
The Potential of Metarhizium anisopliae Blastospores to Control Aedes aegypti Larvae in the Field

 , , , and
, , , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Mosquito Collection and Rearing
2.2. Fungal Isolate
2.3. Conidial Production and Suspensions
2.4. Blastospore Production and Suspension
2.5. Laboratory and Semi-field Bioassays
2.6. Blastospore Persistence under Laboratory Conditions
2.7. Blastospore Virulence When Tested against Larvae under Totally Shaded and Partially Shaded Conditions in the Field
2.8. Statistical Analysis
3. Results
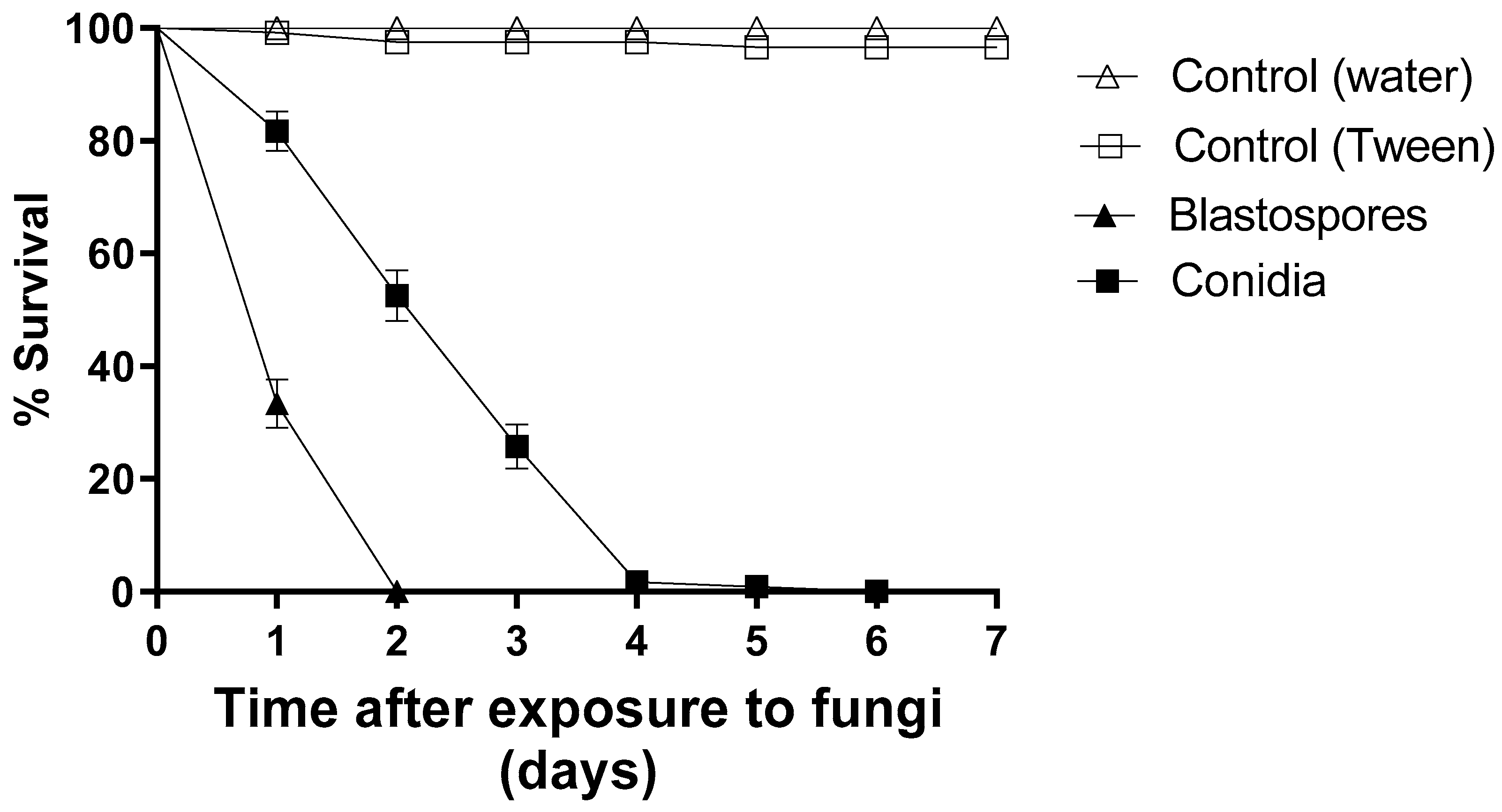
3.1. Comparison of Blastospore and Conidial Virulence under Laboratory Conditions
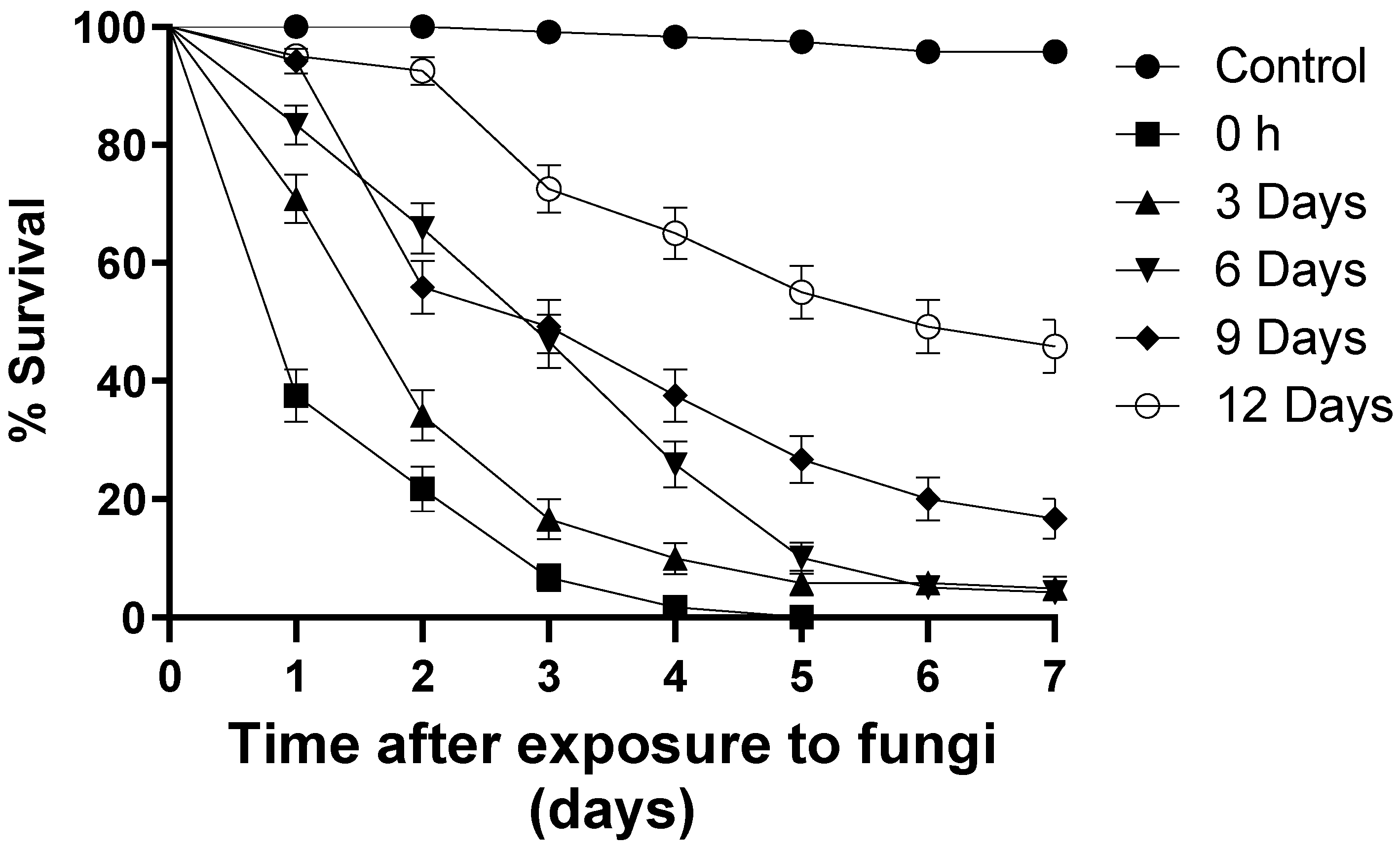
3.2. Blastospore Persistence under Laboratory Conditions
3.3. Assessment of Blastospore Virulence under Semi-Field Conditions: Totally Shaded (TS) and Partially Shaded (PS)
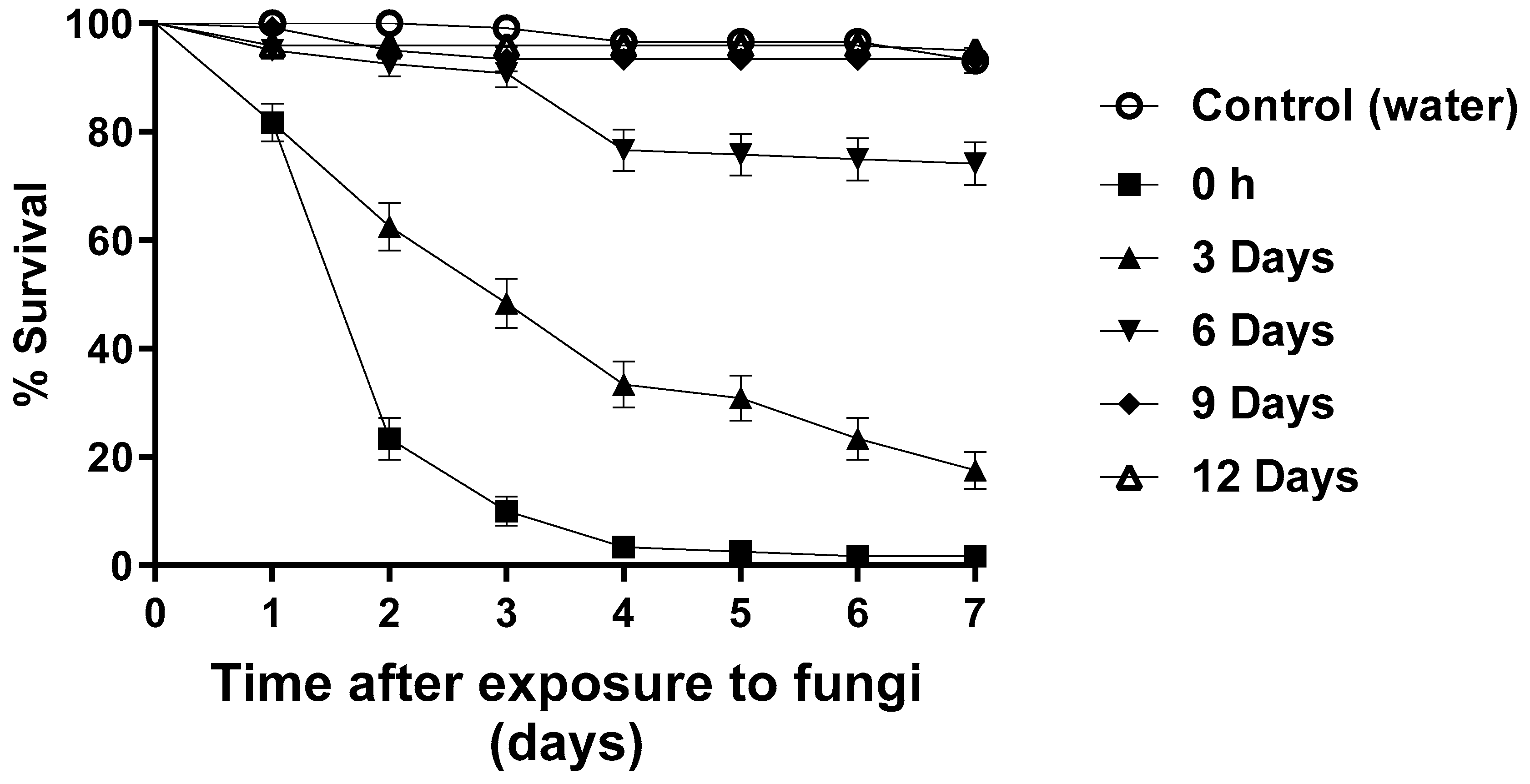
3.4. Blastospore Persistence under Semi-Field Conditions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Powell, J.R. Mosquito-borne human viral diseases: Why Aedes aegypti? Am. J. Trop. Med. Hyg. 2018, 98, 1563–1565. [Google Scholar] [CrossRef] [PubMed]
- Zulfa, R.; Lo, W.C.; Cheng, P.C.; Martini, M.; Chuang, T.W. Updating the insecticide resistance status of Aedes aegypti and Aedes albopictus in Asia: A systematic review and meta-analysis. Trop. Med. Infect. Dis. 2022, 7, 306. [Google Scholar] [CrossRef] [PubMed]
- Estep, A.S.; Sanscrainte, N.D.; Waits, C.M.; Bernard, S.J.; Lloyd, A.M.; Lucas, K.J.; Buckner, E.A.; Vaidyanathan, R.; Morreale, R.; Conti, L.A.; et al. Quantification of permethrin resistance and kdr alleles in Florida strains of Aedes aegypti (L.) and Aedes albopictus (Skuse). PLoS Negl. Trop. Dis. 2018, 12, e0006544. [Google Scholar] [CrossRef] [PubMed]
- Al-Amin, H.M.; Johora, F.T.; Irish, S.R.; Hossainey, M.R.H.; Vizcaino, L.; Paul, K.K.; Khan, W.A.; Haque, R.; Alam, M.S.; Lenhart, A. Insecticide resistance status of Aedes aegypti in Bangladesh. Parasit. Vectors 2020, 13, 622. [Google Scholar] [CrossRef]
- Parker, C.; Ramirez, D.; Thomas, C.; Connelly, C.R. Baseline susceptibility status of Florida populations of Aedes aegypti (Diperta: Culicide) and Aedes albopictus. J. Med. Entomol. 2020, 57, 1550–1559. [Google Scholar] [CrossRef]
- Silalahi, C.N.; Tu, W.C.; Chang, N.T.; Singham, G.V.; Ahmad, I.; Neoh, K.B. Insecticide resistance profiles and synergism of field Aedes aegypti from Indonesia. PLoS Negl. Trop. Dis. 2022, 16, e0010501. [Google Scholar] [CrossRef]
- Butt, T.M.; Greenfield, B.P.J.; Greig, C.; Maffeis, T.G.; Taylor, J.W.D.; Piasecka, J.; Dudley, E.; Abdulla, A.; Dubovskiy, I.M.; Garrido-Jurado, I.; et al. Metarhizium anisopliae pathogenesis of mosquito larvae: A verdict of accidental death. PLoS ONE 2013, 13, e81686. [Google Scholar] [CrossRef]
- Gomes, S.A.; Paula, A.R.; Ribeiro, A.; Moraes, C.O.P.; Santos, J.W.A.B.; Silva, C.P.; Samuels, R.I. Neem oil increases the efficiency of the entomopathogenic fungus Metarhizium anisopliae for the control of Aedes aegypti (Diptera: Culicidae) larvae. Parasit. Vectors 2015, 8, 669. [Google Scholar] [CrossRef]
- Noskov, Y.A.; Polenogova, O.V.; Yaroslavtseva, O.N.; Belevich, O.E.; Yurchenko, Y.A.; Chertkova, E.A.; Kryukova, N.A.; Kryukov, V.Y.; Glupov, V.V. Combined effect of the entomopathogenic fungus Metarhizium robertsii and avermectins on the survival and immune response of Aedes aegypti larvae. Peer J. 2019, 7, e7931. [Google Scholar] [CrossRef]
- Rodrigues, J.; Bergamini, C.; Montalva, C.; Humber, R.A.; Luz, C. Simple method to detect and to isolate entomopathogenic fungi (Hypocreales) from mosquito larvae. J. Invertebr. Pathol. 2021, 182, 107581. [Google Scholar] [CrossRef]
- Amóra, S.S.A.; Bevilaqua, C.M.L.; Feijó, F.M.C.; Pereira, R.H.M.A.; Alves, N.D.; Freire, F.A.M.; Kamimura, M.T.; de Oliveira, D.M.; Luna-Alves Lima, E.A.; Rocha, M.F.G. The effects of the fungus Metarhizium anisopliae var. acridum on different stages of Lutzomyia longipalpis (Diptera: Psychodidae). Acta Trop. 2010, 113, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Castellanos-Sánchez, P.; Falconi-Agapito, F.; Pariona, N.; Paredes-Esquivel, C. Isolation of Beauveria bassiana Pr-11 from Andean orthopterans and its effectiveness against Chagas disease vectors in Peru. Trop. Biomed. 2020, 37, 864–870. [Google Scholar] [CrossRef] [PubMed]
- Figueredo, L.A.; Luna, R.L.N.; Miranda, D.E.O.; Silva, F.J.; Otranto, D.; Cafarchia, C.; Figueiredo, R.C.B.Q.; Dantas-Torres, F.; Brandão-Filho, S.P. Beauveria bassiana (Hypocreales: Cordycipitaceae) reduces the survival time of Lutzomyia longipalpis (Diptera: Psychodidae), the main vector of the visceral leishmaniasis agent in the Americas. J. Med. Entomol. 2020, 13, 2025–2029. [Google Scholar] [CrossRef] [PubMed]
- Marciano, A.F.; Mascarin, G.M.; Franco, R.F.F.; Golo, P.S.; Jaronski, S.T.; Fernandes, É.K.K.; Bittencourt, V.R.E.P. Innovative granular formulation of Metarhizium robertsii microsclerotia and blastospores for cattle tick control. Sci. Rep. 2021, 11, 4972. [Google Scholar] [CrossRef] [PubMed]
- Alkhaibari, A.M.; Carolino, A.T.; Yavasoglu, S.I.; Maffeis, T.; Matosso, T.C.; James, C.B.; Bull, J.C.; Samuels, R.I.; Butt, T.M. Metarhizium brunneum blastospore pathogenesis in Aedes aegypti larvae: Attack on several fronts accelerates mortality. PLoS Pathog. 2016, 12, e1005715. [Google Scholar] [CrossRef]
- Alkhaibari, A.M.; Carolino, A.T.; Bull, J.C.; Samuels, R.I.; Butt, T.M. Differential pathogenicity of Metarhizium blastospores and conidia against larvae of three mosquito species. J. Med. Entomol. 2017, 54, 696–704. [Google Scholar] [CrossRef]
- Alkhaibari, A.M.; Lord, A.M.; Maffeis, T.; Bull, J.C.; Olivares, F.L.; Samuels, R.I.; Butt, T.M. Highly specific host-pathogen interactions influence Metarhizium brunneum blastospore virulence against Culex quinquefasciatus larvae. Virulence 2018, 9, 1449–1467. [Google Scholar] [CrossRef]
- Alkhaibari, A.M.; Wood, M.J.; Yavasoglu, S.I.; Bull, J.C.; Butt, T.M. Optimizing the application timing and dosage of Metarhizium brunneum (Hypocreales: Clavicipitaceae) as a biological control agent of Aedes aegypti (Diptera: Culicidae) larvae. J. Med. Entomol. 2023, 60, 339–345. [Google Scholar] [CrossRef]
- Ramirez, J.L.; Muturi, E.J.; Barletta, A.B.F.; Rooney, A.P. The Aedes aegypti IMD pathway is a critical component of the mosquito antifungal immune response. Dev. Comp. Immunol. 2019, 95, 1–9. [Google Scholar] [CrossRef]
- Lovett, B.; Bilgo, E.; Millogo, S.A.; Ouattarra, A.K.; Sare, I.; Gnambani, E.J.; Dabire, R.K.; Diabate, A.; St Leger, R.J. Transgenic Metarhizium rapidly kills mosquitoes in a malaria-endemic region of Burkina Faso. Science 2019, 364, 894–897. [Google Scholar] [CrossRef]
- Bitencourt, R.O.B.; Mallet, J.R.S.; Mesquita, M.; Gôlo, P.S.; Fiorotti, J.; Bittencourt, R.E.P.; Pontes, E.G.; Angelo, I.C. Larvicidal activity, route of interaction and ultrastructural changes in Aedes aegypti exposed to entomopathogenic fungi. Acta Trop. 2021, 213, 105732. [Google Scholar] [CrossRef] [PubMed]
- Bitencourt, R.O.B.; Faria, F.S.; Marchesini, P.; Santos-Mallet, J.R.; Camargo, M.G.; Bittencourt, V.R.E.P.; Pontes, E.G.; Pereira, D.B.; Chaves, D.S.A.; Angelo, I.C. Entomopathogenic fungi and Schinus molle essential oil: The combination of two eco-friendly agents against Aedes aegypti larvae. J. Invertebr. Pathol. 2022, 194, 107827. [Google Scholar] [CrossRef] [PubMed]
- Mannino, M.C.; Huarte-Bonnet, C.; Davyt-Colo, B.; Pedrini, N. Is the insect cuticle the only entry gate for fungal infection? Insights into alternative modes of action of entomopathogenic fungi. J. Fungi. 2019, 5, 33. [Google Scholar] [CrossRef]
- Carolino, A.T.; Teodoro, T.B.P.; Gomes, S.A.; Silva, C.P.; Samuels, R.I. Production of conidia using different culture media modifies the virulence of the entomopathogenic fungus Metarhizium against Aedes aegypti larvae. J. Vector Borne Dis. 2021, 58, 346–351. [Google Scholar] [CrossRef]
- Paula, A.R.; Brito, E.S.; Pereira, C.R.; Carrera, M.P.; Samuels, R.I. Susceptibility of adult Aedes aegypti (Diptera: Culicidae) to infection by Metarhizium anisopliae and Beauveria bassiana: Prospects for Dengue vector control. Biocontrol Sci. Technol. 2008, 18, 1017–1025. [Google Scholar] [CrossRef]
- Paula, A.R.; Silva, L.E.I.; Ribeiro, A.; Silva, G.A.; Silva, C.P.; Butt, T.M.; Samuels, R.I. Metarhizium anisopliae blastospores are highly virulent to adult Aedes aegypti, an important arbovirus vector. Parasit. Vectors 2021, 14, 555. [Google Scholar] [CrossRef]
- Carolino, A.T.; Gomes, S.A.; Teodoro, T.B.P.; Mattoso, T.C.; Samuels, R.I. Aedes aegypti pupae are highly susceptible to infection by Metarhizium anisopliae blastospores. J. Pure Appl. Microbiol. 2019, 13, 1629–1634. [Google Scholar] [CrossRef]
- Jaronski, S.T.; Mascarin, G.M. Mass production of fungal entomopathogens. In Microbial Control of Insect and Mite Pests; Lacey, L.A., Ed.; Academic Press: Amsterdam, The Netherlands, 2017; Volume 9, pp. 141–155. [Google Scholar] [CrossRef]
- Mascarin, G.M.; Jackson, M.A.; Kobori, N.N.; Behle, R.W.; Delalibera Júnior, Í. Liquid culture fermentation for rapid production of desiccation tolerant blastospores of Beauveria bassiana and Isaria fumosorosea strains. J. Invertebr. Pathol. 2015, 127, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Miranpuri, G.S.; Khachatourians, G.G. Larvicidal activity of blastospores and conidiospores of Beauveria bassiana (strain GK 2016) against age groups of Aedes aegypti. Vet. Parasitol. 1990, 37, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Greenfield, B.P.J.; Lord, A.M.; Dudley, E.; Butt, T.M. Conidia of the insect pathogenic fungus, Metarhizium anisopliae, fail to adhere to mosquito larval cuticle. R. Soc. Open Sci. 2014, 1, 140193. [Google Scholar] [CrossRef]
- Daoust, R.A.; Roberts, D.W. Virulence of natural and insect-passaged strains of Metarhizium anisopliae to mosquito larvae. J. Invertebr. Pathol. 1982, 40, 107–117. [Google Scholar] [CrossRef]
- Blanford, S.; Chan, B.H.K.; Jenkins, N.; Sim, D.; Turner, R.J.; Read, A.F.; Thomas, M.B. Fungal pathogen reduces potential for malaria transmission. Science 2005, 308, 1638–1641. [Google Scholar] [CrossRef] [PubMed]
- Soares, G.G., Jr.; Pinnock, D.E. Effect of temperature on germination, growth, and infectivity of the mosquito pathogen Tolypocladium cylindrosporum (Deuteromycotina; Hyphomycetes). J. Invertebr. Pathol. 1984, 43, 242–247. [Google Scholar] [CrossRef]
- WHO (World Health Organization). Control: Environmental management. In Guidelines for Dengue Surveillance and Mosquito Control; WHO Library Cataloguing in Publication Data; World Health Organization Regional Office for the Western Pacific Manila: Manila, Philippines, 2003; pp. 23–28. ISBN 92-9061-068-9. [Google Scholar]
- Jackson, M.A.; Mcguire, M.R.; Lacey, L.A.; Wraight, S.P. Liquid culture production of desiccation tolerant blastospores of the bioinsecticidal fungus Paecilomyces fumosoroseus. Mycol. Res. 1997, 101, 35–41. [Google Scholar] [CrossRef]
- Iwanicki, N.S.; Ferreira, B.O.; Mascarin, G.M.; Delalibera, Í., Jr. Modified Adamek’s medium renders high yields of Metarhizium robertsii blastospores that are desiccation tolerant and infective to cattle-tick larvae. Fungal Biol. 2018, 122, 883–890. [Google Scholar] [CrossRef]
- Mascarin, G.M.; Kobori, N.N.; Jackson, M.A.; Dunlap, C.A.; Delalibera, Í., Jr. Nitrogen sources affect productivity, desiccation tolerance and storage stability of Beauveria bassiana blastospores. J. Appl. Microbiol. 2018, 124, 810–820. [Google Scholar] [CrossRef]
- Lorenz, S.-C.; Humbert, P.; Patel, A.V. Chitin increases drying survival of encapsulated Metarhizium pemphigi blastospores for Ixodes ricinus control. Ticks Tick Borne Dis. 2022, 11, 101537. [Google Scholar] [CrossRef]
- Mascarin, G.M.; Jackson, M.A.; Kobori, N.N.; Behle, R.W.; Dunlap, C.W.; Delalibera, Í., Jr. Glucose concentration alters dissolved oxygen levels in liquid cultures of Beauveria bassiana and affects formation and bioefficacy of blastospores. Appl. Microbiol. Biotechnol. 2015, 99, 6653–6665. [Google Scholar] [CrossRef]
- Leger, R.J.S.; Staples, R.C.; Roberts, D.W. Cloning and regulatory analysis of starvation-stress gene, ssgA, encoding a hydrophobin-like protein from the entomopathogenic fungus, Metarhizium anisopliae. Gene 1992, 120, 119–124. [Google Scholar] [CrossRef]
- Wong, J.; Stoddard, S.T.; Astete, H.; Morrison, A.C.; Scott, T.W. Oviposition site selection by the dengue vector Aedes aegypti and its implications for dengue control. PLoS Negl. Trop. Dis. 2011, 5, e1015. [Google Scholar] [CrossRef]
- Rangel, D.E.N.; Braga, G.U.L.; Flint, S.D.; Anderson, A.J.; Roberts, D.W. Variations in UV-B tolerance and germination speed of Metarhizium anisopliae conidia produced on insects and artificial substrates. J. Invertebr. Pathol. 2004, 81, 77–83. [Google Scholar] [CrossRef]
- Carolino, A.T.; Paula, A.R.; Silva, C.P.; Butt, T.M.; Samuels, R.I. Monitoring persistence of the entomopathogenic fungus Metarhizium anisopliae under simulated field conditions with the aim of controlling adult Aedes aegypti (Diptera: Culicidae). Parasit. Vectors 2014, 7, 2–7. [Google Scholar] [CrossRef]
- Falvo, M.L.; Pereira-Junior, R.A.; Rodrigues, J.; Lastra, C.C.L.; García, J.J.; Fernandes, E.K.K.; Luz, C. UV-B radiation reduces in vitro germination of Metarhizium anisopliae s.l. but does not affect virulence in fungus-treated Aedes aegypti adults and development on dead mosquitoes. J. Appl. Microbiol. 2016, 121, 1710–1717. [Google Scholar] [CrossRef] [PubMed]
- Braga, G.U.L.; Flint, S.D.; Miller, C.D.; Anderson, A.J.; Roberts, D.W. Variability in response to UV-B among species and strains of Metarhizium isolated from sites at latitudes from 61° N to 54° S. J. Invertebr. Pathol. 2001, 78, 98–108. [Google Scholar] [CrossRef]
- Reddy, N.P.; Khan, P.A.A.; Devi, K.U.; Victor, J.S.; Sharma, H.C. Assessment of the suitability of Tinopal as an enhancing adjuvant in formulations of the insect pathogenic fungus Beauveria bassiana (Bals.) Vuillemin. Pest Manag. Sci. 2008, 9, 909–915. [Google Scholar] [CrossRef] [PubMed]
- Luz, C.; Silva, I.G.; Magalhães, B.P.; Cordeiro, C.M.T.; Tigano, M.S. Control of Triatoma infestans (Klug) (Reduviidae: Triatominae) with Beauveria bassiana (Bals.) Vuill.: Preliminary assays on formulation and application in the field. Biological Control An. Soc. Entomol. Bras. 1999, 98, 101–110. [Google Scholar] [CrossRef]
- Inglis, G.D.; Goettel, M.S.; Johnson, D.L. Influence of ultraviolet light protectants on persistence of the entomopathogenic fungus, Beauveria bassiana. Biol. Control 1995, 5, 581–590. [Google Scholar] [CrossRef]
- Braga, G.U.L.; Rangel, D.E.N.; Fernandes, E.K.K.; Flint, S.D.; Roberts, D.W. Molecular and physiological effects of environmental UV radiation on fungal conidia. Curr. Genet. 2015, 61, 405–425. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | % Survival (±SD) | S50 |
|---|---|---|
| Blastospores TS | 5.8 ± 2.1 c | 1 |
| Blastospores PS | 43.3 ± 4.5 b | 2 |
| Control TS | 97.5 ± 1.3 a | ND |
| Control PS | 98.3 ± 1.1 a | ND |
| Treatments | % Survival ± SD | S50 |
|---|---|---|
| 0 h | 23.3 ± 3.8 c | 2 |
| 3 days | 62.5 ± 4.4 b | 3 |
| 6 days | 92.5 ± 2.4 a | ND |
| 9 days | 95 ± 1.9 a | ND |
| 12 days | 95.8 ± 1.8 a | ND |
| Control (water) | 100 a | ND |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gomes, S.A.; Carolino, A.T.; Teodoro, T.B.P.; Silva, G.A.; Bitencourt, R.d.O.B.; Silva, C.P.; Alkhaibari, A.M.; Butt, T.M.; Samuels, R.I. The Potential of Metarhizium anisopliae Blastospores to Control Aedes aegypti Larvae in the Field. J. Fungi 2023, 9, 759. https://doi.org/10.3390/jof9070759
Gomes SA, Carolino AT, Teodoro TBP, Silva GA, Bitencourt RdOB, Silva CP, Alkhaibari AM, Butt TM, Samuels RI. The Potential of Metarhizium anisopliae Blastospores to Control Aedes aegypti Larvae in the Field. Journal of Fungi. 2023; 9(7):759. https://doi.org/10.3390/jof9070759
Chicago/Turabian StyleGomes, Simone Azevedo, Aline Teixeira Carolino, Thais Berçot Pontes Teodoro, Gerson Adriano Silva, Ricardo de Oliveira Barbosa Bitencourt, Carlos Peres Silva, Abeer M. Alkhaibari, Tariq M. Butt, and Richard Ian Samuels. 2023. "The Potential of Metarhizium anisopliae Blastospores to Control Aedes aegypti Larvae in the Field" Journal of Fungi 9, no. 7: 759. https://doi.org/10.3390/jof9070759
APA StyleGomes, S. A., Carolino, A. T., Teodoro, T. B. P., Silva, G. A., Bitencourt, R. d. O. B., Silva, C. P., Alkhaibari, A. M., Butt, T. M., & Samuels, R. I. (2023). The Potential of Metarhizium anisopliae Blastospores to Control Aedes aegypti Larvae in the Field. Journal of Fungi, 9(7), 759. https://doi.org/10.3390/jof9070759