
Velvet Family Members Regulate Pigment Synthesis of the Fruiting Bodies of Auricularia cornea


Abstract
1. Introduction
2. Materials and Methods
2.1. Test Strains
2.2. Media
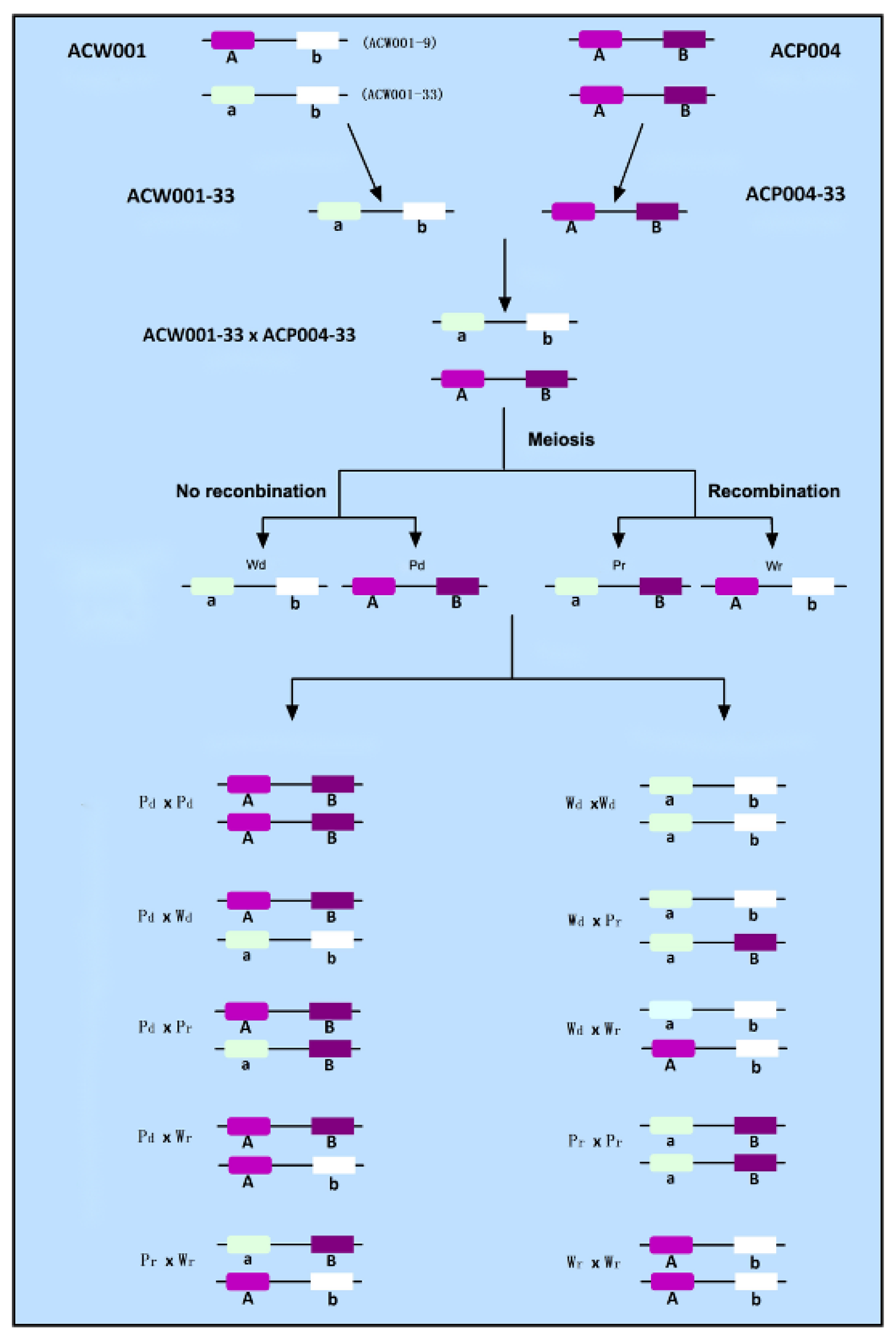
2.3. Preparation of Genetic Population
2.4. Fine Mapping of Genes That Control Color
2.5. Function Prediction of Candidate Genes
2.6. Functional Verification of Candidate Genes
3. Results
3.1. Coloration of the FBs of A. cornea Is Not Controlled by a Single Locus
3.2. Acquisition of Recombinant Monokaryotic Strains
3.3. Fine Color Typing of the Mapping Population
3.4. Coloration of the FBs of A. cornea Is Controlled by Two Loci
3.5. Analysis of Color Locus Composition
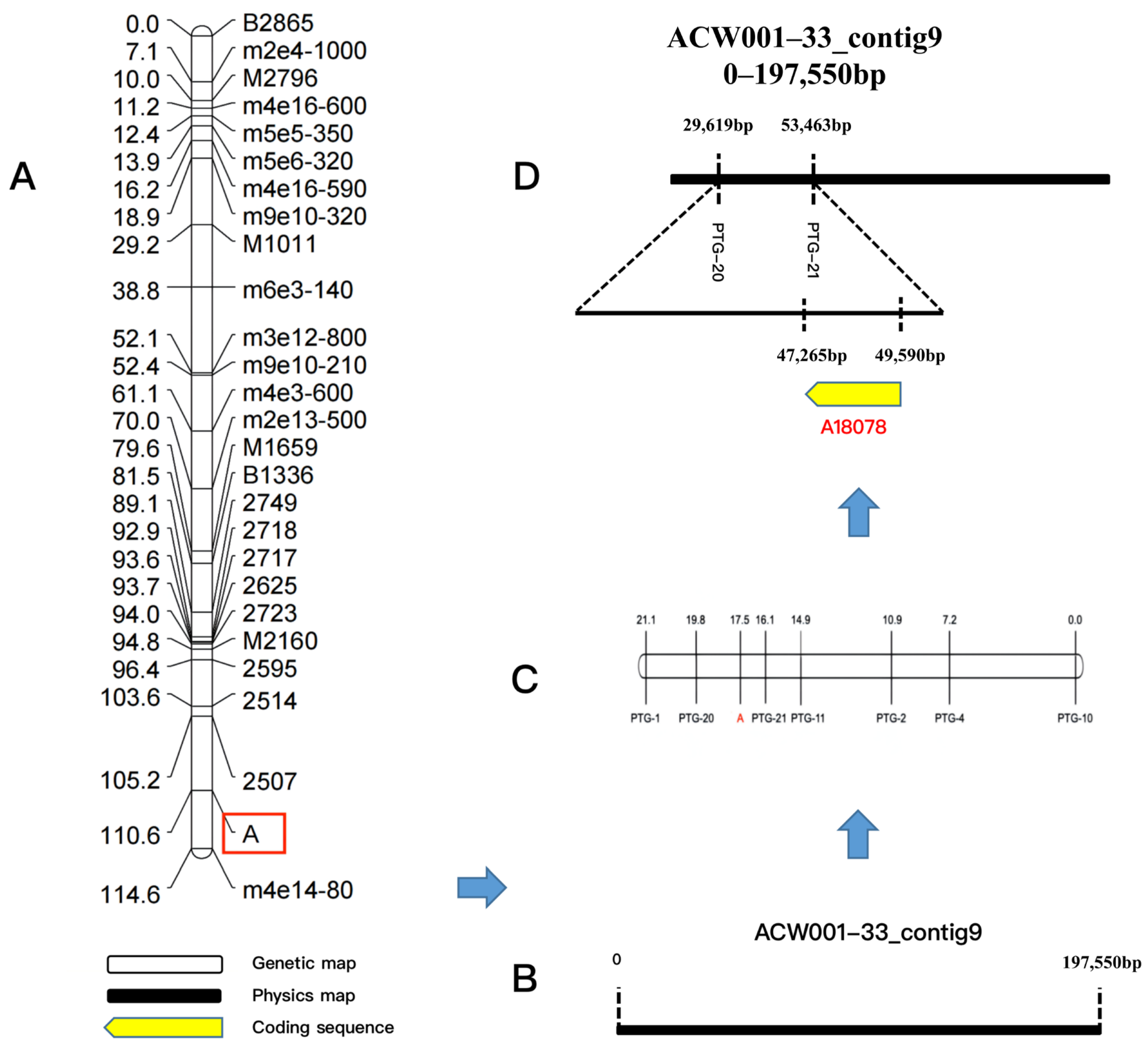
3.6. Fine Mapping of Candidate Genes for Color Control at Locus A
3.7. Prediction of Candidate Functional Genes for Color Control
3.8. Verification of the Functions of Candidate Genes for Color Control
4. Discussion
4.1. Roles of Mating Types in Genetic Populations
4.2. Color Segregation Ratio of FBs among Different Genetic Populations and Genetic Relationship between Two Color Control Loci
4.3. Distorted Segregation of Mating-Type Loci and Color Loci
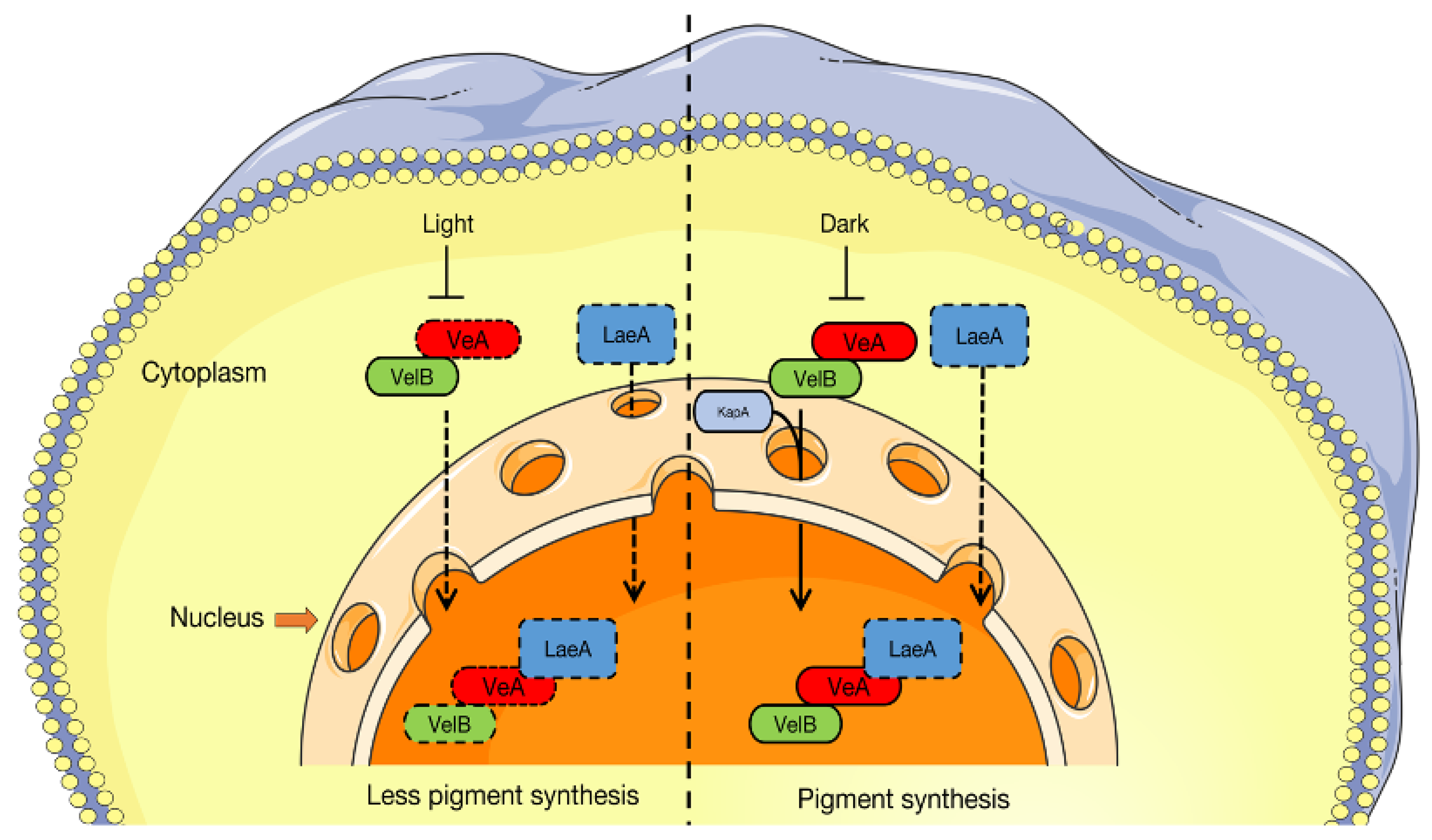
4.4. Regulatory Mechanism of Velvet Family in the Pigment Synthesis Pathway
4.5. Mechanistic Differences in Pigment Synthesis Regulated by the A and B Loci
4.6. Positional Cloning Technology Based on Genetic Linkage Maps for Mapping of Functional Genes
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Callac, P.; Moquet, F.; Imbernon, M.; Lafargue, G.; Mamoun, M.; Olivier, J.M. Evidence for PPC1, a Determinant of the Pilei-Pellis Color of Agaricus bisporus Fruitbodies. Fungal Genet. Biol. 1998, 23, 181–188. [Google Scholar] [CrossRef]
- Miller, R.; Robbins, W.; Kananen, D. Inheritance of sporophore color and “wild” morphology in Agaricus bisporus. Mushroom Sci. 1976, 9, 39–45. [Google Scholar]
- Xie, B.; Liu, W.; Wang, X.; Jiang, Y. Molecular marker for fruit-body color gene of Flammulina velutipes. J. Fujian Agric. For. Univ. 2004, 33, 363–368. [Google Scholar]
- Kong, W.; You, C.; Yoo, Y.; Kim, G.; Kim, K. Research Articles: Molecular Marker Related to Fruitbody Color of Flammulina velutipes. Microbiology 2004, 23, 6–10. [Google Scholar]
- Wang, G.; Chen, L.; Tang, W.; Wang, Y.; Zhang, Q.; Wang, H.; Zhou, X.; Wu, H.; Guo, L.; Dou, M.; et al. Identifying a melanogenesis-related candidate gene by a high-quality genome assembly and population diversity analysis in Hypsizygus marmoreus. J. Genet. Genom. 2021, 48, 75. [Google Scholar] [CrossRef]
- Sheng, C. Studies on the Inheritance of Fruit Body Color and Molecular Markers of Color Traits in Pleurotus cornucopiae. Msater’s Thesis, Ji Lin Agricultural University, Changchun, China, 2012. [Google Scholar]
- Qing, Y.; Yu, M.; Luo, X.; Jiang, N.; Ge, S. Isolation, purification and structural characterisation of bioactive polysaccharide APPIIA from Auricularia polytricha. Mycosystema 2009, 28, 813–818. [Google Scholar]
- Wu, C.; Chen, Q. Anticoagulant and Hypolipidemic Effects of Auricularia polytricha. J. China Pharm. Univ. 1991, 03, 164–166. [Google Scholar]
- Zhang, D.; Chen, G.; Pan, Y.; Zheng, S. Study on anti-tumor effect of intracellular polysaccharide from Auricularia polytricha. Edible Fungi 2014, 36, 75–77. [Google Scholar]
- Zhao, S.; Liu, Y.; Xu, F.; Wang, S.; Geng, X. The hypolipidemic function of Auricularia polytricha protein extract. Mod. Food Sci. Technol. 2013, 29, 941–944. [Google Scholar]
- Ren, Z.; Fang, H.; Meng, X.; Zhang, K.; Dai, J.; Li, S.; Li, Z.; Li, X. Breeding of a New White Auricularia polytricha Variety “Yumuer”. Mol. Plant Breed. 2018, 16, 954–959. [Google Scholar] [CrossRef]
- Wang, P.; Yao, F.; Lu, L.; Fang, M.; Zhang, Y.; Khan, A.; Kong, X.; Yu, J.; Jiang, W.; Kitamoto, Y.; et al. Map-based cloning of genes encoding key enzymes for pigment synthesis in Auricularia cornea. Fungal Biol. 2019, 123, 843–853. [Google Scholar] [CrossRef]
- Kofler, R.; Schlötterer, C.; Lelley, T. SciRoKo: A new tool for whole genome microsatellite search and investigation. Bioinformatics 2007, 23, 1683–1685. [Google Scholar] [CrossRef]
- Ooijen, J.; Van, J. JoinMap 4, Software for the Calculation of Genetic Linkage Maps in Experimental Populations. 2006. Available online: https://docplayer.net/35204534-Joinmap-software-for-the-calculation-of-genetic-linkage-maps-in-experimental-populations-j-w-van-ooijen.html (accessed on 21 May 2021).
- Gil, R.; Lea, C. An alternative pathway to beta-carotene formation in plant chromoplasts discovered by map-based. Proc. Natl. Acad. Sci. USA 2000, 97, 11102–11107. [Google Scholar]
- Brückner, A.; Polge, C.; Lentze, N.; Auerbach, D.; Schlattner, U. Yeast two-hybrid, a powerful tool for systems biology. Int. J. Mol. Sci. 2009, 10, 2763–2788. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef]
- Pertea, M.; Pertea, G.; Antonescu, C.; Chang, T.; Mendell, J.; Salzberg, S. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef]
- Lan, N.; Zhang, H.; Hu, C.; Wang, W.; Calvo, A.; Harris, S.; Chen, S.; Li, S. Coordinated and Distinct Functions of Velvet Proteins in Fusarium verticillioides. Eukaryot Cell 2014, 13, 909–918. [Google Scholar] [CrossRef]
- Özgür, B.; Braus, G. Coordination of secondary metabolism and development in fungi: The velvet family of regulatory proteins. FEMS Microbiol. Rev. 2012, 36, 1–24. [Google Scholar]
- Sarikaya-Bayram, Ö.; Bayram, Ö.; Feussner, K.; Kim, J.; Kim, H.; Kaever, A.; Feussner, I.; Chae, K.; Han, D.; Han, K. Membrane-Bound Methyltransferase Complex VapA-VipC-VapB Guides Epigenetic Control of Fungal Development. Dev. Cell 2014, 29, 406–420. [Google Scholar] [CrossRef]
- Rechsteiner, M.; Rogers, S. PEST sequences and regulation by proteolysis. Trends Biochem. Sci. 1996, 21, 267–271. [Google Scholar] [CrossRef]
- Bayram, O.; Krappmann, S.; Ni, M.; Bok, J.; Helmstaedt, K.; Valerius, O.; Braus-Stromeyer, S.; Kwon, N.; Keller, N.; Yu, J.; et al. VelB/VeA/LaeA Complex Coordinates Light Signal with Fungal Development and Secondary Metabolism. Science 2008, 320, 1504–1506. [Google Scholar] [CrossRef]
- Duncan, E. Microevolution in Auricularia polytricha. Mycologia 1972, 64, 394. [Google Scholar] [CrossRef]
- Luo, X. Study on Polarity of Auricularia auricula and Auricularia polytricha. Acta Mycol. Sin. 1988, 01, 56–61. [Google Scholar] [CrossRef]
- Lu, L.; Yao, F.; Wang, P.; Fang, M.; Lu, J. Construction of a genetic linkage map and QTL mapping of agronomic traits in Auricularia auricula-judae. J. Microbiol. 2017, 55, 792–799. [Google Scholar] [CrossRef]
- Harushima, Y.; Kurata, N.; Yano, M.; Nagamura, Y.; Sasaki, T.; Minobe, Y.; Nakagahra, M. Detection of segregation distortions in Anindica-japonicarice cross using a high-resolution molecular map. Theor. Appl. Genet. 1996, 92, 145–150. [Google Scholar] [CrossRef]
- Mangelsdorf, P.; Jones, D. The expression of mendelian factors in the gametophyte of maize. Genetics 1926, 11, 423–455. [Google Scholar] [CrossRef]
- Byrne, M.; Murrell, J.; Allen, B.; Moran, G. An integrated genetic linkage map for eucalypts using RFLP, RAPD and isozyme markers. Theor. Appl. Genet. 1995, 91, 869–875. [Google Scholar] [CrossRef]
- Helentjaris, M. Inheritance of RAPDs in F1 hybrids of corn. TAG Theor. Appl. Genet. 1993, 85, 961–968. [Google Scholar]
- Fauré, S.; Noyer, J.; Horry, J.; Bakry, F.; Lanaud, C.; de León, D.G. A molecular marker-based linkage map of diploid bananas (Musa acuminata). Theor. Appl. Genet. 1993, 87, 517–526. [Google Scholar] [CrossRef]
- Raper, C. B-Mating-type genes influence survival of nuclei separated from heterokaryons of Schizophyllum. Exp. Mycol. 1985, 9, 149–160. [Google Scholar] [CrossRef]
- Ahmed, Y.; Gerke, J.; Park, H.; Bayram, O.; Neumann, P.; Ni, M.; Dickmanns, A.; Kim, S.; Yu, J.; Braus, G.; et al. The velvet family of fungal regulators contains a DNA-binding domain structurally similar to NF-kappaB. PLoS Biol. 2013, 11, e1001750. [Google Scholar] [CrossRef]
- Cary, J.; Harris-Coward, P.; Ehrlich, K.; Di-Mavungu, J.; Malysheva, S.; De-Saeger, S.; Dowd, P.; Shantappa, S.; Martens, S.; Calvo, A. Functional characterization of a veA-dependent polyketide synthase gene in Aspergillus flavus necessary for the synthesis of asparasone, a sclerotium-specific pigment. Fungal Genet. Biol. 2014, 64, 25–35. [Google Scholar] [CrossRef]
- Mukherjee, G.; Mishra, T.; Deshmukh, S. Fungal Pigments: An Overview. In Developments in Fungal Biology and Applied Mycology; Springer: Berlin/Heidelberg, Germany, 2017; pp. 525–541. [Google Scholar]
- Chang, P.; Scharfenstein, L.; Li, P.; Ehrlich, K. Aspergillus flavus VelB acts distinctly from VeA in conidiation and may coordinate with FluG to modulate sclerotial production. Fungal Genet. Biol. 2013, 58–59, 71–79. [Google Scholar] [CrossRef]
- Kopke, K.; Hoff, B.; Bloemendal, S.; Katschorowski, A.; Kamerewerd, J.; Kuck, U. Members of the Penicillium chrysogenum velvet complex play functionally opposing roles in the regulation of penicillin biosynthesis and conidiation. Eukaryot Cell 2013, 12, 299–310. [Google Scholar] [CrossRef]
- Park, H.; Nam, T.; Han, K.; Kim, S.; Yu, J. VelC Positively Controls Sexual Development in Aspergillus nidulans. PLoS ONE 2014, 9, e89883. [Google Scholar] [CrossRef]
- Jiang, J.; Liu, X.; Yin, Y.; Ma, Z.; Yin, L. Involvement of a Velvet Protein FgVeA in the Regulation of Asexual Development, Lipid and Secondary Metabolisms and Virulence in Fusarium graminearum. PLoS ONE 2011, 6, e28291. [Google Scholar] [CrossRef]
- Park, H.; Ni, M.; Jeong, K.; Kim, Y.; Yu, J. The Role, Interaction and Regulation of the Velvet Regulator VelB in Aspergillus nidulans. PLoS ONE 2012, 7, e45935. [Google Scholar] [CrossRef]
- Tanksley, S.D.; Ganal, M.W.; Martin, G.B. Chromosome landing: A paradigm for map-based gene cloning in plants with large genomes. Trends Genet. 1995, 11, 63–68. [Google Scholar] [CrossRef]
- He, J.P.; Ruan, S.L.; Zhu, S.J. The application and evaluation of map-based gene isolation in crops. Hereditas 2010, 32, 903. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Monokaryon | Pd | Pr | Wd | Wr |
|---|---|---|---|---|
| Pd | P | P | P | P |
| Pr | P | W | W | P |
| Wd | P | W | W | W |
| Wr | P | P | W | W |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, X.; Lu, L.; Zhang, Y.; Fang, M.; Shao, K.; Sun, X.; Yao, F.; Wang, P. Velvet Family Members Regulate Pigment Synthesis of the Fruiting Bodies of Auricularia cornea. J. Fungi 2023, 9, 412. https://doi.org/10.3390/jof9040412
Ma X, Lu L, Zhang Y, Fang M, Shao K, Sun X, Yao F, Wang P. Velvet Family Members Regulate Pigment Synthesis of the Fruiting Bodies of Auricularia cornea. Journal of Fungi. 2023; 9(4):412. https://doi.org/10.3390/jof9040412
Chicago/Turabian StyleMa, Xiaoxu, Lixin Lu, Youmin Zhang, Ming Fang, Kaisheng Shao, Xu Sun, Fangjie Yao, and Peng Wang. 2023. "Velvet Family Members Regulate Pigment Synthesis of the Fruiting Bodies of Auricularia cornea" Journal of Fungi 9, no. 4: 412. https://doi.org/10.3390/jof9040412
APA StyleMa, X., Lu, L., Zhang, Y., Fang, M., Shao, K., Sun, X., Yao, F., & Wang, P. (2023). Velvet Family Members Regulate Pigment Synthesis of the Fruiting Bodies of Auricularia cornea. Journal of Fungi, 9(4), 412. https://doi.org/10.3390/jof9040412