
Fungal Ligninolytic Enzymes and Their Application in Biomass Lignin Pretreatment


Abstract
1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lignocellulosic Materials | Cellulose (%) | Hemicellulose (%) | Lignin (%) |
|---|---|---|---|
| Natural biomass sources | |||
| Hardwood | 40–55 | 24–40 | 18–25 |
| Oak | 43.2 | 21.9 | 35.4 |
| Pine | 45.6 | 24 | 26.8 |
| Natural hay | 44.9 | 31.4 | 12 |
| Leaves | 15–20 | 80–85 | 0 |
| Reed | 34–36 | 26–27 | 21 |
| Switchgrass | 31–45 | 20–31 | 12–18 |
| Agricultural biomass by-products or residues | |||
| Barley straw | 31–45 | 27–38 | 14–19 |
| Wheat straw | 33–38 | 26–32 | 17–19 |
| Rye straw | 33–35 | 27–30 | 16–19 |
| Oat straw | 31–37 | 27–38 | 16–19 |
| Silage | 39.27 | 25.96 | 9.02 |
| Hemp | 53.86 | 10.6 | 8.76 |
| Rapeseed | 20–35 | 15–22 | 15–23 |
| Industrial residues and waste | |||
| Willow sawdust | 35.6 | 21.5 | 28.7 |
| Paper | 85–99 | 0 | 0–15 |
| Newspaper | 40–55 | 25–40 | 18–30 |
| Waste papers from chemical pulps | 60–70 | 10–20 | 5–10 |
| Primary wastewater solids | 8–15 | NA | 24–29 |
| Sugarcane bagasse | 44 | 28 | 21 |
2. Materials and Methods
2.1. Microorganisms
2.2. Media Conditions and Screening of Ligninolytic Enzymes
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Correction Statement
References
- Moshood, T.D.; Nawanir, G.; Mahmud, F. Microalgae biofuels production: A systematic review on socioeconomic prospects of microalgae biofuels and policy implications. Environ. Chall. 2021, 5, 100207. [Google Scholar] [CrossRef]
- Tsita, K.G.; Pilavachi, P.A. Evaluation of next generation biomass derived fuels for the transport sector. Energy Policy 2013, 62, 443–455. [Google Scholar] [CrossRef]
- Statistical Review of World Energy 2022. BP p.l.c. Available online: https://www.bp.com/en/global/corporate/energy-economics/statistical-review-of-world-energy.html (accessed on 3 February 2023).
- European Commission. 2050 Long-Term Strategy. 2020. Available online: https://climate.ec.europa.eu/eu-action/climate-strategies-targets/2050-long-term-strategy_en (accessed on 3 February 2023).
- Internation Energy Agency. Statistics Report Key World Energy Statistics 2021; Internation Energy Agency: Paris, France, 2021. [Google Scholar]
- Atiwesh, G.; Parrish, C.C.; Banoub, J.; Le, T.T. Lignin degradation by microorganisms: A review. Biotechnol. Prog. 2022, 38, e3226. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Lü, X. Chapter 5—Composition of plant biomass and its impact on pretreatment. In Advances in 2nd Generation of Bioethanol Production; Woodhead Publishing Series in Energy; Lü, X., Ed.; Woodhead Publishing: Cambridge, UK, 2021; pp. 71–85. [Google Scholar] [CrossRef]
- Kirk, T. Lignin Biodegradation: Microbiology, Chemistry, and Potential Applications; CRC Press: Boca Raton, FL, USA, 2018. [Google Scholar] [CrossRef]
- Hatakeyama, H.; Hatakeyama, T. Lignin Structure, Properties, and Applications. Biopolymers 2009, 232, 1–63. [Google Scholar]
- Sjulander, N.; Kikas, T. Two-Step Pretreatment of Lignocellulosic Biomass for High-Sugar Recovery from the Structural Plant Polymers Cellulose and Hemicellulose. Energies 2022, 15, 8898. [Google Scholar] [CrossRef]
- Ruiz-Dueñas, F.J.; Martínez, Á.T. Microbial degradation of lignin: How a bulky recalcitrant polymer is efficiently recycled in nature and how we can take advantage of this. Microb. Biotechnol. 2009, 2, 164–177. [Google Scholar] [CrossRef]
- Kumar, A.; Chandra, R. Ligninolytic enzymes and its mechanisms for degradation of lignocellulosic waste in environment. Heliyon 2020, 6, e03170. [Google Scholar] [CrossRef]
- Bajpai, P. (Ed.) Chapter 3—Sources of lignocellulosic biomass. In Lignocellulosic Biomass in Biotechnology; Elsevier: Amsterdam, The Netherlands, 2022; pp. 25–36. [Google Scholar] [CrossRef]
- Ferdeș, M.; Dincă, M.N.; Moiceanu, G.; Zăbavă, B.Ș.; Paraschiv, G. Microorganisms and Enzymes Used in the Biological Pretreatment of the Substrate to Enhance Biogas Production: A Review. Sustainability 2020, 12, 7205. [Google Scholar] [CrossRef]
- Rezania, S.; Din, M.F.; Mohamad, S.E.; Sohaili, J.; Taib, S.M.; Yusof, M.B.M.; Kamyab, H.; Darajeh, N.; Ahsan, A. Review on pretreatment methods and ethanol production from cellulosic water hyacinth. Bioresources 2017, 12, 2108–2124. [Google Scholar] [CrossRef]
- Tayyab, M. Bioethanol production from lignocellulosic biomass by enviornment-friendly pretreatment methods: A review. Appl. Ecol. Environ. Res. 2018, 16, 225–249. [Google Scholar] [CrossRef]
- Madadi, M.; Penga, C.; Abbas, A. Advances in Genetic Manipulation of Lignocellulose to Reduce Biomass Recalcitrance and Enhance Biofuel Production in Bioenergy Crops. J. Plant Biochem. Physiol. 2017, 5, 182. [Google Scholar] [CrossRef]
- Ajala, E.O.; Ighalo, J.O.; Ajala, M.A.; Adeniyi, A.G.; Ayanshola, A.M. Sugarcane bagasse: A biomass sufficiently applied for improving global energy, environment and economic sustainability. Bioresour. Bioprocess. 2021, 8, 87. [Google Scholar] [CrossRef]
- Gilbertson, R.L. Wood-Rotting Fungi of North America. Mycologia 1980, 72, 1–49. [Google Scholar] [CrossRef]
- Blanchette, R.A. Degradation of the lignocellulose complex in wood. Can. J. Bot. 1995, 73, 999–1010. [Google Scholar] [CrossRef]
- Otjen, L.; Blanchette, R.; Effland, M.; Leatham, G. Assessment of 30 White Rot Basidiomycetes for Selective Lignin Degradation. Holzforschung 1987, 41, 343–349. [Google Scholar] [CrossRef]
- Sigoillot, J.-C.; Berrin, J.-G.; Bey, M.; Lesage-Meessen, L.; Levasseur, A.; Lomascolo, A.; Record, E.; Uzan-Boukhris, E. Fungal Strategies for Lignin Degradation. Adv. Bot. Res. 2012, 61, 263–308. [Google Scholar] [CrossRef]
- Shi, J.; Chinn, M.S.; Sharmashivappa, R. Microbial pretreatment of cotton stalks by solid state cultivation of Phanerochaete chrysosporium. Bioresour. Technol. 2008, 99, 6556–6564. [Google Scholar] [CrossRef]
- Dionisi, D.; Anderson, J.A.; Aulenta, F.; McCue, A.; Paton, G. The potential of microbial processes for lignocellulosic biomass conversion to ethanol: A review. J. Chem. Technol. Biotechnol. 2015, 90, 366–383. [Google Scholar] [CrossRef]
- Odier, E.; Odier, E.; Janin, G.; Janin, G.; Monties, B.; Monties, B.; Odier, E.; Odier, E.; Janin, G.; Janin, G.; et al. Poplar Lignin Decomposition by Gram-Negative Aerobic Bacteria. Appl. Environ. Microbiol. 1981, 41, 337–341. [Google Scholar] [CrossRef]
- Giroux, H.; Vidal, P.; Bouchard, J.; Lamy, F. Degradation of Kraft Indulin Lignin by Streptomyces viridosporus and Streptomyces badius. Appl. Environ. Microbiol. 1988, 54, 3064–3070. [Google Scholar] [CrossRef]
- Zimmermann, W.; Broda, P. Utilization of lignocellulose from barley straw by actinomycetes. Appl. Microbiol. Biotechnol. 1989, 30, 103–109. [Google Scholar] [CrossRef]
- Kerem, Z.; Friesem, D.; Hadar, Y. Lignocellulose Degradation during Solid-State Fermentation: Pleurotus ostreatus versus Phanerochaete chrysosporium. Appl. Environ. Microbiol. 1992, 58, 1121–1127. [Google Scholar] [CrossRef] [PubMed]
- Bari, E.; Taghiyari, H.R.; Naji, H.R.; Schmidt, O.; Ohno, K.M.; Clausen, C.A.; Bakar, E.S. Assessing the destructive behaviors of two white-rot fungi on beech wood. Int. Biodeterior. Biodegrad. 2016, 114, 129–140. [Google Scholar] [CrossRef]
- Suhara, H.; Kodama, S.; Kamei, I.; Maekawa, N.; Meguro, S. Screening of selective lignin-degrading basidiomycetes and biological pretreatment for enzymatic hydrolysis of bamboo culms. Int. Biodeterior. Biodegrad. 2012, 75, 176–180. [Google Scholar] [CrossRef]
- Shirkavand, E.; Baroutian, S.; Gapes, D.J.; Young, B.R. Pretreatment of radiata pine using two white rot fungal strains Stereum hirsutum and Trametes versicolor. Energy Convers. Manag. 2017, 142, 13–19. [Google Scholar] [CrossRef]
- Du, W.; Yu, H.; Song, L.; Zhang, J.; Weng, C.; Ma, F.; Zhang, X. The promoting effect of byproducts from Irpex lacteus on subsequent enzymatic hydrolysis of bio-pretreated cornstalks. Biotechnol. Biofuels 2011, 4, 37. [Google Scholar] [CrossRef] [PubMed]
- Kamei, I.; Hirota, Y.; Meguro, S. Integrated delignification and simultaneous saccharification and fermentation of hard wood by a white-rot fungus, Phlebia sp. MG-60. Bioresour. Technol. 2012, 126, 137–141. [Google Scholar] [CrossRef]
- Lee, J.-W.; Gwak, K.-S.; Park, J.-Y.; Park, M.-J.; Choi, D.-H.; Kwon, M.; Choi, I.-G. Biological pretreatment of softwood Pinus densiflora by three white rot fungi. J. Microbiol. 2007, 45, 485–491. [Google Scholar]
- Öhgren, K.; Bura, R.; Lesnicki, G.; Saddler, J.; Zacchi, G. A comparison between simultaneous saccharification and fermentation and separate hydrolysis and fermentation using steam-pretreated corn stover. Process. Biochem. 2007, 42, 834–839. [Google Scholar] [CrossRef]
- Liu, X.; Hiligsmann, S.; Gourdon, R.; Bayard, R. Anaerobic digestion of lignocellulosic biomasses pretreated with Ceriporiopsis subvermispora. J. Environ. Manag. 2017, 193, 154–162. [Google Scholar] [CrossRef]
- Janusz, G.; Pawlik, A.; Sulej, J.; Świderska-Burek, U.; Jarosz-Wilkołazka, A.; Paszczyński, A. Lignin degradation: Microorganisms, enzymes involved, genomes analysis and evolution. FEMS Microbiol. Rev. 2017, 41, 941–962. [Google Scholar] [CrossRef]
- Wong, D.W.S. Structure and Action Mechanism of Ligninolytic Enzymes. Appl. Biochem. Biotechnol. 2009, 157, 174–209. [Google Scholar] [CrossRef]
- Weng, C.; Peng, X.; Han, Y. Depolymerization and conversion of lignin to value-added bioproducts by microbial and enzymatic catalysis. Biotechnol. Biofuels 2021, 14, 84. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Fueyo, E.; Linde, D.; Almendral, D.; López-Lucendo, M.F.; Ruiz-Dueñas, F.J.; Martínez, A.T. Description of the first fungal dye-decolorizing peroxidase oxidizing manganese(II). Appl. Microbiol. Biotechnol. 2015, 99, 8927–8942. [Google Scholar] [CrossRef] [PubMed]
- Vivanco, J.M.; Baluška, F. (Eds.) Secretions and Exudates in Biological Systems; Springer: Berlin/Heidelberg, Germany, 2012; Volume 12. [Google Scholar] [CrossRef]
- Garcia-Ruiz, E.; Mate, D.M.; Gonzalez-Perez, D.; Molina-Espeja, P.; Camarero, S.; Martínez, A.T.; Ballesteros, A.O.; Alcalde, M. Directed Evolution of Ligninolytic Oxidoreductases: From Functional Expression to Stabilization and Beyond. In Cascade Biocatalysis; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2014; pp. 1–22. [Google Scholar] [CrossRef]
- Hofrichter, M.; Ullrich, R.; Pecyna, M.J.; Liers, C.; Lundell, T. New and classic families of secreted fungal heme peroxidases. Appl. Microbiol. Biotechnol. 2010, 87, 871–897. [Google Scholar] [CrossRef]
- Zámocký, M.; Hofbauer, S.; Schaffner, I.; Gasselhuber, B.; Nicolussi, A.; Soudi, M.; Pirker, K.F.; Furtmüller, P.G.; Obinger, C. Independent evolution of four heme peroxidase superfamilies. Arch. Biochem. Biophys. 2015, 574, 108–119. [Google Scholar] [CrossRef] [PubMed]
- I Colpa, D.; Fraaije, M.W.; van Bloois, E. DyP-type peroxidases: A promising and versatile class of enzymes. J. Ind. Microbiol. Biotechnol. 2014, 41, 1–7. [Google Scholar] [CrossRef]
- Sugano, Y.; Matsushima, Y.; Tsuchiya, K.; Aoki, H.; Hirai, M.; Shoda, M. Degradation pathway of an anthraquinone dye catalyzed by a unique peroxidase DyP from Thanatephorus cucumeris Dec 1. Biodegradation 2009, 20, 433–440. [Google Scholar] [CrossRef]
- Liers, C.; Pecyna, M.J.; Kellner, H.; Worrich, A.; Zorn, H.; Steffen, K.T.; Hofrichter, M.; Ullrich, R. Substrate oxidation by dye-decolorizing peroxidases (DyPs) from wood- and litter-degrading agaricomycetes compared to other fungal and plant heme-peroxidases. Appl. Microbiol. Biotechnol. 2013, 97, 5839–5849. [Google Scholar] [CrossRef]
- Morozova, O.V.; Shumakovich, G.P.; Gorbacheva, M.A.; Shleev, S.V.; Yaropolov, A.I. “Blue” laccases. Biochemistry 2007, 72, 1136–1150. [Google Scholar] [CrossRef]
- Baldrian, P. Fungal laccases—Occurrence and properties. FEMS Microbiol. Rev. 2006, 30, 215–242. [Google Scholar] [CrossRef] [PubMed]
- Youn, H.-D.; Hah, Y.C.; Kang, S.-O. Role of laccase in lignin degradation by white-rot fungi. FEMS Microbiol. Lett. 1995, 132, 183–188. [Google Scholar] [CrossRef]
- Leonowicz, A.; Cho, N.; Luterek, J.; Jarosz-Wilkolazka, A.; Wojtas-Wasilewska, M.; Matuszewska, A.; Hofrichter, M.; Wesenberg, D.; Rogalski, J. Fungal laccase: Properties and activity on lignin. J. Basic Microbiol. 2001, 41, 185–227. [Google Scholar] [CrossRef] [PubMed]
- Kunamneni, A.; Ballesteros, A.; Plou, F.J.; Alcalde, M. Fungal laccase—A versatile enzyme for biotechnological applications. Communicating Current Research and Educational Topics and Trends in Applied Microbiology; Mendez-Vilas, A., Ed.; Formex: Badajoz, Spain, 2007; Volume 1, pp. 233–245. ISBN 978-84-611-9422-3. [Google Scholar]
- Shraddha; Shekher, R.; Sehgal, S.; Kamthania, M.; Kumar, A. Laccase: Microbial Sources, Production, Purification, and Potential Biotechnological Applications. Enzym. Res. 2011, 2011, 217861. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez Couto, S.; Toca-Herrera, J.L. Industrial and biotechnological applications of laccases: A review. Biotechnol. Adv. 2006, 24, 500–513. [Google Scholar] [CrossRef]
- Ragauskas, A.J.; Beckham, G.T.; Biddy, M.J.; Chandra, R.; Chen, F.; Davis, M.F.; Davison, B.H.; Dixon, R.A.; Gilna, P.; Keller, M.; et al. Lignin Valorization: Improving Lignin Processing in the Biorefinery. Science 2014, 344, 1246843. [Google Scholar] [CrossRef]
- Lange, L. Fungal Enzymes and Yeasts for Conversion of Plant Biomass to Bioenergy and High-Value Products. Microbiol. Spectr. 2017, 5, 1029–1048. [Google Scholar] [CrossRef]
- Erden, E.; Ucar, M.C.; Gezer, T.; Pazarlioglu, N.K. Screening for ligninolytic enzymes from autochthonous fungi and applications for decolorization of remazole marine blue. Braz. J. Microbiol. 2009, 40, 346–353. [Google Scholar] [CrossRef]
- Dhouib, A.; Hamza, M.; Zouari, H.; Mechichi, T.; Hmidi, R.; Labat, M.; Martinez, M.J.; Sayadi, S. Screening for Ligninolytic Enzyme Production by Diverse Fungi from Tunisia. World J. Microbiol. Biotechnol. 2005, 21, 1415–1423. [Google Scholar] [CrossRef]
- Longe, L.F.; Couvreur, J.; Grandchamp, M.L.; Garnier, G.; Allais, F.; Saito, K. Importance of Mediators for Lignin Degradation by Fungal Laccase. ACS Sustain. Chem. Eng. 2018, 6, 10097–10107. [Google Scholar] [CrossRef]
- Matsumura, E.; Shin, T.; Murao, S.; Yamamoto, E.; Kawano, T. New Enzymatic Colorimetric Reactions of Benzoic Acid Derivatives with ABTS [2,2′-Azino-di-(3-ethylbenzothiazoline-6-sulfonic acid) diammonium salt] in the Presence of Laccase. Biosci. Biotechnol. Biochem. 1987, 51, 2743–2750. [Google Scholar] [CrossRef]
- Sharma, A.; Aggarwal, N.K.; Yadav, A. First Report of Lignin Peroxidase Production from Alternaria alternata ANF238 Isolated from Rotten Wood Sample. Bioeng. Biosci. 2016, 4, 76–87. [Google Scholar] [CrossRef]
- More, S.S.; Renuka, P.S.; Pruthvi, K.; Swetha, M.; Malini, S.; Veena, S.M. Isolation, Purification, and Characterization of Fungal Laccase from Pleurotus sp. Enzym. Res. 2011, 2011, 248735. [Google Scholar] [CrossRef] [PubMed]
- Kut, K.; Stefaniuk, I.; Bartosz, G.; Sadowska-Bartosz, I. Formation of a Purple Product upon the Reaction of ABTS Radicals with Proteins. Int. J. Mol. Sci. 2023, 24, 8912. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhao, J.; Jiang, J.; Xu, H.; Zhang, N.; Xie, J.; Wei, M. Isolation and Characterization of Bacillus Sp. Capable of Degradating Alkali Lignin. Front. Energy Res. 2021, 9, 807286. [Google Scholar] [CrossRef]
- French, K.E. Assessing the bioenergy potential of grassland biomass from conservation areas in England. Land Use Policy 2019, 82, 700–708. [Google Scholar] [CrossRef]






| Species | Feedstock Used | Lignin Degradation (%) | Time (Days) | Reference |
|---|---|---|---|---|
| Bacteria | ||||
| Pseudomonas spp. | Kraft lignin | 39 | 52 | [23] |
| Poplar wood | 40–52 | 30 | [24] | |
| Kraft lignin | 20 | 40–60 | [23] | |
| Acinetobacter spp. | Poplar wood | 47–57 | 30 | [24,25] |
| Xanthomonas spp. | Poplar wood | 39–48 | 30 | [24,25] |
| Streptomyces badius | Indulin lignin | 3–4 | 35 | [26] |
| Streptomyces viridosporous | Indulin lignin | 3–4 | 35 | [26] |
| Streptomyces cyaneus | Barley straw | 29–52 | 21 | [27] |
| Thermomonospora mesophila | Barley straw | 36–48 | 21 | [24,27] |
| Fungi | ||||
| Pleurotus ostreatus | Cotton stalks | 40 | 30 | [28] |
| Beech wood | 56.5 | 120 | [28] | |
| Phanerochaete chrysosporium | Cotton stalks | 60 | 30 | [28] |
| Cotton stalks | 28 | 14 | [23] | |
| Trametes versicolor spp. | Beech wood | 57.4 | 120 | [29] |
| Bamboo culms | 9–24 | 28 | [30,31] | |
| Irpex lacteus | Cornstalks | 15 | 11.84 | [32] |
| Echinodontium taxodii 2538 | Bamboo culms | 24 | 28 | [30] |
| Phlebia sp. MG-60 | Oak wood | 40.6 | 56 | [33] |
| Ceriporia lacerata | Red pine | 13 | 56 | [34] |
| Stereum hirsutum | Red pine | 15 | 56 | [31] |
| Ceriporiopsis subvermispora | Corn stover | 39.2 | 42 | [35,36] |
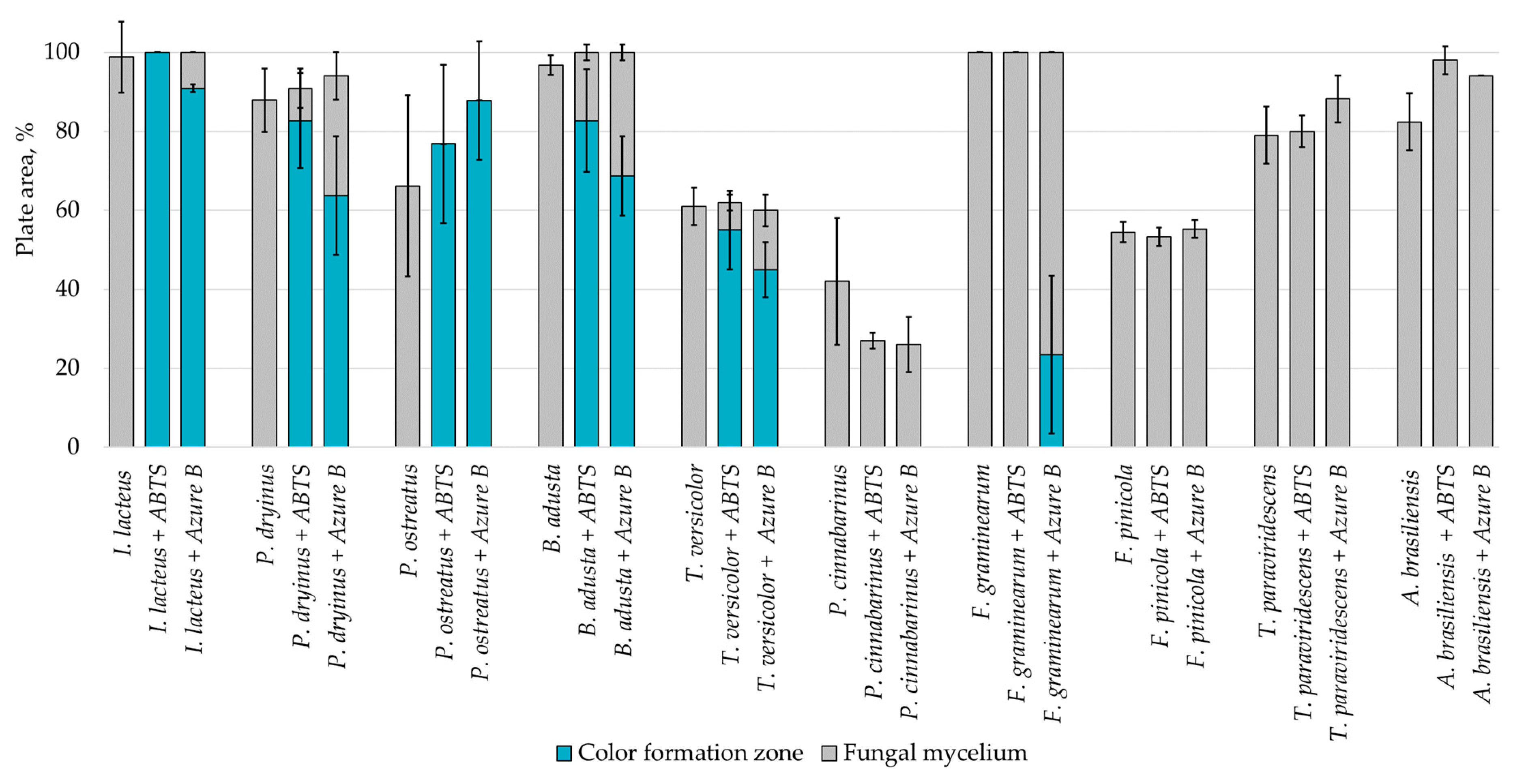
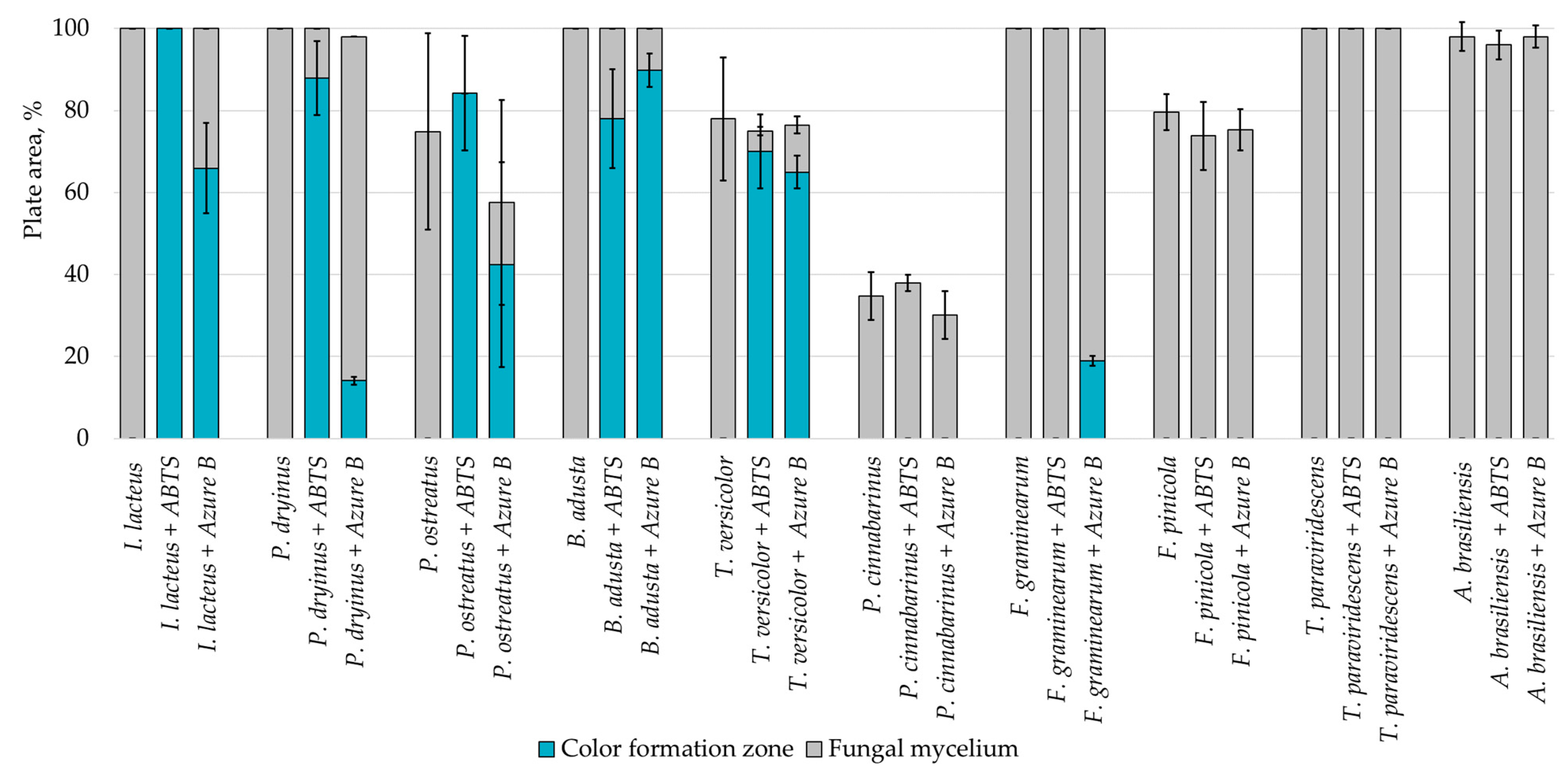
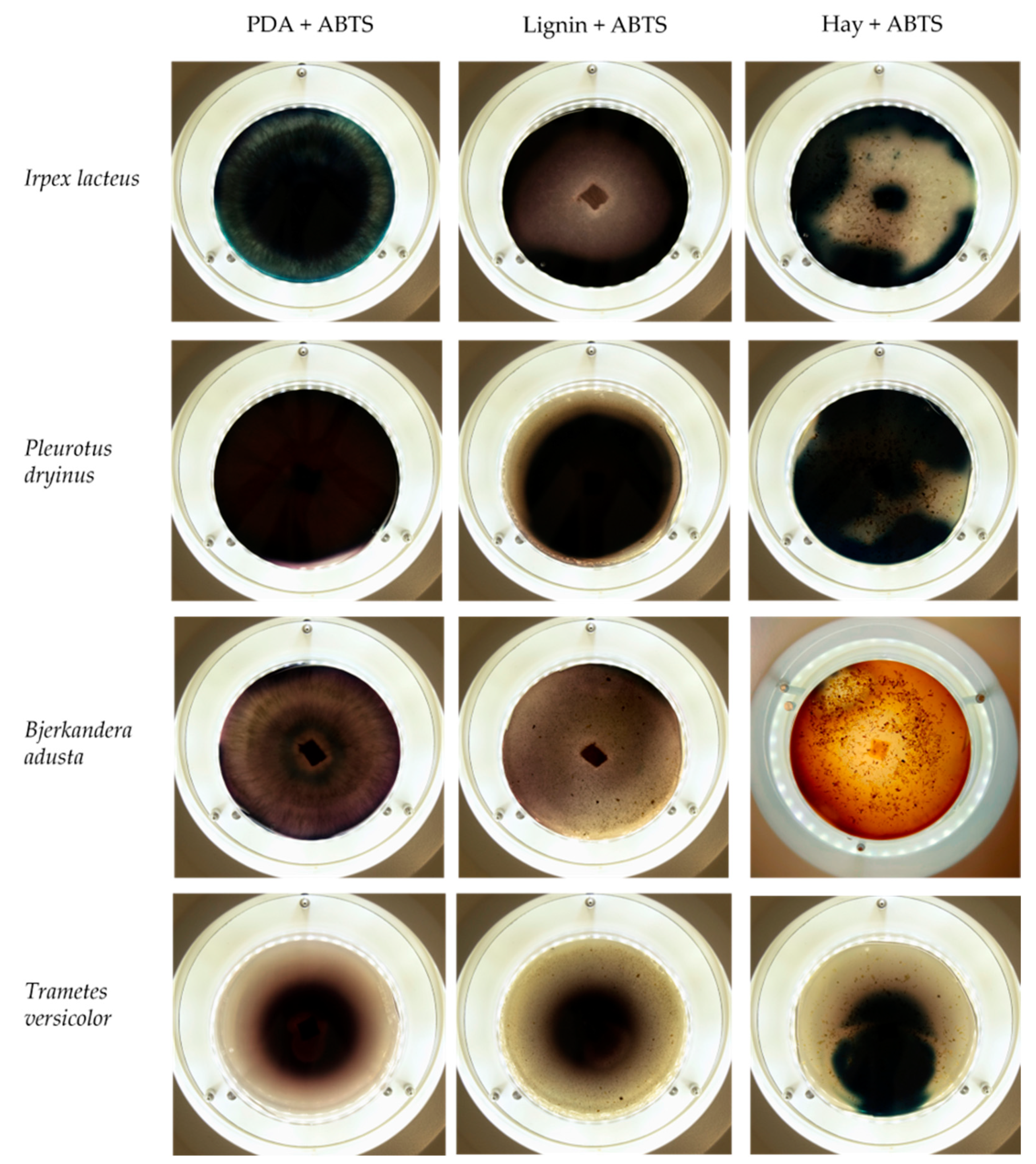
| Substrate | - | 0.1% w/v ABTS | 0.01% w/v Azure B |
|---|---|---|---|
| Potato dextrose agar |  |  |  |
| Lignin |  |  |  |
| Hay |  |  |  |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Civzele, A.; Stipniece-Jekimova, A.A.; Mezule, L. Fungal Ligninolytic Enzymes and Their Application in Biomass Lignin Pretreatment. J. Fungi 2023, 9, 780. https://doi.org/10.3390/jof9070780
Civzele A, Stipniece-Jekimova AA, Mezule L. Fungal Ligninolytic Enzymes and Their Application in Biomass Lignin Pretreatment. Journal of Fungi. 2023; 9(7):780. https://doi.org/10.3390/jof9070780
Chicago/Turabian StyleCivzele, Anna, Alise Anna Stipniece-Jekimova, and Linda Mezule. 2023. "Fungal Ligninolytic Enzymes and Their Application in Biomass Lignin Pretreatment" Journal of Fungi 9, no. 7: 780. https://doi.org/10.3390/jof9070780
APA StyleCivzele, A., Stipniece-Jekimova, A. A., & Mezule, L. (2023). Fungal Ligninolytic Enzymes and Their Application in Biomass Lignin Pretreatment. Journal of Fungi, 9(7), 780. https://doi.org/10.3390/jof9070780