
Self-Assembling Peptide Hydrogels as Functional Tools to Tackle Intervertebral Disc Degeneration



Abstract
:1. Introduction
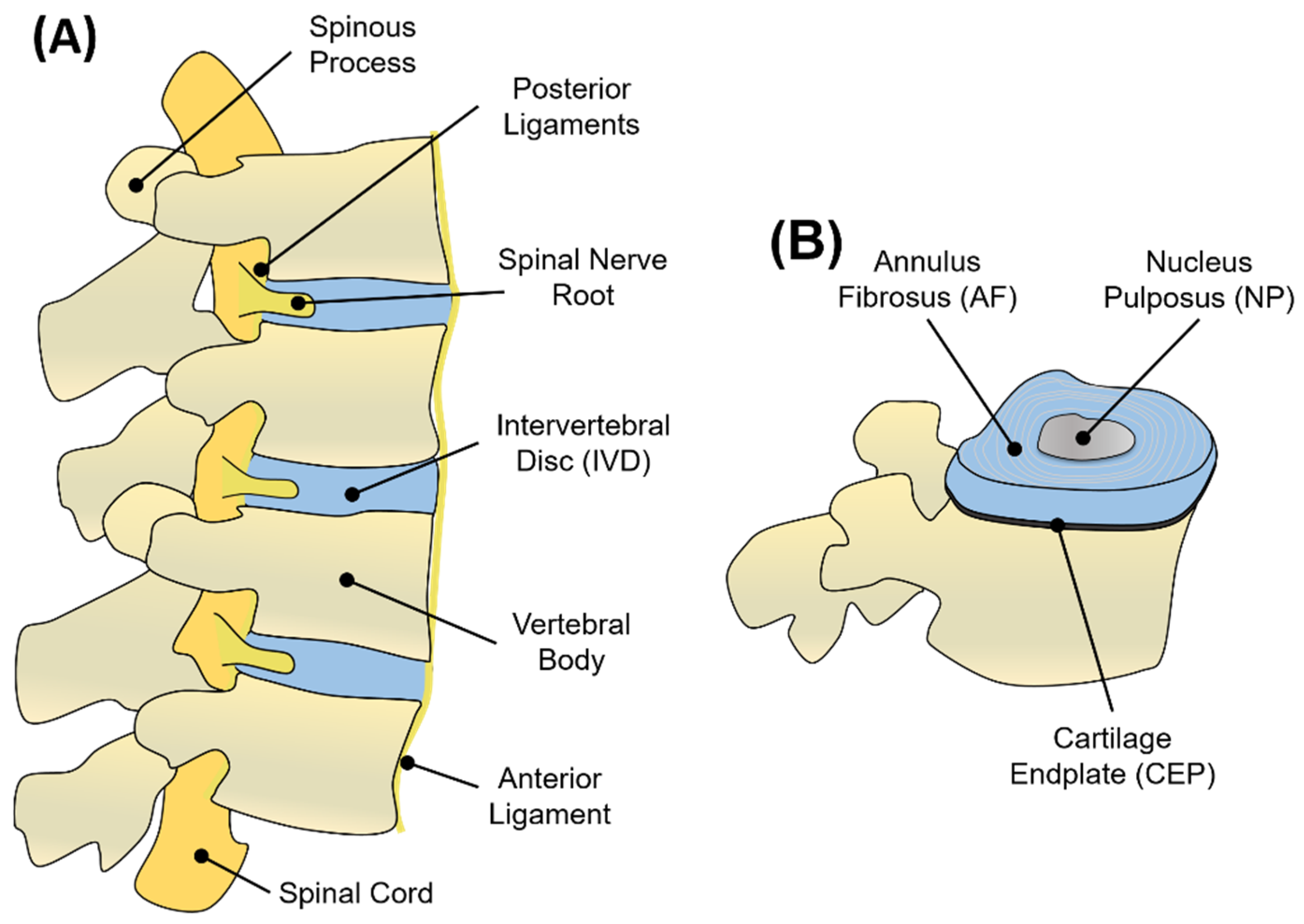
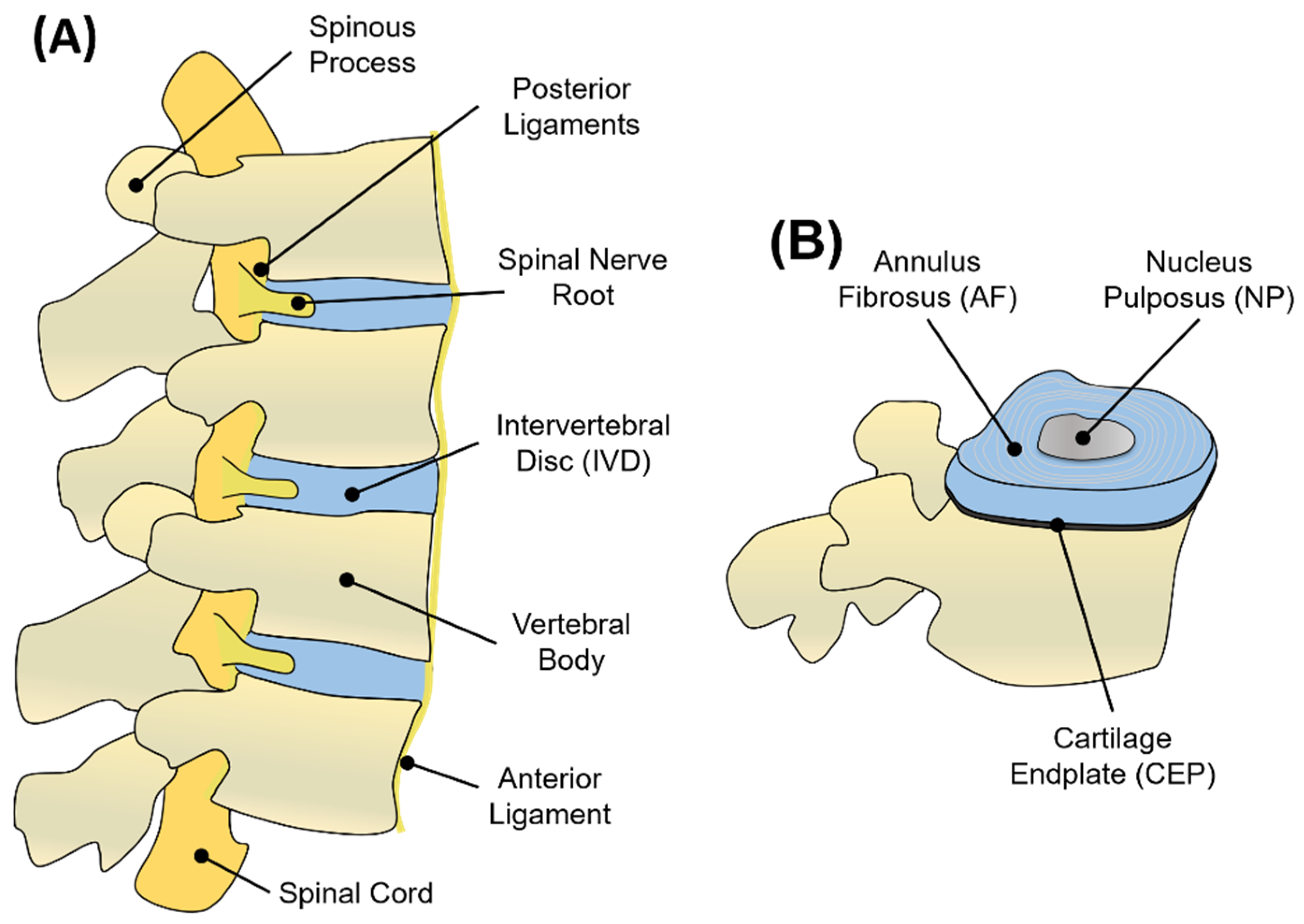
1.1. Structure and Function of the IVD
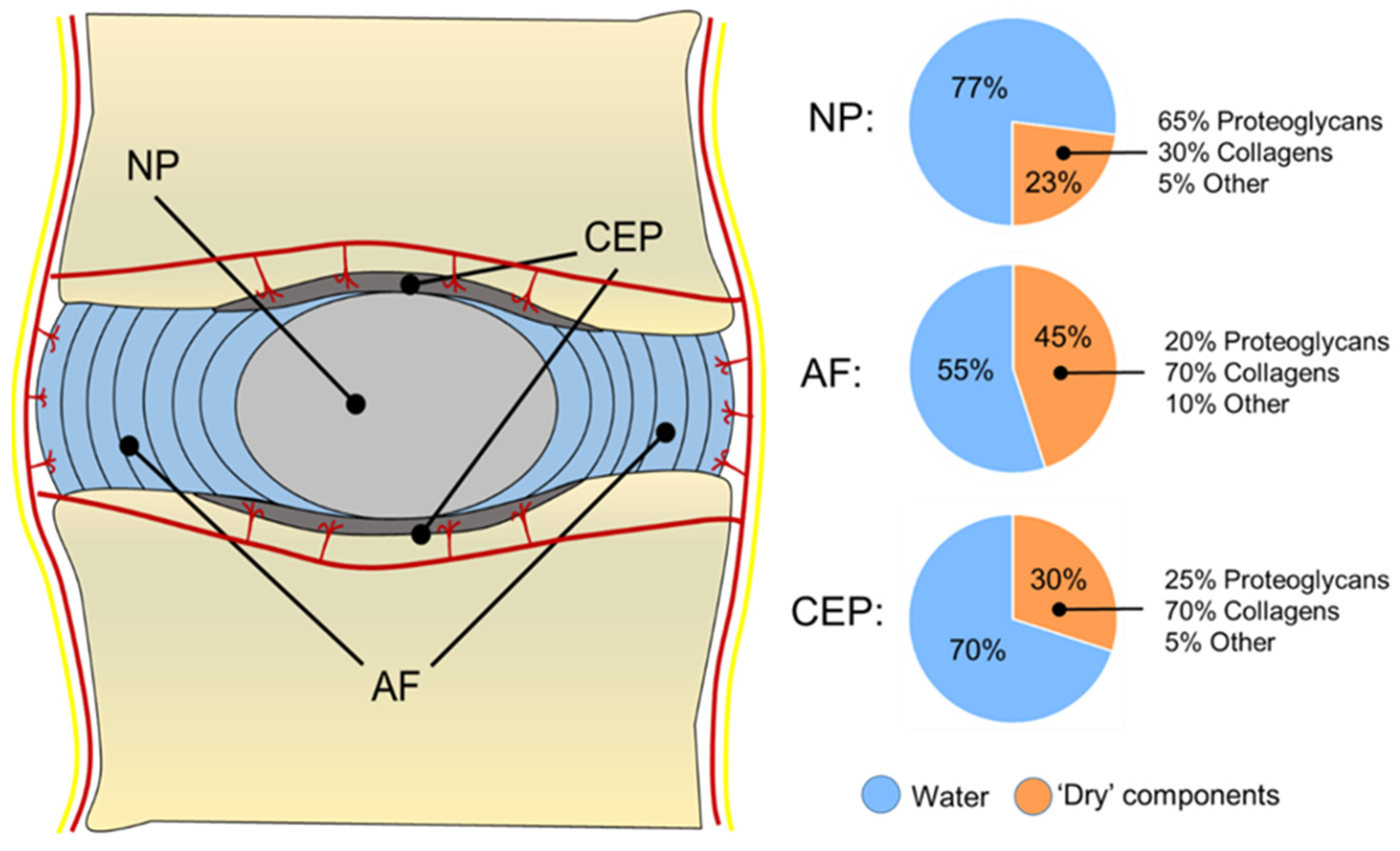
1.2. Nucleus Pulposus (NP)
1.3. Annulus Fibrosus (AF)
1.4. Cartilage Endplate (CEP)
1.5. Anabolic Growth Factors Present in the IVD
2. Pathophysiology of Intervertebral Disc Degeneration
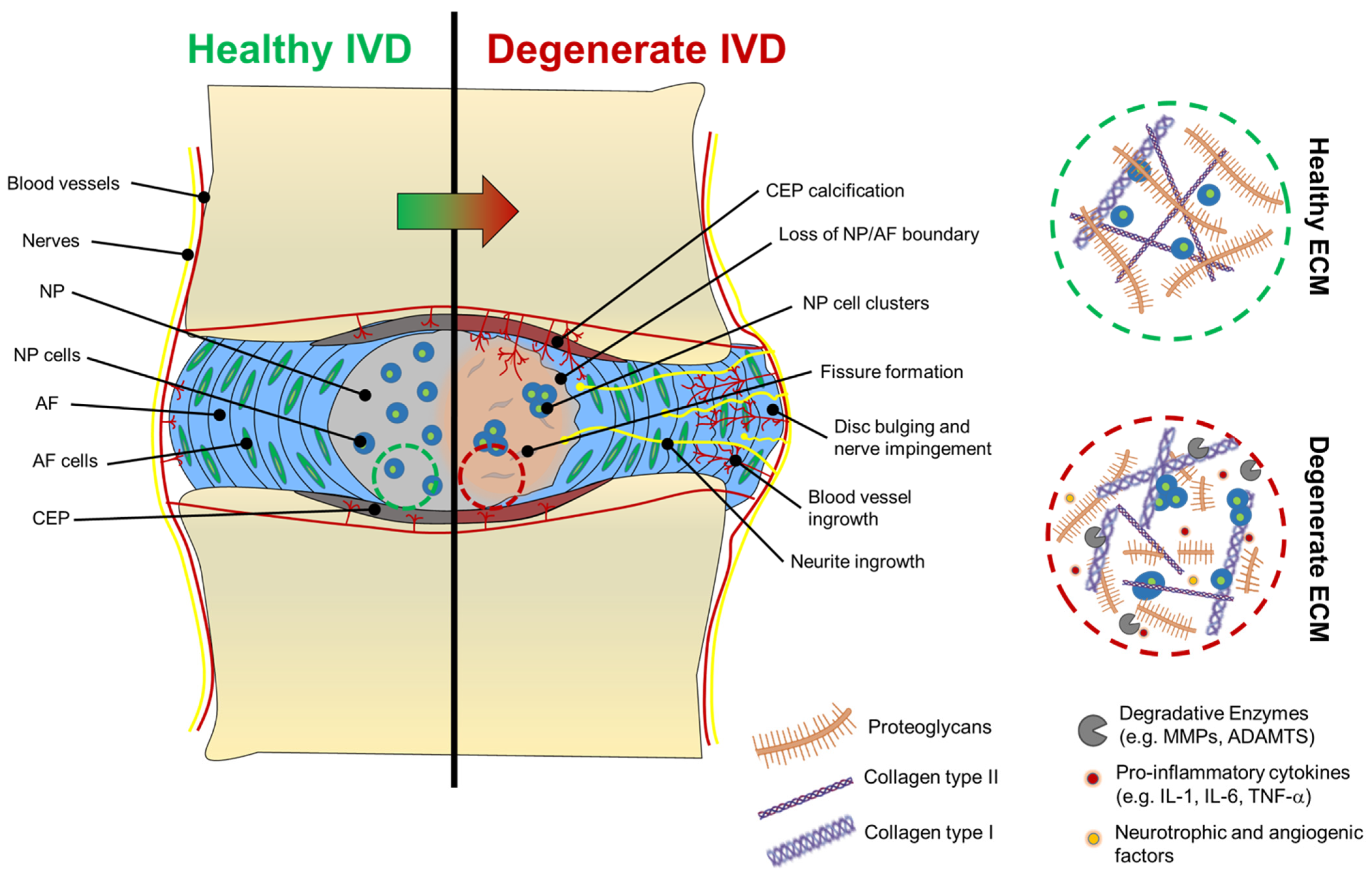
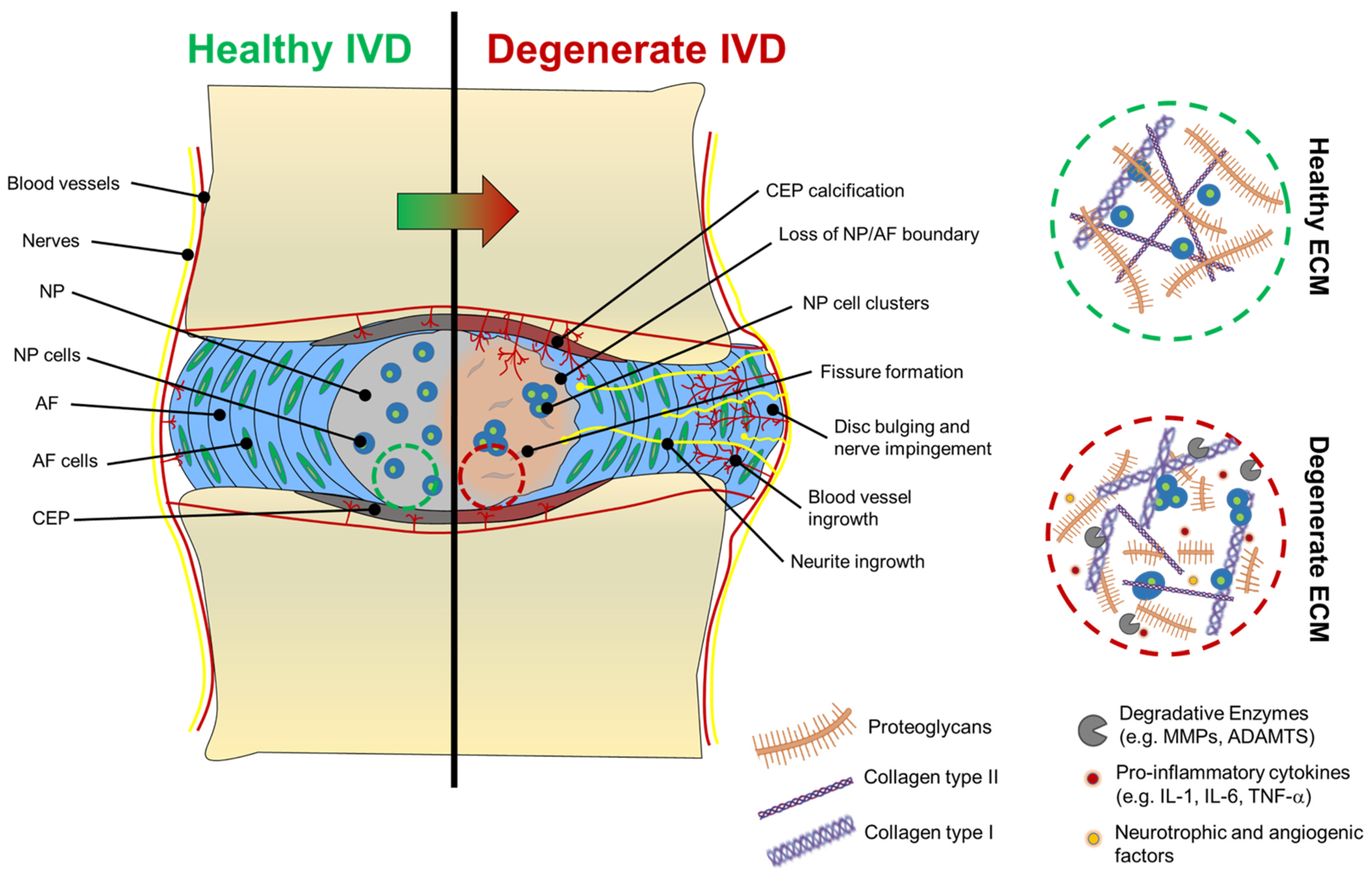
2.1. Alterations in the ECM of Degenerated Discs
2.2. ECM Degradation in Degenerated Discs
2.3. Nerve and Blood Vessel Infiltration in Degenerated Discs
2.4. The Effect of Ageing on Disc Degeneration
2.5. Mechanical Factors for Disc Degeneration
2.6. Genetic Influences on Disc Degeneration
3. Impact and Current Treatments of Disc Degeneration
3.1. Socio-Economic Burden
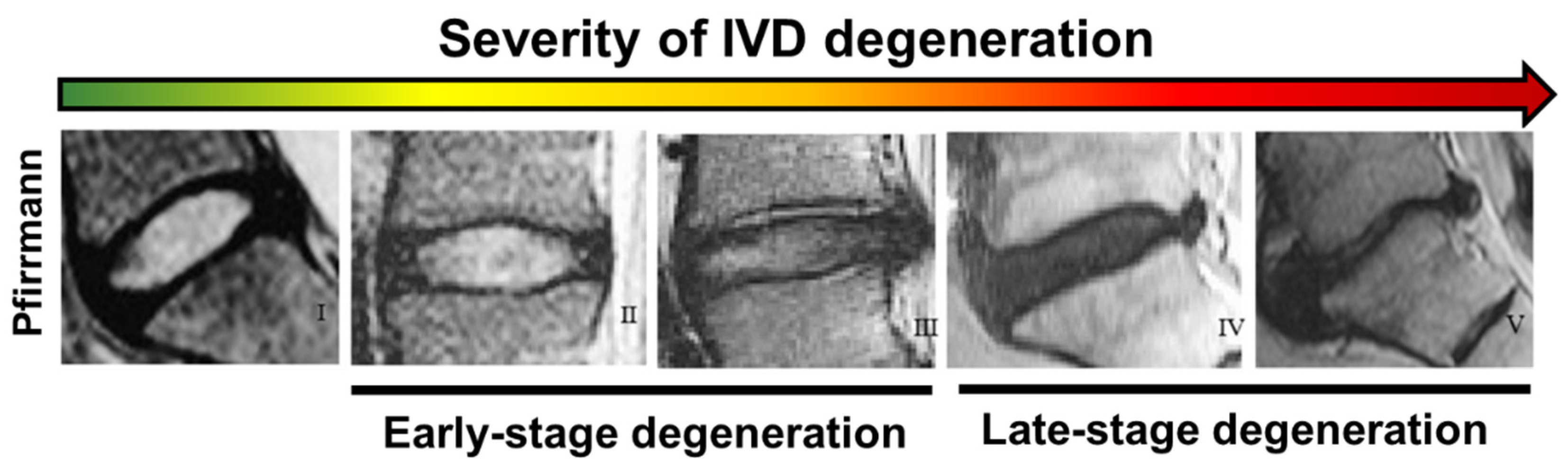
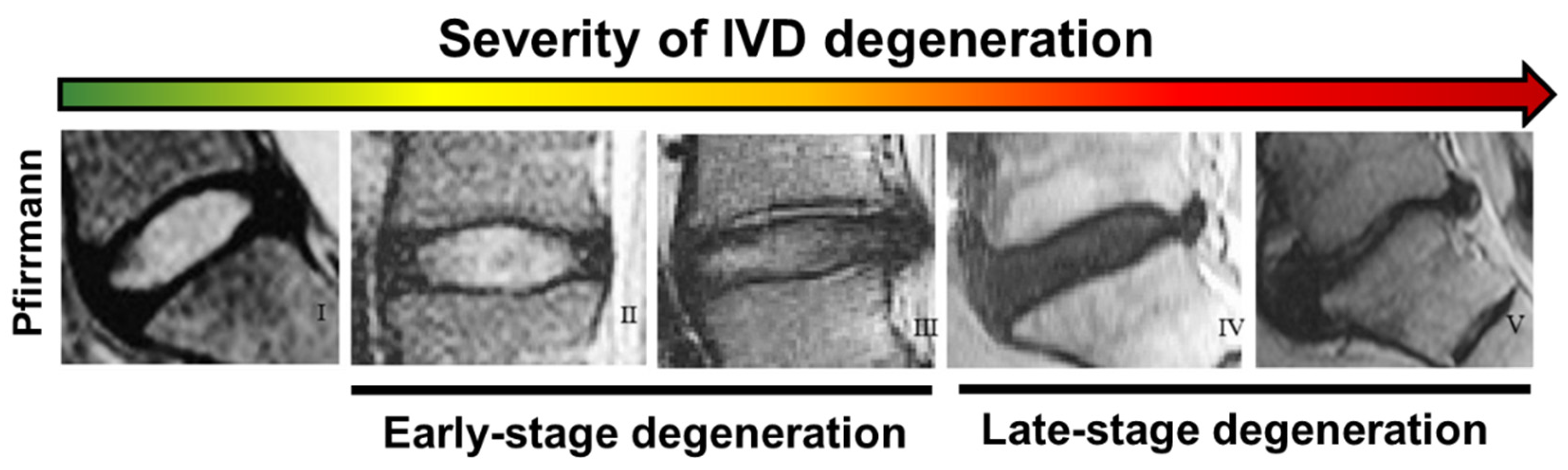
3.2. Grading Scales for the Assessment of Disc Degeneration
3.3. Conservative Therapies for Early Stage Disc Degeneration
3.4. Surgical Approaches for Late-Stage Disc Degeneration
3.5. Cell-Based Therapies for NP Regeneration: Cell Sources
3.6. Cell-Based Therapies for NP Regeneration: Biomaterial Requirements
4. Injectable Hydrogels for NP Regeneration
4.1. Advantages of Using Injectable Hydrogels for NP TE
4.2. Naturally Derived Injectable Hydrogels
4.3. Synthetic Injectable Hydrogels
5. Hydrogels Based on Self-Assembling Peptides
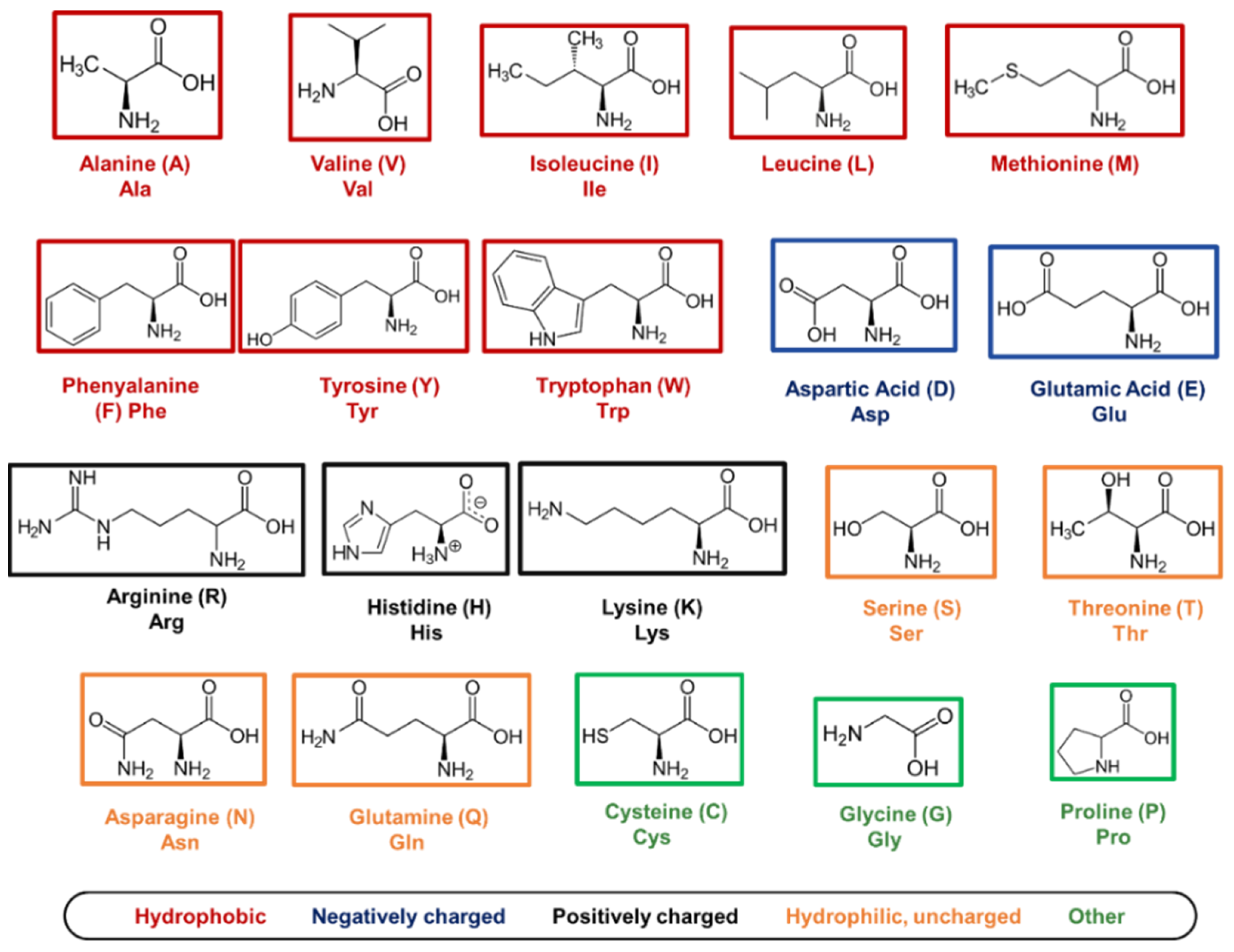
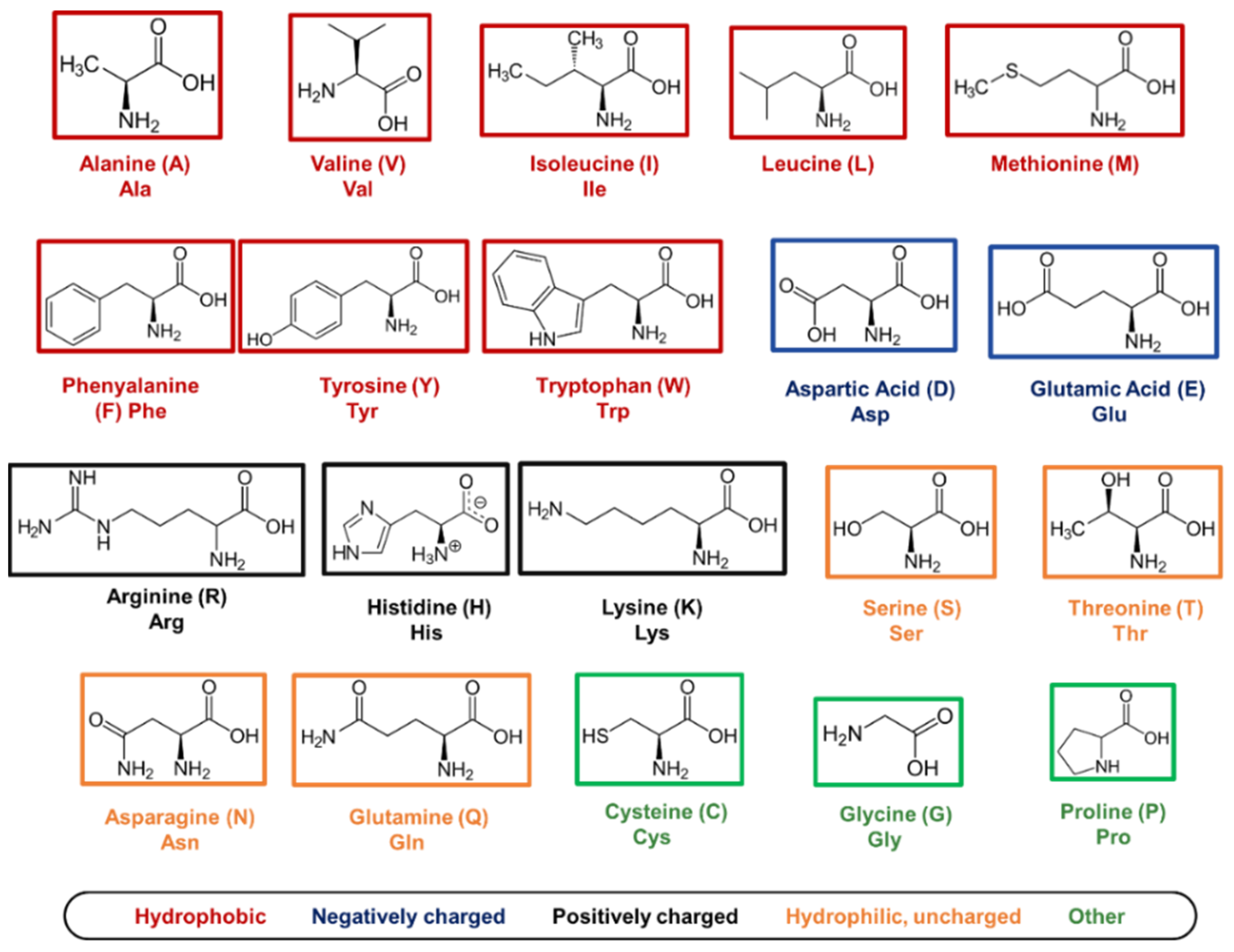
5.1. Amino Acids as Molecular Building Blocks
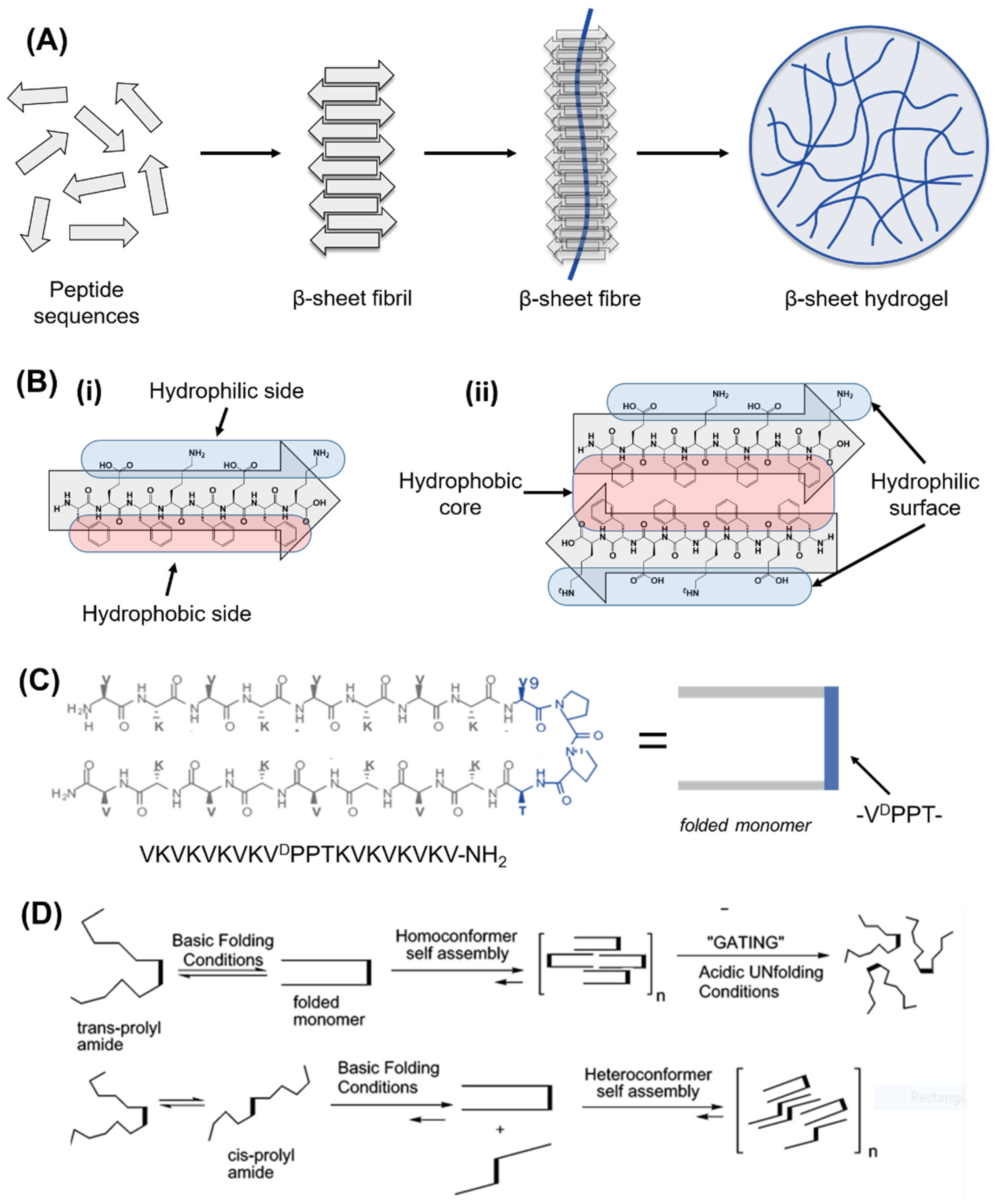
5.2. Self-Assembling Peptide Structures: β-Sheets and β-Hairpins
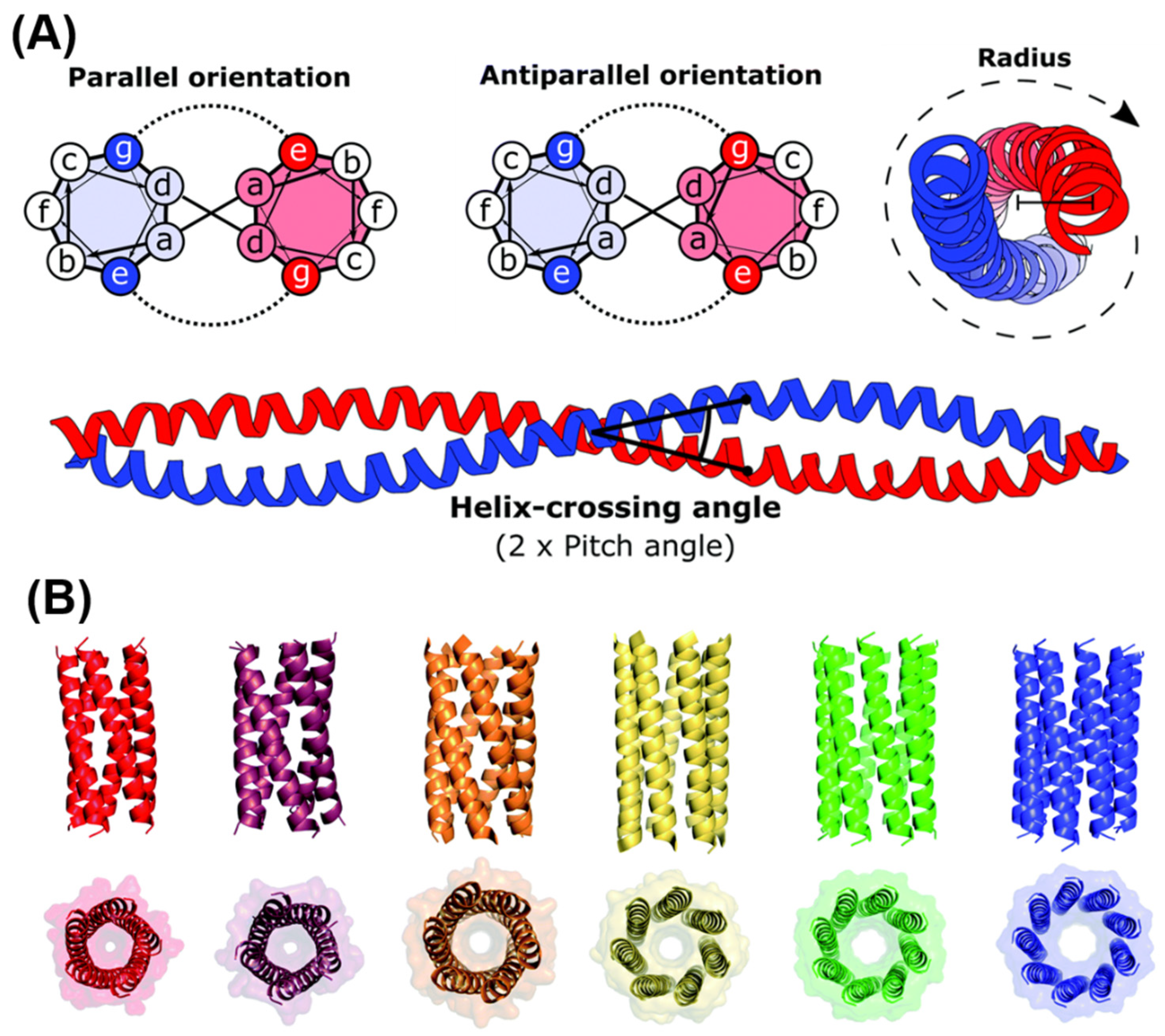
5.3. Self-Assembling Peptide Structures: α-Helices and Coiled Coils
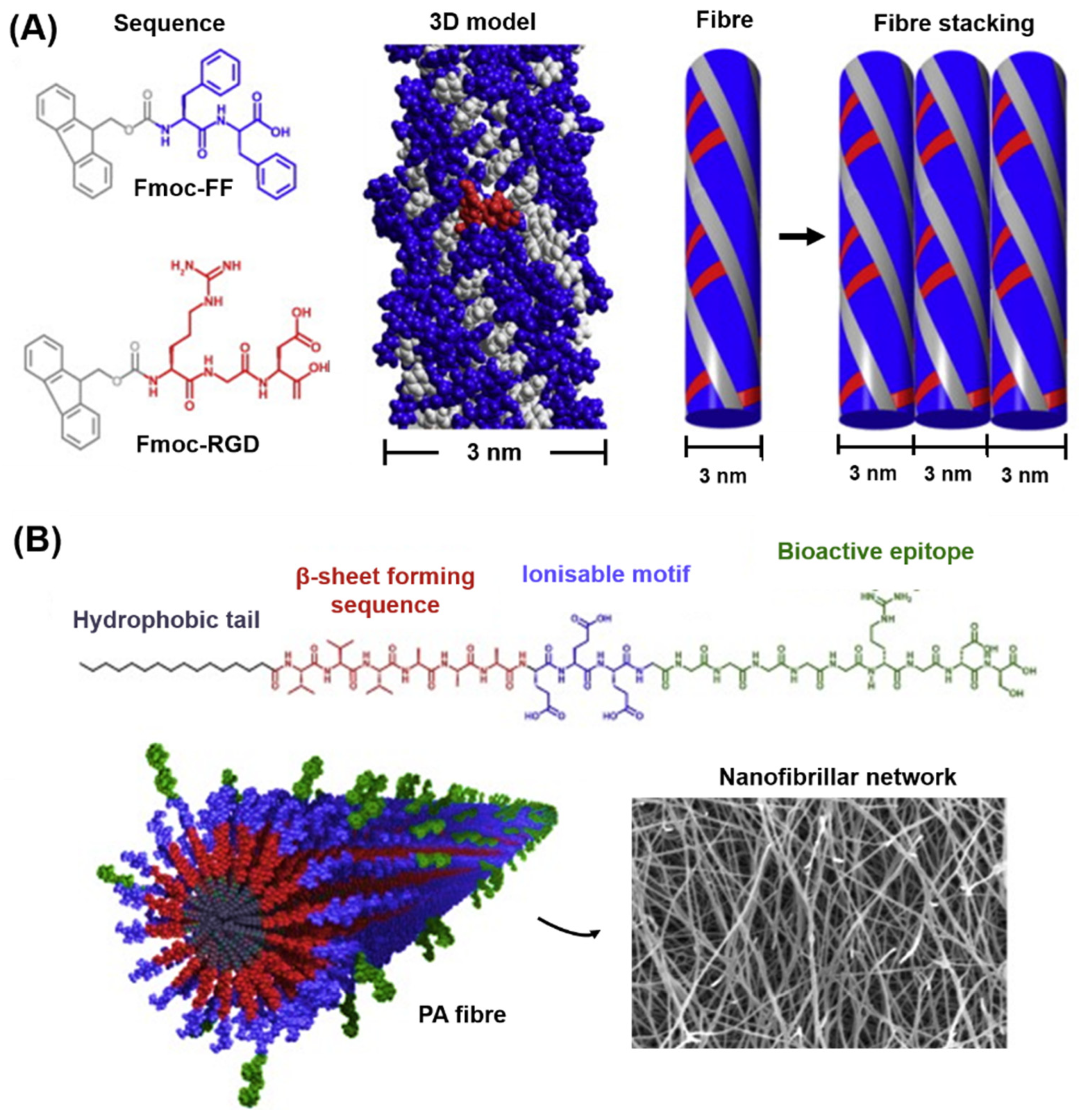
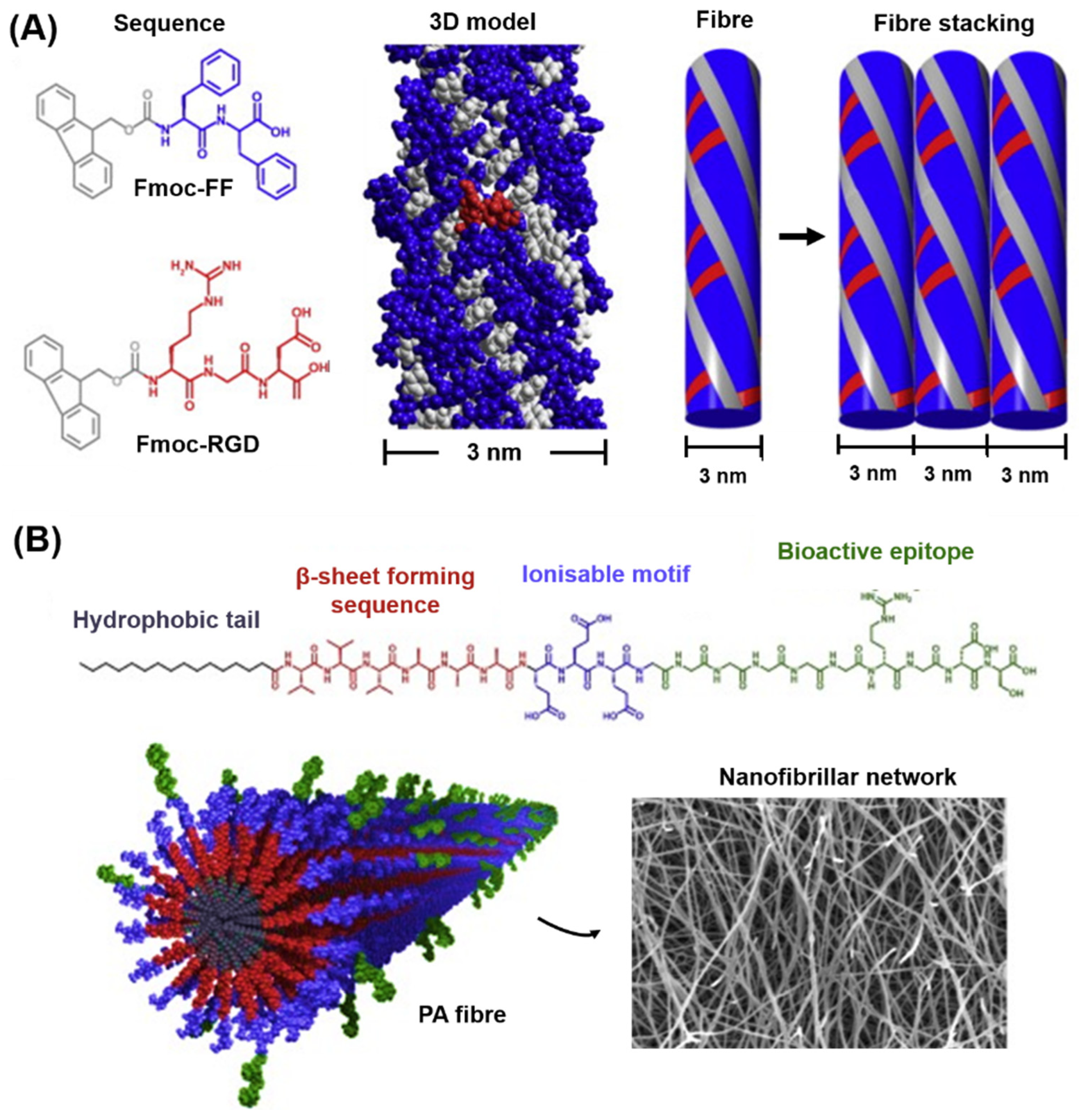
5.4. Short Aromatic Peptides and Peptide Amphiphiles
6. Peptide-Based Hydrogels for IVD Repair
7. Summary and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Robinson, J.S. Sciatica and the lumbar disk syndrome: A historic perspective. South. Med. J. 1983, 76, 232–238. [Google Scholar] [CrossRef] [PubMed]
- Humzah, M.D.; Soames, R.W. Human lntervertebral Disc: Structure and Function. Anat. Rec. 1988, 220, 337–356. [Google Scholar] [CrossRef] [PubMed]
- Raj, P.P. Intervertebral Disc: Pathophysiology-Treatment. Pain Pract. 2008, 8, 18–44. [Google Scholar] [CrossRef]
- Neidlinger-Wilke, C.; Galbusera, F.; Pratsinis, H.; Mavrogonatou, E.; Mietsch, A.; Kletsas, D.; Wilke, H.-J. Mechanical loading of the intervertebral disc: From the macroscopic to the cellular level. Eur. Spine J. Off. Publ. Eur. Spine Soc. Eur. Spinal Deform. Soc. Eur. Sect. Cerv. Spine Res. Soc. 2014, 23 (Suppl. S3), S333–S343. [Google Scholar] [CrossRef] [PubMed]
- Wilke, H.J.; Neef, P.; Caimi, M.; Hoogland, T.; Claes, L.E. New in vivo measurements of pressures in the intervertebral disc in daily life. Spine (Phila. Pa. 1976) 1999, 24, 755–762. [Google Scholar] [CrossRef] [PubMed]
- Newell, N.; Little, J.P.; Christou, A.; Adams, M.A.; Adam, C.J.; Masouros, S.D. Biomechanics of the human intervertebral disc: A review of testing techniques and results. J. Mech. Behav. Biomed. Mater. 2017, 69, 420–434. [Google Scholar] [CrossRef]
- Antoniou, J.; Steffen, T.; Nelson, F.; Winterbottom, N.; Hollander, A.P.; Poole, R.A.; Aebi, M.; Alini, M. The human lumbar intervertebral disc: Evidence for changes in the biosynthesis and denaturation of the extracellular matrix with growth, maturation, ageing, and degeneration. J. Clin. Investig. 1996, 98, 996–1003. [Google Scholar] [CrossRef]
- Iatridis, J.C.; Weidenbaum, M.; Setton, L.A.; Mow, V.C. Is the nucleus pulposus a solid or a fluid? Mechanical behaviors of the nucleus pulposus of the human intervertebral disc. Spine (Phila. Pa. 1976) 1996, 21, 1174–1184. [Google Scholar] [CrossRef]
- Mwale, F.; Roughley, P.; Antoniou, J. Distinction between the extracellular matrix of the nucleus pulposus and hyaline cartilage: A requisite for tissue engineering of intervertebral disc. Eur. Cell. Mater. 2004, 8, 54–58. [Google Scholar] [CrossRef]
- Sztrolovics, R.; Alini, M.; Mort, J.S.; Roughley, P.J. Age-related changes in fibromodulin and lumican in human intervertebral discs. Spine (Phila. Pa. 1976) 1999, 24, 1765–1771. [Google Scholar] [CrossRef]
- Melrose, J.; Ghosh, P.; Taylor, T.K. A comparative analysis of the differential spatial and temporal distributions of the large (aggrecan, versican) and small (decorin, biglycan, fibromodulin) proteoglycans of the intervertebral disc. J. Anat. 2001, 198, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Peter, C.; Roberts, S.; Urban, J.P.G. Elastic fibre organization in the intervertebral discs of the bovine tail. J. Anat. 2002, 201, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Eyre, D.R.; Muir, H. Types I and II collagens in intervertebral disc. Interchanging radial distributions in annulus fibrosus. Biochem. J. 1976, 157, 267–270. [Google Scholar] [CrossRef]
- Inoue, H.; Takeda, T. Three-dimensional observation of collagen framework of lumbar intervertebral discs. Acta Orthop. 1975, 46, 949–956. [Google Scholar] [CrossRef] [Green Version]
- Risbud, M.V.; Schoepflin, Z.R.; Mwale, F.; Kandel, R.A.; Grad, S.; Iatridis, J.C.; Sakai, D.; Hoyland, J.A. Defining the phenotype of young healthy nucleus pulposus cells: Recommendations of the Spine Research Interest Group at the 2014 annual ORS meeting. J. Orthop. Res. 2015, 33, 283–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stockwell, R.A. The interrelationship of cell density and cartilage thickness in mammalian articular cartilage. J. Anat. 1971, 109, 411–421. [Google Scholar]
- Maroudas, A.; Stockwell, R.A.; Nachemson, A.; Urban, J. Factors involved in the nutrition of the human lumbar intervertebral disc: Cellularity and diffusion of glucose in vitro. J. Anat. 1975, 120, 113–130. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1231728/ (accessed on 30 March 2022).
- Minogue, B.M.; Richardson, S.M.; Zeef, L.A.H.; Freemont, A.J.; Hoyland, J.A. Characterization of the human nucleus pulposus cell phenotype and evaluation of novel marker gene expression to define adult stem cell differentiation. Arthritis Rheum. 2010, 62, 3695–3705. [Google Scholar] [CrossRef]
- Minogue, B.M.; Richardson, S.M.; Zeef, L.A.H.; Freemont, A.J.; Hoyland, J.A. Transcriptional profiling of bovine intervertebral disc cells: Implications for identification of normal and degenerate human intervertebral disc cell phenotypes. Arthritis Res. Ther. 2010, 12, R22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nerlich, A.G.; Weiler, C.; Zipperer, J.; Narozny, M.; Boos, N. Immunolocalization of phagocytic cells in normal and degenerated intervertebral discs. Spine (Phila. Pa. 1976) 2002, 27, 2484–2490. [Google Scholar] [CrossRef]
- Shamji, M.F.; Setton, L.A.; Jarvis, W.; So, S.; Chen, J.; Jing, L.; Bullock, R.; Isaacs, R.E.; Brown, C.; Richardson, W.J. Proinflammatory cytokine expression profile in degenerated and herniated human intervertebral disc tissues. Arthritis Rheum. 2010, 62, 1974–1982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urban, J.P.G. The role of the physicochemical environment in determining disc cell behaviour. Biochem. Soc. Trans. 2002, 30, 858–864. [Google Scholar] [CrossRef] [PubMed]
- Holm, S.; Maroudas, A.; Urban, J.P.; Selstam, G.; Nachemson, A. Nutrition of the intervertebral disc: Solute transport and metabolism. Connect. Tissue Res. 1981, 8, 101–119. [Google Scholar] [CrossRef]
- Mwale, F.; Ciobanu, I.; Giannitsios, D.; Roughley, P.; Steffen, T.; Antoniou, J. Effect of oxygen levels on proteoglycan synthesis by intervertebral disc cells. Spine (Phila. Pa. 1976) 2011, 36, E131–E138. [Google Scholar] [CrossRef] [PubMed]
- Marchand, F.; Ahmed, A.M. Investigation of the laminate structure of lumbar disc anulus fibrosus. Spine (Phila. Pa. 1976) 1990, 15, 402–410. [Google Scholar] [CrossRef]
- Taylor, T.K.; Ghosh, P.; Bushell, G.R. The contribution of the intervertebral disk to the scoliotic deformity. Clin. Orthop. Relat. Res. 1981, 156, 79–90. [Google Scholar] [CrossRef]
- Schollmeier, G.; Lahr-Eigen, R.; Lewandrowski, K.U. Observations on fiber-forming collagens in the anulus fibrosus. Spine (Phila. Pa. 1976) 2000, 25, 2736–2741. [Google Scholar] [CrossRef]
- Errington, R.J.; Puustjarvi, K.; White, I.R.; Roberts, S.; Urban, J.P. Characterisation of cytoplasm-filled processes in cells of the intervertebral disc. J. Anat. 1998, 192 Pt 3, 369–378. [Google Scholar] [CrossRef]
- Freemont, A.J.; Peacock, T.E.; Goupille, P.; Hoyland, J.A.; O’Brien, J.; Jayson, M.I. Nerve ingrowth into diseased intervertebral disc in chronic back pain. Lancet 1997, 350, 178–181. [Google Scholar] [CrossRef]
- Purmessur, D.; Freemont, A.J.; Hoyland, J.A. Expression and regulation of neurotrophins in the nondegenerate and degenerate human intervertebral disc. Arthritis Res. Ther. 2008, 10, R99. [Google Scholar] [CrossRef] [Green Version]
- Roberts, S.; Menage, J.; Urban, J.P. Biochemical and structural properties of the cartilage end-plate and its relation to the intervertebral disc. Spine (Phila. Pa. 1976) 1989, 14, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Roberts, S.; McCall, I.W.; Menage, J.; Haddaway, M.J.; Eisenstein, S.M. Does the thickness of the vertebral subchondral bone reflect the composition of the intervertebral disc? Eur. Spine J. 1997, 6, 385–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nachemson, A.; Lewin, T.; Maroudas, A.; Freeman, M.A. In vitro diffusion of dye through the end-plates and the annulus fibrosus of human lumbar inter-vertebral discs. Acta Orthop. Scand. 1970, 41, 589–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, S.; Menage, J.; Duance, V.; Wotton, S.F. Type III collagen in the intervertebral disc. Histochem. J. 1991, 23, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Roberts, S.; Menage, J.; Duance, V.; Wotton, S.; Ayad, S. 1991 Volvo Award in basic sciences. Collagen types around the cells of the intervertebral disc and cartilage end plate: An immunolocalization study. Spine (Phila. Pa. 1976) 1991, 16, 1030–1038. [Google Scholar] [CrossRef]
- Moore, R.J. The vertebral endplate: Disc degeneration, disc regeneration. Eur. Spine J. 2006, 15, 333–337. [Google Scholar] [CrossRef] [Green Version]
- Hynes, R.O. Extracellular matrix: Not just pretty fibrils. Science 2009, 326, 1216–1219. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Liu, S.; Ma, K.; Zhao, L.; Lin, H.; Shao, Z. TGF-β signaling in intervertebral disc health and disease. Osteoarthr. Cartil. 2019, 27, 1109–1117. [Google Scholar] [CrossRef]
- Gruber, H.E.; Fisher, E.C.; Desai, B.; Stasky, A.A.; Hoelscher, G.; Hanley, E.N. Human intervertebral disc cells from the annulus: Three-dimensional culture in agarose or alginate and responsiveness to TGF-beta1. Exp Cell Res. 1997, 235, 13–21. [Google Scholar] [CrossRef]
- Jin, H.; Shen, J.; Wang, B.; Wang, M.; Shu, B.; Chen, D. TGF-β signaling plays an essential role in the growth and maintenance of intervertebral disc tissue. FEBS Lett. 2011, 585, 1209–1215. [Google Scholar] [CrossRef] [Green Version]
- Hodgkinson, T.; Shen, B.; Diwan, A.; Hoyland, J.A.; Richardson, S.M. Therapeutic potential of growth differentiation factors in the treatment of degenerative disc diseases. JOR Spine 2019, 2, e1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richardson, S.M.; Freemont, A.J.; Hoyland, J.A. Pathogenesis of Intervertebral Disc Degeneration. In The Intervertebral Disc; Shapiro, I.M., Risbud, M.V., Eds.; Springer: Vienna, Vienna, 2014; pp. 177–200. [Google Scholar] [CrossRef]
- Duance, V.C.; Crean, J.K.; Sims, T.J.; Avery, N.; Smith, S.; Menage, J.; Eisenstein, S.M.; Roberts, S. Changes in collagen cross-linking in degenerative disc disease and scoliosis. Spine (Phila. Pa. 1976) 1998, 23, 2545–2551. [Google Scholar] [CrossRef] [PubMed]
- le Maitre, C.L.; Pockert, A.; Buttle, D.J.; Freemont, A.J.; Hoyland, J.A. Matrix synthesis and degradation in human intervertebral disc degeneration. Biochem. Soc. Trans. 2007, 35, 652–655. [Google Scholar] [CrossRef] [Green Version]
- Boos, N.; Nerlich, A.G.; Wiest, I.; von der Mark, K.; Aebi, M. Immunolocalization of type X collagen in human lumbar intervertebral discs during ageing and degeneration. Histochem Cell Biol. 1997, 108, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Nachemson, A. Intradiscal Measurements of pH in Patients with Lumbar Rhizopathies. Acta Orthop. Scand. 1969, 40, 23–42. [Google Scholar] [CrossRef] [Green Version]
- Ohshima, H.; Urban, J.P. The effect of lactate and pH on proteoglycan and protein synthesis rates in the intervertebral disc. Spine (Phila. Pa. 1976) 1992, 17, 1079–1082. [Google Scholar] [CrossRef]
- Gilbert, H.T.J.; Hodson, N.; Baird, P.; Richardson, S.M.; Hoyland, J.A. Acidic pH promotes intervertebral disc degeneration: Acid-sensing ion channel -3 as a potential therapeutic target. Sci. Rep. 2016, 6, 37360. [Google Scholar] [CrossRef] [Green Version]
- Cs-Szabo, G.; Juan, D.R.; Turumella, V.; Masuda, K.; Thonar, E.J.; An, H.S. Changes in mRNA and protein levels of proteoglycans of the anulus fibrosus and nucleus pulposus during intervertebral disc degeneration. Spine (Phila. Pa. 1976) 2002, 27, 2212–2219. [Google Scholar] [CrossRef]
- Sztrolovics, R.; Grover, J.; Cs-Szabo, G.; Shi, S.-L.; Zhang, Y.; Mort, J.S.; Roughley, P.J. The characterization of versican and its message in human articular cartilage and intervertebral disc. J. Orthop. Res. 2002, 20, 257–266. [Google Scholar] [CrossRef]
- Oegema, T.R.J.; Johnson, S.L.; Aguiar, D.J.; Ogilvie, J.W. Fibronectin and its fragments increase with degeneration in the human intervertebral disc. Spine (Phila. Pa. 1976) 2000, 25, 2742–2747. [Google Scholar] [CrossRef]
- Aota, Y.; An, H.S.; Homandberg, G.; Thonar, E.J.M.-A.; Andersson, G.B.J.; Pichika, R.; Masuda, K. Differential effects of fibronectin fragment on proteoglycan metabolism by intervertebral disc cells: A comparison with articular chondrocytes. Spine (Phila. Pa. 1976) 2005, 30, 722–728. [Google Scholar] [CrossRef] [PubMed]
- Anderson, D.G.; Li, X.; Tannoury, T.; Beck, G.; Balian, G. A fibronectin fragment stimulates intervertebral disc degeneration in vivo. Spine (Phila. Pa. 1976) 2003, 28, 2338–2345. [Google Scholar] [CrossRef] [PubMed]
- Nagase, H.; Kashiwagi, M. Aggrecanases and cartilage matrix degradation. Arthritis Res. Ther. 2003, 5, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Nagase, H.; Woessner, J.F.J. Matrix metalloproteinases. J. Biol. Chem. 1999, 274, 21491–21494. [Google Scholar] [CrossRef] [Green Version]
- Risbud, M.V. The Intervertebral Disc. Lancet 1951, 257, 275–276. [Google Scholar] [CrossRef]
- Pockert, A.J.; Richardson, S.M.; le Maitre, C.L.; Lyon, M.; Deakin, J.A.; Buttle, D.J.; Freemont, A.J.; Hoyland, J.A. Modified expression of the ADAMTS enzymes and tissue inhibitor of metalloproteinases 3 during human intervertebral disc degeneration. Arthritis Rheum. 2009, 60, 482–491. [Google Scholar] [CrossRef]
- Studer, R.K.; Vo, N.; Sowa, G.; Ondeck, C.; Kang, J. Human nucleus pulposus cells react to IL-6: Independent actions and amplification of response to IL-1 and TNF-alpha. Spine (Phila. Pa. 1976) 2011, 36, 593–599. [Google Scholar] [CrossRef]
- le Maitre, C.L.; Freemont, A.J.; Hoyland, J.A. Localization of degradative enzymes and their inhibitors in the degenerate human intervertebral disc. J Pathol. 2004, 204, 47–54. [Google Scholar] [CrossRef]
- le Maitre, C.L.; Freemont, A.J.; Hoyland, J.A. The role of interleukin-1 in the pathogenesis of human Intervertebral disc degeneration. Arthritis Res. Ther. 2005, 7, R732. [Google Scholar] [CrossRef] [Green Version]
- le Maitre, C.L.; Freemont, A.J.; Hoyland, J.A. A preliminary in vitro study into the use of IL-1Ra gene therapy for the inhibition of intervertebral disc degeneration. Int. J. Exp. Pathol. 2006, 87, 17–28. [Google Scholar] [CrossRef]
- le Maitre, C.L.; Hoyland, J.A.; Freemont, A.J. Interleukin-1 receptor antagonist delivered directly and by gene therapy inhibits matrix degradation in the intact degenerate human intervertebral disc: An in situ zymographic and gene therapy study. Arthritis Res. Ther. 2007, 9, R83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, W.E.B.; Caterson, B.; Eisenstein, S.M.; Hynds, D.L.; Snow, D.M.; Roberts, S. Human intervertebral disc aggrecan inhibits nerve growth in vitro. Arthritis Rheum. 2002, 46, 2658–2664. [Google Scholar] [CrossRef]
- Richardson, S.M.; Doyle, P.; Minogue, B.M.; Gnanalingham, K.; Hoyland, J.A. Increased expression of matrix metalloproteinase-10, nerve growth factor and substance P in the painful degenerate intervertebral disc. Arthritis Res. Ther. 2009, 11, R126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallach, D.; Arumugam, T.U.; Boldin, M.P.; Cantarella, G.; Ganesh, K.A.; Goltsev, Y.; Goncharov, T.M.; Kovalenko, A.V.; Rajput, A.; Varfolomeev, E.E.; et al. How are the regulators regulated? The search for mechanisms that impose specificity on induction of cell death and NF-kappaB activation by members of the TNF/NGF receptor family. Arthritis Res. 2002, 4 (Suppl. S3), S189–S196. [Google Scholar] [CrossRef] [PubMed]
- Cheung, K.M.C.; Karppinen, J.; Chan, D.; Ho, D.W.H.; Song, Y.-Q.; Sham, P.; Cheah, K.S.E.; Leong, J.C.Y.; Luk, K.D.K. Prevalence and pattern of lumbar magnetic resonance imaging changes in a population study of one thousand forty-three individuals. Spine (Phila. Pa. 1976) 2009, 34, 934–940. [Google Scholar] [CrossRef] [PubMed]
- Boos, N.; Weissbach, S.; Rohrbach, H.; Weiler, C.; Spratt, K.F.; Nerlich, A.G. Classification of age-related changes in lumbar intervertebral discs: 2002 Volvo Award in basic science. Spine (Phila. Pa. 1976) 2002, 27, 2631–2644. [Google Scholar] [CrossRef] [PubMed]
- Risbud, M.V.; Fertala, J.; Vresilovic, E.J.; Albert, T.J.; Shapiro, I.M. Nucleus pulposus cells upregulate PI3K/Akt and MEK/ERK signaling pathways under hypoxic conditions and resist apoptosis induced by serum withdrawal. Spine (Phila. Pa. 1976) 2005, 30, 882–889. [Google Scholar] [CrossRef]
- Shen, C.; Yan, J.; Jiang, L.-S.; Dai, L.-Y. Autophagy in rat annulus fibrosus cells: Evidence and possible implications. Arthritis Res. Ther. 2011, 13, R132. [Google Scholar] [CrossRef] [Green Version]
- Ye, W.; Xu, K.; Huang, D.; Liang, A.; Peng, Y.; Zhu, W.; Li, C. Age-related increases of macroautophagy and chaperone-mediated autophagy in rat nucleus pulposus. Connect. Tissue Res. 2011, 52, 472–478. [Google Scholar] [CrossRef] [PubMed]
- Roberts, S.; Evans, E.H.; Kletsas, D.; Jaffray, D.C.; Eisenstein, S.M. Senescence in human intervertebral discs. Eur. Spine J. Off. Publ. Eur. Spine Soc. Eur. Spinal Deform. Soc. Eur. Sect. Cerv. Spine Res. Soc. 2006, 15 (Suppl. S3), S312–S316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- le Maitre, C.L.; Freemont, A.J.; Hoyland, J.A. Accelerated cellular senescence in degenerate intervertebral discs: A possible role in the pathogenesis of intervertebral disc degeneration. Arthritis Res. Ther. 2007, 9, R45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.-W.; Chung, H.-N.; Ha, K.-Y.; Lee, J.-S.; Kim, Y.-Y. Senescence mechanisms of nucleus pulposus chondrocytes in human intervertebral discs. Spine J. 2009, 9, 658–666. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, B.; Zhao, X.; Li, X.; Lou, Z.; Chen, X.; Zhang, F. Iron deficiency accelerates intervertebral disc degeneration through affecting the stability of DNA polymerase epsilon complex. Am. J. Transl. Res. 2018, 10, 3430–3442. [Google Scholar]
- Barbosa, M.C.; Grosso, R.A.; Fader, C.M. Hallmarks of Aging: An Autophagic Perspective. Front. Endocrinol. 2019, 9, 790. [Google Scholar] [CrossRef]
- Puustjarvi, K.; Lammi, M.; Helminen, H.; Inkinen, R.; Tammi, M. Proteoglycans in the intervertebral disc of young dogs following strenuous running exercise. Connect. Tissue Res. 1994, 30, 225–240. [Google Scholar] [CrossRef]
- Iatridis, J.C.; Mente, P.L.; Stokes, I.A.F.; Aronsson, D.D.; Alini, M. Compression-Induced Changes in Intervertebral Disc Properties in a Rat Tail Model. Spine (Phila. Pa. 1976) 1999, 24. [Google Scholar] [CrossRef]
- MacLean, J.J.; Lee, C.R.; Alini, M.; Iatridis, J.C. The effects of short-term load duration on anabolic and catabolic gene expression in the rat tail intervertebral disc. J. Orthop. Res. 2005, 23, 1120–1127. [Google Scholar] [CrossRef] [PubMed]
- Wuertz, K.; Godburn, K.; MacLean, J.J.; Barbir, A.; Donnelly, J.S.; Roughley, P.J.; Alini, M.; Iatridis, J.C. In vivo remodeling of intervertebral discs in response to short- and long-term dynamic compression. J. Orthop. Res. 2009, 27, 1235–1242. [Google Scholar] [CrossRef] [PubMed]
- Maclean, J.J.; Lee, C.R.; Alini, M.; Iatridis, J.C. Anabolic and catabolic mRNA levels of the intervertebral disc vary with the magnitude and frequency of in vivo dynamic compression. J. Orthop. Res. 2004, 22, 1193–1200. [Google Scholar] [CrossRef] [PubMed]
- Handa, T.; Ishihara, H.; Ohshima, H.; Osada, R.; Tsuji, H.; Obata, K. Effects of hydrostatic pressure on matrix synthesis and matrix metalloproteinase production in the human lumbar intervertebral disc. Spine (Phila. Pa. 1976) 1997, 22, 1085–1091. [Google Scholar] [CrossRef]
- Neidlinger-Wilke, C.; Wurtz, K.; Urban, J.P.G.; Borm, W.; Arand, M.; Ignatius, A.; Wilke, H.-J.; Claes, L.E. Regulation of gene expression in intervertebral disc cells by low and high hydrostatic pressure. Eur. Spine J. Off. Publ. Eur. Spine Soc. Eur. Spinal Deform. Soc. Eur. Sect. Cerv. Spine Res. Soc. 2006, 15 (Suppl. S3), S372–S378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- le Maitre, C.L.; Frain, J.; Fotheringham, A.P.; Freemont, A.J.; Hoyland, J.A. Human cells derived from degenerate intervertebral discs respond differently to those derived from non-degenerate intervertebral discs following application of dynamic hydrostatic pressure. Biorheology 2008, 45, 563–575. [Google Scholar] [CrossRef]
- Gilbert, H.T.J.; Hoyland, J.A.; Millward-Sadler, S.J. The response of human anulus fibrosus cells to cyclic tensile strain is frequency-dependent and altered with disc degeneration. Arthritis Rheum. 2010, 62, 3385–3394. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, H.T.J.; Hoyland, J.A.; Freemont, A.J.; Millward-Sadler, S.J. The involvement of interleukin-1 and interleukin-4 in the response of human annulus fibrosus cells to cyclic tensile strain: An altered mechanotransduction pathway with degeneration. Arthritis Res. Ther. 2011, 13, R8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varlotta, G.P.; Brown, M.D.; Kelsey, J.L.; Golden, A.L. Familial predisposition for herniation of a lumbar disc in patients who are less than twenty-one years old. J. Bone Jt. Surg. Am. 1991, 73, 124–128. Available online: https://journals.lww.com/jbjsjournal/Abstract/1991/73010/Familial_predisposition_for_herniation_of_a_lumbar.16.aspx (accessed on 4 March 2022). [CrossRef]
- MacGregor, A.J.; Andrew, T.; Sambrook, P.N.; Spector, T.D. Structural, psychological, and genetic influences on low back and neck pain: A study of adult female twins. Arthritis Rheum. 2004, 51, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Li, S.W.; Prockop, D.J.; Helminen, H.; Fassler, R.; Lapvetelainen, T.; Kiraly, K.; Peltarri, A.; Arokoski, J.; Lui, H.; Arita, M. Transgenic mice with targeted inactivation of the Col2 alpha 1 gene for collagen II develop a skeleton with membranous and periosteal bone but no endochondral bone. Genes Dev. 1995, 9, 2821–2830. [Google Scholar] [CrossRef] [Green Version]
- Kimura, T.; Nakata, K.; Tsumaki, N.; Miyamoto, S.; Matsui, Y.; Ebara, S.; Ochi, T. Progressive degeneration of articular cartilage and intervertebral discs. An experimental study in transgenic mice bearing a type IX collagen mutation. Int. Orthop. 1996, 20, 177–181. [Google Scholar] [CrossRef]
- Watanabe, H.; Nakata, K.; Kimata, K.; Nakanishi, I.; Yamada, Y. Dwarfism and age-associated spinal degeneration of heterozygote cmd mice defective in aggrecan. Proc. Natl. Acad. Sci. USA 1997, 94, 6943–6947. [Google Scholar] [CrossRef] [Green Version]
- Rajasekaran, S.; Kanna, R.M.; Senthil, N.; Raveendran, M.; Cheung, K.M.C.; Chan, D.; Subramaniam, S.; Shetty, A.P. Phenotype variations affect genetic association studies of degenerative disc disease: Conclusions of analysis of genetic association of 58 single nucleotide polymorphisms with highly specific phenotypes for disc degeneration in 332 subjects. Spine J. 2013, 13, 1309–1320. [Google Scholar] [CrossRef]
- Stankiewicz, P.; Sen, P.; Bhatt, S.S.; Storer, M.; Xia, Z.; Bejjani, B.A.; Ou, Z.; Wiszniewska, J.; Driscoll, D.J.; Maisenbacher, M.K.; et al. Genomic and genic deletions of the FOX gene cluster on 16q24.1 and inactivating mutations of FOXF1 cause alveolar capillary dysplasia and other malformations. Am. J. Hum. Genet. 2009, 84, 780–791. [Google Scholar] [CrossRef] [Green Version]
- Smith, C.A.; Tuan, R.S. Functional involvement of Pax-1 in somite development: Somite dysmorphogenesis in chick embryos treated with Pax-1 paired-box antisense oligodeoxynucleotide. Teratology 1995, 52, 333–345. [Google Scholar] [CrossRef]
- Wallin, J.; Wilting, J.; Koseki, H.; Fritsch, R.; Christ, B.; Balling, R. The role of Pax-1 in axial skeleton development. Development 1994, 120, 1109–1121. [Google Scholar] [CrossRef] [PubMed]
- Stewart, W.F.; Ricci, J.A.; Chee, E.; Morganstein, D.; Lipton, R. Lost productive time and cost due to common pain conditions in the US workforce. JAMA 2003, 290, 2443–2454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vlaeyen, J.W.S.; Maher, C.G.; Wiech, K.; van Zundert, J.; Meloto, C.B.; Diatchenko, L.; Battié, M.C.; Goossens, M.; Koes, B.; Linton, S.J. Low back pain. Nat. Rev. Dis. Prim. 2018, 4, 52. [Google Scholar] [CrossRef]
- de Schepper, E.I.T.; Damen, J.; van Meurs, J.B.J.; Ginai, A.Z.; Popham, M.; Hofman, A.; Koes, B.W.; Bierma-Zeinstra, S.M. The association between lumbar disc degeneration and low back pain: The influence of age, gender, and individual radiographic features. Spine (Phila. Pa. 1976) 2010, 35, 531–536. [Google Scholar] [CrossRef] [PubMed]
- Ravindra, V.M.; Senglaub, S.S.; Rattani, A.; Dewan, M.C.; Härtl, R.; Bisson, E.; Park, K.B.; Shrime, M.G. Degenerative Lumbar Spine Disease: Estimating Global Incidence and Worldwide Volume. Glob. Spine J. 2018, 8, 784–794. [Google Scholar] [CrossRef] [Green Version]
- Hoy, D.; Brooks, P.; Blyth, F.; Buchbinder, R. The Epidemiology of low back pain. Best Pract. Res. Clin. Rheumatol. 2010, 24, 769–781. [Google Scholar] [CrossRef] [PubMed]
- Maniadakis, N.; Gray, A. The economic burden of back pain in the UK. Pain 2000, 84, 95–103. [Google Scholar] [CrossRef]
- Hoy, D.; March, L.; Brooks, P.; Blyth, F.; Woolf, A.; Bain, C.; Williams, G.; Smith, E.; Vos, T.; Barendregt, J.; et al. The global burden of low back pain: Estimates from the Global Burden of Disease 2010 study. Ann. Rheum. Dis. 2014, 73, 968–974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfirrmann, C.W.A.; Metzdorf, A.; Zanetti, M.; Hodler, J.; Boos, N. Magnetic Resonance Classification of Lumbar Intervertebral Disc Degeneration. Spine (Phila. Pa. 1976) 2001, 26, 1873–1878. [Google Scholar] [CrossRef]
- Radek, M.; Pacholczyk-Sienicka, B.; Jankowski, S.; Albrecht, Ł.; Grodzka, M.; Depta, A.; Radek, A. Assessing the correlation between the degree of disc degeneration on the Pfirrmann scale and the metabolites identified in HR-MAS NMR spectroscopy. Magn. Reson. Imaging 2016, 34, 376–380. [Google Scholar] [CrossRef] [PubMed]
- Griffith, J.F.; Wang, Y.-X.J.; Antonio, G.E.; Choi, K.C.; Yu, A.; Ahuja, A.T.; Leung, P.C. Modified Pfirrmann Grading System for Lumbar Intervertebral Disc Degeneration. Spine (Phila. Pa. 1976) 2007, 32, E708–E712. [Google Scholar] [CrossRef] [PubMed]
- Sobajima, S.; Kompel, J.F.; Kim, J.S.; Wallach, C.J.; Robertson, D.D.; Vogt, M.T.; Kang, J.D.; Gilbertson, L.G. A slowly progressive and reproducible animal model of intervertebral disc degeneration characterized by MRI, X-ray, and histology. Spine (Phila. Pa. 1976) 2005, 30, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Schenk, P.; Läubli, T.; Hodler, J.; Klipstein, A. Magnetic resonance imaging of the lumbar spine: Findings in female subjects from administrative and nursing professions. Spine (Phila. Pa. 1976) 2006, 31, 2701–2706. [Google Scholar] [CrossRef] [PubMed]
- Waldenberg, C.; Hebelka, H.; Brisby, H.; Lagerstrand, K.M. MRI histogram analysis enables objective and continuous classification of intervertebral disc degeneration. Eur. Spine J. 2018, 27, 1042–1048. [Google Scholar] [CrossRef]
- Wu, P.H.; Kim, H.S.; Jang, I.-T. Intervertebral Disc Diseases PART 2: A Review of the Current Diagnostic and Treatment Strategies for Intervertebral Disc Disease. Int. J. Mol. Sci. 2020, 21, 2135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luan, S.; Wan, Q.; Luo, H.; Li, X.; Ke, S.; Lin, C.; Wu, Y.; Wu, S.; Ma, C. Running exercise alleviates pain and promotes cell proliferation in a rat model of intervertebral disc degeneration. Int. J. Mol. Sci. 2015, 16, 2130–2144. [Google Scholar] [CrossRef]
- Maroon, J.C.; Bost, J.W. Omega-3 fatty acids (fish oil) as an anti-inflammatory: An alternative to nonsteroidal anti-inflammatory drugs for discogenic pain. Surg. Neurol. 2006, 65, 326–331. [Google Scholar] [CrossRef] [PubMed]
- Stuber, K.; Sajko, S.; Kristmanson, K. Efficacy of glucosamine, chondroitin, and methylsulfonylmethane for spinal degenerative joint disease and degenerative disc disease: A systematic review. J. Can. Chiropr. Assoc. 2011, 55, 47–55. [Google Scholar]
- Lanas, A. Nonsteroidal antiinflammatory drugs and cyclooxygenase inhibition in the gastrointestinal tract: A trip from peptic ulcer to colon cancer. Am. J. Med. Sci. 2009, 338, 96–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brune, K.; Patrignani, P. New insights into the use of currently available non-steroidal anti-inflammatory drugs. J. Pain Res. 2015, 8, 105–118. [Google Scholar] [CrossRef] [Green Version]
- Atchison, J.W.; Herndon, C.M.; Rusie, E. NSAIDs for musculoskeletal pain management:current perspectives and novel strategies to improve safety. J. Manag. Care Pharm. 2013, 19, S3–S19. [Google Scholar] [PubMed] [Green Version]
- Errico, T.J. Lumbar disc arthroplasty. Clin. Orthop. Relat. Res. 2005, 435, 106–117. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.C.; Zotti, M.G.T.; Osti, O.L. Operative Management of Lumbar Degenerative Disc Disease. Asian Spine J. 2016, 10, 801–819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serhan, H.; Mhatre, D.; Defossez, H.; Bono, C.M. Motion-preserving technologies for degenerative lumbar spine: The past, present, and future horizons. SAS J. 2011, 5, 75–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zigler, J.; Delamarter, R.; Spivak, J.M.; Linovitz, R.J.; Danielson, G.O.I.I.I.; Haider, T.T.; Cammisa, F.; Zuchermann, J.; Balderston, R.; Kitchel, S.; et al. Results of the Prospective, Randomized, Multicenter Food and Drug Administration Investigational Device Exemption Study of the ProDisc®-L Total Disc Replacement Versus Circumferential Fusion for the Treatment of 1-Level Degenerative Disc Disease. Spine (Phila. Pa. 1976) 2007, 32, 1155–1162. [Google Scholar] [CrossRef] [PubMed]
- Link, H.D. History, design and biomechanics of the LINK SB Charité artificial disc. In Arthroplasty of the Spine; Gunzburg, R., Mayer, H.M., Szpalski, M., Aebi, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2004; pp. 36–43. [Google Scholar] [CrossRef] [Green Version]
- Guerrero, J.; Häckel, S.; Croft, A.S.; Hoppe, S.; Albers, C.E.; Gantenbein, B. The nucleus pulposus microenvironment in the intervertebral disc: The fountain of youth? Eur. Cell. Mater. 2021, 41, 707–738. [Google Scholar] [CrossRef] [PubMed]
- Chou, W.-Y.; Hsu, C.-J.; Chang, W.-N.; Wong, C.-Y. Adjacent segment degeneration after lumbar spinal posterolateral fusion with instrumentation in elderly patients. Arch. Orthop. Trauma Surg. 2002, 122, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Park, P.; Garton, H.J.; Gala, V.C.; Hoff, J.T.; McGillicuddy, J.E. Adjacent Segment Disease after Lumbar or Lumbosacral Fusion: Review of the Literature. Spine (Phila. Pa. 1976) 2004, 29, 1938–1944. [Google Scholar] [CrossRef] [PubMed]
- Leong, J.C.; Chun, S.Y.; Grange, W.J.; Fang, D. Long-term results of lumbar intervertebral disc prolapse. Spine (Phila. Pa. 1976) 1983, 8, 793–799. [Google Scholar] [CrossRef] [PubMed]
- Guigui, P.; Wodecki, P.; Bizot, P.; Lambert, P.; Chaumeil, G.; Deburge, A. Long-term influence of associated arthrodesis on adjacent segments in the treatment of lumbar stenosis: A series of 127 cases with 9-year follow-up. Rev. Chir. Orthop. Reparatrice Appar. Mot. 2000, 86, 546–557. [Google Scholar] [PubMed]
- Kuslich, S.D.; Danielson, G.; Dowdle, J.D.; Sherman, J.; Fredrickson, B.; Yuan, H.; Griffith, S.L. Four-year follow-up results of lumbar spine arthrodesis using the Bagby and Kuslich lumbar fusion cage. Spine (Phila. Pa. 1976) 2000, 25, 2656–2662. [Google Scholar] [CrossRef] [PubMed]
- Deyo, R.A.; Mirza, S.K.; Martin, B.I.; Kreuter, W.; Goodman, D.C.; Jarvik, J.G. Trends, major medical complications, and charges associated with surgery for lumbar spinal stenosis in older adults. JAMA 2010, 303, 1259–1265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buzhor, E.; Leshansky, L.; Blumenthal, J.; Barash, H.; Warshawsky, D.; Mazor, Y.; Shtrichman, R. Cell-based therapy approaches: The hope for incurable diseases. Regen. Med. 2014, 9, 649–672. [Google Scholar] [CrossRef] [PubMed]
- Richardson, S.M.; Hoyland, J.A.; Mobasheri, R.; Csaki, C.; Shakibaei, M.; Mobasheri, A. Mesenchymal stem cells in regenerative medicine: Opportunities and challenges for articular cartilage and intervertebral disc tissue engineering. J. Cell. Physiol. 2010, 222, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Pittenger, M.F.; Mackay, A.M.; Beck, S.C.; Jaiswal, R.K.; Douglas, R.; Mosca, J.D.; Moorman, M.A.; Simonetti, D.W.; Craig, S.; Marshak, D.R. Multilineage potential of adult human mesenchymal stem cells. Science 1999, 284, 143–147. [Google Scholar] [CrossRef] [Green Version]
- Erices, A.; Conget, P.; Minguell, J.J. Mesenchymal progenitor cells in human umbilical cord blood. Br. J. Haematol. 2000, 109, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Tuli, R.; Tuli, S.; Nandi, S.; Wang, M.L.; Alexander, P.G.; Haleem-Smith, H.; Hozack, W.J.; Manner, P.A.; Danielson, K.G.; Tuan, R.S. Characterization of multipotential mesenchymal progenitor cells derived from human trabecular bone. Stem Cells. 2003, 21, 681–693. [Google Scholar] [CrossRef] [PubMed]
- Zuk, P.A.; Zhu, M.; Mizuno, H.; Huang, J.; Futrell, J.W.; Katz, A.J.; Benhaim, P.; Lorenz, H.P.; Hedrick, M.H. Multilineage cells from human adipose tissue: Implications for cell-based therapies. Tissue Eng. 2001, 7, 211–228. [Google Scholar] [CrossRef] [Green Version]
- Young, H.E.; Steele, T.A.; Bray, R.A.; Hudson, J.; Floyd, J.A.; Hawkins, K.; Thomas, K.; Austin, T.; Edwards, C.; Cuzzourt, J.; et al. Human reserve pluripotent mesenchymal stem cells are present in the connective tissues of skeletal muscle and dermis derived from fetal, adult, and geriatric donors. Anat. Rec. 2001, 264, 51–62. [Google Scholar] [CrossRef]
- Jankowski, R.J.; Deasy, B.M.; Huard, J. Muscle-derived stem cells. Gene Ther. 2002, 9, 642–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alsalameh, S.; Amin, R.; Gemba, T.; Lotz, M. Identification of mesenchymal progenitor cells in normal and osteoarthritic human articular cartilage. Arthritis Rheum. 2004, 50, 1522–1532. [Google Scholar] [CrossRef]
- Clarke, L.E.; McConnell, J.C.; Sherratt, M.J.; Derby, B.; Richardson, S.M.; Hoyland, J.A. Growth differentiation factor 6 and transforming growth factor-beta differentially mediate mesenchymal stem cell differentiation, composition, and micromechanical properties of nucleus pulposus constructs. Arthritis Res. Ther. 2014, 16, R67. [Google Scholar] [CrossRef] [Green Version]
- Strassburg, S.; Richardson, S.M.; Freemont, A.J.; Hoyland, J.A. Co-culture induces mesenchymal stem cell differentiation and modulation of the degenerate human nucleus pulposus cell phenotype. Regen. Med. 2010, 5, 701–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strassburg, S.; Hodson, N.W.; Hill, P.I.; Richardson, S.M.; Hoyland, J.A. Bi-Directional Exchange of Membrane Components Occurs during Co-Culture of Mesenchymal Stem Cells and Nucleus Pulposus Cells. PLoS ONE 2012, 7, e33739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panebianco, C.J.; Meyers, J.H.; Gansau, J.; Hom, W.W.; Iatridis, J.C. Balancing biological and biomechanical performance in intervertebral disc repair: A systematic review of injectable cell delivery biomaterials. Eur. Cell. Mater. 2020, 40, 239–258. [Google Scholar] [CrossRef] [PubMed]
- Hohaus, C.; Ganey, T.M.; Minkus, Y.; Meisel, H.J. Cell transplantation in lumbar spine disc degeneration disease. Eur. Spine J. 2008, 17, 492–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nomura, T.; Mochida, J.; Okuma, M.; Nishimura, K.; Sakabe, K. Nucleus pulposus allograft retards intervertebral disc degeneration. Clin. Orthop. Relat. Res. 2001, 389, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Nassr, A.; Lee, J.Y.; Bashir, R.S.; Rihn, J.A.; Eck, J.C.; Kang, J.D.; Lim, M.R. Does incorrect level needle localization during anterior cervical discectomy and fusion lead to accelerated disc degeneration? Spine (Phila. Pa. 1976) 2009, 34, 189–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daly, C.; Ghosh, P.; Jenkin, G.; Oehme, D.; Goldschlager, T. A Review of Animal Models of Intervertebral Disc Degeneration: Pathophysiology, Regeneration, and Translation to the Clinic. Biomed Res. Int. 2016, 2016, 5952165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alini, M.; Eisenstein, S.M.; Ito, K.; Little, C.; Kettler, A.A.; Masuda, K.; Melrose, J.; Ralphs, J.; Stokes, I.; Wilke, H.J. Are animal models useful for studying human disc disorders/degeneration? Eur. Spine J. 2008, 17, 2–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iatridis, J.C.; Setton, L.A.; Weidenbaum, M.; Mow, V.C. The viscoelastic behavior of the non-degenerate human lumbar nucleus pulposus in shear. J. Biomech. 1997, 30, 1005–1013. [Google Scholar] [CrossRef]
- Cortes, D.H.; Jacobs, N.T.; DeLucca, J.F.; Elliott, D.M. Elastic, permeability and swelling properties of human intervertebral disc tissues: A benchmark for tissue engineering. J. Biomech. 2014, 47, 2088–2094. [Google Scholar] [CrossRef] [Green Version]
- Sakai, D. Stem cell regeneration of the intervertebral disk, Orthop. Clin. North Am. 2011, 42, 555–562. [Google Scholar] [CrossRef]
- Schol, J.; Sakai, D. Cell therapy for intervertebral disc herniation and degenerative disc disease: Clinical trials. Int. Orthop. 2019, 43, 1011–1025. [Google Scholar] [CrossRef]
- Clouet, J.; Fusellier, M.; Camus, A.; le Visage, C.; Guicheux, J. Intervertebral disc regeneration: From cell therapy to the development of novel bioinspired endogenous repair strategies. Adv. Drug Deliv. Rev. 2019, 146, 306–324. [Google Scholar] [CrossRef] [PubMed]
- Langer, R.; Vacanti, J.P. Tissue engineering. Science 1993, 260, 920–926. [Google Scholar] [CrossRef] [Green Version]
- Burdick, J.A.; Mauck, R.L.; Gerecht, S. To Serve and Protect: Hydrogels to Improve Stem Cell-Based Therapies. Cell Stem Cell 2016, 18, 13–15. [Google Scholar] [CrossRef] [Green Version]
- Schmitz, T.C.; Salzer, E.; Crispim, J.F.; Fabra, G.T.; LeVisage, C.; Pandit, A.; Tryfonidou, M.; le Maitre, C.; Ito, K. Characterization of biomaterials intended for use in the nucleus pulposus of degenerated intervertebral discs. Acta Biomater. 2020, 114, 1–15. [Google Scholar] [CrossRef]
- Mooney, D.J.; Vandenburgh, H. Cell Delivery Mechanisms for Tissue Repair. Cell Stem Cell 2008, 2, 205–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vadalà, G.; Sowa, G.; Hubert, M.; Gilbertson, L.G.; Denaro, V.; Kang, J.D. Mesenchymal stem cells injection in degenerated intervertebral disc: Cell leakage may induce osteophyte formation. J. Tissue Eng. Regen. Med. 2012, 6, 348–355. [Google Scholar] [CrossRef]
- Wuertz, K.; Godburn, K.; Neidlinger-Wilke, C.; Urban, J.; Iatridis, J.C. Behavior of Mesenchymal Stem Cells in the Chemical Microenvironment of the Intervertebral Disc. Spine (Phila. Pa. 1976) 2008, 33, 1843–1849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baumgartner, L.; Wuertz-Kozak, K.; le Maitre, C.L.; Wignall, F.; Richardson, S.M.; Hoyland, J.; Wills, C.R.; Ballester, M.A.G.; Neidlin, M.; Alexopoulos, L.G.; et al. Multiscale Regulation of the Intervertebral Disc: Achievements in Experimental, In Silico, and Regenerative Research. Int. J. Mol. Sci. 2021, 22, 703. [Google Scholar] [CrossRef]
- Gupta, R.C.; Lall, R.; Srivastava, A.; Sinha, A. Hyaluronic Acid: Molecular Mechanisms and Therapeutic Trajectory. Front. Vet. Sci. 2019, 6, 192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dagenais, S. Intra-articular hyaluronic acid (viscosupplementation) for knee osteoarthritis. Issues Emerg. Health Technol. 2006, 98, 1–4. [Google Scholar]
- Isa, I.L.M.; Srivastava, A.; Tiernan, D.; Owens, P.; Rooney, P.; Dockery, P.; Pandit, A. Hyaluronic Acid Based Hydrogels Attenuate Inflammatory Receptors and Neurotrophins in Interleukin-1β Induced Inflammation Model of Nucleus Pulposus Cells. Biomacromolecules 2015, 16, 1714–1725. [Google Scholar] [CrossRef] [PubMed]
- Isa, I.L.M.; Abbah, S.A.; Kilcoyne, M.; Sakai, D.; Dockery, P.; Finn, D.P.; Pandit, A. Implantation of hyaluronic acid hydrogel prevents the pain phenotype in a rat model of intervertebral disc injury. Sci. Adv. 2018, 4, eaaq0597. [Google Scholar] [CrossRef] [Green Version]
- Tokita, Y.; Okamoto, A. Hydrolytic degradation of hyaluronic acid. Polym. Degrad. Stab. 1995, 48, 269–273. [Google Scholar] [CrossRef]
- Quero, L.; Klawitter, M.; Schmaus, A.; Rothley, M.; Sleeman, J.; Tiaden, A.N.; Klasen, J.; Boos, N.; Hottiger, M.O.; Wuertz, K.; et al. Hyaluronic acid fragments enhance the inflammatory and catabolic response in human intervertebral disc cells through modulation of toll-like receptor 2 signalling pathways. Arthritis Res. Ther. 2013, 15, R94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalaf, E.A.G.; Pendyala, M.; Bledsoe, J.G.; Sell, S.A. Characterization and restoration of degenerated IVD function with an injectable, in situ gelling alginate hydrogel: An in vitro and ex vivo study. J. Mech. Behav. Biomed. Mater. 2017, 72, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Baer, A.E.; Wang, J.Y.; Kraus, V.B.; Setton, L.A. Collagen gene expression and mechanical properties of intervertebral disc cell-alginate cultures. J. Orthop. Res. 2001, 19, 2–10. [Google Scholar] [CrossRef]
- Chenite, A.; Chaput, C.; Wang, D.; Combes, C.; Buschmann, M.D.; Hoemann, C.D.; Leroux, J.C.; Atkinson, B.L.; Binette, F.; Selmani, A. Novel injectable neutral solutions of chitosan form biodegradable gels in situ. Biomaterials 2000, 21, 2155–2161. [Google Scholar] [CrossRef]
- Ahmadi, R.; de Bruijn, J.D. Biocompatibility and gelation of chitosan-glycerol phosphate hydrogels. J. Biomed. Mater. Res. A. 2008, 86, 824–832. [Google Scholar] [CrossRef] [PubMed]
- Roughley, P.; Hoemann, C.; DesRosiers, E.; Mwale, F.; Antoniou, J.; Alini, M. The potential of chitosan-based gels containing intervertebral disc cells for nucleus pulposus supplementation. Biomaterials 2006, 27, 388–396. [Google Scholar] [CrossRef]
- Sisson, K.; Zhang, C.; Farach-Carson, M.C.; Chase, D.B.; Rabolt, J.F. Evaluation of cross-linking methods for electrospun gelatin on cell growth and viability. Biomacromolecules 2009, 10, 1675–1680. [Google Scholar] [CrossRef] [PubMed]
- Slaughter, B.V.; Khurshid, S.S.; Fisher, O.Z.; Khademhosseini, A.; Peppas, N.A. Hydrogels in Regenerative Medicine. Adv. Mater. 2009, 21, 3307–3329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thorpe, A.A.; Dougill, G.; Vickers, L.; Reeves, N.D.; Sammon, C.; Cooper, G.; le Maitre, C.L. Thermally triggered hydrogel injection into bovine intervertebral disc tissue explants induces differentiation of mesenchymal stem cells and restores mechanical function. Acta Biomater. 2017, 54, 212–226. [Google Scholar] [CrossRef] [PubMed]
- Thorpe, A.A.; Boyes, V.L.; Sammon, C.; le Maitre, C.L. Thermally triggered injectable hydrogel, which induces mesenchymal stem cell differentiation to nucleus pulposus cells: Potential for regeneration of the intervertebral disc. Acta Biomater. 2016, 36, 99–111. [Google Scholar] [CrossRef] [PubMed]
- Haq, M.A.; Su, Y.; Wang, D. Mechanical properties of PNIPAM based hydrogels: A review. Mater. Sci. Eng. C. 2017, 70, 842–855. [Google Scholar] [CrossRef]
- Woodard, L.N.; Grunlan, M.A. Hydrolytic Degradation and Erosion of Polyester Biomaterials. ACS Macro Lett. 2018, 7, 976–982. [Google Scholar] [CrossRef] [Green Version]
- Tibbitt, M.W.; Anseth, K.S. Hydrogels as extracellular matrix mimics for 3D cell culture. Biotechnol. Bioeng. 2009, 103, 655–663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peppas, N.A.; Hilt, J.Z.; Khademhosseini, A.; Langer, R. Hydrogels in Biology and Medicine: From Molecular Principles to Bionanotechnology. Adv. Mater. 2006, 18, 1345–1360. [Google Scholar] [CrossRef]
- Gough, J.E.; Saiani, A.; Miller, A.F. Peptide hydrogels: Mimicking the extracellular matrix. Bioinspired Biomim. Nanobiomaterials 2012, 1, 4–12. [Google Scholar] [CrossRef]
- Zhang, S. Fabrication of novel biomaterials through molecular self-assembly. Nat. Biotechnol. 2003, 21, 1171–1178. [Google Scholar] [CrossRef]
- Zhang, S.; Holmes, T.C.; DiPersio, C.M.; Hynes, R.O.; Su, X.; Rich, A. Self-complementary oligopeptide matrices support mammalian cell attachment. Biomaterials 1995, 16, 1385–1393. [Google Scholar] [CrossRef]
- Engler, A.J.; Sen, S.; Sweeney, H.L.; Discher, D.E. Matrix Elasticity Directs Stem Cell Lineage Specification. Cell 2006, 126, 677–689. [Google Scholar] [CrossRef] [Green Version]
- Guvendiren, M.; Lu, H.D.; Burdick, J.A. Shear-thinning hydrogels for biomedical applications. Soft Matter 2012, 8, 260–272. [Google Scholar] [CrossRef]
- Morris, O.; Elsawy, M.A.; Fairclough, M.; Williams, K.J.; Mcmahon, A.; Grigg, J.; Forster, D.; Miller, A.F.; Saiani, A.; Prenant, C. In vivo characterisation of a therapeutically relevant self-assembling (18) F-labelled β-sheet forming peptide and its hydrogel using positron emission tomography. J. Labelled Comp. Radiopharm. 2017, 60, 481–488. [Google Scholar] [CrossRef] [Green Version]
- Markey, A.; Workman, V.L.; Bruce, I.A.; Woolford, T.J.; Derby, B.; Miller, A.F.; Cartmell, S.H.; Saiani, A. Peptide hydrogel in vitro non-inflammatory potential. J. Pept. Sci. 2016, 23, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Nagarkar, R.P.; Schneider, J.P. Synthesis and primary characterization of self-assembled peptide-based hydrogels. Methods Mol. Biol. 2008, 474, 61–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gelain, F.; Luo, Z.; Rioult, M.; Zhang, S. Self-assembling peptide scaffolds in the clinic. npj Regen. Med. 2021, 6, 9. [Google Scholar] [CrossRef] [PubMed]
- Ulijn, R.V.; Smith, A.M. Designing peptide based nanomaterials. Chem. Soc. Rev. 2008, 37, 664. [Google Scholar] [CrossRef]
- Aggeli, A.; Nyrkova, I.A.; Bell, M.; Harding, R.; Carrick, L.; McLeish, T.C.B.; Semenov, A.N.; Boden, N. Hierarchical self-assembly of chiral rod-like molecules as a model for peptide β-sheet tapes, ribbons, fibrils, and fibers. Proc. Natl. Acad. Sci. USA 2001, 98, 11857–11862. [Google Scholar] [CrossRef] [Green Version]
- Ding, X.; Zhao, H.; Li, Y.; Lee, A.L.; Li, Z.; Fu, M.; Li, C.; Yang, Y.Y.; Yuan, P. Synthetic peptide hydrogels as 3D scaffolds for tissue engineering. Adv. Drug Deliv. Rev. 2020, 160, 78–104. [Google Scholar] [CrossRef] [PubMed]
- Kisiday, J.; Jin, M.; Kurz, B.; Hung, H.; Semino, C.; Zhang, S.; Grodzinsky, A.J. Self-assembling peptide hydrogel fosters chondrocyte extracellular matrix production and cell division: Implications for cartilage tissue repair. Proc. Natl. Acad. Sci. USA 2002, 99, 9996–10001. [Google Scholar] [CrossRef] [Green Version]
- Kopesky, P.W.; Vanderploeg, E.J.; Sandy, J.S.; Kurz, B.; Grodzinsky, A.J. Self-Assembling Peptide Hydrogels Modulate In Vitro Chondrogenesis of Bovine Bone Marrow Stromal Cells. Tissue Eng. Part A 2010, 16, 465–477. [Google Scholar] [CrossRef] [Green Version]
- Bian, Z.; Sun, J. Development of a KLD-12 polypeptide/TGF-β1-tissue scaffold promoting the differentiation of mesenchymal stem cell into nucleus pulposus-like cells for treatment of intervertebral disc degeneration. Int. J. Clin. Exp. Pathol. 2015, 8, 1093–1103. [Google Scholar]
- Tripathi, J.K.; Pal, S.; Awasthi, B.; Kumar, A.; Tandon, A.; Mitra, K.; Chattopadhyay, N.; Ghosh, J.K. Variants of self-assembling peptide, KLD-12 that show both rapid fracture healing and antimicrobial properties. Biomaterials 2015, 56, 92–103. [Google Scholar] [CrossRef]
- Saiani, A.; Mohammed, A.; Frielinghaus, H.; Collins, R.; Hodson, N.; Kielty, C.M.; Sherratt, M.J.; Miller, A.F. Self-assembly and gelation properties of [small alpha]-helix versus [small beta]-sheet forming peptides. Soft Matter 2009, 5, 193–202. [Google Scholar] [CrossRef]
- Diaz, L.A.C.; Saiani, A.; Gough, J.E.; Miller, A.F.; Diaz, L.A.C.; Saiani, A.; Gough, J.E.; Miller, A.F.; Diaz, L.A.C.; Saiani, A.; et al. Human osteoblasts within soft peptide hydrogels promote mineralisation in vitro. J. Tissue Eng. 2014, 5, 2041731414539344. [Google Scholar] [CrossRef] [Green Version]
- Mujeeb, A.; Miller, A.F.; Saiani, A.; Gough, J.E. Self-assembled octapeptide scaffolds for in vitro chondrocyte culture. Acta Biomater. 2013, 9, 4609–4617. [Google Scholar] [CrossRef] [PubMed]
- Wan, S.; Borland, S.; Richardson, S.M.; Merry, C.L.R.; Saiani, A.; Gough, J.E.; Wan, S.; Wan, S.; Borland, S.; Richardson, S.M.; et al. Self-Assembling Peptide Hydrogel for Intervertebral Disc Tissue Engineering. Acta Biomater. 2016, 46, 29–40. [Google Scholar] [CrossRef]
- Burgess, K.A.; Frati, C.; Meade, K.; Gao, J.; Diaz, L.C.; Madeddu, D.; Graiani, G.; Cavalli, S.; Miller, A.F.; Oceandy, D.; et al. Functionalised peptide hydrogel for the delivery of cardiac progenitor cells. Mater. Sci. Eng. C 2021, 119, 111539. [Google Scholar] [CrossRef]
- Imere, A.; Ligorio, C.; O’Brien, M.; Wong, J.; Domingos, M.; Cartmell, S. Engineering a cell-hydrogel-fibre composite to mimic the structure and function of the tendon synovial sheath. Acta Biomater. 2021, 119, 140–154. [Google Scholar] [CrossRef]
- Wychowaniec, J.K.; Smith, A.M.; Ligorio, C.; Mykhaylyk, O.O.; Miller, A.F.; Saiani, A. Role of Sheet-Edge Interactions in β-sheet Self-Assembling Peptide Hydrogels. Biomacromolecules 2020, 21, 2285–2297. [Google Scholar] [CrossRef]
- Chiesa, I.; Ligorio, C.; Bonatti, A.F.; de Acutis, A.; Smith, A.M.; Saiani, A.; Vozzi, G.; de Maria, C. Modeling the Three-Dimensional Bioprinting Process of β-Sheet Self-Assembling Peptide Hydrogel Scaffolds. Front. Med. Technol. 2020, 2, 4. [Google Scholar] [CrossRef]
- Schneider, J.P.; Pochan, D.J.; Ozbas, B.; Rajagopal, K.; Pakstis, L.; Kretsinger, J. Responsive hydrogels from the intramolecular folding and self-assembly of a designed peptide. J. Am. Chem. Soc. 2002, 124, 15030–15037. [Google Scholar] [CrossRef]
- Yan, C.; Altunbas, A.; Yucel, T.; Nagarkar, R.P.; Schneider, J.P.; Pochan, D.J. Injectable solid hydrogel: Mechanism of shear-thinning and immediate recovery of injectable β-hairpin peptide hydrogels. Soft Matter 2010, 6, 5143–5156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindsey, S.; Piatt, J.H.; Worthington, P.; Sönmez, C.; Satheye, S.; Schneider, J.P.; Pochan, D.J.; Langhans, S.A. Beta Hairpin Peptide Hydrogels as an Injectable Solid Vehicle for Neurotrophic Growth Factor Delivery. Biomacromolecules 2015, 16, 2672–2683. [Google Scholar] [CrossRef]
- Altunbas, A.; Lee, S.J.; Rajasekaran, S.A.; Schneider, J.P.; Pochan, D.J. Encapsulation of curcumin in self-assembling peptide hydrogels as injectable drug delivery vehicles. Biomaterials 2011, 32, 5906–5914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pauling, L.; Corey, R.B. Compound Helical Configurations of Polypeptide Chains: Structure of Proteins of the α-Keratin Type. Nature 1953, 171, 59–61. [Google Scholar] [CrossRef]
- Banwell, E.F.; Abelardo, E.S.; Adams, D.J.; Birchall, M.A.; Corrigan, A.; Donald, A.M.; Kirkland, M.; Serpell, L.C.; Butler, M.F.; Woolfson, D.N. Rational design and application of responsive α-helical peptide hydrogels. Nat. Mater. 2009, 8, 596–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lapenta, F.; Aupič, J.; Strmšek, Ž.; Jerala, R. Coiled coil protein origami: From modular design principles towards biotechnological applications. Chem. Soc. Rev. 2018, 47, 3530–3542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woolfson, D.N.; Ryadnov, M.G. Peptide-based fibrous biomaterials: Some things old, new and borrowed. Curr. Opin. Chem. Biol. 2006, 10, 559–567. [Google Scholar] [CrossRef]
- Dawson, W.M.; Martin, F.J.O.; Rhys, G.G.; Shelley, K.L.; Brady, R.L.; Woolfson, D.N. Coiled coils 9-to-5: Rational de novo design of α-helical barrels with tunable oligomeric states. Chem. Sci. 2021, 12, 6923–6928. [Google Scholar] [CrossRef] [PubMed]
- MacPhee, C.E.; Woolfson, D.N. Engineered and designed peptide-based fibrous biomaterials. Curr. Opin. Solid State Mater. Sci. 2004, 8, 141–149. [Google Scholar] [CrossRef]
- Mehrban, N.; Zhu, B.; Tamagnini, F.; Young, F.I.; Wasmuth, A.; Hudson, K.L.; Thomson, A.R.; Birchall, M.A.; Randall, A.D.; Song, B.; et al. Functionalized α-Helical Peptide Hydrogels for Neural Tissue Engineering. ACS Biomater. Sci. Eng. 2015, 1, 431–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehrban, N.; Molina, C.P.; Quijano, L.M.; Bowen, J.; Johnson, S.A.; Bartolacci, J.; Chang, J.T.; Scott, D.A.; Woolfson, D.N.; Birchall, M.A.; et al. Host macrophage response to injectable hydrogels derived from ECM and α-helical peptides. Acta Biomater. 2020, 111, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Reches, M.; Gazit, E. Self-assembly of peptide nanotubes and amyloid-like structures by charged-termini-capped diphenylalanine peptide analogues. Isr. J. Chem. 2005, 45, 363–371. [Google Scholar] [CrossRef]
- Smith, A.M.; Williams, R.J.; Tang, C.; Coppo, P.; Collins, R.F.; Turner, M.L.; Saiani, A.; Ulijn, R.V. Fmoc-Diphenylalanine Self Assembles to a Hydrogel via a Novel Architecture Based on π–π Interlocked β-Sheets. Adv. Mater. 2008, 20, 37–41. [Google Scholar] [CrossRef]
- Jayawarna, V.; Smith, A.; Gough, J.E.; Ulijn, R.V. Three-dimensional cell culture of chondrocytes on modified di-phenylalanine scaffolds. Biochem. Soc. Trans. 2007, 35, 535–537. [Google Scholar] [CrossRef] [Green Version]
- Zhou, M.; Smith, A.M.; Das, A.K.; Hodson, N.W.; Collins, R.F.; Ulijn, R.V.; Gough, J.E. Self-assembled peptide-based hydrogels as scaffolds for anchorage-dependent cells. Biomaterials 2009, 30, 2523–2530. [Google Scholar] [CrossRef]
- Jayawarna, V.; Richardson, S.M.; Hirst, A.R.; Hodson, N.W.; Saiani, A.; Gough, J.E.; Ulijn, R.V. Introducing chemical functionality in Fmoc-peptide gels for cell culture. Acta Biomater. 2009, 5, 934–943. [Google Scholar] [CrossRef]
- Mata, A.; Hsu, L.; Capito, R.; Aparicio, C.; Henrikson, K.; Stupp, S.I. Micropatterning of bioactive self-assembling gels. Soft Matter 2009, 5, 1228–1236. [Google Scholar] [CrossRef] [PubMed]
- Webber, M.J.; Tongers, J.; Renault, M.-A.; Roncalli, J.G.; Losordo, D.W.; Stupp, S.I. Development of bioactive peptide amphiphiles for therapeutic cell delivery. Acta Biomater. 2010, 6, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Berndt, P.; Fields, G.B.; Tirrell, M. Synthetic lipidation of peptides and amino acids: Monolayer structure and properties. J. Am. Chem. Soc. 1995, 117, 9515–9522. [Google Scholar] [CrossRef]
- Chow, L.W.; Bitton, R.; Webber, M.J.; Carvajal, D.; Shull, K.R.; Sharma, A.K.; Stupp, S.I. A bioactive self-assembled membrane to promote angiogenesis. Biomaterials 2011, 32, 1574–1582. [Google Scholar] [CrossRef] [Green Version]
- Shah, R.N.; Shah, N.A.; Lim, M.M.d.; Hsieh, C.; Nuber, G.; Stupp, S.I. Supramolecular design of self-assembling nanofibers for cartilage regeneration. Proc. Natl. Acad. Sci. USA 2010, 107, 3293–3298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webber, M.J.; Stupp, S.I. Emerging Peptide Nanomedicine to Regenerate Tissues and Organs. J. Intern. Med. 2010, 267, 71–88. [Google Scholar] [CrossRef]
- Zhou, S.; Hokugo, A.; McClendon, M.; Zhang, Z.; Bakshi, R.; Wang, L.; Segovia, L.A.; Rezzadeh, K.; Stupp, S.I.; Jarrahy, R. Bioactive peptide amphiphile nanofiber gels enhance burn wound healing. Burns 2019, 45, 1112–1121. [Google Scholar] [CrossRef] [PubMed]
- Li, X.-C.; Wu, Y.-H.; Bai, X.-D.; Ji, W.; Guo, Z.-M.; Wang, C.-F.; He, Q.; Ruan, D. BMP7-Based Functionalized Self-Assembling Peptides Protect Nucleus Pulposus-Derived Stem Cells From Apoptosis In Vitro. Tissue Eng. Part A 2016, 22, 1218–1228. [Google Scholar] [CrossRef]
- An, H.S.; Takegami, K.; Kamada, H.; Nguyen, C.M.; Thonar, E.J.-M.A.; Singh, K.; Andersson, G.B.; Masuda, K. Intradiscal administration of osteogenic protein-1 increases intervertebral disc height and proteoglycan content in the nucleus pulposus in normal adolescent rabbits. Spine (Phila. Pa. 1976) 2005, 30, 22–25. [Google Scholar] [CrossRef] [PubMed]
- Gu, T.; Shi, Z.; Wang, C.; Chen, C.; Wu, J.; Wang, D.; Xu, C.; Qing, H.; Dike, R. Human bone morphogenetic protein 7 transfected nucleus pulposus cells delay the degeneration of intervertebral disc in dogs. J. Orthop. Res. Off. Publ. Orthop. Res. Soc. 2017, 35, 1311–1322. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Jia, Z.; Liu, L.; Zhao, Y.; Li, H.; Wang, C.; Tao, H.; Tang, Y.; He, Q.; Ruan, D. Functional Self-Assembled Peptide Nanofibers for Bone Marrow Mesenchymal Stem Cell Encapsulation and Regeneration in Nucleus Pulposus. Artif. Organs 2016, 40, E112–E119. [Google Scholar] [CrossRef]
- Tao, H.; Zhang, Y.; Wang, C.; Zhang, C.; Wang, X.; Wang, D.; Bai, X.; Wen, T.; Xin, H.; Wu, J.; et al. Biological Evaluation of Human Degenerated Nucleus Pulposus Cells in Functionalized Self-Assembling Peptide Nanofiber Hydrogel Scaffold. Tissue Eng. Part A 2014, 20, 1621–1631. [Google Scholar] [CrossRef] [PubMed]
- McKenna, L.A.; Liu, H.; Sansom, P.A.; Dean, M.F. An N-terminal peptide from link protein stimulates proteoglycan biosynthesis in human articular cartilage in vitro. Arthritis Rheum. 1998, 41, 157–162. [Google Scholar] [CrossRef]
- Wang, B.; Wu, Y.; Shao, Z.; Yang, S.; Che, B.; Sun, C.; Ma, Z.; Zhang, Y. Functionalized self-assembling peptide nanofiber hydrogel as a scaffold for rabbit nucleus pulposus cells. J. Biomed. Mater. Res.—Part A 2012, 100, 646–653. [Google Scholar] [CrossRef]
- Ma, K.; Wu, Y.; Wang, B.; Yang, S.; Wei, Y.; Shao, Z. Effect of a synthetic link N peptide nanofiber scaffold on the matrix deposition of aggrecan and type II collagen in rabbit notochordal cells. J. Mater. Sci. Mater. Med. 2013, 24, 405–415. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Zheng, Q.; Wu, Y.; Liu, Y.; Guo, X.; Wu, W. Culture of nucleus pulposus cells from intervertebral disc on self-assembling KLD-12 peptide hydrogel scaffold. Mater. Sci. Eng. C 2010, 30, 975–980. [Google Scholar] [CrossRef]
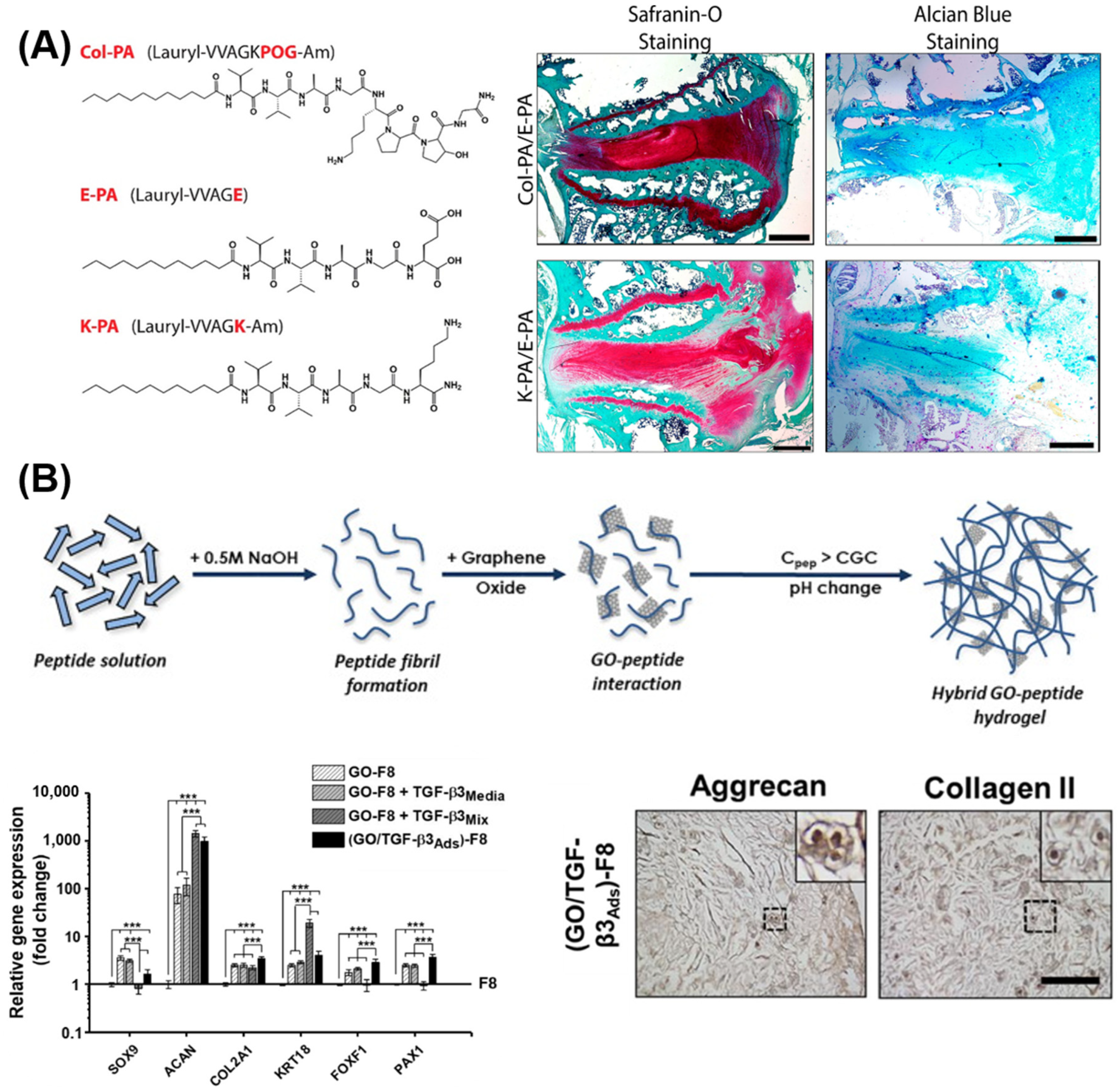
- Ligorio, C.; Zhou, M.; Wychowaniec, J.K.; Zhu, X.; Bartlam, C.; Miller, A.F.; Vijayaraghavan, A.; Hoyland, J.A.; Saiani, A. Graphene oxide containing self-assembling peptide hybrid hydrogels as a potential 3D injectable cell delivery platform for intervertebral disc repair applications. Acta Biomater. 2019, 92, 92–103. [Google Scholar] [CrossRef] [PubMed]
- Ligorio, C.; O’Brien, M.; Hodson, N.W.; Mironov, A.; Iliut, M.; Miller, A.F.; Vijayaraghavan, A.; Hoyland, J.A.; Saiani, A. TGF-β3-loaded graphene oxide—Self-assembling peptide hybrid hydrogels as functional 3D scaffolds for the regeneration of the nucleus pulposus. Acta Biomater. 2021, 127, 116–130. [Google Scholar] [CrossRef]
- Mishra, A.; Loo, Y.; Deng, R.; Chuah, Y.J.; Hee, H.T.; Ying, J.Y.; Hauser, C.A.E. Ultrasmall natural peptides self-assemble to strong temperature-resistant helical fibers in scaffolds suitable for tissue engineering. Nano Today 2011, 6, 232–239. [Google Scholar] [CrossRef]
- Uysal, O.; Arslan, E.; Gulseren, G.; Kilinc, M.C.; Dogan, I.; Ozalp, H.; Caglar, Y.S.; Guler, M.O.; Tekinay, A.B. Collagen Peptide Presenting Nanofibrous Scaffold for Intervertebral Disc Regeneration. ACS Appl. Bio Mater. 2019, 2, 1686–1695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barreto-Henriksson, H.; Llorente, M.; Larsson, A.; Brisby, H.; Gold, J.; Schuster, E.; Ström, A. Determination of mechanical and rheological properties of a cell-loaded peptide gel during ECM production. Int. J. Pharm. 2019, 563, 437–444. [Google Scholar] [CrossRef] [PubMed]
- Moss, I.L.; Gordon, L.; Woodhouse, K.A.; Whyne, C.M.; Yee, A.J.M. A novel thiol-modified hyaluronan and elastin-like polypetide composite material for tissue engineering of the nucleus pulposus of the intervertebral disc. Spine (Phila. Pa. 1976) 2011, 36, 1022–1029. [Google Scholar] [CrossRef] [PubMed]
- Miles, D.E.; Mitchell, E.A.; Kapur, N.; Beales, P.A.; Wilcox, R.K. Peptide:glycosaminoglycan hybrid hydrogels as an injectable intervention for spinal disc degeneration. J. Mater. Chem. B. 2016, 4, 3225–3231. [Google Scholar] [CrossRef] [Green Version]
- Yoon, H.H.; Bhang, S.H.; Kim, T.; Yu, T.; Hyeon, T.; Kim, B.-S. Dual Roles of Graphene Oxide in Chondrogenic Differentiation of Adult Stem Cells: Cell-Adhesion Substrate and Growth Factor-Delivery Carrier. Adv. Funct. Mater. 2014, 24, 6455–6464. [Google Scholar] [CrossRef]
- Zhou, M.; Lozano, N.; Wychowaniec, J.K.; Hodgkinson, T.; Richardson, S.M.; Kostarelos, K.; Hoyland, J.A. Graphene oxide: A growth factor delivery carrier to enhance chondrogenic differentiation of human mesenchymal stem cells in 3D hydrogels. Acta Biomater. 2019, 96, 271–280. [Google Scholar] [CrossRef]
- Raphael, B.; Khalil, T.; Workman, V.L.; Smith, A.; Brown, C.P.; Streuli, C.; Saiani, A.; Domingos, M. 3D cell bioprinting of self-assembling peptide-based hydrogels. Mater. Lett. 2017, 190, 103–106. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Grade | Structure Colour | NP/AF Distinction | Signal Intensity | IVD Height |
|---|---|---|---|---|
| I | Homogenous, bright white | Clear | Hyperintense | Normal |
| II | Inhomogeneous, opaque | Clear | Hyperintense | Normal |
| III | Inhomogeneous and grey | Unclear | Intermediate | Normal to slightly decreased |
| IV | Inhomogeneous, grey to black | Lost | Intermediate to Hypointense | Normal to moderately decreased |
| V | Inhomogeneous, black | Lost | Hypointense | Collapse of disc space |
| Hydrogels | Relevant Examples | Main Advantages | Main Disadvantages |
|---|---|---|---|
| Natural | Hyaluronic Acid, alginate, chitosan, fibrin, gelatin, cellulose and gellan gum | Biodegradable Cell-binding sites Low immunogenicity Relatively cheap | Poor mechanical properties High degradation rates Batch-to-batch variability |
| Synthetic | poly(N-isopropylacrylamide), poly(ethylene glycol), poly(vinyl alcohol), poly(vinyl pyrrolidone) | Tuneable mechanical properties and shapes Tuneable degradation Ease of scalability | Lack of cell-recognition sites Toxic degradation by-products Toxic cross-linkers |
| Hydrogel | Injectable? | Cell Type | Duration | Outcomes | Refs. |
|---|---|---|---|---|---|
| RKP (RADA16 functionalised with KPSS) | N/A | Human NP-derived stem cells | 2 days | High cell proliferation and upregulation of the gene expression of collagen II, aggrecan, and Sox-9 compared to controls | [224] |
| RADA16:RADA-KPSS in 1:1 ratio | Yes | Human MSCs | 14 days | Hydrogels promoted cell proliferation and chemotaxis as well as kept cell viability up to 14 days once injected into an ex vivo cultured disc mode | [227] |
| RAD-RKP (co-assembly of RADA16 with RKP) | Yes | Degenerated human NP cells | 28 days | High cell proliferation and migration. Collagen type II, SOX9, and aggrecan were upregulated, while collagen I was downregulated compared to controls | [228] |
| LN-NS (co-assembly of link N-conjugated RADA16 with RADA16) | Yes | Rabbit NP cells | 14 days | High viability and cell adhesion. Increased gene expression of PGs and collagen type II compared to controls | [230] |
| LN-NS (1:1 mixture of link N-conjugated RADA16 with RADA16) | Yes | Rabbit Notochordal Cells | 14 days | High cell survival rate. Increased gene expression of aggrecan and collagen type II compared to controls | [231] |
| KLD12 | Yes | Rabbit NP cells | 14 days | Increased GAG and collagen type II production over time. NP cells preserved high viability and characteristic shape | [232] |
| KLD12/TGF-β1 | N/A | Rabbit MSCs | 14 days | High cell viability over time. Increased gene expression and protein deposition of aggrecan and collagen type II compared to controls | [190] |
| FEFEFKFK | Yes | Bovine NP Cells | 14 days | Characteristic morphology and high viability over time. Upregulation of collagen type II, aggrecan, cytokeratin-8, cytokeratin-18, SOX9, and FOXF1, CA12 | [195] |
| FEFKFEFK-GO | Yes | Bovine NP cells | 7 days | Increased viability of NP cells and stable metabolic activity over time | [233] |
| FEFKFEFK-(GO + TGF-β3) | Yes | Bovine NP cells | 21 days | TGF-β3-decorated GO flakes induced increased gene expression and matrix deposition over time compared to TGF-β3 ‘free’ in peptide hydrogels or added exogenously. SMAD signalling was preserved when TGF-β3 was anchored on GO flakes, inducing ECM production | [234] |
| Ac-ID3, Ac-LD6, Ac-AD6 | N/A | Porcine NP cells | 2 days | Porcine NP cells showed good viability when incubated with peptides | [235] |
| Col-PA/E-PA (PA hydrogel decorated with collagen-mimic POG motif) | Yes | Rat MSCs | 14 days | In vitro differentiation into NP-like cells. Increased GAGs and collagen deposition compared to controls. Functional recovery of rabbit IVDs after injection in vivo | [236] |
| PuraMatrix® | Yes | Human MSCs | 21 days | High deposition of GAGs accompanied by fast stress relaxation and mechanical properties mimicking native NP tissue | [237] |
| Elastin-like polypeptide/thiol-modified hyaluronan | Yes | Human IVD cells | 21 days | High cell viability and maintenance of anabolic cell phenotype. Good biocompatibility in a preclinical rabbit annular puncture model | [238] |
| P11-4, P11-8, P11-9, P11-12/chondroitin sulphate | Yes | Ex vivo bovine caudal spine model | 1 day | Hybrid hydrogels showed mechanical properties similar to the human NP. Hydrogels injected in a caudal spine model undergo gelation in situ with minimal leakage over time | [239] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ligorio, C.; Hoyland, J.A.; Saiani, A. Self-Assembling Peptide Hydrogels as Functional Tools to Tackle Intervertebral Disc Degeneration. Gels 2022, 8, 211. https://doi.org/10.3390/gels8040211
Ligorio C, Hoyland JA, Saiani A. Self-Assembling Peptide Hydrogels as Functional Tools to Tackle Intervertebral Disc Degeneration. Gels. 2022; 8(4):211. https://doi.org/10.3390/gels8040211
Chicago/Turabian StyleLigorio, Cosimo, Judith A. Hoyland, and Alberto Saiani. 2022. "Self-Assembling Peptide Hydrogels as Functional Tools to Tackle Intervertebral Disc Degeneration" Gels 8, no. 4: 211. https://doi.org/10.3390/gels8040211
APA StyleLigorio, C., Hoyland, J. A., & Saiani, A. (2022). Self-Assembling Peptide Hydrogels as Functional Tools to Tackle Intervertebral Disc Degeneration. Gels, 8(4), 211. https://doi.org/10.3390/gels8040211