
Nutritional, Bioactive, and Flavor Components of Giant Stropharia (Stropharia rugoso-annulata): A Review

Abstract
1. Introduction
2. Nutritional Components of S. rugoso-annulata
2.1. Nutritional Components of S. rugoso-annulata Processed by Different Drying Methods
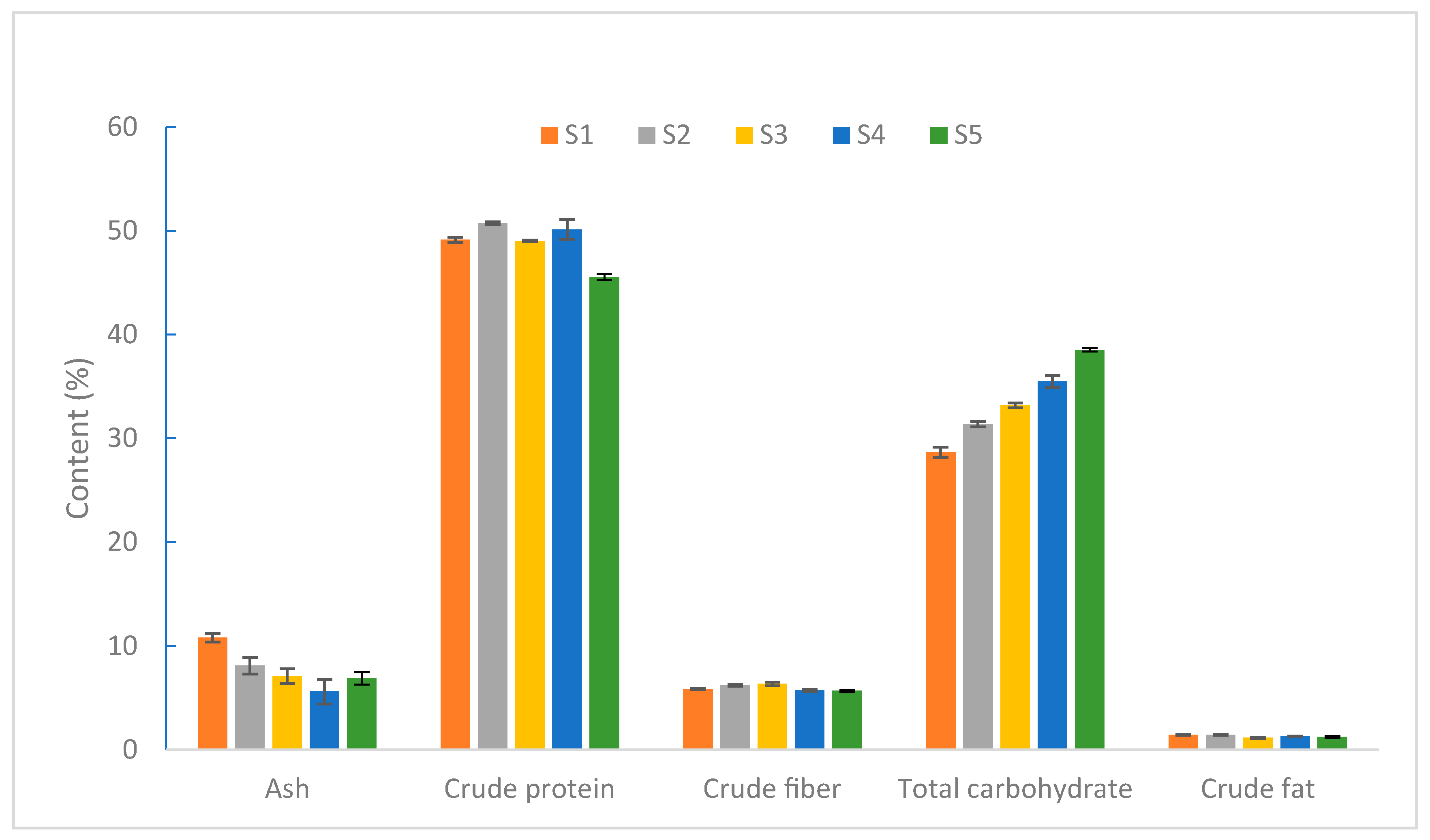
2.2. Nutritional Components of S. rugoso-annulata at Different Developmental Stages
2.3. Nutritional Components of Different Parts of S. rugoso-annulata
3. Bioactive Components of S. rugoso-annulata
3.1. Soluble Polysaccharides
3.2. Other Bioactive Components

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Components | Bioactivity | Optimal Extraction Conditions | Yield | References |
|---|---|---|---|---|
| Protein | Antioxidant activity and scavenging ability on DPPH radicals and hydroxyl radicals | Distilled water, pH = 12, 1:30 (w/v), 60 min, 45 °C | 37.54% | [54] |
| Oligopeptide | Antioxidant, ACE inhibitory activity | Pure water, 1:20 (w/v), ultrasound 120 w–400 w, 20 kHz, 10–35 min | 11.04–23.02% | [60] |
| Ergosterol | Precursor of Vitamin D2, anti-cancer, anti-aging | Ethanol 100%, 1:30 (w/v), 30 min, ultrasound, 3 repeats | 0.23% | [17] |
| Phenol | Antioxidant, antibacterial, anticancer, anti-aging, and inhibition of cholesterol elevation | Ethanol 30%, 1:20 (w/v), 70 °C, 1 h | 11.00% | [55] |
| Ethanol 30%, 1:20 (w/v), 60 °C, 6 min, ultrasound 240 w | 6.71% | |||
| Ethanol 35%, 1:20 (w/v), 2.5 min, microwave 640 w | 5.32% | |||
| Polyphenol | Antioxidation, antivirus, antibacterial | Ethanol 64.68%, 53.1 °C, ultrasound 39.3 min | 1.66% | [66] |
| Flavonoid | Antivirus, antioxidant, antibacterial, and protection of the cardiovascular and cerebrovascular systems. | Ethanol 30%, 1:15 (w/v), ultrasound 120 w, 1 h | 1.14% | [56] |
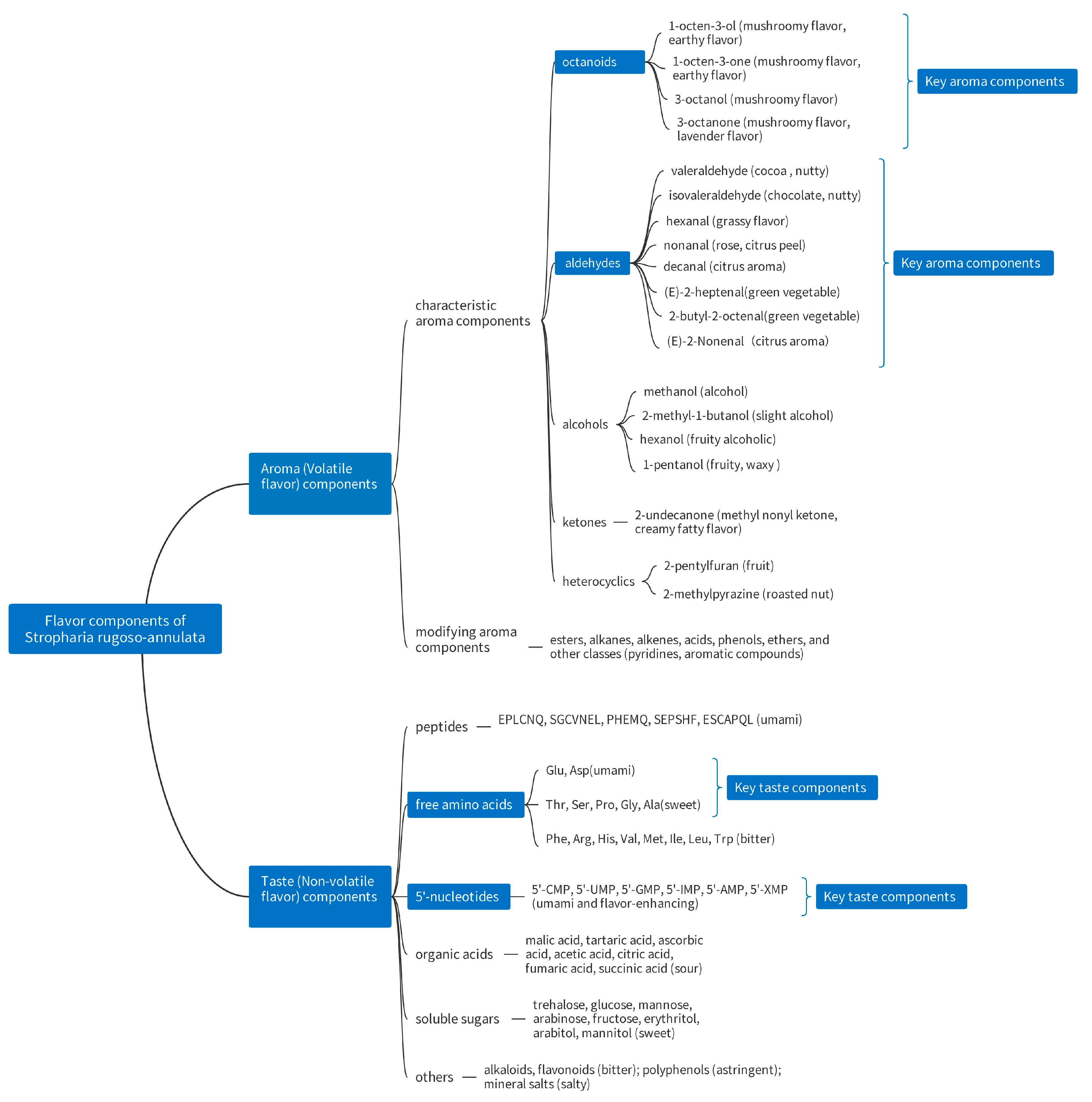
4. Flavor Components of S. rugoso-annulata
4.1. Taste (Non-Volatile Flavor) Components of S. rugoso-annulata
4.1.1. Taste Peptides
4.1.2. Soluble Sugars
4.1.3. Free Amino Acids
4.1.4. 5′-Nucleotides
4.1.5. Organic Acids
4.1.6. Other Taste Components
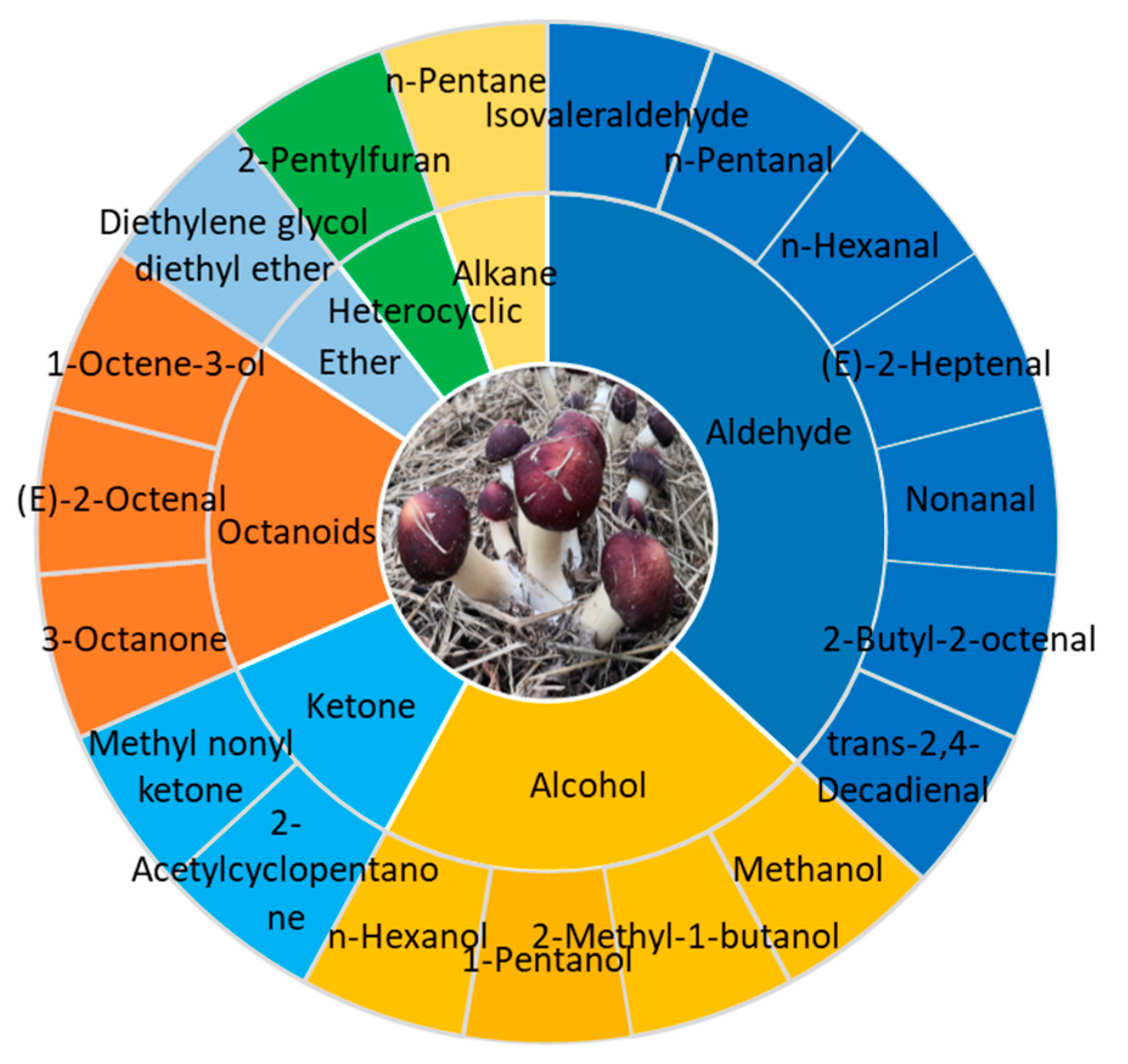
4.2. Aroma (Volatile Flavor) Components of S. rugoso-annulata
4.3. Factors Influencing Flavor Components of S. rugoso-annulata
4.3.1. Taste Components in Different Parts of S. rugoso-annulata
4.3.2. Aroma Components in Different Parts of S. rugoso-annulata
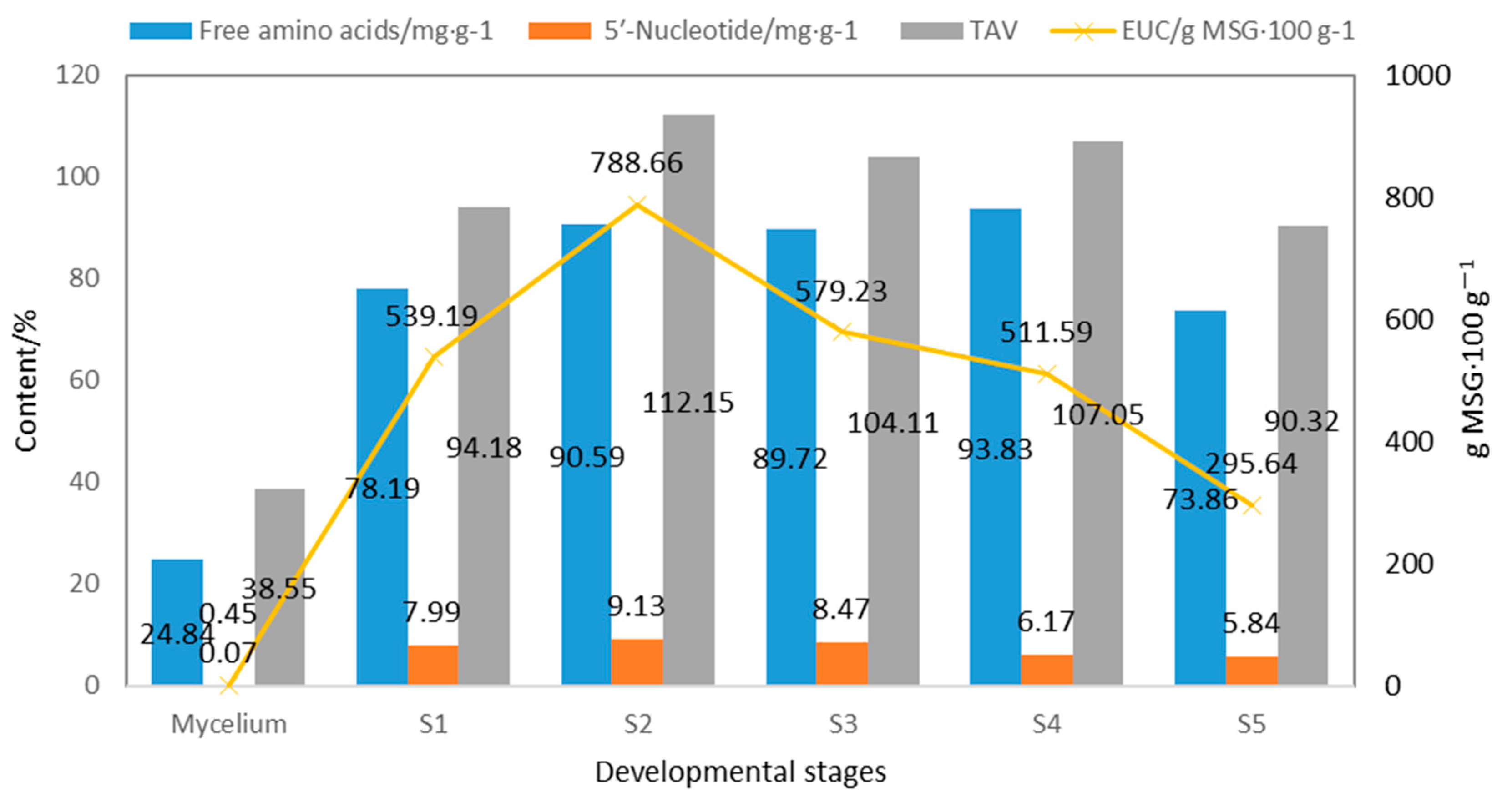
4.3.3. Taste Components of S. rugoso-annulata at Different Developmental Stages
4.3.4. Taste Components of S. rugoso-annulata with Different Processing Methods
4.3.5. Aroma Components of S. rugoso-annulata with Different Processing Methods
4.3.6. Flavor Components of S. rugoso-annulata at Different Temperatures
4.3.7. Comparison of Aroma Components between S. rugoso-annulata and Other Edible Mushrooms
5. Problems and Prospects
5.1. Zero Research Report on Chitin
5.2. High Protein Content Distortion
5.3. Little Research on the Bioactive Components in the Fermentation Mycelium and Liquid of S. rugoso-annulata
5.4. Genes Related to the Metabolism of Nutritional and Flavoring Components Have Not Been Reported
5.5. The Deep-Processed Products Are Almost Blank
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Diego, C.Z.; Pardo-Giménez, A. Edible and Medicinal Mushrooms: Technology and Applications; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2017. [Google Scholar] [CrossRef]
- Jing, B.-N.; Chang, X.; Wei, L.; Xie, X.-Y.; Zhou, Y.; Wang, Z.-Y.; Liu, Y.-Q.; Wang, W. Analysis and Evaluation of Nutrient Components, Bioactive Substances and Heavy Metal Content of Stropharia rugosoannulata in Bo’ai County. Sci. Technol. Food Ind. 2022, 43, 278–285. [Google Scholar] [CrossRef]
- Li, X.-X.; Zhang, Z.-Q.; Wang, L.; Zhao, H.-Q.; Jia, Y.-H.; Ma, X.; Li, J.-Z.; Wang, Y.; Ma, B.-J. Three-phase extraction of polysaccharide from Stropharia rugosoannulata: Process optimization, structural characterization and bioactivities. Front. Immunol. 2023, 13, 994706. [Google Scholar] [CrossRef]
- Wu, J.; Kobori, H.; Kawaide, M.; Suzuki, T.; Choi, J.; Yasuda, N.; Noguchi, K.; Matsumoto, T.; Hirai, H.; Kawagishi, H. Isolation of Bioactive Steroids from the Stropharia Rugosoannulata Mushroom and Absolute Configuration of Strophasterol. Biosci. Biotechnol. Biochem. 2013, 77, 1779–1781. [Google Scholar] [CrossRef]
- Li, W.; Chen, W.; Ma, H.; Wang, J.; Li, Z.; Wang, Q.; Zhang, Z.; Wu, D.; Zhang, J.; Yang, Y. Study on the Relationship Between Structure and Taste Activity of the Umami Peptide of Stropharia Rugosoannulata Prepared by Ultrasound. Ultrason. Sonochem. 2022, 90, 106206. [Google Scholar] [CrossRef]
- Bao, C.-L.-G.; Guan, C.-B.; Xin, M.-H.; Teng, X.; Liu, T.-T.; Wang, D.-W. To analyze the effects of roasting on volatile flavor compounds of Stropharia rugoso-annulate using HS-SPME-GC-MS and electronic nose. Food Sci. 2022, 43, 1–14. [Google Scholar]
- Li, X.-M.; Li, H.-J.; Xiao, X.; Liao, L.; Chen, R.; Xie, Z.-H.; He, Z.-F. Study on the change of volatile flavor substances during the processing of deep-fried and battered mushroom stalks of Stropharia rugoso-annulata. Food Ferment. Ind. 2022, 1–10. [Google Scholar] [CrossRef]
- Stamets, P.; Chilton, J.S. The Mushroom Cultivator-A Practical Guide to Growing Mushrooms at Home. 1983. Available online: http://library.uniteddiversity.coop/Permaculture/Mushroom_Cultivator-A_Practical_Guide_to_Growing_Mushrooms_at_Home.pdf (accessed on 8 October 2021).
- Domondon, D.; Poppe, J.; Griensven, L. Fruit optimization with wastes used for outdoor cultivation of king Stropharia. Neophilologus 2000, 79, 619–628. [Google Scholar]
- Yan, Q.-X.; Huang, M.-X.; Sun, P.; Cheng, S.-X.; Zhang, Q.; Dai, H. Steroids, fatty acids and ceramide from the mushroom Stropharia rugosoannulata Farlow apud Murrill. Biochem. Syst. Ecol. 2020, 88, 103963. [Google Scholar] [CrossRef]
- China Edible Fungi Association. Statistical Survey of Edible Fungi in China in 2021. Edible Fungi China 2023, 42, 118–127. [Google Scholar] [CrossRef]
- Chen, W.-C.; Li, W.; Wu, D.; Zhang, Z.; Chen, H.; Li, Z.-P.; Wang, C.-G.; Yang, Y. Multilevel analysis and evaluation of dried flavor quality of Stropharia rugoso-annulata based on component profile data. Food Sci. 2022, 44, 1–12. [Google Scholar]
- Li, W.; Feng, J.; Ma, H.-L.; Chen, W.-C.; Wu, D.; Zhang, Z.; Yang, Y. Analysis of the characteristic flavor components and flavor characteristics of the fermentation of Stropharia rugoso-annulata based on targeted metabolite assay. J. Food Saf. Qual. 2022, 13, 2736–2744. [Google Scholar] [CrossRef]
- Lu, Q.; Wang, S.; Xue, S.; Yang, D.; Li, L. Comparison of Non-volatile Flavor Compounds in Stropharia rugosoannulata Soup Processed by Different Methods. J. Food Sci. Technol. 2022, 59, 1. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Xue, S.-J.; Yang, D.; Wang, S.-H.; Li, L. Effects of Different Processing Methods on the Volatile Components and Antioxidant Ability of the Water Extracts from Stropharia rugosoannulata. Sci. Technol. Food Ind. 2021, 42, 41–48. [Google Scholar] [CrossRef]
- Huang, J.-X.; Yuan, S.-N.; Pan, J.; Zheng, D.-H.; Chen, J.-M.; Li, J.; Gui, Q.; Zhou, L.-J. Difference of Primary Nutritional Ingredient between Lentinus edodes, Pleurotus ostreatus and Stropharia rugosoannulata Grown Mainly with Rubber Wood Dust. Chin. J. Trop. Crops 2018, 39, 1625–1629. [Google Scholar] [CrossRef]
- Yan, Q.-X. Study on Screening of Active Components and Quality Standard of Stropharia Rugosoannulata. Master’s Thesis, Guangxi University of Traditional Chinese Medicine, Nanning, China, 2019. Chinese Master’s Theses Full-Text Database. Available online: https://kns.cnki.net/kcms/detail/detail.aspx?dbname=CMFD2022&filename=1021126383.nh&dbcode=CMFD (accessed on 9 October 2021).
- Li, S.-R.; Wang, L.; Ni, S.-J.; Wang, H.-H.; Liu, X.-F.; Liu, C.-X. The Amino Acids Content of Different Part of Stropharia rugoso-annulata and Their Nutrition Evaluation. Food Res. Dev. 2017, 38, 95–99. [Google Scholar] [CrossRef]
- Liu, M.-M.; Zhang, X.-L.; Xu, L.-L.; Jiang, P. Analysis of Amino Acids Content and Food Safety Assessment of Stropharia rugosoannulata Cultivated in the Imitated Wild Environment under Forest. Edible Fungi China 2021, 40, 67–70. [Google Scholar] [CrossRef]
- Wei, J.; Zhao, Q.-Q.; Wang, X.-L.; Zhang, G.-Q.; Cheng, J.-H.; Chen, Q.-J. Effects of Different Formulations and Cultivation Techniques on Nutrient Quality of Stropharia rugoso-annulata. Vegetables 2022, 8, 51–54. [Google Scholar]
- Wang, X.-W.; Zhan, W.; Tao, M.-X.; Ji, Z.-S.; Liu, Y.; Wang, F.; Cheng, G.-Y. Analysis of the nutritional and antioxidant components of Stropharia rugoso-annulata. Edible Fungi 2007, 171, 62–63. [Google Scholar]
- Zeng, J.-Y.; Zhao, B.; Bi, W.-Y.; Liu, C.-Y.; Zhang, G.-C. Effects of Na2SeO3 and ZnSO4 on Liquid Fermentation of Stropharia rugoso-annulata Mycelia. J. Jilin Agric. Univ. 2018, 40, 171–177. [Google Scholar] [CrossRef]
- Wang, D. Study on the Accumulation of Heavy Metal Lead and Cadmium and the Interaction Effect of Selenium Supplementation in the Stropharia rugosoannulata. Master’s Thesis, Anhui Agricultural University, Hefei, China, 2018. Available online: https://kns.cnki.net/kcms/detail/detail.aspx?dbname=CMFD201901&filename=1018318645.NH&dbcode=CMFD (accessed on 10 October 2021).
- Zhao, Y.; Ren, Y.-F.; Chen, M.-J.; Zha, L.; Song, X.-X.; Guo, L.-G.; Wang, C.-G. Analysis of fatty acid composition and content in three strains of Stropharia rugoso-annulata by GC-MS. Food Mach. 2018, 34, 65–67+72. [Google Scholar] [CrossRef]
- Li, Z.-P.; Yu, C.-X.; Ren, Y.-F.; Chen, M.-J.; Zha, L.; Yang, H.-L.; Song, X.-X.; Zhao, Y. Effect of Heat Stress on Fatty Acids in Stropharia rugosoannulata Mycelia. Acta Edulis Fungi 2020, 27, 45–50. [Google Scholar] [CrossRef]
- Lu, Q.; Xue, S.-J.; Yang, D.; Wang, S.-H.; Li, L. Effects of Three Extraction Methods on Antioxidant Properties of Crude Polysaccharides from Stropharia Rugoso. Food Sci. Technol. 2021, 46, 171–178. [Google Scholar] [CrossRef]
- Hu, S.; Feng, X.; Huang, W.; Ibrahim, S.A.; Liu, Y. Effects of Drying Methods on Non-volatile Taste Components of Stropharia rugoso-annulata Mushrooms. LWT 2020, 127, 109428. [Google Scholar] [CrossRef]
- Wang, Q.; Zhao, Y.; Feng, X.; Ibrahim, S.A.; Huang, W.; Liu, Y. Effects of Drying on the Structural Characteristics and Antioxidant Activities of Polysaccharides from Stropharia rugosoannulata. J. Food Sci. Technol. 2021, 58, 3622–3631. [Google Scholar] [CrossRef]
- Liu, L.-P.; Qian, W.-C.; Zhan, P.F.; Song, W.-M.; Wen, Y.; Wang, Q.-H. Effect of Different Substrates and Drying Methods on the Nutritional Composition of Stropharia rugosoannulata. J. Southwest Univ. Nat. Sci. Ed. 2018, 40, 8–13. [Google Scholar] [CrossRef]
- Yu, H.-P.; Hu, S.; Huang, W.; Wang, Y.; Liu, Y. Effects of Drying Process on the Tasty Components in Stropharia rugoso-annulata. Sci. Technol. Food Ind. 2021, 42, 251–256. [Google Scholar] [CrossRef]
- Qin, Y.-C.; Wu, D.-P.; Wang, L.-L.; Fang, R.; He, L.; Wang, Y.-B.; Qian, H.; Liu, B.-T. Effects of Various Drying Methods on Volatile Composition of Stropharia rugosoannulata by Headspace-Solid Phase Microextraction-Gas Chromatography-Mass Spectrometry. Food Sci. 2022, 43, 273–280. [Google Scholar] [CrossRef]
- Chen, R.-R.; Li, W.; Wu, D.; Mao, C.-F.; Bao, D.-P.; Chen, W.-C.; Zhang, J.-S.; Yang, Y. Nutrients, Texture and Taste Characteristics of Stropharia rugosoannulata During Growth and Development. Acta Edulis Fungi 2022, 29, 42–54. [Google Scholar] [CrossRef]
- Brodzinska, Z.; Lasota, W. Chemical composition of cultivated mushrooms Part I. Stropharia rugoso-annulata Farlow ex. Murr. Bromatol. I Chem. Toksykol. 1981, 14, 229–238. [Google Scholar]
- Jiang, H. Studies on the Degradation and Cultivation under Forest of Stropharia rugoso-annulata. Master’s Thesis, Xinjiang Agricultural University, Ürümqi, China, 2020. Chinese Master’s Theses Full-Text Database. Available online: https://kns.cnki.net/kcms/detail/detail.aspx?dbname=CMFDTEMP&filename=1022002887.nh&dbcode=CMFD (accessed on 11 October 2021).
- Yang, F.-T.; Long, S.-B.; Long, J.-Q.; Ma, X.-T.; Wang, K. A Culture Technique of Edible Fungi (Stropharia rugosoannulata Farl. ex Murrill)-paddy Rice Rotation in Guizhou Province. Tillage Cultiv. 2021, 41, 96–98. [Google Scholar] [CrossRef]
- Wang, L.; Ni, S.-J.; Li, S.-R.; Wang, H.-H.; Jiang, X.-X.; Liu, X.-F.; Pan, Y. Analysis of the Different Parts of Base Nutrition of Stropharia rugoso-annulata. Heilongjiang Agric. Sci. 2016, 269, 143–145. [Google Scholar]
- Wei, L.; Wang, W.; Xie, X.-Y.; Zhou, Y.; Liu, Y.-Q.; Ma, Y.-N.; Ning, E.-J.; Wang, T.; Li, N.-J.; Jing, B.-N. Optimization of Extraction Process of Polysaccharides from Stropharia rugosoannulata in Bo’ai County by Response Surface Method and Evalation of Their Antibacterial and Antioxidant Activity. Sci. Technol. Food Ind. 2023, 1–12. [Google Scholar] [CrossRef]
- Xie, X.-Y.; Chang, X.; Liu, Y.-Q.; Jing, B.-N.; Han, H.-Y.; Wei, L.; Zhou, Y.; Wang, W. Optimization of fermentation conditions and antioxidant activity of polysaccharides from fruiting body of Stropharia rugosoannulata. China Food Addit. 2022, 33, 55–61. [Google Scholar] [CrossRef]
- Jia, J.; Xie, X.-C.; Song, Y.; Sun, Y.; Xu, Y.-J.; Yao, S. Optimizing Culture Medium and Antioxidant Activity of Extracellular Polysaccharides in Liquid Fermentation of Stropharia rugosoannulata. North. Hortic. 2021, 495, 105–115. [Google Scholar]
- He, P.-X.; Geng, L.-J.; Wang, J.-Z.; Xu, C.-P. Production, purfication, molecular characterization and bioactivities of exopolysaccharides produced by the wine cap culinary-medicinal mushroom, Stropharia rugosoannulata 2# (higher Basidiomycetes). Int. J. Med. Mushrooms 2012, 14, 365–376. [Google Scholar]
- Jin, M.; Zhang, W.; Zhang, X.; Huang, Q.; Chen, H.; Ye, M. Characterization, Chemical Modification and Bioactivities of a Polysaccharide from Stropharia rugosoannulata. Process Biochem. 2023, 128, 30–39. [Google Scholar] [CrossRef]
- Jiang, L. The Research on Preparation, Structural Identification and Biological Activity of Polysaccharide from Stropharia rugosoannulata (SR-1) and Polysaccharide from Tricholoma lascivum (Fr.) Gillet (TLG-1). Master’s Thesis, China West Normal University, Nanchong, China, 2019. Available online: https://kns.cnki.net/kcms/detail/detail.aspx?dbname=CMFD202001&filename=1019625385.NH&dbcode=CMFD (accessed on 12 October 2021).
- Zhai, X.; Zhao, A.; Geng, L.; Xu, C. Fermentation Characteristics and Hypoglycemic Activity of an Exopolysaccharide Produced by Submerged Culture of Stropharia rugosoannulata #2. Ann. Microbiol. 2012, 63, 1013–1020. [Google Scholar]
- Li, X.; Cui, W.; Cui, Y.; Song, X.; Jia, L.; Zhang, J. Stropharia rugoso-annulata Acetylated Polysaccharides Alleviate Nafld Via Nrf2/jnk1/ampk Signaling Pathways. Int. J. Biol. Macromol. 2022, 215, 560–570. [Google Scholar] [CrossRef]
- Wang, X.-W. Nutrition Components Analyse, Extraction and Antioxidant Properties of Polysaccharide of Stropharia rugoso-annulata. Master’s Thesis, Nanjing Normal University, Nanjing, China, 2007. Available online: https://kns.cnki.net/kcms/detail/detail.aspx?dbname=CMFD2007&filename=2007107796.NH&dbcode=CMFD (accessed on 13 October 2021).
- Xiao, W.-F.; Chai, W.-G.; Qiu, J.-R.; Xin, Y.; Ruan, S.-L. Proteomic Analysis Reveals Mechanisms of Stropharia rugosoannulata Polysaccharides Promoting Kiwifruit Growth and High Temperature Resistance. J. Nucl. Agric. Sci. 2022, 36, 94–104. [Google Scholar] [CrossRef]
- Xiao, W.-F.; Ni, S.; Ruan, S.-L.; Chen, H.-Z.; Xin, Y.; Qiu, J.-R.; Chai, W.-G. Effects of Fruiting Body Polysaccharides from 9 Edible Fungi on Growth of Rice Seedling. Acta Edulis Fungi 2020, 27, 69–74. [Google Scholar] [CrossRef]
- Pozdnyakova, N.; Schlosser, D.; Dubrovskaya, E.; Balandina, S.; Sigida, E.; Grinev, V.; Turkovskaya, O. The Degradative Activity and Adaptation Potential of the Litter-decomposing Fungus Stropharia rugosoannulata. World J. Microbiol. Biotechnol. 2018, 34, 133. [Google Scholar] [CrossRef]
- Liu, Y.; Hu, C.; Feng, X.; Cheng, L.; Ibrahim, S.A.; Wang, C.; Huang, W. Isolation, Characterization and Antioxidant of Polysaccharides from Stropharia rugosoannulata. Int. J. Biol. Macromol. 2019, 155, 883–889. [Google Scholar] [CrossRef]
- Wang, M.-X.; Sheng, Z.-C.; Chen, X.-L. Optimization of Extraction and Characterization of Polysaccharide from Stropharia rugosoannulata Fermentation Mycelium. North. Hortic. 2020, 01, 111–116. [Google Scholar] [CrossRef]
- Qian, C.-Q.; Lin, W.-F.; Wu, D.-P.; Liu, N.-Y. Study on the Extraction of Polysaccharides from Stropharia rugoso-annulata by Alkaline Extraction and Its Scavenging Oxygen Free Radical. Mod. Food 2021, 03, 99–102. [Google Scholar] [CrossRef]
- Miao, Y.-Z. Separation and Purification of Se-Polysaccharide from Stropharia rugoso-annulata and Its Antioxidant Activities. Master’s Thesis, Shandong Agricultural University, Tai’an, China, 2009. Available online: https://kns.cnki.net/kcms/detail/detail.aspx?dbname=CMFD2010&filename=2009234859.NH&dbcode=CMFD (accessed on 14 October 2021).
- Chen, J.-C.; Weng, M.-J.; Lai, P.-F.; Li, Y.-B.; Zhou, X.-H.; Shen, H.-S. Distribution of Stropharia rugoso-annulata Polysaccharides Molecular Weight and Component Sugar. Sci. Agric. Sin. 2011, 44, 2109–2117. [Google Scholar]
- Cui, X.-R.; Wang, L.; Shi, F.-F.; Pan, Y.; Jia, H.-L.; Liu, X.-F.; Li, S.-R.; Song, H.-B. Extraction and antioxidant activity of Stropharia rugosoannulata protein. J. Food Saf. Qual. 2018, 9, 5949–5956. [Google Scholar]
- Huang, S. Study on the Extraction and Antioxidation of Polyphenol Compounds of Stropharia rugosoannulata Farlow. Master’s Thesis, Fujian Agriculture and Forestry University, Fuzhou, China, 2010. Chinese Master’s Thesis Full-Text Database. Available online: https://kns.cnki.net/kcms/detail/detail.aspx?dbname=CMFD2012&filename=1011038174.nh&dbcode=CMFD (accessed on 15 October 2021).
- Chen, C.; Huang, J.; Ding, L. Study on Ultrasonic Wave Assisted Extraction of Flavonoids from Stropharia. Food Res. Dev. 2015, 36, 116–119. [Google Scholar] [CrossRef]
- Wu, J.; Tokuyama, S.; Nagai, K.; Yasuda, N.; Noguchi, K.; Matsumoto, T.; Hirai, H.; Kawagishi, H. Strophasterols a to D with an Unprecedented Steroid Skeleton: From the Mushroom Stropharia rugosoannulata. Angew. Chem. Int. Ed. 2012, 51, 10820–10822. [Google Scholar] [CrossRef]
- Sato, S.; Taguchi, Y.; Kuwahara, S. Synthesis and Stereochemistry of Glaucoposterol a and Strophasterol D. Tetrahedron 2020, 76, 131129. [Google Scholar] [CrossRef]
- Zhang, W.; Tian, G.; Geng, X.; Zhao, Y.; Ng, T.; Zhao, L.; Wang, H. Isolation and Characterization of a Novel Lectin from the Edible Mushroom Stropharia rugosoannulata. Molecules 2014, 19, 19880–19891. [Google Scholar] [CrossRef]
- Li, W.; Chen, W.; Ma, H.; Wu, D.; Zhang, Z.; Yang, Y. Structural Characterization and Angiotensin-converting Enzyme (ace) Inhibitory Mechanism of Stropharia rugosoannulata Mushroom Peptides Prepared by Ultrasound. Ultrason. Sonochem. 2022, 88, 106074. [Google Scholar] [CrossRef] [PubMed]
- Lv, G.-Y.; Song, T.-T.; He, W.-Q.; Zhang, Y.-Y.; Cai, W.-M.; Zhang, Z.-F. Comparison of antioxidant activities and constituent analysis of different polar extracts from Stropharia rugosoannulata. Mycosystema 2022, 41, 999–1007. [Google Scholar]
- Lei, P.; Zhang, W.; Men, X. Review on Anti-tumor Effect of Triterpene Acid Compounds. J. Cancer Res. Ther. 2014, 10, 14. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Suzuki, T.; Kawagishi, H. An unusual sterol from the mushroom Stropharia rugosoannulata. Tetrahedron Lett. 2013, 54, 4900–4902. [Google Scholar] [CrossRef]
- Wu, J.; Fushimi, K.; Tokuyama, S.; Ohno, M.; Miwa, T.; Koyama, T.; Yazawa, K.; Nagai, K.; Matsumoto, T.; Hirai, H.; et al. Functional-food Constituents in the Fruiting Bodies of Stropharia rugosoannulata. Biosci. Biotechnol. Biochem. 2011, 75, 1631–1634. [Google Scholar] [CrossRef]
- Yan, Q.-X.; Luo, X.-L.; Li, C.-Q.; Zhou, Y.-X.; Qin, X.-M.; Qin, J.; Dai, H. Anti-fatigue Effect of Stropharia rugosoannulata Fruiting Body Extracts Derived from Different Solvents. Acta Edulis Fungi 2018, 25, 65–70. [Google Scholar] [CrossRef]
- Hu, D.-S.; Li, J.; Zhang, Y.; Zhong, D.-M.; Fang, X.-F.; Liao, Y.-Z.; Su, G.-G. Optimization of Extraction Conditions of Polyphenols from Stropharia rugosoannulata Cultivated under Forest. Edible Fungi China 2022, 41, 65–70. [Google Scholar] [CrossRef]
- Sun, L.; Zhang, Z.; Xin, G.; Sun, B.; Bao, X.; Wei, Y.; Zhao, X.; Xu, H. Advances in umami taste and aroma of edible mushrooms. Trends Food Sci. Technol. 2020, 96, 176–187. [Google Scholar] [CrossRef]
- Rao, K.-C.; Huang, W.; Wang, Y.; Liu, Y. Effects of Storage Temperature on the Postharvest Quality and Volatile Flavor Components of Stropharia rugoso-annulata. Sci. Technol. Food Ind. 2023, 44, 369–378. [Google Scholar] [CrossRef]
- Zhu, Y.; Chen, J.; Chen, X.; Chen, D.; Deng, S. Use of Relative Odor Activity Value (roav) to Link Aroma Profiles to Volatile Compounds: Application to Fresh and Dried Eel (Muraenesox cinereus). Int. J. Food Prop. 2020, 23, 2257–2270. [Google Scholar] [CrossRef]
- Li, J.-L.; Yang, Y.; Li, W.; Chen, W.-C.; Liu, X.-F. Aroma Change and Its Relationship with Key Enzymatic Reactions in Drying Process of Stropharia rugosoannulata. J. Food Sci. Technol. 2023, 41, 30–42. [Google Scholar] [CrossRef]
- Liu, Y.; Meng, F.; Tang, P.; Huang, D.; Li, Q.; Lin, M. Widely Targeted Metabolomics Analysis of the Changes to Key Non-volatile Taste Components in Stropharia rugosoannulata under Different Drying Methods. Front. Nutr. 2022, 9, 884400. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Shang, X.-D.; Wang, X.-Y.; Wang, C.-H.; Liu, J.-H.; Chen, W.-Z.; Wang, R.-J.; Xu, N. Comparison of Volatile Components in Different Parts of Fruiting Body of Stropharia rugoso-annulata. Mod. Food Sci. Technol. 2022, 38, 271–281+196. [Google Scholar] [CrossRef]
- Li, W.; Chen, W.-C.; Ma, H.-L.; Wu, D.; Zhang, Z.; Yang, Y. Ultrasonic Preparation of Stropharia rugosoannulata Peptides and Analysis of Their Taste Characteristics and Pharmacological Activities. Acta Edulis Fungi 2022, 29, 81–94. [Google Scholar] [CrossRef]
- Li, W.; Chen, W.-C.; Wu, D.; Zhang, Z.; Yang, Y. Taste peptides derived from Stropharia rugosoannulata fermentation mycelium and molecular docking to the taste receptor T1R1/T1R3. Front. Nutr. 2022, 9, 960218. [Google Scholar] [CrossRef]
- Chen, W.; Li, W.; Wu, D.; Zhang, Z.; Chen, H.; Zhang, J.; Wang, C.; Wu, T.; Yang, Y. Characterization of Novel Umami-active Peptides from Stropharia rugoso-annulata Mushroom and in Silico Study on Action Mechanism. J. Food Compos. Anal. 2022, 110, 104530. [Google Scholar] [CrossRef]
- Li, N.; Prescott, J.; Wu, Y.; Barzi, F.; Yu, X.; Zhao, L.; Neal, B. The Effects of a Reduced-sodium, High-potassium Salt Substitute on Food Taste and Acceptability in Rural Northern China. Br. J. Nutr. 2008, 101, 1088–1093. [Google Scholar] [CrossRef]
- Gu, Z.; Yang, Y. Research progress in flavor components of edible fungus. Sci. Technol. Food Ind. 2013, 34, 363–367. [Google Scholar] [CrossRef]
- Yin, C.-M.; Fan, X.-Z.; Shi, D.-F.; Fan, Z.; Cheng, W.; Gao, H. Flavor Compounds Analysis of 5 Fresh Mushrooms Using HS-SPME-GC-MS and HPLC. Sci. Technol. Food Ind. 2019, 40, 254–260. [Google Scholar] [CrossRef]
- Yu, C.-X.; Zhao, Y.; Chen, M.-J.; Wang, H.; Li, Z.-P.; Pan, G.-F.; Feng, A.-P. Analysis of volatile of flavor components in Volvariella volvacea fruiting bodies cultivated on different substrates. Acta Edulis Fungi 2019, 26, 37–44. [Google Scholar] [CrossRef]
- Anon. Biology of Sulfur; CRC Press: Boca Raton, FL, USA, 1996; pp. 30–51. [Google Scholar]
- Bowman, S.M.; Free, S.J. The Structure and Synthesis of the Fungal Cell Wall. Bioessays 2006, 28, 799–808. [Google Scholar] [CrossRef] [PubMed]
- Crini, G. Chitin and Chitosan; Elsevier: Amsterdam, The Netherlands, 2022. [Google Scholar]
- Deguchi, S.; Tsujii, K.; Horikoshi, K. In Situ Microscopic Observation of Chitin and Fungal Cells with Chitinous Cell Walls in Hydrothermal Conditions. Sci. Rep. 2015, 5, 11907. [Google Scholar] [CrossRef] [PubMed]
- Alimi, B.A.; Pathania, S.; Wilson, J.; Duffy, B.; Frias, J.M.C. Extraction, Quantification, Characterization, and Application in Food Packaging of Chitin and Chitosan from Mushrooms: A Review. Int. J. Biol. Macromol. 2023, 237, 124195. [Google Scholar] [CrossRef] [PubMed]
- Jedidi, I.K.; Ayoub, I.K.; Philippe, T.; Bouzouita, N. Chemical Composition and Nutritional Value of Three Tunisian Wild Edible Mushrooms; Springer Science and Business Media LLC: New York, NY, USA, 2017; Volume 11, pp. 2069–2075. [Google Scholar]
- DD ISO/TS 17837: 2008; Milk and Milk Products. Determination of Nitrogen Content and Crude Protein Calculation. Kjeldahl Method. BSI British Standards: London, UK, 2018.
- Cheung, P.C.K. The Nutritional and Health Benefits of Mushrooms. Nutr. Bull. 2010, 35, 292–299. [Google Scholar] [CrossRef]
- Elisashvili, V. Submerged Cultivation of M edicinal Mushrooms: Bioprocesses and Products (review). Int. J. Med. Mushrooms 2012, 14, 211–239. [Google Scholar] [CrossRef]
- Li, Y.-R.; Chen, Z.-L.; Wen, L.-H.; Sun, C.-Q.; Li, R.-J.; Lin, M.; Chen, X.-J.; Zhang, C.-R.; Meng, F.-B.; Huang, D.-M.; et al. Pollution-free cultivation technology of Stropharia rugosoannulata mushrooms in Guizhou. Agro-Tech. Serv. 2020, 37, 52–54+56. [Google Scholar]





| Parts of S. rugoso-annulata | Contents of Nutritional Components | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Crude Protein (%) | Total Amino Acid (%) | Ash (%) | Crude Fat (%) | Total Carbohydrate (%) | Crude Fiber (%) | Mn (mg·kg−1) | Zn (mg·kg−1) | Ca (mg·kg−1) | Mg (mg·kg−1) | |
| Pileus | 26.07–34.60 * | 17.51–28.19 * | 9.95–11.36 * | 2.43–3.83 * | 46.24–49.10 | 5.05–6.97 | 7.3 * | 91.4 * | 183.3 | 688.7 |
| Stipe | 12.49–22.60 | 6.68–20.40 | 7.71–8.88 | 1.31–2.16 | 58.91–60.80 * | 5.48–11.26 * | 4.3 | 35.2 | 218.8 * | 1211.3 * |
| Source | Monosaccharide Components/Molar Ratio | Structure Characteristic | Molecular Weight | Optimal Extraction Conditions | Yield | References |
|---|---|---|---|---|---|---|
| Fruiting body | Glucose, galactose, fucose (61.48:35.19:3.33) | The main chain was composed of [→3)-Glcp-(1→] and [→3,6)-Glcp-(1→], and the branch chain was composed of [→6)-Galp-(1→], T-Glcp and a small amount of T-Fucp exists at the end | 22.907 kDa | 80 °C, 3 h, 1:30 (w/v) | 3.46% | [41] |
| Fruiting body | D-fructose, D-glucose, D-xylose, two unidentified monosaccharides | Mainly pyranose conformation, with both α- and β-glycosidic bonds | 22 kDa | 73.8 °C, 22.7 min, 1:26.2 (w/v) | 7.64% | [53] |
| Fruiting body | Galactose, glucose (3:1) | A skeleton structure of (1,6)-α-D-Galp with branches at 2-O and the branches consisting of a (1→6,4)-β-D-Glcp and two →2)-α-D-Glcp | 13.28 kDa | 100 °C, 6 h, 1:20 (w/v) | —— | [42] |
| Fruiting body | SRP-1 (neutral polysaccharides) consist of galactose, glucose, and mannose (4.11:56.26:1); SRP-2 (acidic polysaccharides) consist of galactose, glucose, glucuronic acid, mannose, and ribose (1.41:23.24:1.22:1:3.03) | Both neutral polysaccharides and acidic polysaccharides have a (1→6)-α-D-glucan backbone; neutral polysaccharides have both α-type and β-type glycosidic linkages whereas acidic polysaccharides have a dominant β-type constitution | —— | Room temperature, microwave 640 W, 2 min, 1:50 (w/v) | —— | [49] |
| Fruiting body | The same four types of monosaccharides: mannose, glucose, galactose, and arabinose; FSRP–1 (2.46:25.39:7.70:1); HSRP–1 (1.53:31.02:4.34:1); FSRP–2 (3.55:19.19:6.20:1); HSRP–2 (0.37:13.79:5.63:1) | Neutral polysaccharides FSRP-1 and HSRP-1 have the same (1→6)-α-D-Galp as the backbone chain, acidic polysaccharides FSRP-2 and HSRP-2 have the same (1→3)-β-D-Glcp as the backbone chain. FSRP-1 contains two types of side chains, (1→6)-β-D-Glcp and (1→3)-β-D-Glcp, HSRP-1 only contains the (1→3)-β-D-Glcp side chain. Both FSRP-2 and HSRP-2 contain the same (1→4)-α-D-Galp side chain | —— | room temperature, microwave 640 W, 2 min, 1:50 (w/v) | —— | [28] |
| Fruiting body | —— | Pyranose conformation | 2456–8992 kDa | 100 °C, 2 h, 1:30 (w/v) | 6.22–10.7% | [26] |
| Fruiting body | Glucose, galactose, glucuronic acid, fructose, xylose, fucose, arabinose, rhamnose (35.79%:26.80%:9.92%:8.65%:7.92%:4.19%:3.46%:3.26%) | —— | 27.52 kDa | 100 °C, 2 h, 1:30 (w/v) | 6.85% | [3] |
| Mycelium (intracellular) | Mannose, ribose, rhamnose, glucose, xylose, arabinose, fructose (0.76:1.64:0.65:1.00:1.24:1.18:0.20) | Pyranose conformation, β-glycosidic bonds | —— | 82 °C, 3.5 h, 1:26 (w/v) | 15.12% | [50] |
| Mycelium (extracellular) | Mannose, glucose, galactose (44.4%:7.7%:1.2%) | —— | —— | —— | —— | [48] |
| Mycelium (extracellular) | —— | β-glycosidic bonds | 5.305 kDa | (Liquid medium per liter) 60.0 g sucrose, 6.0 g tryptone, 5 mM KH2PO4, and initial pH 7.0, 28 °C | 9.967 g/L | [40] |
| Samples | Processing Methods | Flavor Components | References | |
|---|---|---|---|---|
| Fresh fruiting body | —— | Non-volatile | Volatile | [12] |
| Dried fruiting body | HAD (50 °C), VFD, MWD, ND (20 ± 4 °C) | Non-volatile | Volatile | [12] |
| Fresh fruiting body | Post-harvest storage (≤1 h, 0 °C) | —— | Volatile | [31] |
| Fresh pileus and stipe | Post-harvest storage (≤1 h, 0 °C) | —— | Volatile | [31] |
| Dried fruiting body | HAD (30 °C, 50 °C, 70 °C, 90 °C), VFD | —— | Volatile | [31] |
| Fruiting body from fresh to dried | HAD (55 °C, 0–12 h) | —— | Volatile | [70] |
| Dried fruiting body at different developmental stages | HAD (50 °C) | Non-volatile | —— | [32] |
| Dried fruiting body | HAD (50 °C), VFD, MWD | Non-volatile | —— | [30] |
| Dried pileus and stipe | NAD (20–30 °C), HAD (50 °C), VFD | Non-volatile | —— | [27] |
| Fresh pileus and stipe | —— | Non-volatile | —— | [71] |
| Dried pileus and stipe | HAD (55 °C), VFD, MVD | Non-volatile | —— | [71] |
| Fresh fruiting body | Post-harvest storage (0 °C, 15 °C, 12 d) | —— | Volatile | [68] |
| Fresh stipe | Battered and deep-fried (180 °C) | —— | Volatile | [7] |
| Dried pileus and stipe | HAD and then VFD | —— | Volatile | [72] |
| Dried fruiting body | Roasted (100 °C,120 °C,140 °C,160 °C, 180 °C, 5 min) | —— | Volatile | [6] |
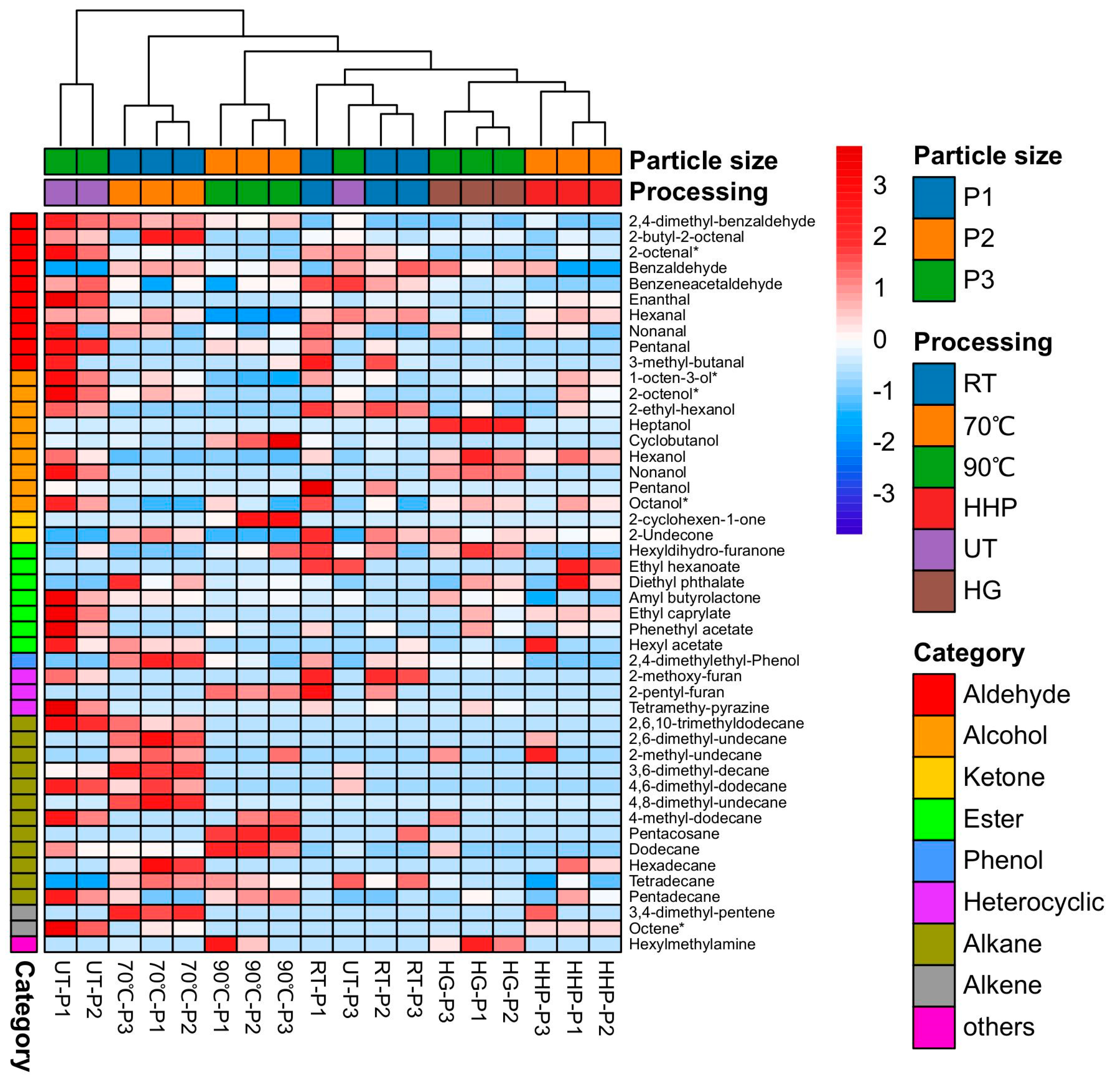
| Soup (water extracts of fruiting body powder) | Thermal treatment (70 °C, 90 °C), non-thermal treatment (RT, UT, HG, HHP) | —— | Volatile | [15] |
| Soup (water extracts of fruiting body powder) | Thermal treatment (70 °C, 90 °C), non-thermal treatment (RT, UT, HG, HHP) | Non-volatile | —— | [14] |
| Fermentation mycelium | VFD | Non-volatile | —— | [13] |
| Extracellular fluid of fermentation mycelium | —— | Non-volatile | Volatile | [13] |
| Samples | Number | Components | Taste Characteristics | Components and Taste Threshold/mmol·L−1 | References |
|---|---|---|---|---|---|
| Fruiting body | 5 | Pentapeptide to heptapeptide | Umami | EPLCNQ (0.178), SGCVNEL (0.174), PHEMQ (0.390), SEPSHF (0.356), ESCAPQL (0.167) | [75] |
| Fruiting body | 47 | Octapeptide to undecapeptide | Umami | GQEDYDRLRPL (0.117), VVVGTPGRVF (0.403), ESPERPFL (0.254), HLYHPVPIEE (0.640) | [5] |
| Fermented mycelium | 748 | Heptapeptide to tetracosapeptide | Umami, salty | —— | [74] |
| Samples | Components | Total Content | Component of Highest Content | References |
|---|---|---|---|---|
| Dried fruiting body (HAD, VFD, MWD, ND) | Trehalose, glucose, mannose, fructose, mannitol, erythritol, arabitol | 10.01–13.06% | Trehalose (7.82–11.03%) | [12] |
| Dried pileus (HAD, VFD, NAD) | Arabinose, trehalose, glucose | 5.56–26.89% | Arabinose (5.46–26.54%) | [27] |
| Dried stipe (HAD, VFD, NAD) | Arabinose, trehalose, glucose | 11.94–33.19% | Arabinose (11.90–32.94%) | [27] |
| Soup (water extracts of fruiting body powder) | Glucose, fucose, xylose, mannose, fructose, ribose | 0.37–3.05% | Glucose (0.20–2.78%) | [14] |
| Fermentation mycelium (VFD) | Glucose, glucuronic acid, rhamnose, galactose, ribose | 8.32–8.65% | Glucose (7.92–8.25%) | [13] |
| Extracellular liquid of fermentation mycelium | Glucose, glucuronic acid, rhamnose, galactose, ribose, arabinose, fucose | 0.38–1.06 mg/mL | Glucose (0.34–1.02 mg/mL) | [13] |
| Samples | Components | Total Content | Component of Highest Content | References |
|---|---|---|---|---|
| Dried fruiting body (HAD) | Glu, Asp, Thr, Ser, Pro, Gly, Ala, Phe, Arg, His, Val, Met, Ile, Leu, Trp, Lys, Tyr, Cys, Gln, Asn | 7.39–9.38% | Glu (0.64–0.99%) | [32] |
| Dried fruiting body (HAD, VFD, MWD, ND) | Glu, Asp, Thr, Ser, Pro, Gly, Ala, Phe, Arg, His, Val, Met, Ile, Leu, Trp, Lys, Tyr, Cys | 3.07–4.61% | Glu (0.47–0.77%) | [12] |
| Dried fruiting body (HAD, VFD, MWD) | Glu, Asp, Thr, Ser, Pro, Gly, Ala, Phe, Arg, His, Val, Met, Ile, Leu, Trp, Lys, Tyr, Cys, GABA, Orn, Tau | 6.46–7.36% | Thr (1.55–1.90%) | [30] |
| Dried pileus (HAD, VFD, NAD) | Glu, Asp, Thr, Ser, Pro, Gly, Ala, Phe, Arg, His, Val, Met, Ile, Leu, Trp, Lys, Tyr, Cys, GABA, Orn | 3.72–5.11% | Thr (0.73–1.94%) | [27] |
| Dried stipe (HAD, VFD, NAD) | Glu, Asp, Thr, Ser, Pro, Gly, Ala, Phe, Arg, His, Val, Met, Ile, Leu, Trp, Lys, Tyr, Cys, GABA, Orn | 2.48–4.51% | Thr (0.67–0.89%) | [27] |
| Soup (water extracts of fruiting body powder) | Glu, Asp, Thr, Ser, Gly, Ala, Phe, His, Val, Ile, Leu, Trp, Lys, Tyr, Cys | 4.10–12.36% | Leu (1.19–2.15%) | [14] |
| Fermented mycelium (VFD) | Glu, Asp, Thr, Ser, Pro, Gly, Ala, Phe, Arg, His, Val, Met, Ile, Leu, Trp, Lys, Tyr, Cys, Gln, Asn | 2.27–2.48% | Arg (0.51–0.52%) | [13] |
| Extracellular liquid of fermented mycelium | Glu, Asp, Thr, Ser, Pro, Gly, Ala, Phe, Arg, His, Val, Met, Ile, Leu, Lys, Tyr, Cys, Gln, Asn | 20.85–25.58 μg/mL | Ser (5.79–6.27 μg/mL) | [13] |
| Samples | Components | Total Content | Component of Highest Content | References |
|---|---|---|---|---|
| Dried fruiting body (HAD, VFD, MWD, ND) | G, I, A, X | 0.47–0.63% | X (0.32%, HAD) I (0.13–0.26%, VFD, MWD, ND) | [12] |
| Dried fruiting body (HAD) | C, U, G, I, A, X | 0.58–0.91% | C (0.16–0.24%) | [32] |
| Dried fruiting body (HAD, VFD, MWD) | C, U, G, I, A, X | 0.88–1.07% | C (0.45–0.57%) | [30] |
| Dried pileus (HAD, VFD, NAD) | C, U, G, I, A | 1.83–2.28% | C (0.96–1.32%) | [27] |
| Dried stipe (HAD, VFD, NAD) | C, U, G, I, A | 0.82–1.19% | C (0.44–0.68%) | [27] |
| Soup (water extracts of fruiting body powder) | C, U, G, I, A, X | 1.46–1.82% | A (0.16–1.45%) | [14] |
| Fermented mycelium (VFD) | C, U, G, I | 0.006–0.0067% | U (0.0023–0.0026%) | [13] |
| Extracellular liquid of fermented mycelium | C, U, G, I, A | 14.82 μg/mL–17.49 μg/mL | U (13.22–15.69 μg/mL) | [13] |
| Samples | Components | Content | Component of Highest Content | References |
|---|---|---|---|---|
| Dried fruiting body (HAD, VFD, MWD) | Malic acid, tartaric acid, ascorbic acid, acetic acid, fumaric acid, succinic acid, citric acid | 17.68–24.14% | Malic acid (9.2–16.77%) | [30] |
| Dried pileus (HAD, VFD, NAD) | Malic acid, tartaric acid, ascorbic acid, acetic acid, fumaric acid, succinic acid | 14.41–16.94% | Malic acid (5.17–5.96%) | [27] |
| Dried stipe (HAD, VFD, NAD) | Malic acid, tartaric acid, ascorbic acid, acetic acid, fumaric acid, succinic acid | 11.10–16.26% | Malic acid (3.77–7.82%) | [27] |
| Soup (water extracts of fruiting body powder) | Citric acid, malic acid, tartaric acid, ascorbic acid, acetic acid, fumaric acid, succinic acid | 4.03–9.84% | Citric acid (2.05–5.73%) | [14] |
| Fermentation mycelium (VFD) | Acetic acid, succinic acid, citric acid | 0.55–0.61% | Acetic acid (0.42%) | [13] |
| Extracellular liquid of fermentation mycelium | Acetic acid, succinic acid, citric acid, malic acid | 68.96–604.89 μg/mL | Acetic acid (14.43–495.13 μg/mL) | [13] |
| Parts of S. rugoso-annulata | Contents of Taste Components | ||||
|---|---|---|---|---|---|
| Soluble Sugars (%) | Free Amino Acids (%) | Organic Acids (%) | 5′-Nucleotides (%) | EUC (g MSG·100 g−1) | |
| Pileus | 5.56–26.89 | 3.72–5.11 * | 14.41–16.94 * | 1.83–2.28 * | 219.37–784.05 * |
| Stipe | 11.94–33.19 * | 2.48–4.51 | 11.10–16.26 | 0.82–1.19 | 76.83–128.07 |
| Mushrooms | Common Names | Main Aroma Components | Characteristic Aroma Components | References |
|---|---|---|---|---|
| S. rugoso-annulata | Giant Stropharia, winecap mushroom | Octanoids, aldehydes, alcohols, ketones | Isobutyraldehyde, hexanal, 1-octen-3-ol, 1-octen-3-one, and 3-octanone | [6,12,70] |
| L. edodes | Shiitake | Sulfur compounds, octanoids, alcohols, ketones | Dimethyl disulfide, dimethyl trisulfide, methanethiol, 1-octen-3-one | [78] |
| V. volvacea | Straw mushroom | Octanoids, aldehydes, ketones | Isobutyraldehyde, hexanal, 1-octen-3-ol, methanethiol, 2-pentylfuran, dimethyl sulfide | [79] |
| Agaricus bisporus | Button mushroom, white mushroom | Octanoids, alcohols, ketones | 1-octen-3-one, 3-octanone, 3-octanol | [78] |
| Pleurotus eryngii | King oyster mushroom | Octanoids, alcohols, ketones | 1-octen-3-one, 1-octen-3-ol | [78] |
| Pleurotus ostreatus | Oyster mushroom | Octanoids, alcohols, ketones | 1-octen-3-one, 1-octen-3-ol, 3-octanone | [78] |
| Flammulina velutipes | Golden needle mushrooms | Octanoids, alcohols, ketones | 1-octen-3-one, 3-octanone, 3-octanol | [78] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, L.; He, C.; Si, C.; Shi, H.; Duan, J. Nutritional, Bioactive, and Flavor Components of Giant Stropharia (Stropharia rugoso-annulata): A Review. J. Fungi 2023, 9, 792. https://doi.org/10.3390/jof9080792
Huang L, He C, Si C, Shi H, Duan J. Nutritional, Bioactive, and Flavor Components of Giant Stropharia (Stropharia rugoso-annulata): A Review. Journal of Fungi. 2023; 9(8):792. https://doi.org/10.3390/jof9080792
Chicago/Turabian StyleHuang, Lei, Chunmei He, Can Si, Hongyu Shi, and Jun Duan. 2023. "Nutritional, Bioactive, and Flavor Components of Giant Stropharia (Stropharia rugoso-annulata): A Review" Journal of Fungi 9, no. 8: 792. https://doi.org/10.3390/jof9080792
APA StyleHuang, L., He, C., Si, C., Shi, H., & Duan, J. (2023). Nutritional, Bioactive, and Flavor Components of Giant Stropharia (Stropharia rugoso-annulata): A Review. Journal of Fungi, 9(8), 792. https://doi.org/10.3390/jof9080792