
Fungal Gut Microbiome in Myasthenia Gravis: A Sub-Analysis of the MYBIOM Study

 , ,
, ,  , and
, and 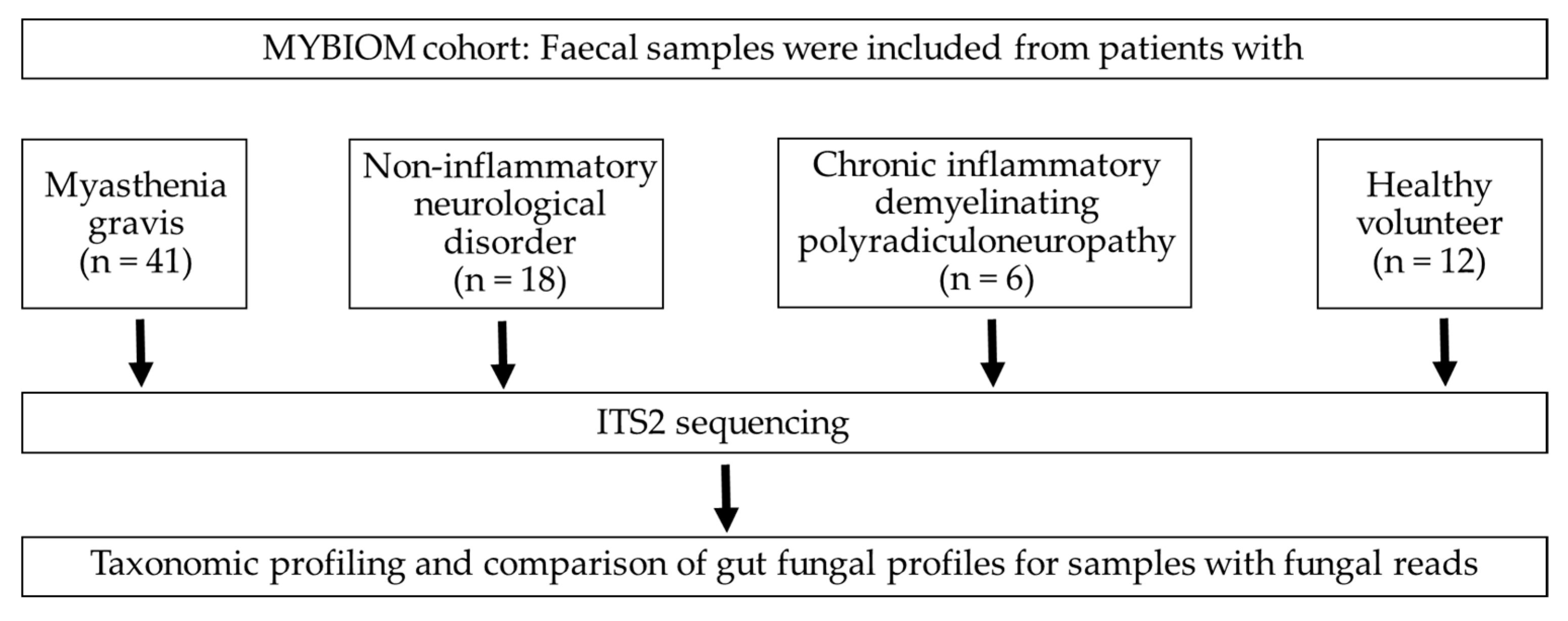{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design and Patients
2.2. Sample Collection, DNA Extraction and Sequencing
2.3. Illumina Demultiplexing
2.4. Processing and Statistical Analysis of Metataxonomic Data
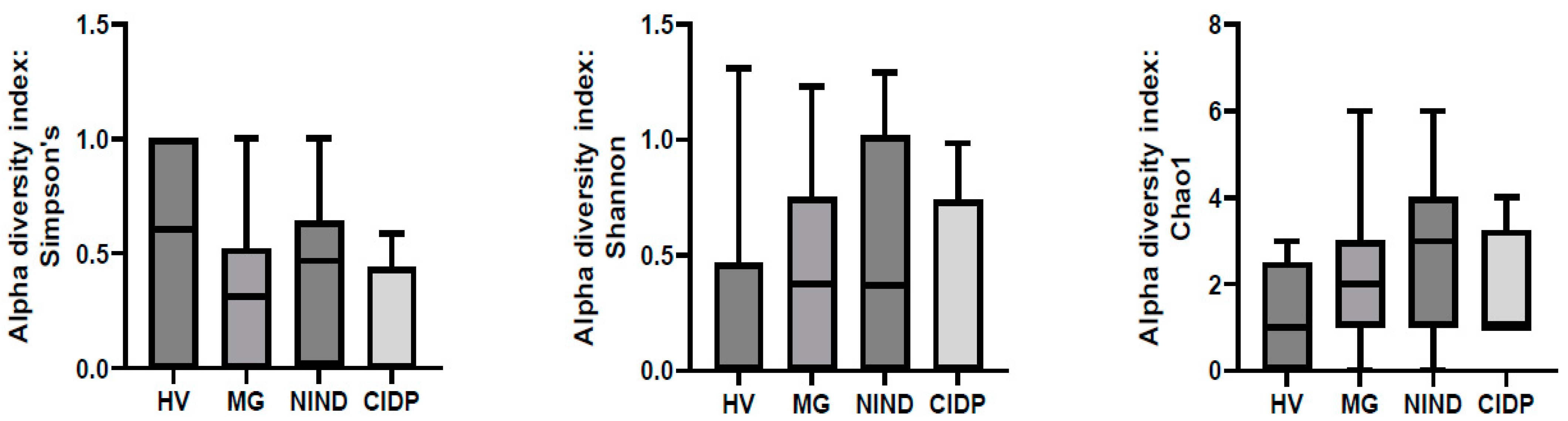
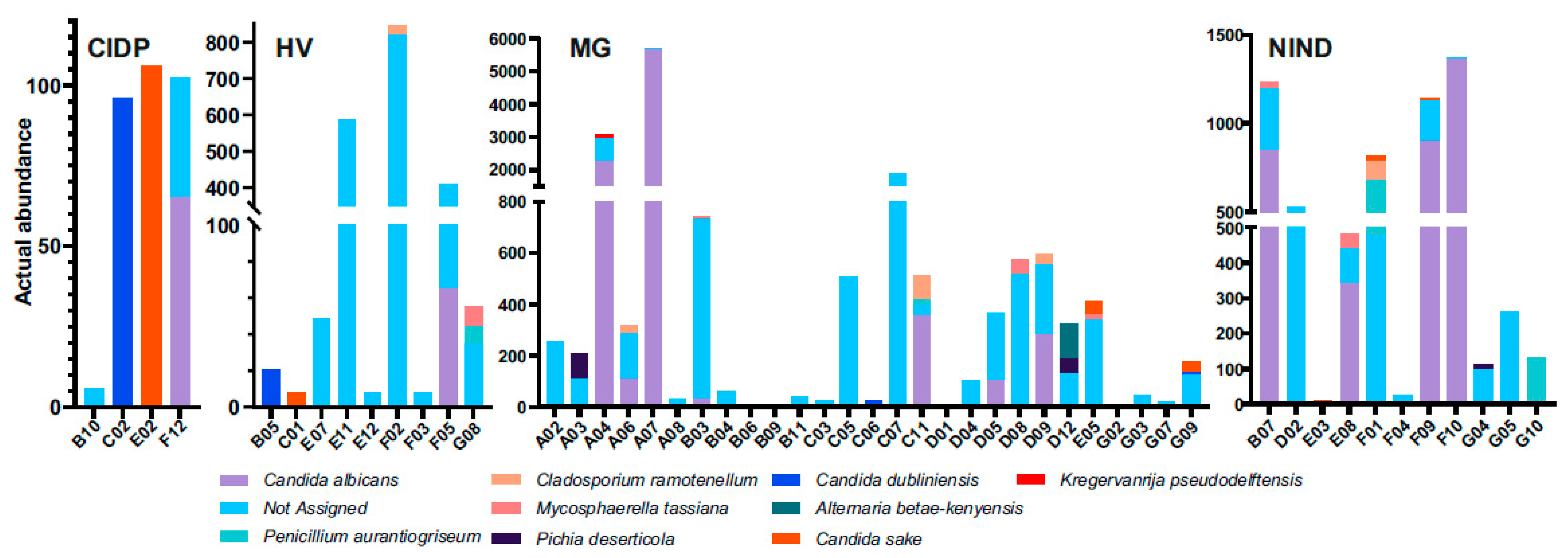
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Koneczny, I.; Herbst, R. Myasthenia Gravis: Pathogenic Effects of Autoantibodies on Neuromuscular Architecture. Cells 2019, 8, 671. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.; Li, L.; Kang, X.; Zhao, Y.; Cai, Y. Gut microbiota and metabolites in myasthenia gravis: Early diagnostic biomarkers and therapeutic strategies. Clin. Immunol. 2022, 245, 109173. [Google Scholar] [CrossRef]
- Totzeck, A.; Ramakrishnan, E.; Schlag, M.; Stolte, B.; Kizina, K.; Bolz, S.; Thimm, A.; Stettner, M.; Marchesi, J.R.; Buer, J.; et al. Gut bacterial microbiota in patients with myasthenia gravis: Results from the MYBIOM study. Ther. Adv. Neurol. Disord. 2021, 14, 17562864211035657. [Google Scholar] [CrossRef] [PubMed]
- Moris, G.; Arboleya, S.; Mancabelli, L.; Milani, C.; Ventura, M.; de Los Reyes-Gavilan, C.G.; Gueimonde, M. Fecal microbiota profile in a group of myasthenia gravis patients. Sci. Rep. 2018, 8, 14384. [Google Scholar] [CrossRef] [PubMed]
- Qiu, D.; Xia, Z.; Jiao, X.; Deng, J.; Zhang, L.; Li, J. Altered Gut Microbiota in Myasthenia Gravis. Front. Microbiol. 2018, 9, 2627. [Google Scholar] [CrossRef] [PubMed]
- Rinaldi, E.; Consonni, A.; Cordiglieri, C.; Sacco, G.; Crasa, C.; Fontana, A.; Morelli, L.; Elli, M.; Mantegazza, R.; Baggi, F. Therapeutic Effect of Bifidobacterium Administration on Experimental Autoimmune Myasthenia Gravis in Lewis Rats. Front. Immunol. 2019, 10, 2949. [Google Scholar] [CrossRef]
- Shah, S.; Locca, A.; Dorsett, Y.; Cantoni, C.; Ghezzi, L.; Lin, Q.; Bokoliya, S.; Panier, H.; Suther, C.; Gormley, M.; et al. Alterations of the gut mycobiome in patients with MS. EBioMedicine 2021, 71, 103557. [Google Scholar] [CrossRef]
- Yadav, M.; Ali, S.; Shrode, R.L.; Shahi, S.K.; Jensen, S.N.; Hoang, J.; Cassidy, S.; Olalde, H.; Guseva, N.; Paullus, M.; et al. Multiple sclerosis patients have an altered gut mycobiome and increased fungal to bacterial richness. PLoS ONE 2022, 17, e0264556. [Google Scholar] [CrossRef]
- Zou, R.; Wang, Y.; Duan, M.; Guo, M.; Zhang, Q.; Zheng, H. Dysbiosis of Gut Fungal Microbiota in Children with Autism Spectrum Disorders. J. Autism Dev. Disord. 2021, 51, 267–275. [Google Scholar] [CrossRef]
- Zhang, X.; Pan, L.Y.; Zhang, Z.; Zhou, Y.Y.; Jiang, H.Y.; Ruan, B. Analysis of gut mycobiota in first-episode, drug-naive Chinese patients with schizophrenia: A pilot study. Behav. Brain Res. 2020, 379, 112374. [Google Scholar] [CrossRef]
- Strati, F.; Calabro, A.; Donati, C.; De Felice, C.; Hayek, J.; Jousson, O.; Leoncini, S.; Renzi, D.; Rizzetto, L.; De Filippo, C.; et al. Intestinal Candida parapsilosis isolates from Rett syndrome subjects bear potential virulent traits and capacity to persist within the host. BMC Gastroenterol. 2018, 18, 57. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Santos, N.; Gaffen, S.L. Th17 cells in immunity to Candida albicans. Cell Host Microbe 2012, 11, 425–435. [Google Scholar] [CrossRef] [PubMed]
- Iliev, I.D.; Funari, V.A.; Taylor, K.D.; Nguyen, Q.; Reyes, C.N.; Strom, S.P.; Brown, J.; Becker, C.A.; Fleshner, P.R.; Dubinsky, M.; et al. Interactions between commensal fungi and the C-type lectin receptor Dectin-1 influence colitis. Science 2012, 336, 1314–1317. [Google Scholar] [CrossRef]
- van der Velden, W.J.; Netea, M.G.; de Haan, A.F.; Huls, G.A.; Donnelly, J.P.; Blijlevens, N.M. Role of the mycobiome in human acute graft-versus-host disease. Biol. Blood Marrow Transplant. 2013, 19, 329–332. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Aschenbrenner, D.; Yoo, J.Y.; Zuo, T. The gut mycobiome in health, disease, and clinical applications in association with the gut bacterial microbiome assembly. Lancet Microbe 2022, 3, e969–e983. [Google Scholar] [CrossRef]
- Melzer, N.; Ruck, T.; Fuhr, P.; Gold, R.; Hohlfeld, R.; Marx, A.; Melms, A.; Tackenberg, B.; Schalke, B.; Schneider-Gold, C.; et al. Clinical features, pathogenesis, and treatment of myasthenia gravis: A supplement to the Guidelines of the German Neurological Society. J. Neurol. 2016, 263, 1473–1494. [Google Scholar] [CrossRef]
- Van den Bergh, P.Y.; Hadden, R.D.; Bouche, P.; Cornblath, D.R.; Hahn, A.; Illa, I.; Koski, C.L.; Leger, J.M.; Nobile-Orazio, E.; Pollard, J.; et al. European Federation of Neurological Societies/Peripheral Nerve Society guideline on management of chronic inflammatory demyelinating polyradiculoneuropathy: Report of a joint task force of the European Federation of Neurological Societies and the Peripheral Nerve Society—First revision. Eur. J. Neurol. 2010, 17, 356–363. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.J.W.T.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press, Inc.: Cambridge, MA, USA, 1990; pp. 315–322. [Google Scholar]
- Chong, J.; Liu, P.; Zhou, G.; Xia, J. Using MicrobiomeAnalyst for comprehensive statistical, functional, and meta-analysis of microbiome data. Nat. Protoc. 2020, 15, 799–821. [Google Scholar] [CrossRef]
- Meklin, T.; Reponen, T.; McKinstry, C.; Cho, S.H.; Grinshpun, S.A.; Nevalainen, A.; Vepsalainen, A.; Haugland, R.A.; Lemasters, G.; Vesper, S.J. Comparison of mold concentrations quantified by MSQPCR in indoor and outdoor air sampled simultaneously. Sci. Total. Environ. 2007, 382, 130–134. [Google Scholar] [CrossRef]
- Schubert, K.; Groenewald, J.Z.; Braun, U.; Dijksterhuis, J.; Starink, M.; Hill, C.F.; Zalar, P.; de Hoog, G.S.; Crous, P.W. Biodiversity in the Cladosporium herbarum complex (Davidiellaceae, Capnodiales), with standardisation of methods for Cladosporium taxonomy and diagnostics. Stud. Mycol. 2007, 58, 105–156. [Google Scholar] [CrossRef]
- Dupuy, A.K.; David, M.S.; Li, L.; Heider, T.N.; Peterson, J.D.; Montano, E.A.; Dongari-Bagtzoglou, A.; Diaz, P.I.; Strausbaugh, L.D. Redefining the human oral mycobiome with improved practices in amplicon-based taxonomy: Discovery of Malassezia as a prominent commensal. PLoS ONE 2014, 9, e90899. [Google Scholar] [CrossRef] [PubMed]
- Pastor, F.J.; Guarro, J. Alternaria infections: Laboratory diagnosis and relevant clinical features. Clin. Microbiol. Infect. 2008, 14, 734–746. [Google Scholar] [CrossRef] [PubMed]
- Bhagat, J.; Kaur, A.; Kaur, R.; Yadav, A.K.; Sharma, V.; Chadha, B.S. Cholinesterase inhibitor (Altenuene) from an endophytic fungus Alternaria alternata: Optimization, purification and characterization. J. Appl. Microbiol. 2016, 121, 1015–1025. [Google Scholar] [CrossRef]
- Wang, Y.; Zeng, Q.G.; Zhang, Z.B.; Yan, R.M.; Wang, L.Y.; Zhu, D. Isolation and characterization of endophytic huperzine A-producing fungi from Huperzia serrata. J. Ind. Microbiol. Biotechnol. 2011, 38, 1267–1278. [Google Scholar] [CrossRef]
- Archer, D.L. Freezing: An underutilized food safety technology? Int. J. Food Microbiol. 2004, 90, 127–138. [Google Scholar] [CrossRef]
- Kurtzman, C.P. New species and new combinations in the yeast genera Kregervanrija gen. nov., Saturnispora and Candida. FEMS Yeast Res. 2006, 6, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.H.J.; Yao, T.; Ma, L.; Ren, X.; Li, L. Application of Pichia Deserticola in Prevention and Treatment of Postharvest Diseases of Fruits. Patent application filed by Beijing Technology and Business University, China CN107988088B, 2017. [Google Scholar]
- Kett, D.H.; Azoulay, E.; Echeverria, P.M.; Vincent, J.L.; FCCM for the Extended Prevalence of Infection in the ICU Study (EPIC II) Group of Investigators. Candida bloodstream infections in intensive care units: Analysis of the extended prevalence of infection in intensive care unit study. Crit. Care Med. 2011, 39, 665–670. [Google Scholar] [CrossRef]
- Nash, A.K.; Auchtung, T.A.; Wong, M.C.; Smith, D.P.; Gesell, J.R.; Ross, M.C.; Stewart, C.J.; Metcalf, G.A.; Muzny, D.M.; Gibbs, R.A.; et al. The gut mycobiome of the Human Microbiome Project healthy cohort. Microbiome 2017, 5, 153. [Google Scholar] [CrossRef]
- Poulain, D. Candida albicans, plasticity and pathogenesis. Crit. Rev. Microbiol. 2015, 41, 208–217. [Google Scholar] [CrossRef]
- Tedersoo, L.; Nilsson, R.H.; Abarenkov, K.; Jairus, T.; Sadam, A.; Saar, I.; Bahram, M.; Bechem, E.; Chuyong, G.; Koljalg, U. 454 Pyrosequencing and Sanger sequencing of tropical mycorrhizal fungi provide similar results but reveal substantial methodological biases. New. Phytol. 2010, 188, 291–301. [Google Scholar] [CrossRef]
- Lee, C.K.; Herbold, C.W.; Polson, S.W.; Wommack, K.E.; Williamson, S.J.; McDonald, I.R.; Cary, S.C. Groundtruthing next-gen sequencing for microbial ecology-biases and errors in community structure estimates from PCR amplicon pyrosequencing. PLoS ONE 2012, 7, e44224. [Google Scholar] [CrossRef]
- Hawksworth, D.L.; Crous, P.W.; Redhead, S.A.; Reynolds, D.R.; Samson, R.A.; Seifert, K.A.; Taylor, J.W.; Wingfield, M.J.; Abaci, O.; Aime, C.; et al. The amsterdam declaration on fungal nomenclature. IMA Fungus 2011, 2, 105–112. [Google Scholar] [CrossRef]
- Fiedorova, K.; Radvansky, M.; Nemcova, E.; Grombirikova, H.; Bosak, J.; Cernochova, M.; Lexa, M.; Smajs, D.; Freiberger, T. The Impact of DNA Extraction Methods on Stool Bacterial and Fungal Microbiota Community Recovery. Front. Microbiol. 2019, 10, 821. [Google Scholar] [CrossRef]
- Angebault, C.; Ghozlane, A.; Volant, S.; Botterel, F.; d’Enfert, C.; Bougnoux, M.E. Combined bacterial and fungal intestinal microbiota analyses: Impact of storage conditions and DNA extraction protocols. PLoS ONE 2018, 13, e0201174. [Google Scholar] [CrossRef] [PubMed]
- Thielemann, N.; Herz, M.; Kurzai, O.; Martin, R. Analyzing the human gut mycobiome—A short guide for beginners. Comput. Struct. Biotechnol. J. 2022, 20, 608–614. [Google Scholar] [CrossRef] [PubMed]
- Lucking, R.; Aime, M.C.; Robbertse, B.; Miller, A.N.; Ariyawansa, H.A.; Aoki, T.; Cardinali, G.; Crous, P.W.; Druzhinina, I.S.; Geiser, D.M.; et al. Unambiguous identification of fungi: Where do we stand and how accurate and precise is fungal DNA barcoding? IMA Fungus 2020, 11, 14. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, R.H.; Anslan, S.; Bahram, M.; Wurzbacher, C.; Baldrian, P.; Tedersoo, L. Mycobiome diversity: High-throughput sequencing and identification of fungi. Nat. Rev. Microbiol. 2019, 17, 95–109. [Google Scholar] [CrossRef]
- Gdanetz, K.; Benucci, G.M.N.; Vande Pol, N.; Bonito, G. CONSTAX: A tool for improved taxonomic resolution of environmental fungal ITS sequences. BMC Bioinform. 2017, 18, 538. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Verhasselt, H.L.; Ramakrishnan, E.; Schlag, M.; Marchesi, J.R.; Buer, J.; Kleinschnitz, C.; Hagenacker, T.; Totzeck, A. Fungal Gut Microbiome in Myasthenia Gravis: A Sub-Analysis of the MYBIOM Study. J. Fungi 2023, 9, 569. https://doi.org/10.3390/jof9050569
Verhasselt HL, Ramakrishnan E, Schlag M, Marchesi JR, Buer J, Kleinschnitz C, Hagenacker T, Totzeck A. Fungal Gut Microbiome in Myasthenia Gravis: A Sub-Analysis of the MYBIOM Study. Journal of Fungi. 2023; 9(5):569. https://doi.org/10.3390/jof9050569
Chicago/Turabian StyleVerhasselt, Hedda Luise, Elakiya Ramakrishnan, Melina Schlag, Julian R Marchesi, Jan Buer, Christoph Kleinschnitz, Tim Hagenacker, and Andreas Totzeck. 2023. "Fungal Gut Microbiome in Myasthenia Gravis: A Sub-Analysis of the MYBIOM Study" Journal of Fungi 9, no. 5: 569. https://doi.org/10.3390/jof9050569
APA StyleVerhasselt, H. L., Ramakrishnan, E., Schlag, M., Marchesi, J. R., Buer, J., Kleinschnitz, C., Hagenacker, T., & Totzeck, A. (2023). Fungal Gut Microbiome in Myasthenia Gravis: A Sub-Analysis of the MYBIOM Study. Journal of Fungi, 9(5), 569. https://doi.org/10.3390/jof9050569