 | miR-466 | 90 | AUGUGUAA | 80 |
| miR-214-5p | 80 | ACAGGCA | 115 | |
| GACAGGCA | 284 | |||
| circ-BOK-15 | miR-545-3p | 89 | UUUGCUGA | 620 |
| miR-4700-5p | 87 | UCCCCAGA | 191 | |
| miR-3617-3p | 83 | UGCUGAUA | 622 | |
| miR-8089 | 83 | UCCCCAGA | 191 | |
| miR-4667-5p | 83 | UCCCCAGA | 191 |
| BOK circRNA | Putative Coding Region (Start–Stop) (nt) | Length of the Deduced Protein Sequence (aa) | Predicted m6A Site Position (nt) 1 | m6A Prediction Confidence Level 1 |
|---|---|---|---|---|
| circ-BOK-1 | 31–435 | 134 | 203/248/310 | High/High/Very high |
| 107–211 | 34 | |||
| circ-BOK-2 | 2–121 | 39 | 117 | Low |
| circ-BOK-3 | 100–306 | 68 | 74/119/181 | High/High/Very high |
| circ-BOK-4 | 25–144 | 39 | 140 | Low |
| circ-BOK-5 | 157–561 | 134 | 79/329/374/436 | Low/High/High/Very high |
| 233–337 | 34 | |||
| circ-BOK-6 | 439–609 | 56 | 210/255/317 | High/High/Very high |
| 38–442 | 134 | |||
| 114–218 | 34 | |||
| circ-BOK-7 | 19–123 | 34 | 115/160/222 | High/High/High |
| circ-BOK-9 | 457–630 | 57 | – | – |
| 92–286 | 64 | |||
| 141–491 | 116 | |||
| circ-BOK-11 | 39–143 | 34 | 135/180/242 | High/High/High |
| circ-BOK-12 | 525–617 | 30 | 277/354/499/544 | Moderate/Low/Moderate/Moderate |
| circ-BOK-13 | 89–193 | 34 | 185/230/292 | High/High/High |
| circ-BOK-14 | 117–221 | 34 | 213/258 | High/High |
| circ-BOK-15 | 55–159 | 34 | 16/203 | Moderate/High |
| 401–844 | 147 | |||
| 525–809 | 94 | |||
| 273–428 | 51 | |||
| circ-BOK-17 | 301–405 | 34 | 147/397/442 | Low/High/High |
| circ-BOK-19 | 84–290 | 68 | 58/103/165 | High/High/Very high |
| circ-BOK-22 | 127–231 | 34 | 223/268 | High/Very high |
| BOK Exon 1 | Primer Direction | Primer Sequence (5′→3′) | |
|---|---|---|---|
| First-round PCR | 2nd coding exon | Sense | CTGTGGTGACCGATGCGTTC |
| Antisense | TGCAGGGAGATGTGCAGCT | ||
| 3rd coding exon | Sense | TGCCTGGGGGAGTTCGT | |
| Antisense | CCTGCCTCACACAGTCCA | ||
| 4th coding exon | Sense | CTTCTTCGTGCTGCTGCCA | |
| Antisense | CCGAAGCTGCAGAGTGCA | ||
| Nested PCR | 2nd coding exon | Sense | TGGCTGGCCACATCTTCTC |
| Antisense | GCCACGTTGCGGTAGAC | ||
| 3rd coding exon | Sense | GCAAGACCCTGGCAACCT | |
| Antisense | GCATACAGGGACACCACCTT | ||
| 4th coding exon | Sense | GAGATGAGCTGCCCACCTG | |
| Antisense | GCTGACCACACACTTGAGGAC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kontos, C.K.; Hadjichambi, D.; Papatsirou, M.; Karousi, P.; Christodoulou, S.; Sideris, D.C.; Scorilas, A. Discovery and Comprehensive Characterization of Novel Circular RNAs of the Apoptosis-Related BOK Gene in Human Ovarian and Prostate Cancer Cells, Using Nanopore Sequencing. Non-Coding RNA 2023, 9, 57. https://doi.org/10.3390/ncrna9050057
Kontos CK, Hadjichambi D, Papatsirou M, Karousi P, Christodoulou S, Sideris DC, Scorilas A. Discovery and Comprehensive Characterization of Novel Circular RNAs of the Apoptosis-Related BOK Gene in Human Ovarian and Prostate Cancer Cells, Using Nanopore Sequencing. Non-Coding RNA. 2023; 9(5):57. https://doi.org/10.3390/ncrna9050057
Chicago/Turabian StyleKontos, Christos K., Despina Hadjichambi, Maria Papatsirou, Paraskevi Karousi, Spyridon Christodoulou, Diamantis C. Sideris, and Andreas Scorilas. 2023. "Discovery and Comprehensive Characterization of Novel Circular RNAs of the Apoptosis-Related BOK Gene in Human Ovarian and Prostate Cancer Cells, Using Nanopore Sequencing" Non-Coding RNA 9, no. 5: 57. https://doi.org/10.3390/ncrna9050057
APA StyleKontos, C. K., Hadjichambi, D., Papatsirou, M., Karousi, P., Christodoulou, S., Sideris, D. C., & Scorilas, A. (2023). Discovery and Comprehensive Characterization of Novel Circular RNAs of the Apoptosis-Related BOK Gene in Human Ovarian and Prostate Cancer Cells, Using Nanopore Sequencing. Non-Coding RNA, 9(5), 57. https://doi.org/10.3390/ncrna9050057