Long Non-Coding RNAs Associated with Heterochromatin Function in Immune Cells in Psychosis

 ,
,
Abstract
:1. Introduction
2. Results
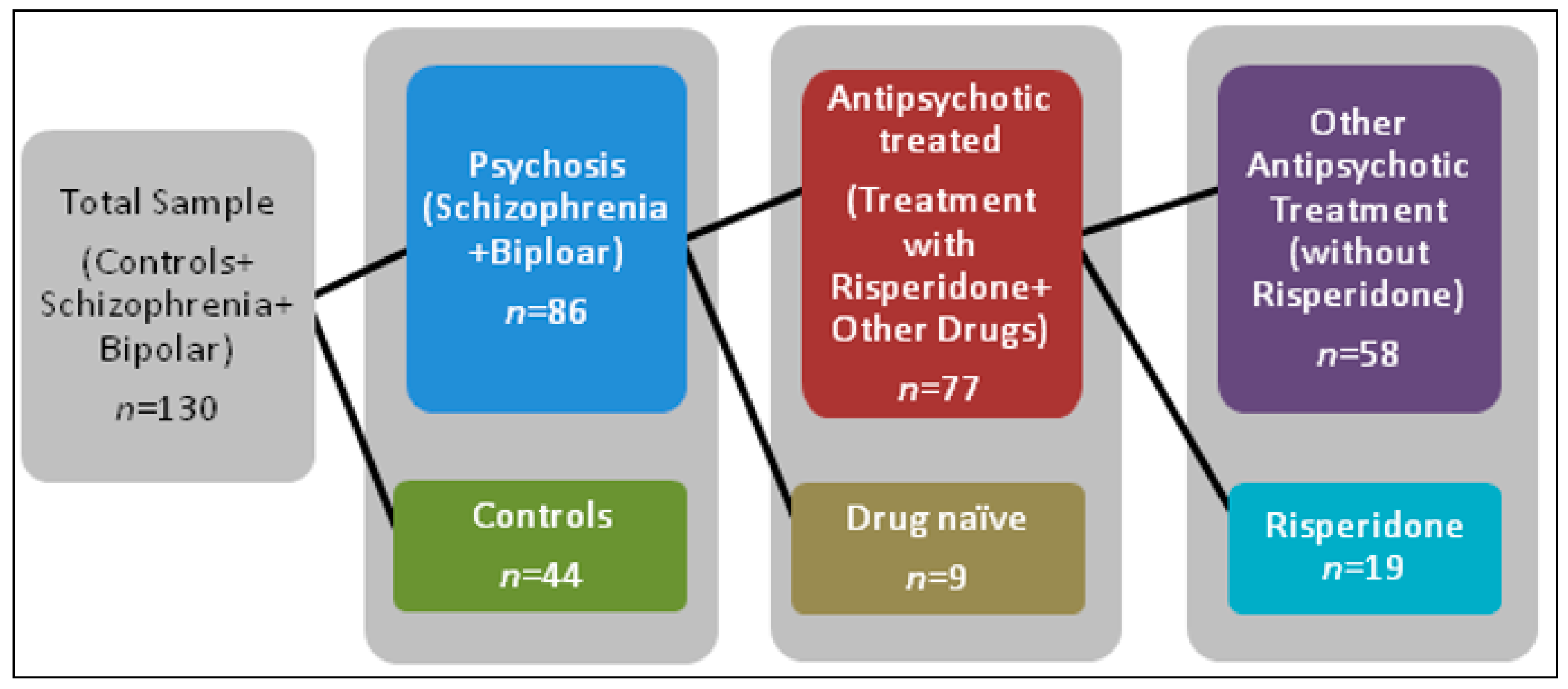
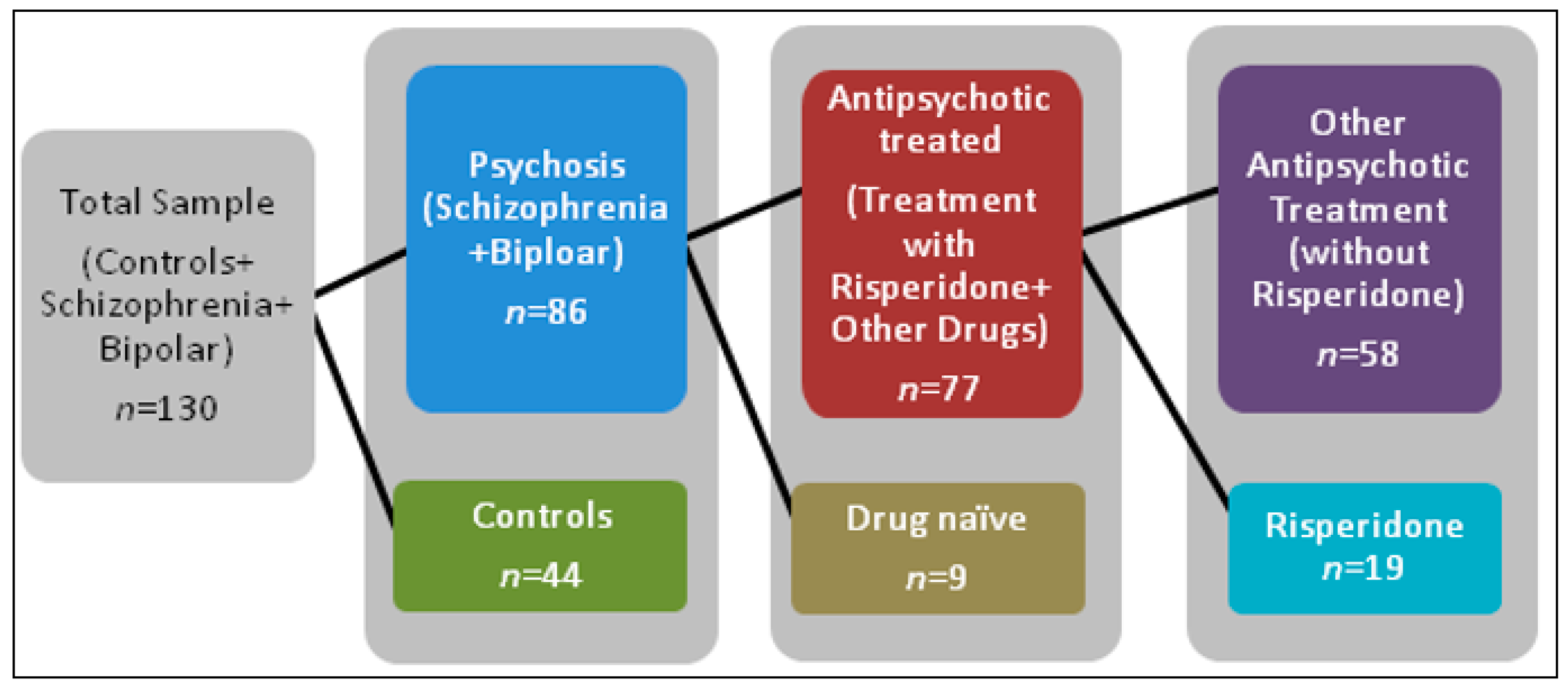
2.1. Demographics
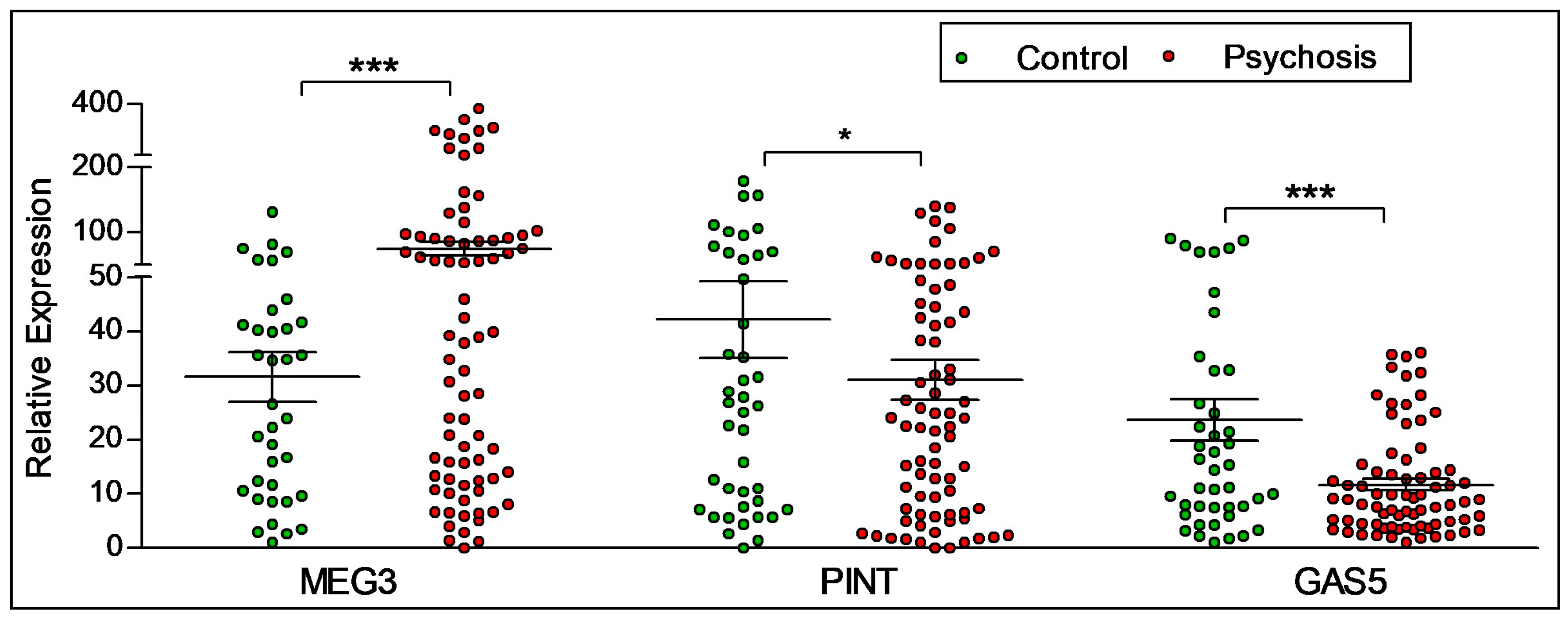
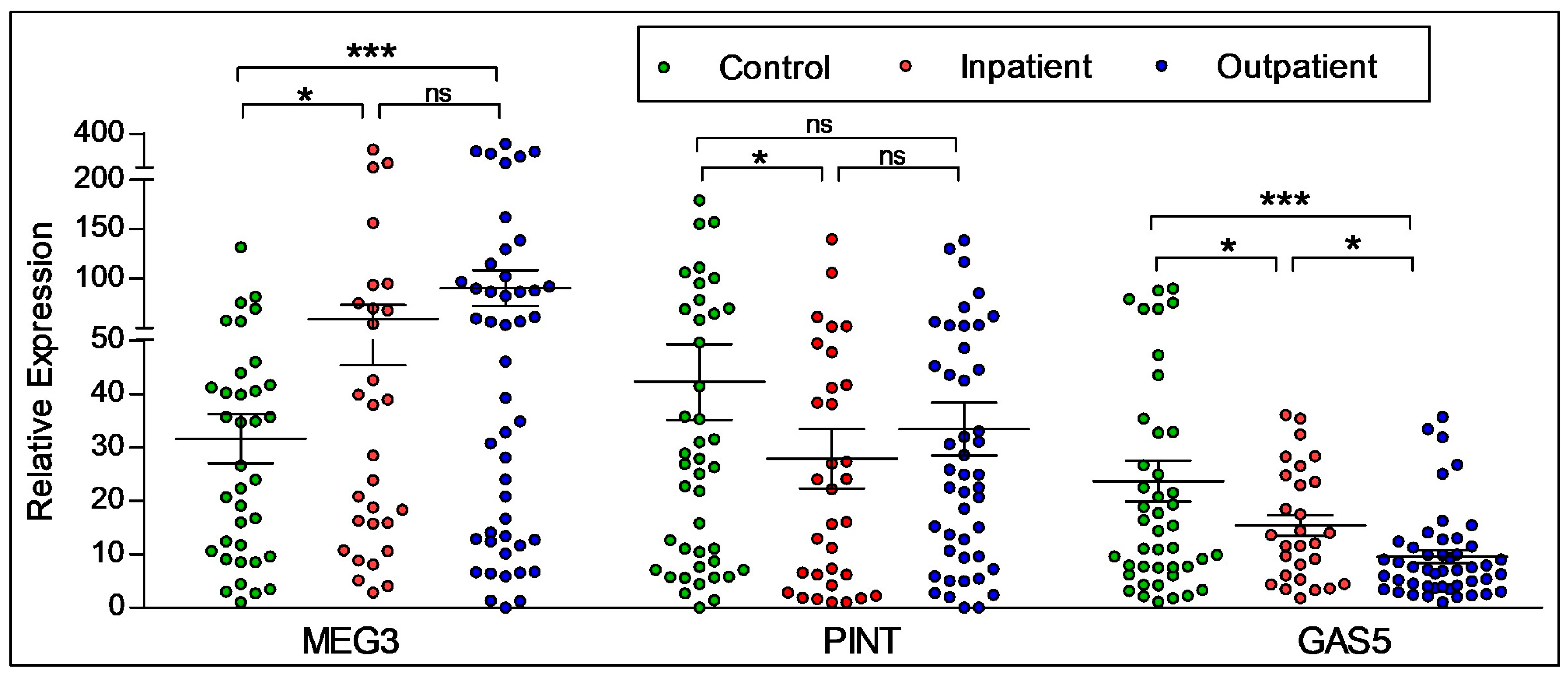
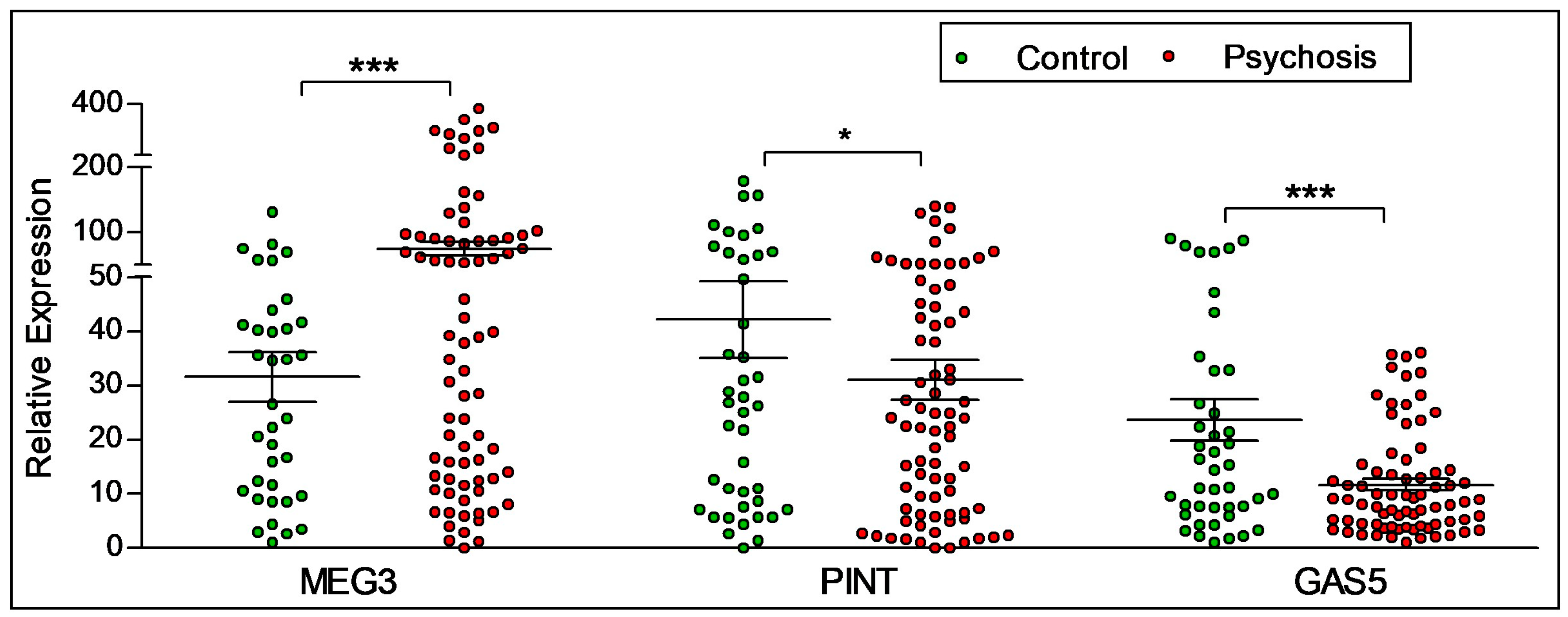
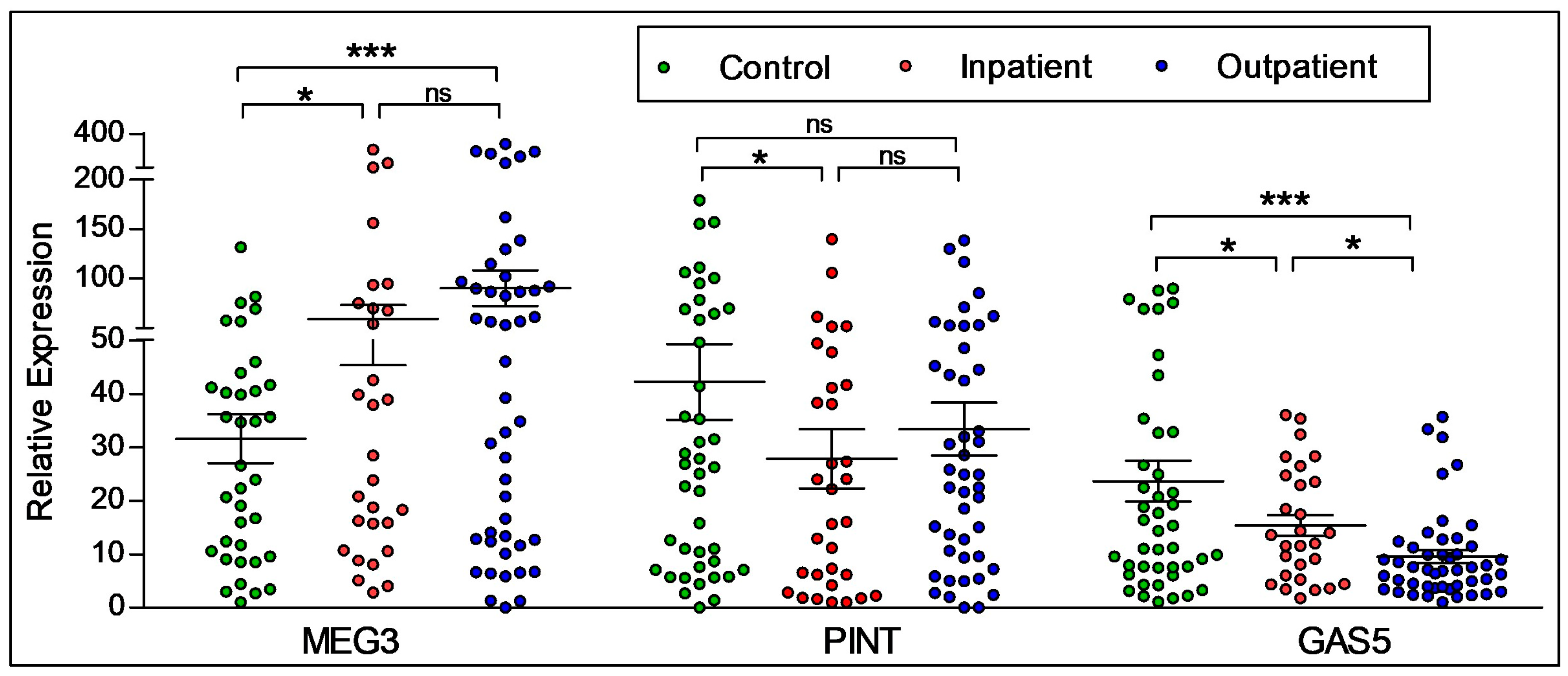
2.2. Diagnostic Differences
2.3. Co-Expression of lncRNAs
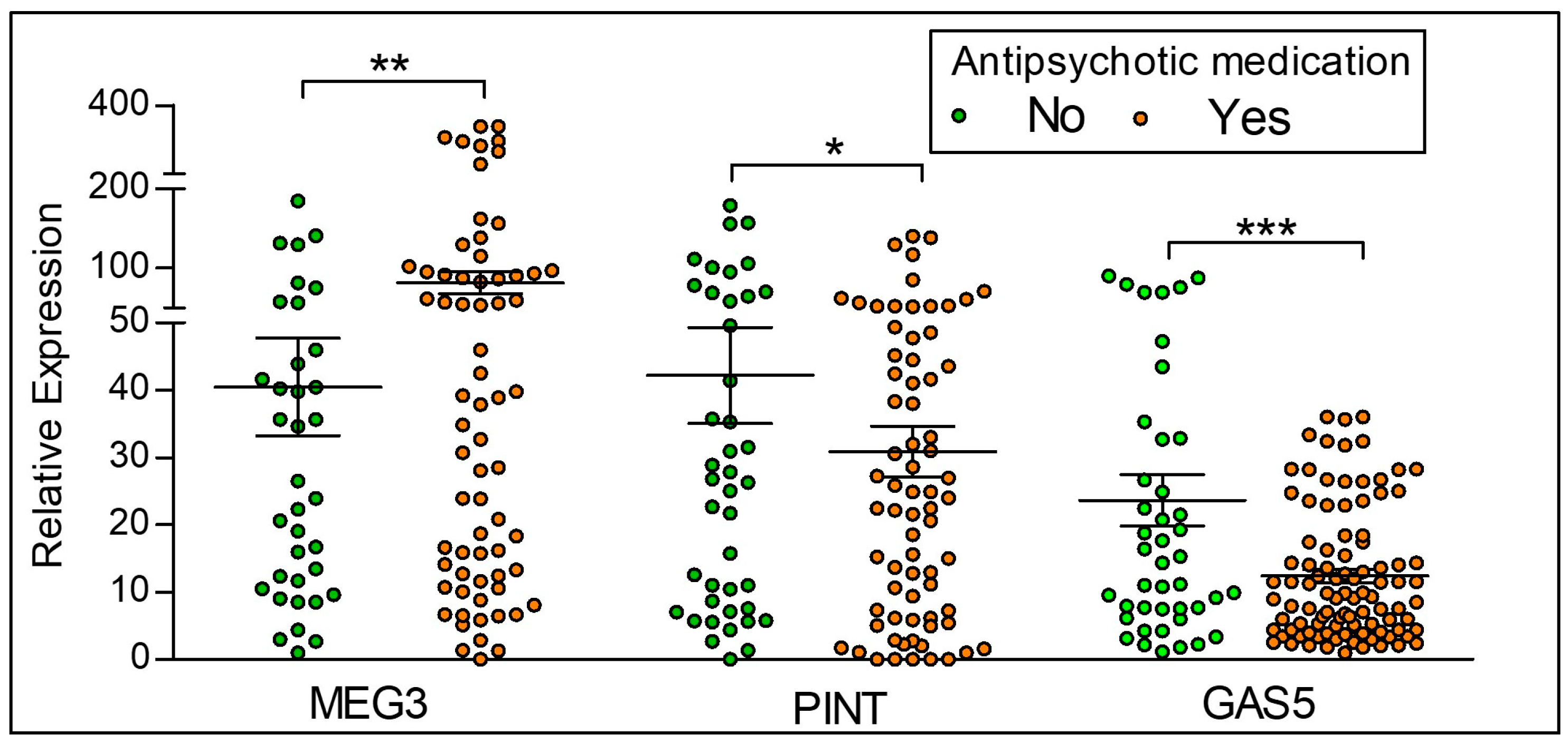
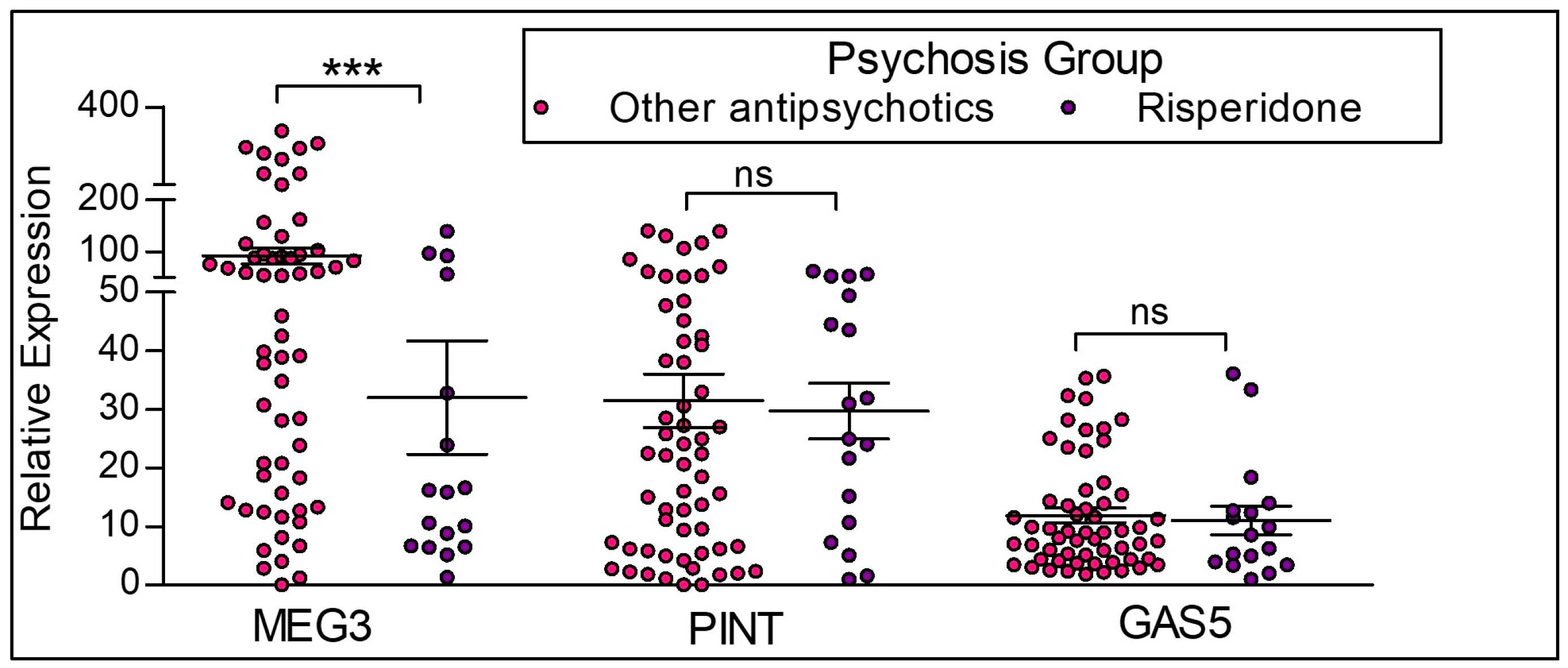
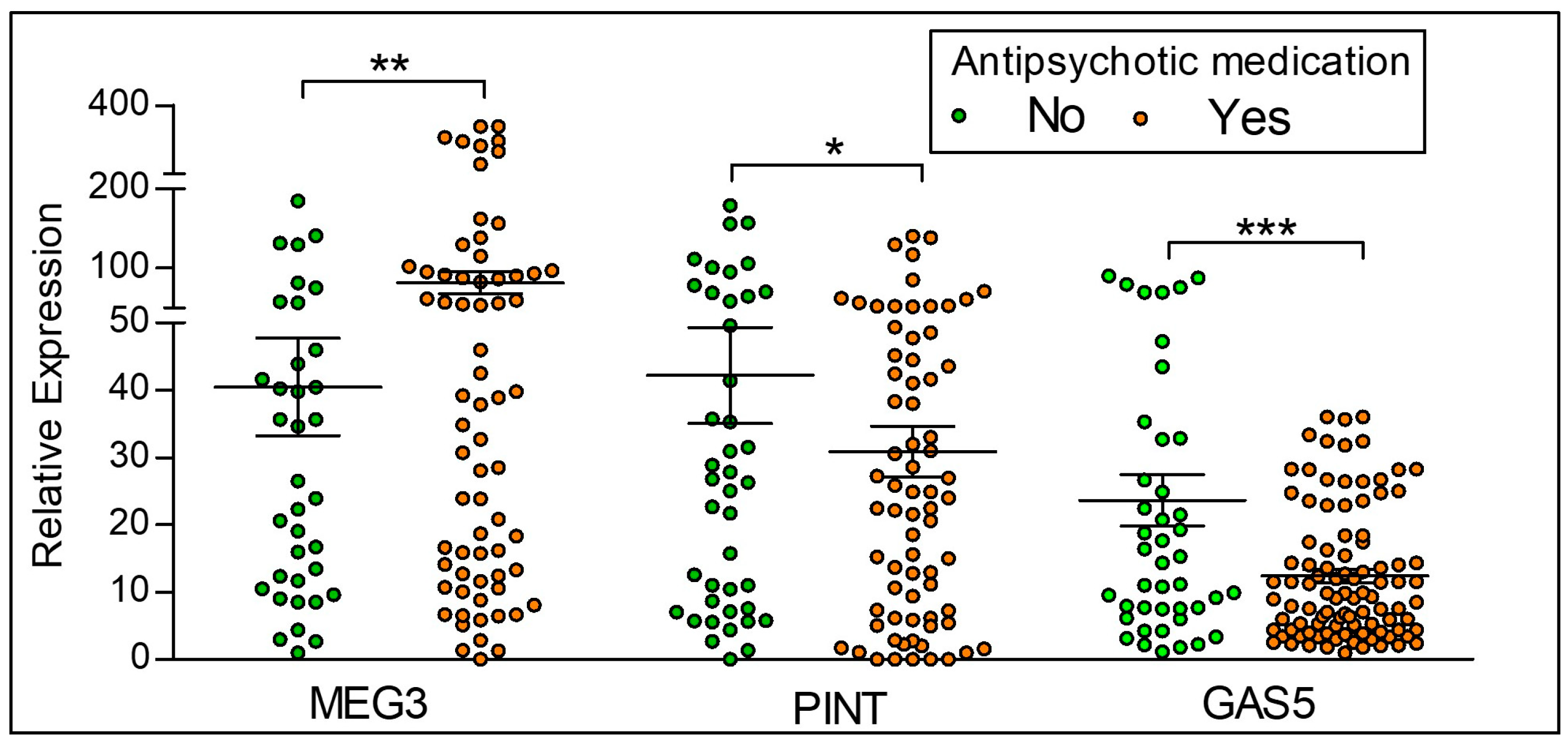
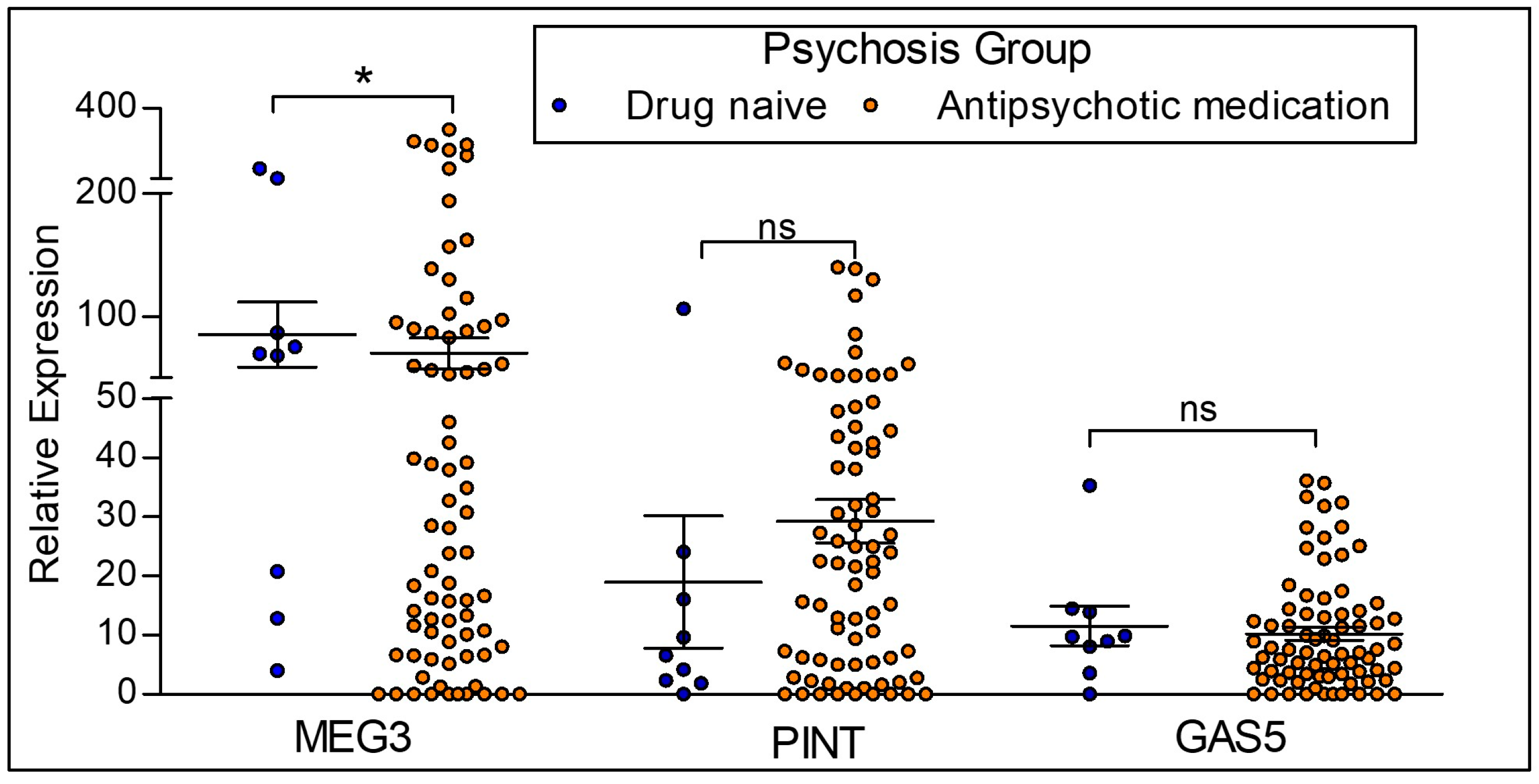
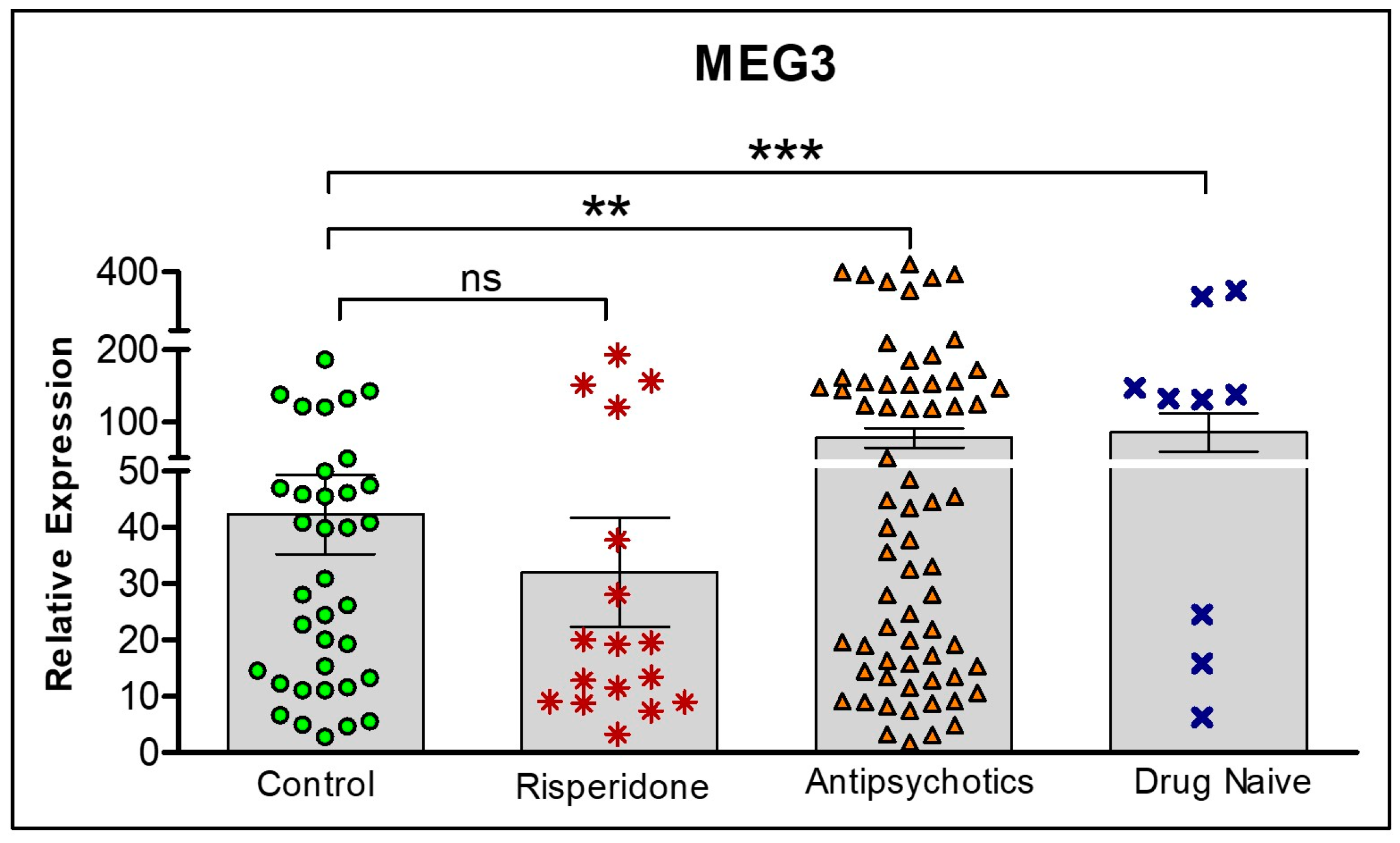
2.4. Differences in the Expression of lncRNAs Based on Antipsychotic Treatment Status in the Clinical Sample
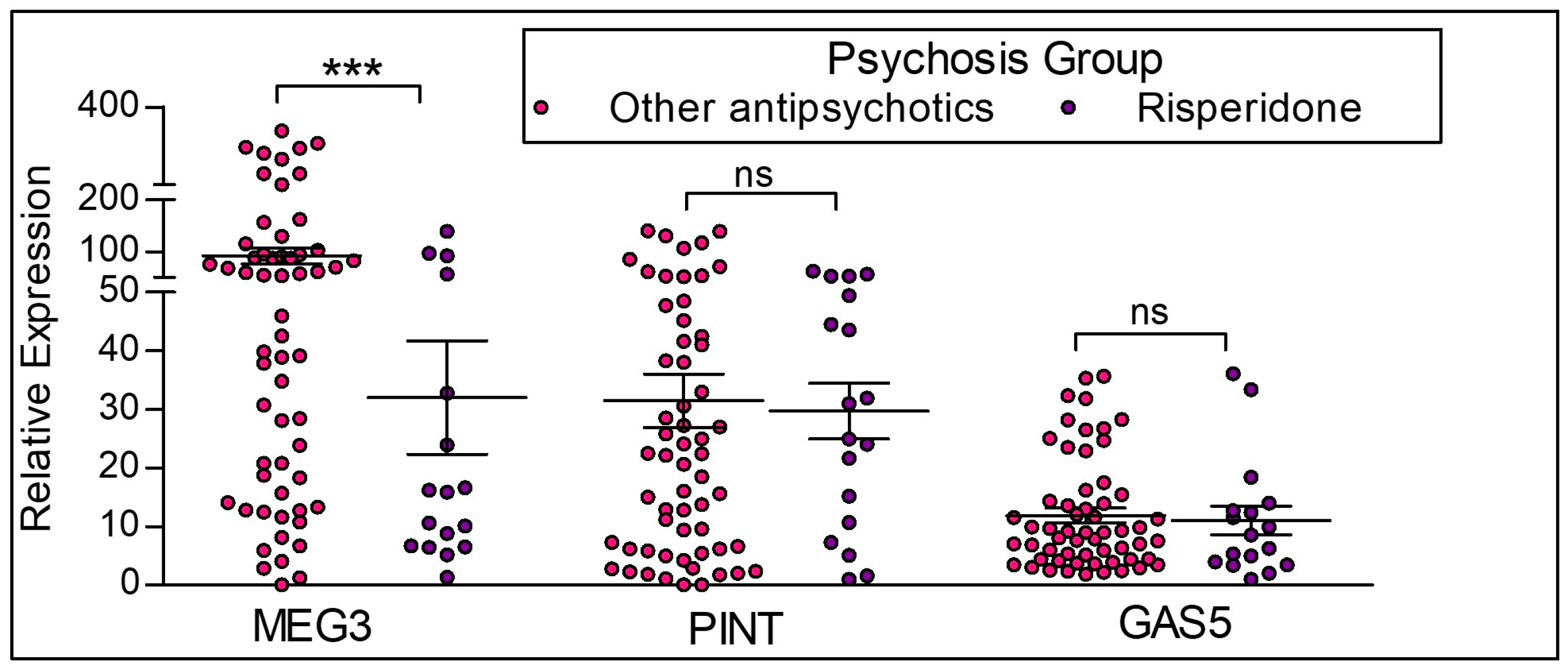
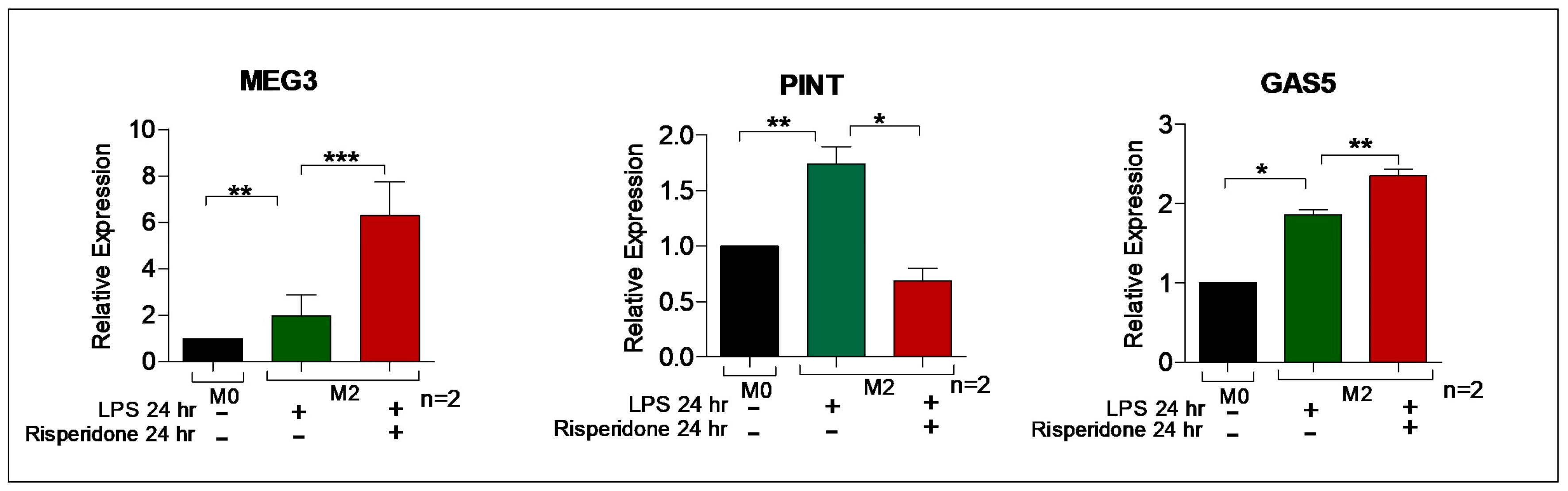
2.5. Effect of Antipsychotics on the expression of lncRNAs in the M0 and M2 macrophages
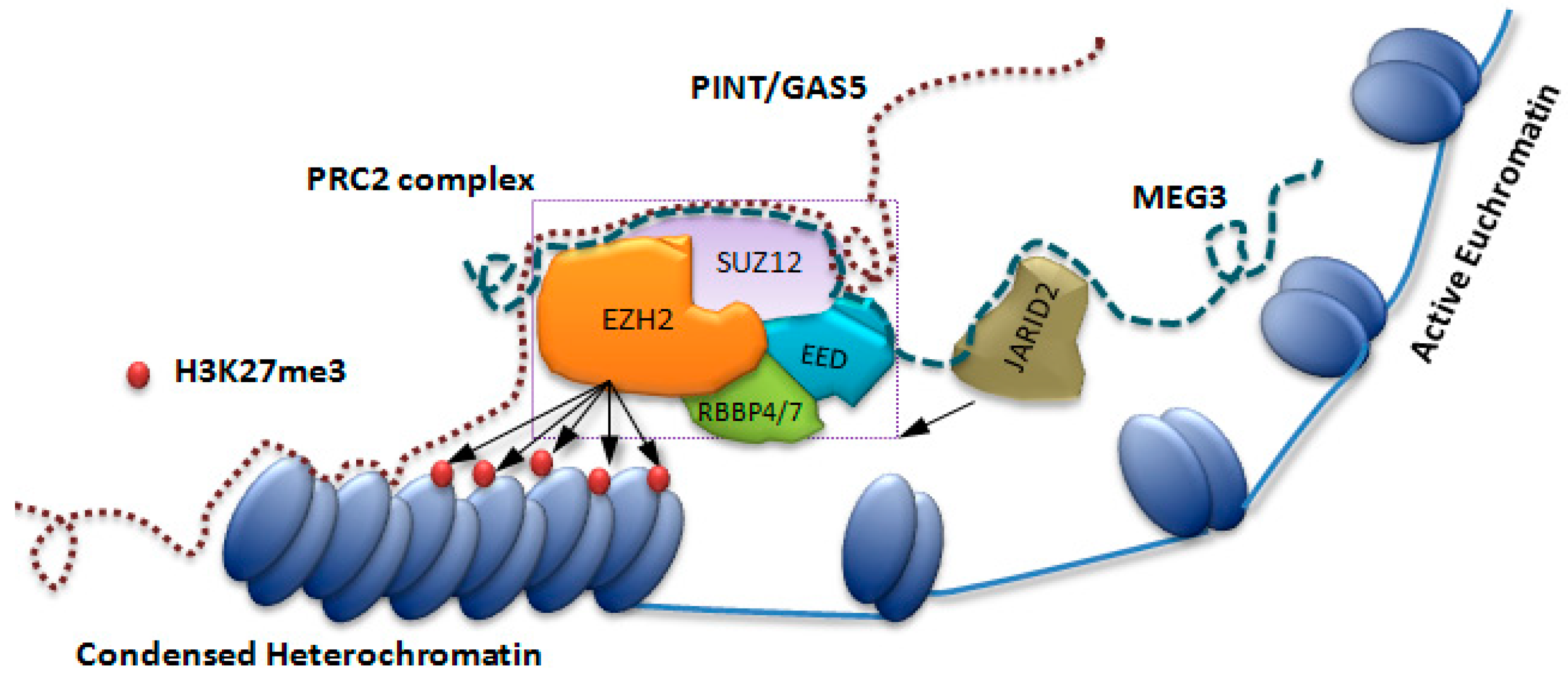
3. Discussion
4. Materials and Methods
4.1. Participant Information and Clinical Measures
4.2. Sample Collection, Processing and Quantitative real-time PCR (qRT-PCR)
4.3. Primary Clinical Measures
4.4. THP1-Derived M2tol Polarized Macrophages and Risperidone Treatment
4.5. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Beumer, W.; Gibney, S.M.; Drexhage, R.C.; Pont-Lezica, L.; Doorduin, J.; Klein, H.C.; Steiner, J.; Connor, T.J.; Harkin, A.; Versnel, M.A.; et al. The immune theory of psychiatric diseases: A key role for activated microglia and circulating monocytes. J. Leukoc. Biol. 2012, 92, 959–975. [Google Scholar] [CrossRef] [PubMed]
- Goldsmith, D.R.; Rapaport, M.H.; Miller, B.J. A meta-analysis of blood cytokine network alterations in psychiatric patients: Comparisons between schizophrenia, bipolar disorder and depression. Mol. Psychiatry 2016, 21, 1696–1709. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, S.; Fitzgerald, K.A. Cytokines and Long Noncoding RNAs. Cold Spring Harb. Perspect. Biol. 2018, 10, a028589. [Google Scholar] [CrossRef] [PubMed]
- Marchese, F.P.; Raimondi, I.; Huarte, M. The multidimensional mechanisms of long noncoding RNA function. Genome Biol. 2017, 18, 206. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.P.; Rosen, C.; Kartan, S.; Guidotti, A.; Costa, E.; Grayson, D.R.; Chase, K. Valproic acid and chromatin remodeling in schizophrenia and bipolar disorder: Preliminary results from a clinical population. Schizophr. Res. 2006, 88, 227–231. [Google Scholar] [CrossRef] [PubMed]
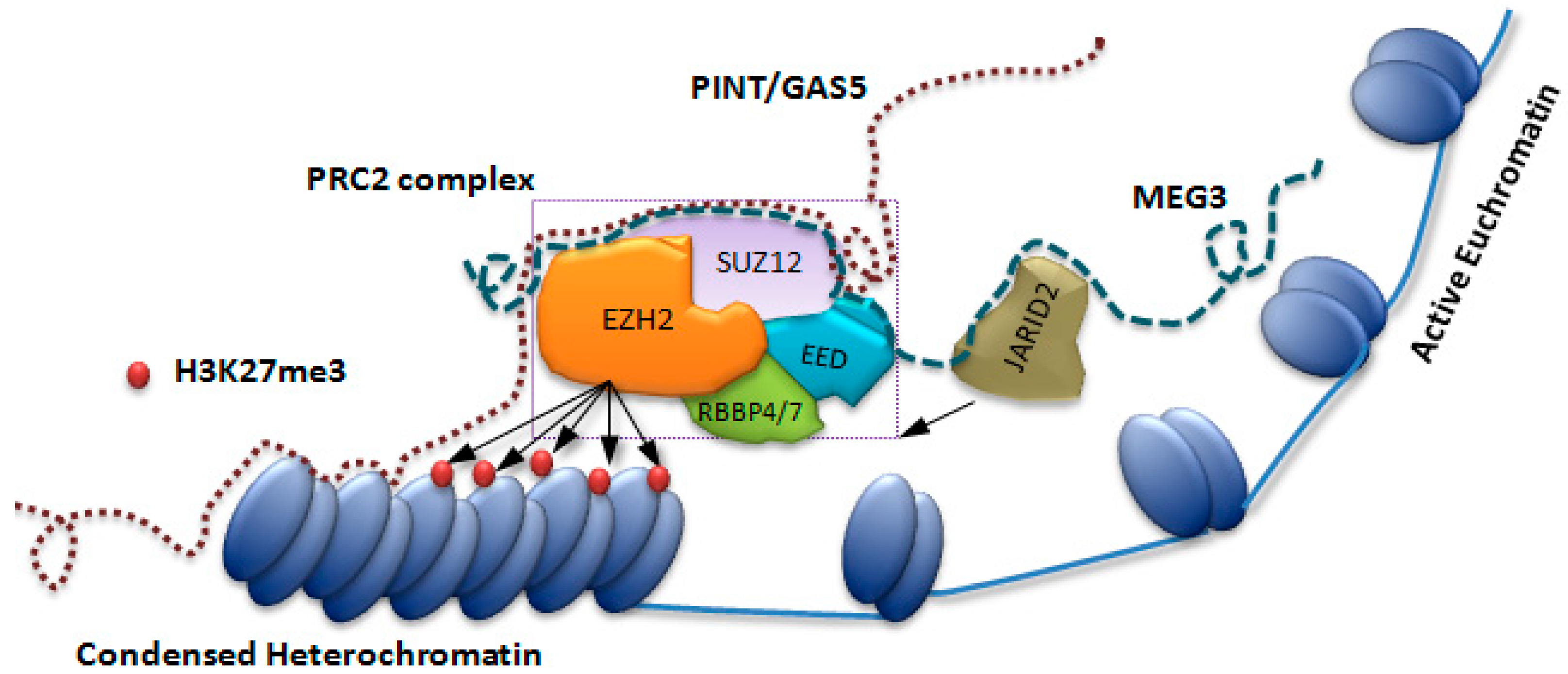
- Margueron, R.; Reinberg, D. The Polycomb complex PRC2 and its mark in life. Nature 2011, 469, 343–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, M.-C.; Manor, O.; Wan, Y.; Mosammaparast, N.; Wang, J.K.; Lan, F.; Shi, Y.; Segal, E.; Chang, H.Y. Long Noncoding RNA as Modular Scaffold of Histone Modification Complexes. Science 2010, 329, 689–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.C.; Chang, H.Y. Molecular Mechanisms of Long Noncoding RNAs. Mol. Cell 2011, 43, 904–914. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, S.; Bonasio, R.; Saldaña-Meyer, R.; Yoshida, T.; Son, J.; Nishino, K.; Umezawa, A.; Reinberg, D. Interactions between JARID2 and Noncoding RNAs Regulate PRC2 Recruitment to Chromatin. Mol. Cell 2014, 53, 290–300. [Google Scholar] [CrossRef] [PubMed]
- Melbourne, J.K.; Chase, K.A.; Feiner, B.; Rosen, C.; Sharma, R.P. Long non-coding and endogenous retroviral RNA levels are associated with proinflammatory cytokine mRNA expression in peripheral blood cells: Implications for schizophrenia. Psychiatry Res. 2018, 262, 465–468. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.P.; University of Illinois at Chicago, Chicago, IL, USA. Genome Wide Sequencing of RNA in CD14 Monocytes. Personal communication, 2017. [Google Scholar]
- Mondal, T.; Subhash, S.; Vaid, R.; Enroth, S.; Uday, S.; Reinius, B.; Mitra, S.; Mohammed, A.; James, A.R.; Hoberg, E.; et al. MEG3 long noncoding RNA regulates the TGF-β pathway genes through formation of RNA–DNA triplex structures. Nat. Commun. 2015, 6, 7743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marín-Béjar, O.; Marchese, F.P.; Athie, A.; Sánchez, Y.; González, J.; Segura, V.; Huang, L.; Moreno, I.; Navarro, A.; Monzó, M.; et al. Pint lincRNA connects the p53 pathway with epigenetic silencing by the Polycomb repressive complex 2. Genome Biol. 2013, 14, R104. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Guo, Z.; Zhao, Y.; Jin, Y.; An, L.; Wu, B.; Liu, Z.; Chen, X.; Chen, X.; Zhou, H.; et al. Genetic polymorphisms of lncRNA-p53 regulatory network genes are associated with concurrent chemoradiotherapy toxicities and efficacy in nasopharyngeal carcinoma patients. Sci. Rep. 2017, 7, 8320. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Yu, Z.; Fang, X.; Liu, M.; Pu, Y.; Shao, Q.; Wang, D.; Zhao, X.; Huang, A.; Xiang, Z.; et al. LncRNA GAS5 inhibits microglial M2 polarization and exacerbates demyelination. EMBO Rep. 2017, 18, 1801–1816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pawar, K.; Hanisch, C.; Palma Vera, S.E.; Einspanier, R.; Sharbati, S. Down regulated lncRNA MEG3 eliminates mycobacteria in macrophages via autophagy. Sci. Rep. 2016, 6, 19416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Zhang, X.; Zhou, Y.; Ansell, P.J.; Klibanski, A. Cyclic AMP stimulates MEG3 gene expression in cells through a cAMP-response element (CRE) in the MEG3 proximal promoter region. Int. J. Biochem. Cell Biol. 2006, 38, 1808–1820. [Google Scholar] [CrossRef] [PubMed]
- Kino, T.; Hurt, D.E.; Ichijo, T.; Nader, N.; Chrousos, G.P. Noncoding RNA Gas5 Is a Growth Arrest- and Starvation-Associated Repressor of the Glucocorticoid Receptor. Sci. Signal. 2010, 3, ra8. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Chen, X.; Zhang, X.; Duan, X.; Pan, T.; Hu, Q.; Zhang, Y.; Zhong, F.; Liu, J.; Zhang, H.; et al. An Lnc RNA (GAS5)/snoRNA-derived piRNA induces activation of TRAIL gene by site-specifically recruiting MLL/COMPASS-like complexes. Nucleic Acids Res. 2015, 43, 3712–3725. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhong, Y.; Wang, Y.; Zhang, X.; Batista, D.L.; Gejman, R.; Ansell, P.J.; Zhao, J.; Weng, C.; Klibanski, A. Activation of p53 by MEG3 non-coding RNA. J. Biol. Chem. 2007, 282, 24731–24742. [Google Scholar] [CrossRef] [PubMed]
- Prinz, M.; Priller, J. The role of peripheral immune cells in the CNS in steady state and disease. Nat. Neurosci. 2017, 20, 136–144. [Google Scholar] [CrossRef] [PubMed]
- Herz, J.; Filiano, A.J.; Smith, A.; Yogev, N.; Kipnis, J. Myeloid Cells in the Central Nervous System. Immunity 2017, 46, 943–956. [Google Scholar] [CrossRef] [PubMed]
- Nitsche, A.; Stadler, P.F. Evolutionary clues in lncRNAs. Wiley Interdiscip. Rev. Rna 2017, 8, e1376. [Google Scholar] [CrossRef] [PubMed]
- DSM-IV-TR Casebook: A Learning Companion to the Diagnostic and Statistical Manual of Mental Disorders; Spitzer, R.L.; Gibbon, M.; Skodol, A.E.; Williams, J.B.W.; First, M.B. (Eds.) American Psychiatric Publishing, Inc.: Arlington, VA, USA, 2002; Volume 1, ISBN 1-58562-266-4. [Google Scholar]
- Chase, K.A.; Feiner, B.; Rosen, C.; Gavin, D.P.; Sharma, R.P. Characterization of peripheral cannabinoid receptor expression and clinical correlates in schizophrenia. Psychiatry Res. 2016, 245, 346–353. [Google Scholar] [CrossRef] [PubMed]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3. [Google Scholar] [CrossRef] [Green Version]
- Kay, S.R.; Fiszbein, A.; Opler, L.A. The Positive and Negative Syndrome Scale (PANSS) for Schizophrenia. Schizophr. Bull. 1987, 13, 261–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindenmayer, J.P.; Bernstein-Hyman, R.; Grochowski, S. Five-factor model of schizophrenia. Initial validation. J. Nerv. Ment. Dis. 1994, 182, 631–638. [Google Scholar] [CrossRef] [PubMed]
- Lehoux, C.; Gobeil, M.-H.; Lefèbvre, A.-A.; Maziade, M.; Roy, M.-A. The Five-Factor Structure of the PANSS: A Critical Review of its Consistency Across Studies. Clin. Schizophr. Relat. Psychoses 2009, 3, 103–110. [Google Scholar] [CrossRef]
- Genin, M.; Clement, F.; Fattaccioli, A.; Raes, M.; Michiels, C. M1 and M2 macrophages derived from THP-1 cells differentially modulate the response of cancer cells to etoposide. BMC Cancer 2015, 15, 577. [Google Scholar] [CrossRef] [PubMed]
- Pena, O.M.; Pistolic, J.; Raj, D.; Fjell, C.D.; Hancock, R.E.W. Endotoxin Tolerance Represents a Distinctive State of Alternative Polarization (M2) in Human Mononuclear Cells. J. Immunol. 2011, 186, 7243–7254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porta, C.; Rimoldi, M.; Raes, G.; Brys, L.; Ghezzi, P.; Di Liberto, D.; Dieli, F.; Ghisletti, S.; Natoli, G.; De Baetselier, P.; et al. Tolerance and M2 (alternative) macrophage polarization are related processes orchestrated by p50 nuclear factor B. Proc. Natl. Acad. Sci. USA 2009, 106, 14978–14983. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | Psychosis | Group Differences | ||||
|---|---|---|---|---|---|---|
| Total (n) | 44 (33.8%) | 86 (66.2%) | ||||
| Sex | Female (n) | 25 (41.0%) | 36 (59.0%) | χ2(3) = 2.615, p = 0.106 | ||
| Male (n) | 19 (27.5%) | 50 (72.5%) | ||||
| Race | Caucasian, non-Hispanic (n) | 12 (54.5%) | 10 (45.5%) | χ2(3) = 22.494, p < 0.001 | ||
| Black, non-Hispanic (n) | 18 (22.5%) | 62 (77.5%) | ||||
| Asian or other Pacific Islander (n) | 10 (83.3%) | 2 (16.7%) | ||||
| Hispanic (n) | 4 (25.0%) | 12 (75.0%) | ||||
| Antipsychotic Use | Yes (n) | 0 | 77 | |||
| No (n) | 44 | 9 | ||||
| Other medication * | Yes (n) | 4 | 9 | |||
| No (n) | 40 | 77 | ||||
| Smoking | Yes(n) | 7 | 39 | χ2(3) = 11.034, p = 0.001 | ||
| No (n) | 37 | 47 | ||||
| Age (Mean ± SD) | 38.77 ± 12.82 | 40.21 ± 12.98 | t(128) = −0.600, p = 0.55 | |||
| BMI (Mean ± SD) | 28.93 ± 8.1 | 30.8 ± 8.15 | t(128) = 1.241, p = 0.217 | |||
| Mean | SD | Mean | SD | |||
| Age at onset of illness | ~ | ~ | 21.32 | 7.88 | ||
| Duration of untreated psychosis | ~ | ~ | 2.97 | 5.85 | ||
| Duration of illness | ~ | ~ | 19.33 | 14.35 | ||
| N = 130 | Control (n = 44) | Psychosis (n = 86) | Group Differences | |||||
|---|---|---|---|---|---|---|---|---|
| Inpatient(n = 33) | Outpatient(n = 53) | |||||||
| PANSS Five-Factor Scores | Mean ± SD | Range | Mean ± SD | Range | Mean ± SD | Range | Inpatient and Outpatient † | Control, Inpatient and Outpatient ‡ |
| Positive | 4.11 ± 0.49 | 4–7 | 18.27 ± 2.39 | 13–23 | 13.49 ± 4.40 | 5–21 | t(84) = 5.73 *** | F(2,127) = 218.20 *** |
| Negative | 6.55 ± 7.56 | 6–15 | 20.76 ± 7.56 | 6–36 | 15.60 ± 5.91 | 6–27 | t(84) = 3.58 *** | F(2,127) = 68.56 *** |
| Cognitive | 5.48 ± 1.07 | 5–10 | 17.34 ± 4.09 | 8–25 | 12.00 ± 4.01 | 5–21 | t(84) = 5.95 *** | F(2,127) = 121.25 *** |
| Excitement | 5.66 ± 1.98 | 4–13 | 10.91 ± 3.41 | 6–28 | 9.34 ± 2.78 | 4–16 | t(84) = 2.33 * | F(2,127) = 39.35 *** |
| Depression | 7.55 ± 2.71 | 5–14 | 15.69 ± 2.80 | 10–23 | 13.83 ± 3.33 | 6–22 | t(84) = 2.68 ** | F(2,127) = 83.173 *** |
| PINT | GAS5 | |
|---|---|---|
| MEG3 | * r = 0.237 p = 0.015 | * r = 0.080 p = 0.422 |
| PINT | * r = 0.222 p = 0.016 |
| GAPDH | 5′-CGAGATCCCTCCAAAATCAA-3′ | 5′-TTCACACCCATGACGAACAT-3′ |
| ACTB | 5′s-TGAAGGTAGTTTCGTGGATGC-3′ | 5′-TCCCTGGAGAAGAGCTACGA-3′ |
| MEG3 | 5′-GCGGAGAGCAGAGAGGGA-3′ | 5′-AGGAGAGACCCGGGTGAG-3′ |
| PINT | 5′-CCATCTGGAGTTTCTCTGCCT-3′ | 5′-GGTAAGACTCTGTCTTCAGCTGTTA-3′ |
| GAS-5 | 5′-TCCTGTGAGGTATGGTGCTGG-3′ | 5-AACTTGCCTGGACCAGCTTA-3′ |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sudhalkar, N.; Rosen, C.; Melbourne, J.K.; Park, M.R.; Chase, K.A.; Sharma, R.P. Long Non-Coding RNAs Associated with Heterochromatin Function in Immune Cells in Psychosis. Non-Coding RNA 2018, 4, 43. https://doi.org/10.3390/ncrna4040043
Sudhalkar N, Rosen C, Melbourne JK, Park MR, Chase KA, Sharma RP. Long Non-Coding RNAs Associated with Heterochromatin Function in Immune Cells in Psychosis. Non-Coding RNA. 2018; 4(4):43. https://doi.org/10.3390/ncrna4040043
Chicago/Turabian StyleSudhalkar, Niyati, Cherise Rosen, Jennifer K. Melbourne, Mi Rae Park, Kayla A. Chase, and Rajiv P. Sharma. 2018. "Long Non-Coding RNAs Associated with Heterochromatin Function in Immune Cells in Psychosis" Non-Coding RNA 4, no. 4: 43. https://doi.org/10.3390/ncrna4040043