

Characterization of the Community of Black Meristematic Fungi Inhabiting the External White Marble of the Florence Cathedral


Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling and Cultivation of Meristematic Fungi
2.2. DNA Extraction and PCR Amplification
2.3. Phylogenetic Analysis
2.4. Thermal Preferences, Salt Tolerance, and Acid Test
2.5. Sensitivity Test to Selected Biocides
2.6. Statistical Analyses
3. Results
3.1. Cultivation and Identification of Meristematic Fungi
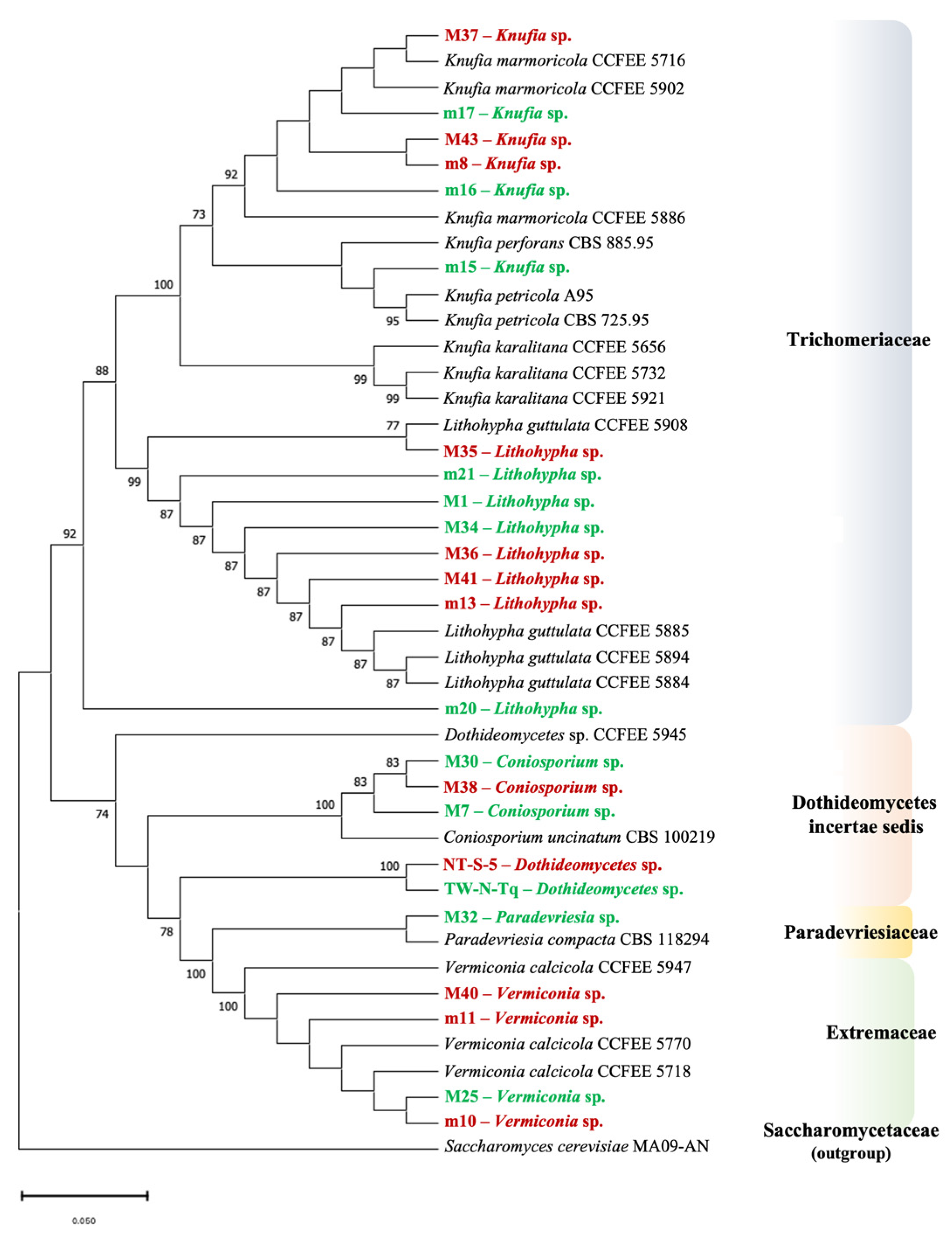
3.2. Multi-Locus Phylogeny
3.3. Physiological Analyses on RIF Strains
3.3.1. Thermal Preferences
3.3.2. Salt Tolerance
3.3.3. Carbonate Solubilization Test
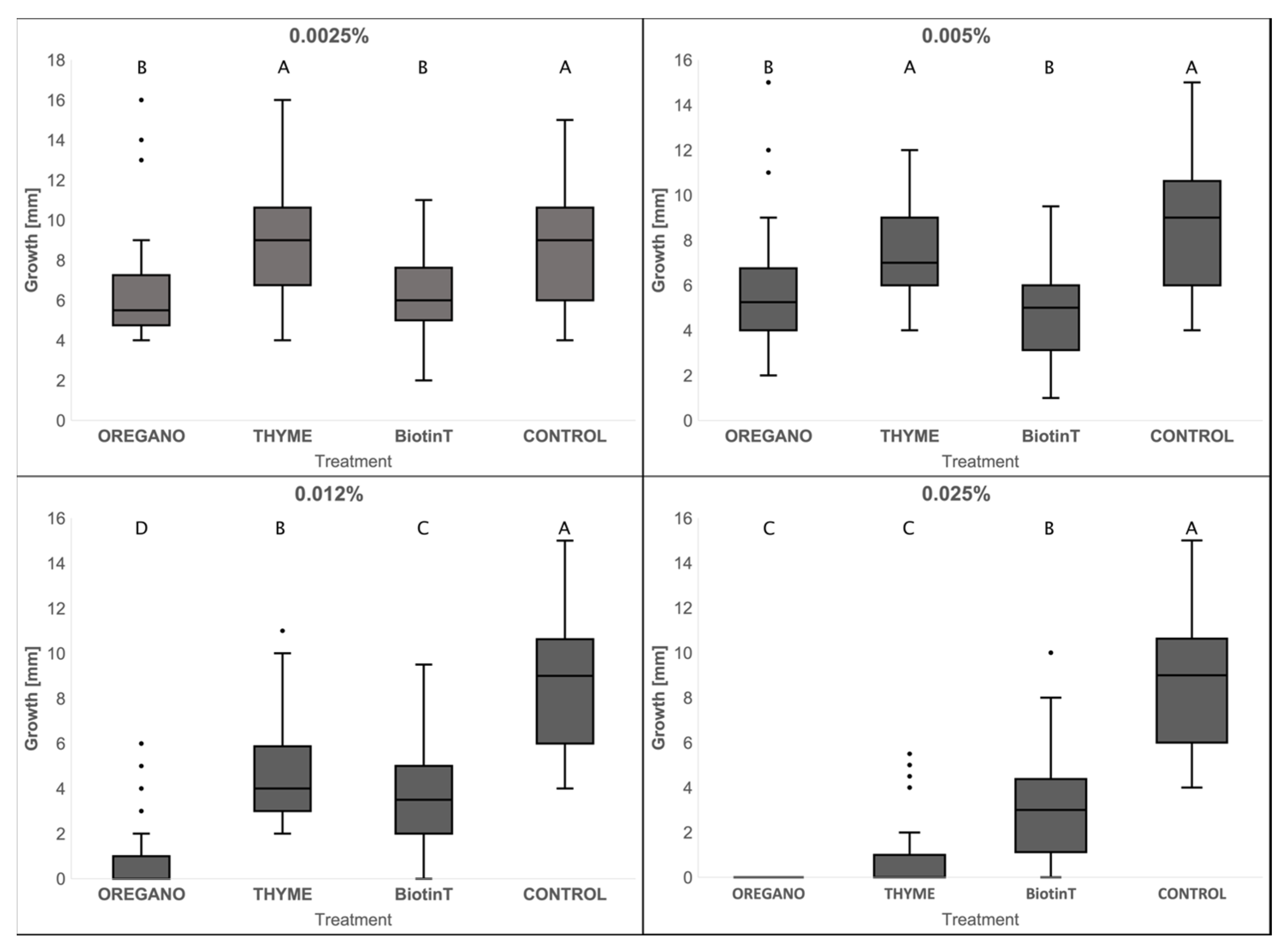
3.3.4. Sensitivity to Biocides
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zanardini, E.; Abbruscato, P.; Ghedini, N.; Realini, M.; Sorlini, C. Infuence of Atmospheric Pollutants on the Biodeterioration of Stone. Int. Biodeterior. Biodegrad. 2000, 45, 35–42. [Google Scholar] [CrossRef]
- Santo, A.P.; Cuzman, O.A.; Petrocchi, D.; Pinna, D.; Salvatici, T.; Perito, B. Black on White: Microbial Growth Darkens the External Marble of Florence Cathedral. Appl. Sci. 2021, 11, 6163. [Google Scholar] [CrossRef]
- Coleine, C.; Selbmann, L. Black Fungi Inhabiting Rock Surfaces. In Life at Rock Surfaces: Challenged by Extreme Light, Temperature and Hydration Fluctuations; Büdel, B., Friedl, T., Eds.; Walter de Gruyter GmbH & Co KG: Berlin, Germany, 2021; pp. 57–77. [Google Scholar] [CrossRef]
- Sterflinger, K. Black Yeasts and Meristematic Fungi: Ecology, Diversity and Identification. In Biodiversity and Ecophysiology of Yeasts; Springer: Berlin/Heidelberg, Germany, 2006; pp. 501–514. [Google Scholar] [CrossRef]
- Selbmann, L.; de Hoog, G.S.; Zucconi, L.; Isola, D.; Ruisi, S.; Gerrits van den Ende, A.H.G.; Ruibal, C.; De Leo, F.; Urzì, C.; Onofri, S. Drought Meets Acid: Three New Genera in a Dothidealean Clade of Extremotolerant Fungi. Stud. Mycol. 2008, 61, 1–20. [Google Scholar] [CrossRef]
- Marvasi, M.; Donnarumma, F.; Frandi, A.; Mastromei, G.; Sterflinger, K.; Tiano, P.; Perito, B. Black Microcolonial Fungi as Deteriogens of Two Famous Marble Statues in Florence, Italy. Int. Biodeterior. Biodegrad. 2012, 68, 36–44. [Google Scholar] [CrossRef]
- Onofri, S.; De La Torre, R.; De Vera, J.P.; Ott, S.; Zucconi, L.; Selbmann, L.; Scalzi, G.; Venkateswaran, K.J.; Rabbow, E.; Sánchez Iñigo, F.J.; et al. Survival of Rock-Colonizing Organisms after 1.5 Years in Outer Space. Astrobiology 2012, 12, 508–516. [Google Scholar] [CrossRef]
- Zakharova, K.; Tesei, D.; Marzban, G.; Dijksterhuis, J.; Wyatt, T.; Sterflinger, K. Microcolonial Fungi on Rocks: A Life in Constant Drought? Mycopathologia 2013, 175, 537–547. [Google Scholar] [CrossRef]
- Sterflinger, K.; Krumbein, W.E. Multiple Stress Factors Affecting Growth of Rock-inhabiting Black Fungi. Bot. Acta 1995, 108, 490–496. [Google Scholar] [CrossRef]
- Sterflinger, K. Temperature and NaCl-Tolerance of Rock-Inhabiting Meristematic Fungi. Antonie Van Leeuwenhoek 1998, 74, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Ruibal, C.; Gueidan, C.; Selbmann, L.; Gorbushina, A.A.; Crous, P.W.; Groenewald, J.Z.; Muggia, L.; Grube, M.; Isola, D.; Schoch, C.L.; et al. Phylogeny of Rock-Inhabiting Fungi Related to Dothideomycetes. Stud. Mycol. 2009, 64, 123–133. [Google Scholar] [CrossRef]
- Gorbushina, A.A. Life on the Rocks. Environ. Microbiol. 2007, 9, 1613–1631. [Google Scholar] [CrossRef]
- Kejžar, A.; Gobec, S.; PlemenitaŠ, A.; Lenassi, M. Melanin Is Crucial for Growth of the Black Yeast Hortaea Werneckii in Its Natural Hypersaline Environment. Fungal Biol. 2013, 117, 368–379. [Google Scholar] [CrossRef]
- Schumacher, J.; Gorbushina, A.A. Light Sensing in Plant- and Rock-Associated Black Fungi. Fungal Biol. 2020, 124, 407–417. [Google Scholar] [CrossRef]
- Tesei, D. Black Fungi Research: Out-of-This-World Implications. Encyclopedia 2022, 2, 212–229. [Google Scholar] [CrossRef]
- Onofri, S.; Zucconi, L.; Isola, D.; Selbmann, L. Rock-Inhabiting Fungi and Their Role in Deterioration of Stone Monuments in the Mediterranean Area. Plant Biosyst. 2014, 148, 384–391. [Google Scholar] [CrossRef]
- Sterflinger, K.; De Baere, R.; De Hoog, G.S.; De Wachter, R.; Krumbein, W.E.; Haase, G. Coniosporium Perforans and C. apollinis, Two New Rock-Inhabiting Fungi Isolated from Marble in the Sanctuary of Delos (Cyclades, Greece); Kluwer Academic Publishers: Norwell, MA, USA, 1997; Volume 72. [Google Scholar] [CrossRef]
- Gorbushina, A.A.; Krumbein, W.E. Subaerial Microbial Mats and Their Effects on Soil and Rock. In Microbial Sediments; Riding, R.E., Awramik, S.M., Eds.; Springer: Berlin/Heidelberg, Germany, 2000; pp. 161–170. [Google Scholar] [CrossRef]
- De Leo, F.; Antonelli, F.; Pietrini, A.M.; Ricci, S.; Urzì, C. Study of the Euendolithic Activity of Black Meristematic Fungi Isolated from a Marble Statue in the Quirinale Palace’s Gardens in Rome, Italy. Facies 2019, 65, 18. [Google Scholar] [CrossRef]
- Isola, D.; Zucconi, L.; Cecchini, A.; Caneva, G. Dark-Pigmented Biodeteriogenic Fungi in Etruscan Hypogeal Tombs: New Data on Their Culture-Dependent Diversity, Favouring Conditions, and Resistance to Biocidal Treatments. Fungal Biol. 2021, 125, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Isola, D.; Bartoli, F.; Meloni, P.; Caneva, G.; Zucconi, L. Black Fungi and Stone Heritage Conservation: Ecological and Metabolic Assays for Evaluating Colonization Potential and Responses to Traditional Biocides. Appl. Sci. 2022, 12, 2038. [Google Scholar] [CrossRef]
- Sterflinger, K. Fungi: Their Role in Deterioration of Cultural Heritage. Fungal Biol. Rev. 2010, 24, 47–55. [Google Scholar] [CrossRef]
- Isola, D.; Zucconi, L.; Onofri, S.; Caneva, G.; de Hoog, G.S.; Selbmann, L. Extremotolerant Rock Inhabiting Black Fungi from Italian Monumental Sites. Fungal Divers. 2016, 76, 75–96. [Google Scholar] [CrossRef]
- Isola, D.; Selbmann, L.; Meloni, P.; Maracci, E.; Onofri, S.; Zucconi, L. Detrimental Rock Black Fungi and Biocides: A Study on the Monumental Cemetery of Cagliari. In Science and Technology for the Conservation of Cultural Heritage; Rogerio-Candelera, M.A., Lazzari, M., Cano, E., Eds.; CRC Press Taylor & Francis Group: Boca Raton, FL, USA, 2013; pp. 83–86. Available online: https://hdl.handle.net/11584/53815 (accessed on 21 May 2023).
- Isola, D. Biodiversità, Filogenesi Ed Evoluzione Dei Funghi Neri Delle Rocce. Doctoral Thesis, Università degli Studi della Tuscia, Viterbo, Italy, 2010. Available online: http://hdl.handle.net/2067/1068 (accessed on 21 May 2023).
- Egidi, E.; De Hoog, G.S.; Isola, D.; Onofri, S.; Quaedvlieg, W.; De Vries, M.; Verkley, G.J.M.; Stielow, J.B.; Zucconi, L.; Selbmann, L. Phylogeny and Taxonomy of Meristematic Rock-Inhabiting Black Fungi in the Dothideomycetes Based on Multi-Locus Phylogenies. Fungal Divers. 2014, 65, 127–165. [Google Scholar] [CrossRef]
- Liu, B.; Fu, R.; Wu, B.; Liu, X.; Xiang, M. Rock-Inhabiting Fungi: Terminology, Diversity, Evolution and Adaptation Mechanisms. Mycology 2022, 13, 1–31. [Google Scholar] [CrossRef] [PubMed]
- De Leo, F.; Marchetta, A.; Urzì, C. Black Fungi on Stone-Built Heritage: Current Knowledge and Future Outlook. Appl. Sci. 2022, 12, 3969. [Google Scholar] [CrossRef]
- Wollenzien, U.; De Hoog, G.S.; Krumbein, W.; Uijthof, J.M.J. Sarcinomyces petricola, a New Microcolonial Fungus from Marble in the Mediterranean Basin; Kluwer Academic Publishers: Norwell, MA, USA, 1997; Volume 71. [Google Scholar] [CrossRef]
- Sert, H.; Sümbül, H.; Sterflinger, K. Microcolonial Fungi from Antique Marbles in Perge/Side/Termessos (Antalya/Turkey). Antonie Van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2007, 91, 217–227. [Google Scholar] [CrossRef]
- Checcucci, A.; Borruso, L.; Petrocchi, D.; Perito, B. Diversity and Metabolic Profile of the Microbial Communities Inhabiting the Darkened White Marble of Florence Cathedral. Int. Biodeterior. Biodegrad. 2022, 171, 105420. [Google Scholar] [CrossRef]
- Cappitelli, F.; Villa, F. Novel Antibiofilm Non-Biocidal Strategies. In Microorganisms in the Deterioration and Preservation of Cultural Heritage; Springer International Publishing: Cham, Switzerland, 2021. [Google Scholar] [CrossRef]
- Fidanza, M.R.; Caneva, G. Natural Biocides for the Conservation of Stone Cultural Heritage: A Review. J. Cult. Herit. 2019, 38, 271–286. [Google Scholar] [CrossRef]
- Stupar, M.; Grbić, M.L.; Džamić, A.; Unković, N.; Ristić, M.; Jelikić, A.; Vukojević, J. Antifungal Activity of Selected Essential Oils and Biocide Benzalkonium Chloride against the Fungi Isolated from Cultural Heritage Objects. S. Afr. J. Bot. 2014, 93, 118–124. [Google Scholar] [CrossRef]
- Bruno, L.; Rugnini, L.; Spizzichino, V.; Caneve, L.; Canini, A.; Neil, T.; Ellwood, W. Biodeterioration of Roman Hypogea: The Case Study of the Catacombs of SS. Marcellino and Pietro (Rome, Italy). Ann. Microbiol. 2019, 69, 1023–1032. [Google Scholar] [CrossRef]
- Santo, A.P.; Agostini, B.; Cuzman, O.A.; Michelozzi, M.; Salvatici, T.; Perito, B. Essential Oils to Contrast Biodeterioration of the External Marble of Florence Cathedral. Sci. Total Environ. 2023, 877, 162913. [Google Scholar] [CrossRef] [PubMed]
- Santo, A.P.; Agostini, B.; Checcucci, A.; Pecchioni, E.; Perito, B. An Interdisciplinary Study of Biodeterioration of the External Marbles of Santa Maria Del Fiore Cathedral, Florence (IT). In IOP Conference Series: Materials Science and Engineering; IOP Publishing Ltd.: Bristol, UK, 2020; Volume 949. [Google Scholar] [CrossRef]
- Kumar, S.; Nei, M.; Dudley, J.; Tamura, K. MEGA: A Biologist-Centric Software for Evolutionary Analysis of DNA and Protein Sequences. Brief. Bioinform. 2008, 9, 299–306. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The Neighbor-Joining Method: A New Method for Reconstructing Phylogenetic Trees’. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Kimura, M. Possibility of Extensive Neutral Evolution under Stabilizing Selection with Special Reference to Nonrandom Usage of Synonymous Codons. Genetics 1981, 78, 5773–5777. [Google Scholar] [CrossRef]
- Trovão, J.; Portugal, A. Current Knowledge on the Fungal Degradation Abilities Profiled through Biodeteriorative Plate Essays. Appl. Sci. 2021, 11, 4196. [Google Scholar] [CrossRef]
- Jurado, V.; Gonzalez-Pimentel, J.L.; Hermosin, B.; Saiz-Himenez, C. Biodeterioration of Salón de Reinos, Museo Nacional Del Prado, Madrid, Spain. Appl. Sci. 2021, 11, 8858. [Google Scholar] [CrossRef]
- Selbmann, L.; Isola, D.; Egidi, E.; Zucconi, L.; Gueidan, C.; De Hoog, G.S.; Onofri, S. Mountain Tips as Reservoirs for New Rock-Fungal Entities: Saxomyces Gen. Nov. and Four New Species from the Alps. Fungal Divers. 2014, 65, 167–182. [Google Scholar] [CrossRef]
- Sterflinger, K. Fungi as Geologic Agents. Geomicrobiol. J. 2000, 17, 97–124. [Google Scholar] [CrossRef]
- Gerrits, R.; Pokharel, R.; Breitenbach, R.; Radnik, J.; Feldmann, I.; Schuessler, J.A.; von Blanckenburg, F.; Gorbushina, A.A.; Schott, J. How the Rock-Inhabiting Fungus K. Petricola A95 Enhances Olivine Dissolution through Attachment. Geochim. Cosmochim. Acta 2020, 282, 76–97. [Google Scholar] [CrossRef]
- Tonon, C.; Breitenbach, R.; Voigt, O.; Turci, F.; Gorbushina, A.A.; Favero-Longo, S.E. Hyphal Morphology and Substrate Porosity—Rather than Melanization- Drive Penetration of Black Fungi into Carbonate Substrates. J. Cult. Herit. 2021, 48, 244–253. [Google Scholar] [CrossRef]
- Caldeira, A.T. Green Mitigation Strategy for Cultural Heritage Using Bacterial Biocides. In Microorganisms in the Deterioration and Preservation of Cultural Heritage; Edith, J., Ed.; Springer: Berlin/Heidelberg, Germany, 2021; p. 137. [Google Scholar] [CrossRef]
- Gostinčar, C.; Grube, M.; Gunde-Cimerman, N. Evolution of Fungal Pathogens in Domestic Environments? Fungal Biol. 2011, 115, 1008–1018. [Google Scholar] [CrossRef]
- Saad, N.Y.; Muller, C.D.; Lobstein, A. Major Bioactivities and Mechanism of Action of Essential Oils and Their Components. Flavour Fragr. J. 2013, 28, 269–279. [Google Scholar] [CrossRef]
- Algburi, A.; Comito, N.; Kashtanov, D.; Dicks, L.M.T.; Chikindas, M.L. Control of Biofilm Formation: Antibiotics and Beyond. Appl. Environ. Microbiol. 2017, 83, e02508-16. [Google Scholar] [CrossRef]
- Napoli, E.; Di Vito, M. Toward a New Future for Essential Oils. Antibiotics 2021, 10, 207. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Area | Strain | Organism with the Best ITS Match (Accession Number) | Identity (%) | Organism with the Best LSU Match (Accession Number) | Identity (%) |
|---|---|---|---|---|---|
| NW | TW-N-Tq | Dothideomycetes sp. (OQ319946) | 99.45 | Dothideomycetes sp. (OQ302485) | 96.12 |
| M1 | Lithohypha guttulata (MW361305) | 100 | Lithohypha aloicola (OQ302490) | 100 | |
| M7 | Coniosporium uncinatum (OQ319953) | 98.33 | Coniosporium uncinatum (OQ302491) | 100 | |
| M25 | Vermiconia calcicola (OQ319958) | 98.20 | Vermiconia calcicola (OQ302495) | 100 | |
| M30 | Coniosporium uncinatum (OQ319954) | 99.24 | Coniosporium uncinatum (OQ302493) | 100 | |
| M32 | Paradevriesia pseudoamericana (OQ319966) | 95.94 | Paradevriesia americana (OQ302498) | 99.27 | |
| M34 | Lithohypha guttulata (OQ319948) | 99.46 | Lithohypha aloicola (OQ302486) | 100 | |
| m15 | Knufia marmoricola (OQ319960) | 99.58 | Knufia marmoricola (OQ302499) | 100 | |
| m16 | Knufia marmoricola (OQ319964) | 100 | Knufia marmoricola (OQ302500) | 99.65 | |
| m17 | Knufia marmoricola (OQ319961) | 100 | Knufia marmoricola (OQ302501) | 100 | |
| m20 | Lithohypha guttulata (OQ319952) | 100 | Coniosporium uncinatum (OQ302505) | 99.52 | |
| m21 | Lithohypha guttulata (OQ319947) | 99.81 | Lithohypha guttulata (OQ302504) | 100 | |
| SE | NT-S-5 * | Dothideomycetes sp. (MW361320) | 99.45 | Dothideomycetes sp. (OQ302484) | 99.66 |
| M35 | Lithohypha sp. (OQ319956) | 99.44 | Lithohypha guttulata (OQ302487) | 99.65 | |
| M36 | Lithohypha guttulata (OQ319949) | 99.81 | Lithohypha aloicola (OQ302488) | 99.83 | |
| M37 | Knufia marmoricola (OQ319965) | 99.15 | Knufia marmoricola (OQ302497) | 100 | |
| M38 | Coniosporium uncinatum (OQ319955) | 98.52 | Coniosporium uncinatum (OQ302492) | 100 | |
| M40 | Vermiconia calcicola (OQ319957) | 100 | Vermiconia calcicola (OQ302494) | 100 | |
| M41 | Lithohypha guttulata (OQ319950) | 99.82 | Lithohypha guttulata (OQ302489) | 100 | |
| M43 | Knufia petricola (OQ319962) | 100 | Knufia marmoricola (OQ302496) | 100 | |
| m8 | Knufia marmoricola (OQ319963) | 99.60 | Knufia sp. (OQ302502) | 99.65 | |
| m10 | Vermiconia calcicola (OQ319967) | 100 | Vermiconia calcicola (OQ302506) | 99.65 | |
| m11 | Vermiconia calcicola (OQ319959) | 99.18 | Vermiconia calcicola (OQ302507) | 99.82 | |
| m13 | Lithohypha guttulata (OQ319951) | 99.81 | Lithohypha guttulata (OQ302503) | 99.82 |
| Growth after 28 Days (mm ± SD) | |||||
|---|---|---|---|---|---|
| Strain | Genus | 5 °C | 15 °C | 20 °C | 30 °C * |
| m8 | Knufia sp. | 7.83 ± 0.98 | 9.75 ± 1.78 | 7.25 ± 2.53 | 1.38 ± 0.48 |
| m10 | Vermiconia sp. | 3.92 ± 0.66 | 3.92 ± 0.38 | 5.02 ± 1.54 | 4.38 ± 0.75 |
| m15 | Knufia sp. | 7.17 ± 0.93 | 8.33 ± 0.52 | 10.42 ± 1.20 | 4.83 ± 1.59 |
| m16 | Knufia sp. | 8.50 ± 2.05 | 8.00 ± 0.71 | 13.00 ± 1.52 | 4.88 ± 1.31 |
| m20 | Lithohypha sp. | 5.38 ± 1.46 | 4.95 ± 1.25 | 5.67 ± 1.17 | 3.88 ± 0.63 |
| m21 | Lithohypha sp. | 6.13 ± 0.43 | 6.33 ± 1.51 | 14.00 ± 1.63 | 5.50 ± 2.12 |
| M30 | Coniosporium sp. | 4.08 ± 1.20 | 5.17 ± 1.47 | 7.08 ± 0.92 | 4.50 ± 1.08 |
| TW-N-Tq | Dothideomycetes sp. | 6.67 ± 1.94 | 8.58 ± 2.42 | 9.17 ± 1.69 | 9.50 ± 2.68 |
| Growth after 28 Days (mm ± SD) | ||||
|---|---|---|---|---|
| Strain | Genus | Control | 5% NaCl * | % Decrease in Growth |
| m8 | Knufia sp. | 8.75 ± 2.83 | 1.92 ± 0.49 | 78.1 |
| m10 | Vermiconia sp. | 5.30 ± 1.56 | 4.08 ± 1.02 | 22.9 |
| m15 | Knufia sp. | 11.75 ± 1.50 | 1.92 ± 0.92 | 83.7 |
| m16 | Knufia sp. | 9.25 ± 0.80 | 1.92 ± 0.66 | 79.3 |
| m20 | Lithohypha sp. | 5.00 ± 1.00 | 2.17 ± 0.52 | 56.7 |
| m21 | Lithohypha sp. | 9.25 ± 0.63 | 0.00 ± 0.00 | 100.0 |
| M30 | Coniosporium sp. | 9.00 ± 1.56 | 1.63 ± 0.75 | 81.9 |
| TW-N-Tq | Dothideomycetes sp. | 8.50 ± 1.42 | 0.00 | 100.0 |
| % Decrease in Growth | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EO-Origanum (% v/v) | EO-Thymus (% v/v) | BiotinT (% v/v) | ||||||||||
| Strain | 0.0025% | 0.005% | 0.012% | 0.025% | 0.0025% | 0.005% | 0.012% | 0.025% | 0.0025% | 0.005% | 0.012% | 0.025% |
| m8 | 42 | 45 | 100 | 100 | 33 | 8 | 59 | 100 | 69 | 68 | 81 | 77 |
| m10 | 0 | 0 | 77 | 100 | 0 | 0 | 0 | 79 | 0 | 0 | 12 | 26 |
| m15 | 51 | 55 | 100 | 100 | 37 | 32 | 64 | 100 | 32 | 79 | 77 | 94 |
| m16 | 25 | 28 | 100 | 100 | 0 | 12 | 44 | 100 | 28 | 40 | 72 | 72 |
| m20 | 53 | 66 | 92 | 100 | 36 | 54 | 58 | 91 | 54 | 54 | 56 | 56 |
| m21 | 0 | 0 | 56 | 100 | 0 | 4 | 2 | 57 | 6 | 18 | 31 | 35 |
| M30 | 4 | 12 | 100 | 100 | 0 | 4 | 39 | 100 | 0 | 0 | 20 | 33 |
| TW-N-Tq | 38 | 43 | 100 | 100 | 20 | 10 | 57 | 100 | 37 | 45 | 75 | 98 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Berti, L.; Marvasi, M.; Perito, B. Characterization of the Community of Black Meristematic Fungi Inhabiting the External White Marble of the Florence Cathedral. J. Fungi 2023, 9, 665. https://doi.org/10.3390/jof9060665
Berti L, Marvasi M, Perito B. Characterization of the Community of Black Meristematic Fungi Inhabiting the External White Marble of the Florence Cathedral. Journal of Fungi. 2023; 9(6):665. https://doi.org/10.3390/jof9060665
Chicago/Turabian StyleBerti, Letizia, Massimiliano Marvasi, and Brunella Perito. 2023. "Characterization of the Community of Black Meristematic Fungi Inhabiting the External White Marble of the Florence Cathedral" Journal of Fungi 9, no. 6: 665. https://doi.org/10.3390/jof9060665
APA StyleBerti, L., Marvasi, M., & Perito, B. (2023). Characterization of the Community of Black Meristematic Fungi Inhabiting the External White Marble of the Florence Cathedral. Journal of Fungi, 9(6), 665. https://doi.org/10.3390/jof9060665