
Field-Crop Soils in Eastern France: Coldspots of Azole-Resistant Aspergillus fumigatus
 ,
,  , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Environmental Sampling
2.2. Physicochemical Analysis of Soils
2.3. Analysis of Soil Fungicide Levels
2.3.1. Extraction and Analysis
2.3.2. Experiment Validation
2.4. Isolation and Identification of A. fumigatus from Soils
2.5. Assessment of Fungal Load by qPCR
2.6. Statistical Analysis
3. Results
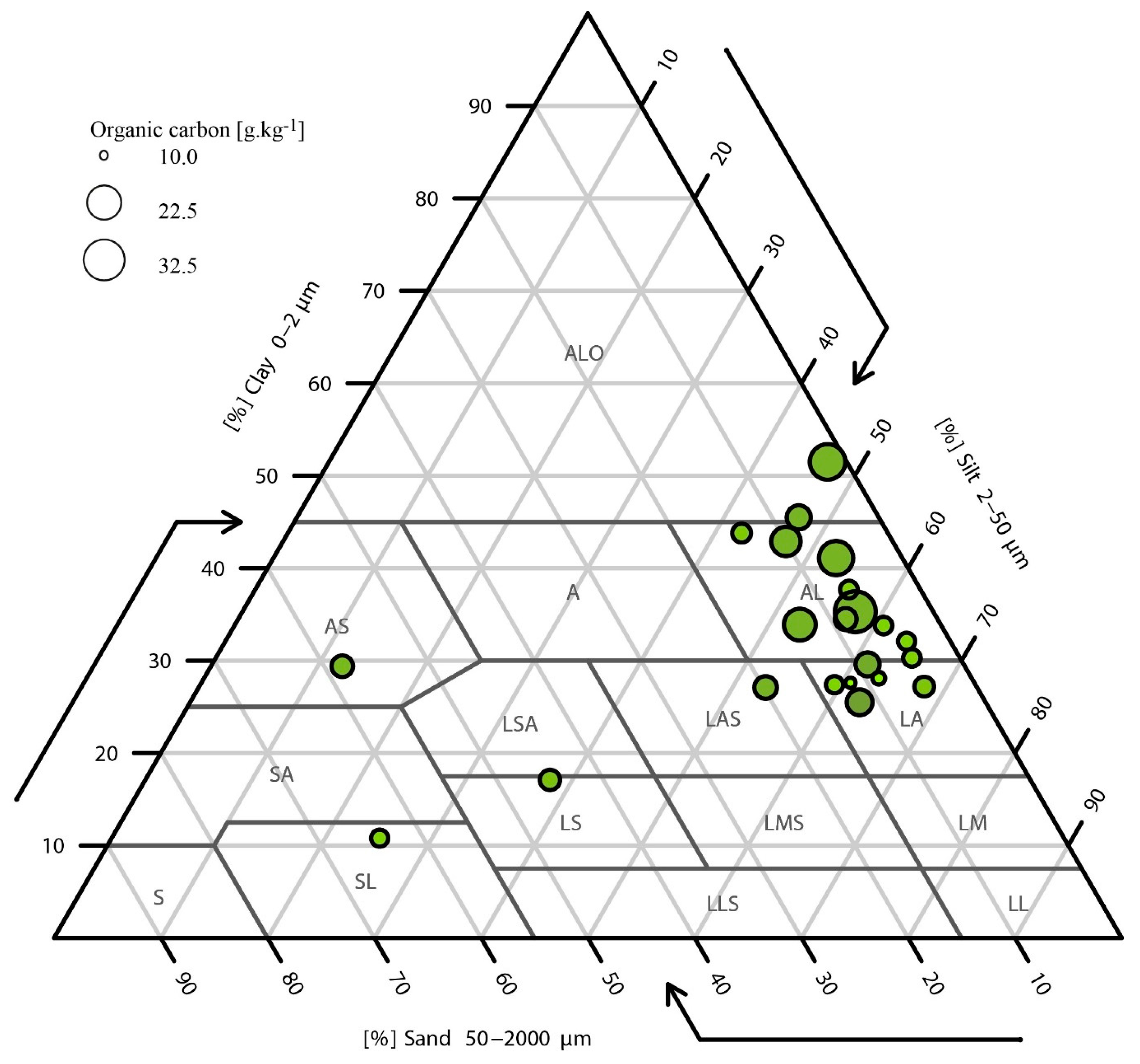
3.1. Physicochemical and Fungicide Analyses of Soils
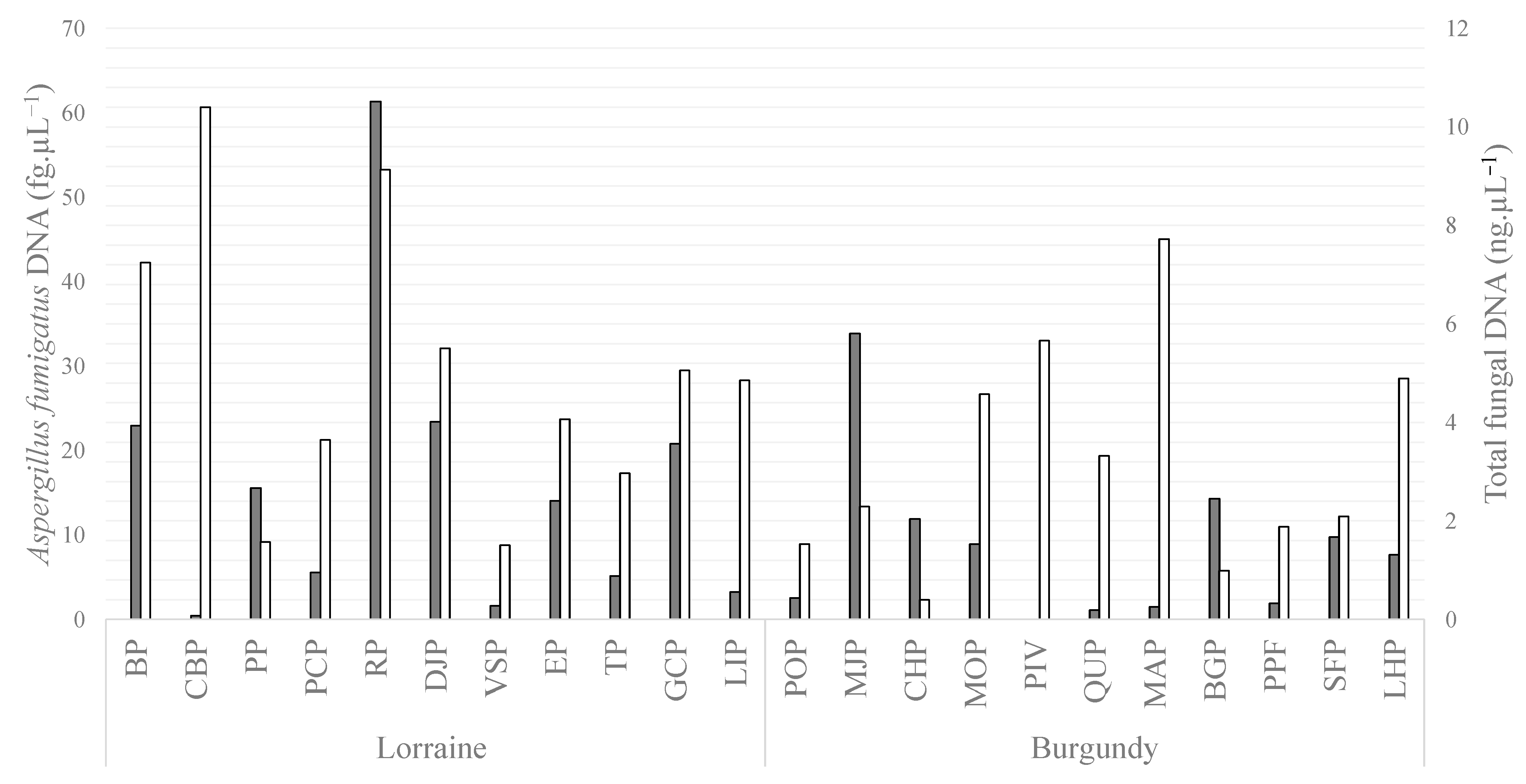
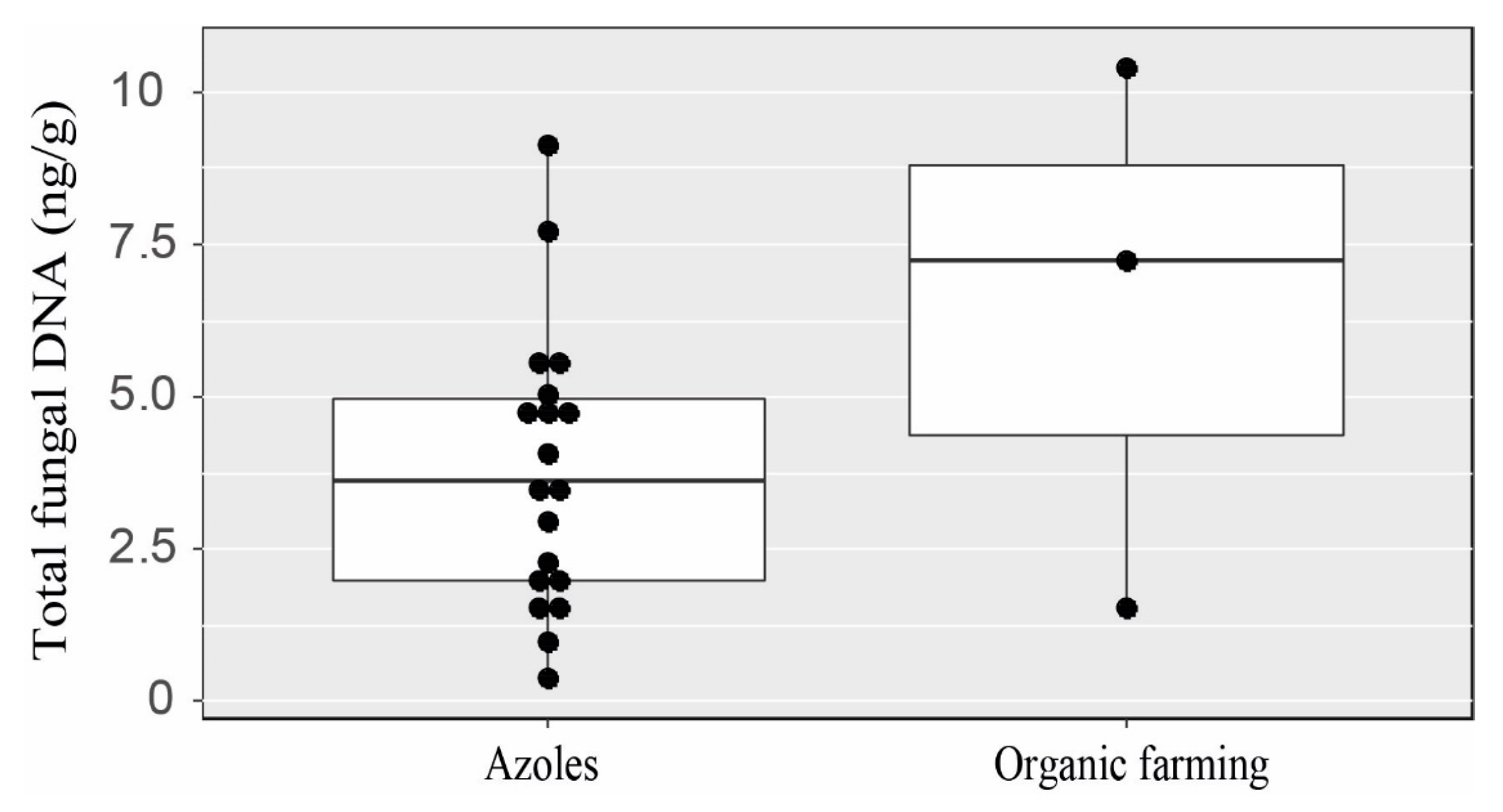
3.2. Fungal Analysis of Soils
3.3. Characterisation of the Fungal Environment
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chowdhary, A.; Meis, J.F. Emergence of azole resistant Aspergillus fumigatus and One Health: Time to implement environmental stewardship. Environ. Microbiol. 2018, 20, 1299–1301. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, J.C. Aspergillus fumigatus: Growth and virulence. Med. Mycol. 2006, 44 (Suppl. S1), S77–S81. [Google Scholar] [CrossRef] [PubMed]
- O’Gorman, C.M. Airborne Aspergillus fumigatus conidia: A risk factor for aspergillosis. Fungal Biol. Rev. 2011, 25, 151–157. [Google Scholar] [CrossRef]
- Steinmann, J.; Hamprecht, A.; Vehreschild, M.J.; Cornely, O.A.; Buchheidt, D.; Spiess, B.; Koldehoff, M.; Buer, J.; Meis, J.F.; Rath, P.M. Emergence of azole-resistant invasive aspergillosis in HSCT recipients in Germany. J. Antimicrob. Chemother. 2015, 70, 1522–1526. [Google Scholar] [CrossRef] [PubMed]
- Snelders, E.; van der Lee, H.A.L.; Kuijpers, J.; Rijs, A.J.M.M.; Varga, J.; Samson, R.A.; Mellado, E.; Donders, A.R.T.; Melchers, W.J.G.; Verweij, P.E. Emergence of Azole Resistance in Aspergillus fumigatus and Spread of a Single Resistance Mechanism. PLoS Med. 2008, 5, e219. [Google Scholar] [CrossRef] [PubMed]
- Verweij, P.E.; Ananda-Rajah, M.; Andes, D.; Arendrup, M.C.; Bruggemann, R.J.; Chowdhary, A.; Cornely, O.A.; Denning, D.W.; Groll, A.H.; Izumikawa, K.; et al. International expert opinion on the management of infection caused by azole-resistant Aspergillus fumigatus. Drug Resist. Updat. 2015, 21–22, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Rubio, R.; Gonzalez-Jimenez, I.; Lucio, J.; Mellado, E. Characterization of Aspergillus fumigatus cross-resistance between clinical and DMI azole drugs. Appl. Environ. Microbiol. 2021, 87, e02539-20. [Google Scholar] [CrossRef] [PubMed]
- Snelders, E.; Camps, S.M.T.; Karawajczyk, A.; Schaftenaar, G.; Kema, G.H.J.; van der Lee, H.A.; Klaassen, C.H.; Melchers, W.J.G.; Verweij, P.E. Triazole Fungicides Can Induce Cross-Resistance to Medical Triazoles in Aspergillus fumigatus. PLoS ONE 2012, 7, e31801. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, J.; Abdolrasouli, A.; Dunne, K.; Sewell, T.R.; Zhang, Y.; Ballard, E.; Brackin, A.P.; van Rhijn, N.; Chown, H.; Tsitsopoulou, A.; et al. Population genomics confirms acquisition of drug-resistant Aspergillus fumigatus infection by humans from the environment. Nat. Microbiol. 2022, 7, 663–674. [Google Scholar] [CrossRef] [PubMed]
- Hagiwara, D.; Watanabe, A.; Kamei, K.; Goldman, G.H. Epidemiological and Genomic Landscape of Azole Resistance Mechanisms in Aspergillus Fungi. Front. Microbiol. 2016, 7, 1382. [Google Scholar] [CrossRef] [PubMed]
- Meneau, I.; Coste, A.T.; Sanglard, D. Identification of Aspergillus fumigatus multidrug transporter genes and their potential involvement in antifungal resistance. Med. Mycol. 2016, 54, 616–627. [Google Scholar] [CrossRef]
- Berger, S.; El Chazli, Y.; Babu, A.F.; Coste, A.T. Azole Resistance in Aspergillus fumigatus: A Consequence of Antifungal Use in Agriculture? Front. Microbiol. 2017, 8, 1024. [Google Scholar] [CrossRef]
- Schoustra, S.E.; Debets, A.J.M.; Rijs, A.; Zhang, J.; Snelders, E.; Leendertse, P.C.; Melchers, W.J.G.; Rietveld, A.G.; Zwaan, B.J.; Verweij, P.E. Environmental Hotspots for Azole Resistance Selection of Aspergillus fumigatus, the Netherlands. Emerg. Infect. Dis. 2019, 25, 1347–1353. [Google Scholar] [CrossRef]
- Fraaije, B.; Atkins, S.; Hanley, S.; Macdonald, A.; Lucas, J. The Multi-Fungicide Resistance Status of Aspergillus fumigatus Populations in Arable Soils and the Wider European Environment. Front. Microbiol. 2020, 11, 599233. [Google Scholar] [CrossRef]
- Rocchi, S.; Godeau, C.; Crini, G.; Snelders, E. Emergence of a Pathogenic Fungus Resistant to Triazole Antifungal Drugs. In Emerging Contaminants Vol. 1: Occurrence and Impact; Morin-Crini, N., Lichtfouse, E., Crini, G., Eds.; Springer International Publishing: Cham, Switerland, 2021; pp. 165–206. [Google Scholar]
- Sewell, T.R.; Zhang, Y.; Brackin, A.P.; Shelton, J.M.G.; Rhodes, J.; Fisher, M.C. Elevated prevalence of azole resistant Aspergillus fumigatus in urban versus rural environments in the United Kingdom. Antimicrob. Agents Chemother. 2019, 63, e00548-19. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Lopez Jimenez, L.; Snelders, E.; Debets, A.J.M.; Rietveld, A.G.; Zwaan, B.J.; Verweij, P.E.; Schoustra, S.E. Dynamics of Aspergillus fumigatus in Azole Fungicide-Containing Plant Waste in the Netherlands (2016–2017). Appl. Environ. Microbiol. 2021, 87, e02295-20. [Google Scholar] [CrossRef]
- Barber, A.E.; Riedel, J.; Sae-Ong, T.; Kang, K.; Brabetz, W.; Panagiotou, G.; Deising, H.B.; Kurzai, O. Effects of Agricultural Fungicide Use on Aspergillus fumigatus Abundance, Antifungal Susceptibility, and Population Structure. mBio 2020, 11, e02213-20. [Google Scholar] [CrossRef]
- Moeys, J. Soiltexture: Functions for Soil Texture Plot, Classification and Transformation, R Package version 1.5.1. 2018. Available online: https://rdrr.io/cran/soiltexture/ (accessed on 29 March 2023).
- Anastassiades, M.; Mastovská, K.; Lehotay, S.J. Evaluation of analyte protectants to improve gas chromatographic analysis of pesticides. J. Chromatogr. A 2003, 1015, 163–184. [Google Scholar] [CrossRef]
- Van der Linden, J.W.; Camps, S.M.; Kampinga, G.A.; Arends, J.P.; Debets-Ossenkopp, Y.J.; Haas, P.J.; Rijnders, B.J.; Kuijper, E.J.; van Tiel, F.H.; Varga, J.; et al. Aspergillosis due to voriconazole highly resistant Aspergillus fumigatus and recovery of genetically related resistant isolates from domiciles. Clin. Infect. Dis. 2013, 57, 513–520. [Google Scholar] [CrossRef]
- Bader, O.; Tunnermann, J.; Dudakova, A.; Tangwattanachuleeporn, M.; Weig, M.; Gross, U. Environmental Isolates of Azole-Resistant Aspergillus fumigatus in Germany. Antimicrob. Agents Chemother. 2015, 59, 4356–4359. [Google Scholar] [CrossRef] [PubMed]
- Lamoth, F. Aspergillus fumigatus-Related Species in Clinical Practice. Front. Microbiol. 2016, 7, 683. [Google Scholar] [CrossRef] [PubMed]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [PubMed]
- Guinea, J. Updated EUCAST Clinical Breakpoints against Aspergillus, Implications for the Clinical Microbiology Laboratory. J. Fungi 2020, 6, 343. [Google Scholar] [CrossRef]
- Haugland, R.A.; Varma, M.; Wymer, L.J.; Vesper, S.J. Quantitative PCR Analysis of Selected Aspergillus, Penicillium and Paecilomyces Species. Syst. Appl. Microbiol. 2004, 27, 198–210. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.M.; Kachur, S.; Dwan, M.G.; Abraham, A.G.; Aziz, M.; Hsueh, P.R.; Huang, Y.T.; Busch, J.D.; Lamit, L.J.; Gehring, C.A.; et al. FungiQuant: A broad-coverage fungal quantitative real-time PCR assay. BMC Microbiol. 2012, 12, 255. [Google Scholar] [CrossRef] [PubMed]
- Knapp, J.; Umhang, G.; Poulle, M.L.; Millon, L. Development of a Real-Time PCR for a Sensitive One-Step Coprodiagnosis Allowing both the Identification of Carnivore Feces and the Detection of Toxocara spp. and Echinococcus multilocularis. Appl. Environ. Microbiol. 2016, 82, 2950–2958. [Google Scholar] [CrossRef] [PubMed]
- Godeau, C.; Reboux, G.; Scherer, E.; Laboissiere, A.; Lechenault-Bergerot, C.; Millon, L.; Rocchi, S. Azole-resistant Aspergillus fumigatus in the hospital: Surveillance from flower beds to corridors. Am. J. Infect. Control 2020, 48, 702–704. [Google Scholar] [CrossRef] [PubMed]
- Komarek, M.; Cadkova, E.; Chrastny, V.; Bordas, F.; Bollinger, J.C. Contamination of vineyard soils with fungicides: A review of environmental and toxicological aspects. Environ. Int. 2010, 36, 138–151. [Google Scholar] [CrossRef] [PubMed]
- Čadková, E.; Komárek, M.; Kaliszová, R.; Vaněk, A.; Balíková, M. Tebuconazole Sorption in Contrasting Soil Types. Soil Sediment Contam. Int. J. 2013, 22, 404–414. [Google Scholar] [CrossRef]
- Sun, T.; Miao, J.; Saleem, M.; Zhang, H.; Yang, Y.; Zhang, Q. Bacterial compatibility and immobilization with biochar improved tebuconazole degradation, soil microbiome composition and functioning. J. Hazard. Mater. 2020, 398, 122941. [Google Scholar] [CrossRef]
- Meite, F.; Granet, M.; Imfeld, G. Ageing of copper, zinc and synthetic pesticides in particle-size and chemical fractions of agricultural soils. Sci. Total Environ. 2022, 824, 153860. [Google Scholar] [CrossRef] [PubMed]
- Rocchi, S.; Poncot, M.; Morin-Crini, N.; Laboissiere, A.; Valot, B.; Godeau, C.; Lechenault-Bergerot, C.; Reboux, G.; Crini, G.; Millon, L. Determination of azole fungal residues in soils and detection of Aspergillus fumigatus-resistant strains in market gardens of Eastern France. Environ. Sci. Pollut. Res. Int. 2018, 25, 32015–32023. [Google Scholar] [CrossRef]
- Alvarez-Moreno, C.; Lavergne, R.A.; Hagen, F.; Morio, F.; Meis, J.F.; Le Pape, P. Azole-resistant Aspergillus fumigatus harboring TR(34)/L98H, TR(46)/Y121F/T289A and TR(53) mutations related to flower fields in Colombia. Sci. Rep. 2017, 7, 45631. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Moreno, C.; Lavergne, R.A.; Hagen, F.; Morio, F.; Meis, J.F.; Le Pape, P. Fungicide-driven alterations in azole-resistant Aspergillus fumigatus are related to vegetable crops in Colombia, South America. Mycologia 2019, 111, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.J.; Wang, H.C.; Lee, J.C.; Lo, H.J.; Dai, C.T.; Chou, P.H.; Ko, W.C.; Chen, Y.C. Azole-resistant Aspergillus fumigatus isolates carrying TR(3)(4)/L98H mutations in Taiwan. Mycoses 2015, 58, 544–549. [Google Scholar] [CrossRef] [PubMed]
- Jeanvoine, A.; Rocchi, S.; Reboux, G.; Crini, N.; Crini, G.; Millon, L. Azole-resistant Aspergillus fumigatus in sawmills of Eastern France. J. Appl. Microbiol. 2017, 123, 172–184. [Google Scholar] [CrossRef]
- Silva, V.; Mol, H.G.J.; Zomer, P.; Tienstra, M.; Ritsema, C.J.; Geissen, V. Pesticide residues in European agricultural soils—A hidden reality unfolded. Sci. Total Environ. 2019, 653, 1532–1545. [Google Scholar] [CrossRef]
- Pelosi, C.; Bertrand, C.; Daniele, G.; Coeurdassier, M.; Benoit, P.; Nélieu, S.; Lafay, F.; Bretagnolle, V.; Gaba, S.; Vulliet, E.; et al. Residues of currently used pesticides in soils and earthworms: A silent threat? Agric. Ecosyst. Environ. 2021, 305, 107167. [Google Scholar] [CrossRef]
- Vaezi, A.; Fakhim, H.; Javidnia, J.; Khodavaisy, S.; Abtahian, Z.; Vojoodi, M.; Nourbakhsh, F.; Badali, H. Pesticide behavior in paddy fields and development of azole-resistant Aspergillus fumigatus: Should we be concerned? J. Mycol. Med. 2018, 28, 59–64. [Google Scholar] [CrossRef]
- Lavergne, R.A.; Chouaki, T.; Hagen, F.; Toublanc, B.; Dupont, H.; Jounieaux, V.; Meis, J.F.; Morio, F.; Le Pape, P. Home Environment as a Source of Life-Threatening Azole-Resistant Aspergillus fumigatus in Immunocompromised Patients. Clin. Infect. Dis. 2017, 64, 76–78. [Google Scholar] [CrossRef]
- Rocchi, S.; Daguindau, E.; Grenouillet, F.; Deconinck, E.; Bellanger, A.P.; Garcia-Hermoso, D.; Bretagne, S.; Reboux, G.; Millon, L. Azole-resistant Aspergillus fumigatus isolate with the TR34/L98H mutation in both a fungicide-sprayed field and the lung of a hematopoietic stem cell transplant recipient with invasive aspergillosis. J. Clin. Microbiol. 2014, 52, 1724–1726. [Google Scholar] [CrossRef]
- Chowdhary, A.; Kathuria, S.; Randhawa, H.S.; Gaur, S.N.; Klaassen, C.H.; Meis, J.F. Isolation of multiple-triazole-resistant Aspergillus fumigatus strains carrying the TR/L98H mutations in the cyp51A gene in India. J. Antimicrob. Chemother. 2012, 67, 362–366. [Google Scholar] [CrossRef] [PubMed]
- Cao, D.; Wang, F.; Yu, S.; Dong, S.; Wu, R.; Cui, N.; Ren, J.; Xu, T.; Wang, S.; Wang, M.; et al. Prevalence of Azole-Resistant Aspergillus fumigatus is Highly Associated with Azole Fungicide Residues in the Fields. Environ. Sci. Technol. 2021, 55, 3041–3049. [Google Scholar] [CrossRef] [PubMed]
- Mortensen, K.L.; Mellado, E.; Lass-Flörl, C.; Rodriguez-Tudela, J.L.; Johansen, H.K.; Arendrup, M.C. Environmental study of azole-resistant Aspergillus fumigatus and other aspergilli in Austria, Denmark, and Spain. Antimicrob. Agents Chemother. 2010, 54, 4545–4549. [Google Scholar] [CrossRef] [PubMed]
- Prigitano, A.; Venier, V.; Cogliati, M.; De Lorenzis, G.; Esposto, M.C.; Tortorano, A.M. Azole-resistant Aspergillus fumigatus in the environment of northern Italy, May 2011 to June 2012. Eurosurveillance 2014, 19, 20747. [Google Scholar] [CrossRef]
- Jeanvoine, A.; Rocchi, S.; Reboux, G.; Grenouillet, F.; Benassarou, M.; Chirouze, C.; Millon, L. Sinus aspergillosis due to an azole-resistant Aspergillus fumigatus strain carrying the TR34/L98H mutation in immunocompetent host. Infect. Dis. 2016, 48, 765–766. [Google Scholar] [CrossRef] [PubMed]
- Zoran, T.; Sartori, B.; Sappl, L.; Aigner, M.; Sánchez-Reus, F.; Rezusta, A.; Chowdhary, A.; Taj-Aldeen, S.J.; Arendrup, M.C.; Oliveri, S.; et al. Corrigendum: Azole-Resistance in Aspergillus terreus and Related Species: An Emerging Problem or a Rare Phenomenon? Front. Microbiol. 2019, 9, 03245. [Google Scholar] [CrossRef]
- Zhang, W.; Xu, J.; Dong, F.; Liu, X.; Zhang, Y.; Wu, X.; Zheng, Y. Effect of tetraconazole application on the soil microbial community. Environ. Sci. Pollut. Res. 2014, 21, 8323–8332. [Google Scholar] [CrossRef]
- Rocchi, S.; Godeau, C.; Scherer, E.; Reboux, G.; Millon, L. One year later: The effect of changing azole-treated bulbs for organic tulips bulbs in hospital environment on the azole-resistant Aspergillus fumigatus rate. Med. Mycol. 2021, 59, 741–743. [Google Scholar] [CrossRef]
- Burks, C.; Darby, A.; Gómez Londoño, L.; Momany, M.; Brewer, M.T. Azole-resistant Aspergillus fumigatus in the environment: Identifying key reservoirs and hotspots of antifungal resistance. PLoS Pathog. 2021, 17, e1009711. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Structural Analysis | Chemical Analysis | Mycological Analysis | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Plot ID | Crop | Area (ha) | Azole Molecules Sprayed over the Two Years before Sampling | OM (%) | pH | Silt (%) | Sand (%) | Clay (%) | TBC (ng/g) | EPC (ng/g) | Aspergillus fumigatus/g of Soil | |
| Lorraine | BP | wheat | 3.0 | Organic farming | 6.2 | 7.9 | 56.9 | 8.6 | 34.5 | 6.0 | <QL | 17 |
| CBP | wheat | 3.1 | Organic farming | 5.6 | 5.9 | 53.1 | 19.8 | 27.1 | 6.2 | <QL | 0 | |
| PP | wheat | 20.5 | TBC, PTC, MTC | 7.3 | 7.2 | 47.1 | 10 | 42.9 | 7.1 | 4.2 | 0 | |
| PCP | wheat | 9.3 | TBC, PTC, MTC | 4.5 | 5.9 | 37.9 | 45 | 17.1 | 7.1 | <QL | 17 | |
| RP | wheat | 12.6 | TBC, PTC, MTC | 6.3 | 6.3 | 61.4 | 9 | 29.6 | 8.1 | 4.6 | 17 | |
| DJP | wheat | 22.6 | TBC, PTC, MTC | 5.5 | 5.9 | 65.2 | 4.5 | 30.3 | 19.1 | <QL | 467 | |
| VSP | wheat | 7.9 | TBC, PTC, MTC | 3.7 | 5.2 | 63.8 | 4.1 | 32.1 | 7.0 | 3.0 | 367 | |
| EP | wheat | 7.2 | TBC, PTC, MTC | 5.1 | 6.2 | 67.9 | 4.9 | 27.2 | 6.6 | 3.4 | 0 | |
| TP | wheat | 1.5 | TBC, PTC, MTC | 10.1 | 6.8 | 46.7 | 1.8 | 51.5 | 6.2 | <QL | 150 | |
| GCP | wheat | 12.1 | TBC, PTC | 7.6 | 7.9 | 47 | 7.5 | 45.5 | 6.0 | 3.2 | 17 | |
| LIP | wheat | 7.3 | TBC, PTC, MTC | 4.9 | 6.6 | 25.1 | 64.1 | 10.8 | 7.0 | <QL | 33 | |
| Burgundy | POP | fallow | 16.3 | Organic farming | 4.8 | 7.6 | 55.6 | 6.7 | 37.7 | 5.6 | <QL | 83 |
| MJP | maize | 4.4 | No azole | 5.1 | 8 | 59.4 | 13.2 | 27.4 | 5.7 | <QL | 67 | |
| CHP | maize | 3.0 | No azole | 3.4 | 6.8 | 60.8 | 11.6 | 27.6 | 5.9 | <QL | 17 | |
| MOP | maize | 8.9 | No azole | 6.9 | 7.8 | 62.7 | 11.8 | 25.5 | 5.5 | <QL | 433 | |
| PIV | wheat | 16.0 | PTC | 4.3 | 6.9 | 63.2 | 8.7 | 28.1 | 5.9 | <QL | 0 | |
| QUP | wheat | 12.0 | TBC, PTC, DFC | 5.8 | 7.9 | 42.5 | 13.7 | 43.8 | 11.0 | <QL | 100 | |
| MAP | wheat | 6.4 | TBC, PTC, DFC | 5.1 | 6.5 | 60.8 | 5.4 | 33.8 | 10.7 | <QL | 50 | |
| BGP | wheat | 2.9 | TBC, PTC | 9.1 | 7.9 | 57.4 | 7.3 | 35.3 | 9.8 | <QL | 50 | |
| PPF | oilseed rape | 7.4 | MTC, PTC (90 g/L) | 7.4 | 7.6 | 52.7 | 6.2 | 41.1 | 8.1 | <QL | 0 | |
| SFP | oilseed rape | 16.3 | MTC, PTC | 7.7 | 7.7 | 52.9 | 13.2 | 33.9 | 12.2 | <QL | 600 | |
| LHP | oilseed rape | 11.5 | MTC, PTC | 7.6 | 7.7 | 12.3 | 58.3 | 29.4 | 14.7 | <QL | 83 | |
| Samples | Number of ARAf per Sample (Culture Media) | Aspergillus fumigatus qPCR, Mean Quantity (ng/µL) | Pan Fungal qPCR, Mean Quantity (ng/µL) |
|---|---|---|---|
| A22 | 0 | 0.000959 | 270.5 |
| D3 | 1 | 0.0246 | 74.85 |
| D9 | 3 | 0.05935 | 44.1 |
| D2 | 4 | 0.07415 | 49.3 |
| D10 | 8 | 0.0943 | 98.1 |
| A27 | 5 | 0.10265 | 113 |
| D8 | 8 | 0.1175 | 203.5 |
| median quantity (qPCR) | 0.07415 | 98.1 | |
| mean quantity (qPCR) | 0.0676441 | 121.9 | |
| minimal quantity (qPCR) | 0.000959 | 44.1 | |
| maximal quantity (qPCR) | 0.1175 | 270.5 | |
| Determination Coefficient | QL (ng/g) | RSD (%) | Recovery (%) | |
|---|---|---|---|---|
| Propiconazole | 0.9987 | 9.0 | 17 | 102 |
| Tebuconazole | 0.9981 | 2.7 | 16 | 107 |
| Epoxiconazole | 0.9985 | 2.5 | 20 | 91 |
| Bromuconazole | 0.9983 | 7.6 | 21 | 94 |
| Difenoconazole | 0.9972 | 13.8 | 21 | 97 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Godeau, C.; Morin-Crini, N.; Crini, G.; Guillemin, J.-P.; Voisin, A.-S.; Dousset, S.; Rocchi, S. Field-Crop Soils in Eastern France: Coldspots of Azole-Resistant Aspergillus fumigatus. J. Fungi 2023, 9, 618. https://doi.org/10.3390/jof9060618
Godeau C, Morin-Crini N, Crini G, Guillemin J-P, Voisin A-S, Dousset S, Rocchi S. Field-Crop Soils in Eastern France: Coldspots of Azole-Resistant Aspergillus fumigatus. Journal of Fungi. 2023; 9(6):618. https://doi.org/10.3390/jof9060618
Chicago/Turabian StyleGodeau, Chloé, Nadia Morin-Crini, Grégorio Crini, Jean-Philippe Guillemin, Anne-Sophie Voisin, Sylvie Dousset, and Steffi Rocchi. 2023. "Field-Crop Soils in Eastern France: Coldspots of Azole-Resistant Aspergillus fumigatus" Journal of Fungi 9, no. 6: 618. https://doi.org/10.3390/jof9060618
APA StyleGodeau, C., Morin-Crini, N., Crini, G., Guillemin, J.-P., Voisin, A.-S., Dousset, S., & Rocchi, S. (2023). Field-Crop Soils in Eastern France: Coldspots of Azole-Resistant Aspergillus fumigatus. Journal of Fungi, 9(6), 618. https://doi.org/10.3390/jof9060618