
Three New Species of Lactifluus (Basidiomycota, Russulaceae) from Guizhou Province, Southwest China

 , , , , and
, , , , and
Abstract
1. Introduction
2. Material and Methods
2.1. Study Site and Specimen Collection
2.2. Morphological Analysis
2.3. DNA Extraction, Amplification, and Sequencing
2.4. Sequence Alignment and Phylogenetic Analysis
2.5. Phylogenetic Analyses
3. Results
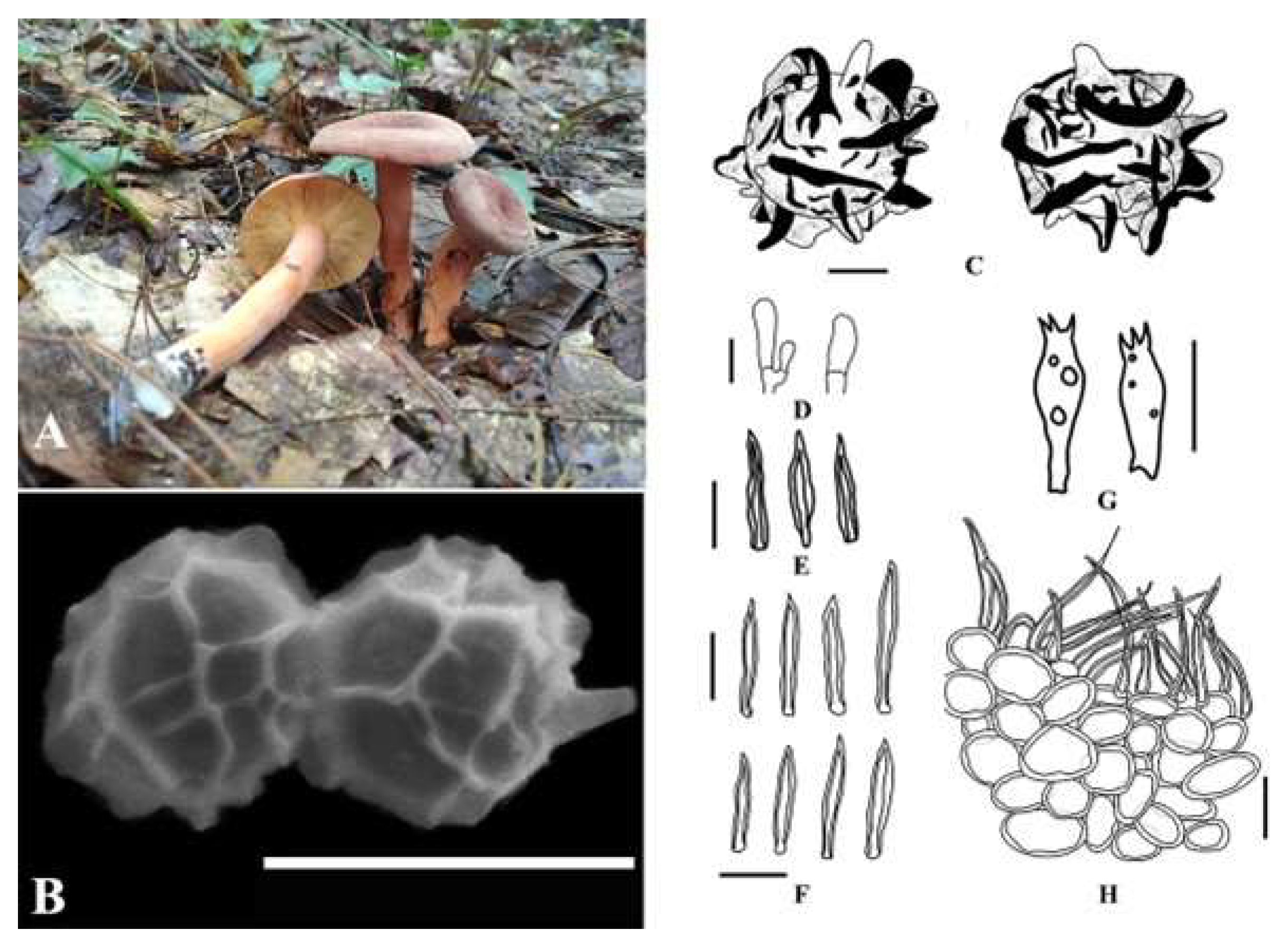
Taxonomy
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Buyck, B.; Hofstetter, V.; Eberhardt, U.; Verbeken, A.; Kauff, F. Walking the thin line between Russula and Lactarius: The dilemma of Russula subsect. Ochricompactae. Fungal Divers. 2008, 28, 15–40. [Google Scholar]
- Buyck, B.; Hofstetter, V.; Verbeken, A.; Walleyn, R. Proposal 1919: To conserve Lactarius nom. cons. (Basidiomycota) with a conserved type. Mycotaxon 2010, 111, 504–508. [Google Scholar] [CrossRef]
- McNeill, J.; Turland, N.J.; Monro, A.M.; Lepschi, B.J. XVIII International botanical congress: Preliminary mail vote and report of congress action on nomenclature proposals. Taxon 2011, 60, 1507–1520. [Google Scholar] [CrossRef]
- Tedersoo, L.; Bahram, M.; Jairus, T.; Bechem, E.; Chinoya, S.; Mpumba, R.; Leal, M.; Randrianjohany, E.; Razafimandimbison, S.; Sadam, A.; et al. Spatial structure and the effects of host and soil environments on communities of ectomycorrhizal fungi in wooded savannas and rain forests of Continental Africa and Madagascar. Mol. Ecol. 2011, 20, 3071–3080. [Google Scholar] [CrossRef] [PubMed]
- Tedersoo, L.; Nilsson, R.H.; Abarenkov, K.; Jairus, T.; Sadam, A.; Saar, I.; Bahram, M.; Bechem, E.; Chuyong, G.; Koljalg, U. 454 Pyrosequencing and Sanger sequencing of tropical mycorrhizal fungi provide similar results but reveal substantial methodological biases. New Phytologist. 2010, 188, 291–301. [Google Scholar] [CrossRef]
- De Crop, E. Global Phylogeny and Evolutionary History of the Genus Lactifluus. Ph. D. Thesis, Ghent University, Ghent, Belgium, 2016. [Google Scholar]
- De Crop, E.; Nuytinck, J.; Van de Putte, K.; Wisitrassameewong, K.; Hackel, J.; Stubbe, D.; Hyde, K.D.; Roy, M.; Halling, R.E.; Moreau, P.A.; et al. A multi-gene phylogeny of Lactifluus (Basidiomycota, Russulales) translated into a new infrageneric classification of the genus. Persoonia 2017, 38, 58–80. [Google Scholar] [CrossRef]
- De Crop, E.; Delgat, L.; Nuytinck, J.; Halling, R.E.; Verbeken, A. A short story of nearly everything in Lactifluus (Russulaceae). Fungal Sys. Evol. 2021, 7, 133–164. [Google Scholar] [CrossRef]
- Hyde, K.D.; Norphanphoun, C.; Abreu, V.P.; Bazzicalupo, A.; Chethana, K.W.T.; Clericuzio, M.; Dayarathne, M.C.; Dissanayake, A.J.; Ekanayaka, A.H.; He, M.Q.; et al. Fungal diversity notes 603–708: Taxonomic and phylogenetic notes on genera and species. Fungal Divers. 2017, 87, 1–235. [Google Scholar] [CrossRef]
- Boonmee, S.; Wanasinghe, D.N.; Calabon, M.S. Huanraluek, N.; Chandrasiri, S.K.U.; Jones, G.E.B.; Rossi, W.; Leonardi, M.; Singh, S.K.; Rana, S.; et al. Fungal diversity notes 1387-1511: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2021, 111, 1–335. [Google Scholar] [CrossRef]
- Verbeken, A.; Nuytinck, J. Not every milkcap is a Lactarius. Scr. Bot. Belgica. 2013, 51, 162–168. [Google Scholar]
- Delgat, L.; Dierickx, G.; Wilde, S.D.; Angelini, C.; De Crop, E.; De Lange, R.; Halling, R.; Manz, C.; Nuytinck, J.; Verbeken, A. Looks can be deceiving: The deceptive milkcaps (Lactifluus, Russulaceae) exhibit low morphological variance but harbour high genetic diversity. IMA Fungus 2019, 10, 14. [Google Scholar] [CrossRef]
- Stubbe, D.; Nuytinck, J.; Verbeken, A. Critical assessment of the Lactarius gerardii species complex (Russulales). Fungal Biol. 2010, 114, 271–283. [Google Scholar] [CrossRef]
- Stubbe, D.; Le, H.T.; Wang, X.H.; Nuytinck, J.; Van de Putte, K.; Verbeken, A. The Australasian species of Lactarius subgenus Gerardii (Russulales). Fungal Divers. 2012, 52, 141–167. [Google Scholar] [CrossRef]
- Van de Putte, K.; Nuytinck, J.; Stubbe, D.; Huyen, T.L.; Verbeken, A. Lactarius volemus sensu lato (Russulales) from Northern Thailand: Morphological and phylogenetic species concepts explored. Fungal Divers. 2010, 45, 99–130. [Google Scholar] [CrossRef]
- Van de Putte, K.; Nuytinck, J.; Das, K.; Verbeken, A. Exposing hidden diversity by concordant genealogies and morphology–a study of the Lactifluus volemus (Russulales) species complex in Sikkim Himalaya (India). Fungal Divers. 2012, 55, 171–194. [Google Scholar] [CrossRef]
- Van de Putte, K.; Nuytinck, J.; De Crop, E.; Verbeken, A. Lactifluus volemus in Europe: Three species in one-Revealed by a multilocus genealogical approach, Bayesian species delimitation and morphology. Fungal Biol. 2016, 120, 1–25. [Google Scholar] [CrossRef]
- De Crop, E.; Nuytinck, J.; Van de Putte, K.; Lecomte, M.; Eberhardt, U.; Verbeken, A. Lactifluus piperatus (Russulales, Basidiomycota) and allied species in Western Europe and a preliminary overview of the group worldwide. Mycol. Prog. 2014, 13, 493–511. [Google Scholar] [CrossRef]
- De Crop, E.; Kobeke, V.; Wilde, S.D.; Njouonkou, A.L.; Kesel, A.D.; Verbeken, A. Lactifluus foetens and Lf. albomembranaceus sp. nov. (Russulaceae): Look-alike milkcaps from gallery forests in tropical Africa. Phytotaxa 2016, 277, 159–170. [Google Scholar] [CrossRef]
- Wang, X.H.; Buyck, B.; Verbeken, A.; Hansen, K. Revisiting the morphology and phylogeny of Lactifluus with three new lineages from Southern China. Mycologia 2015, 107, 941–958. [Google Scholar] [CrossRef]
- Lee, H.; Wissitrassameewong, K.; Park, M.S.; Verbeken, A.; Eimes, J.; Lim, Y.W. Taxonomic revision of the genus Lactarius (Russulales, Basidiomycota) in Korea. Fungal Divers. 2019, 95, 275–335. [Google Scholar] [CrossRef]
- Eberhardt, U. Molecular kinship analyses of the agaricoid Russulaceae: Correspondence with mycorrhizal anatomy and sporocarp features in the genus Russula. Mycol. Prog. 2002, 1, 201–223. [Google Scholar] [CrossRef]
- Miller, S.L.; Buyck, B. Molecular phylogeny of the genus Russula in Europe with a comparison of modern infrageneric classifications. Mycol. Res. 2002, 106, 259–276. [Google Scholar] [CrossRef]
- Song, Y.; Zhang, J.B.; Li, J.W.; Xia, S.Y.; Qiu, L.H. Phylogenetic and morphological evidence for Lactifluus robustus sp. nov. (Russulaceae) from Southern China. Nova Hedwigia 2017, 105, 519–528. [Google Scholar] [CrossRef]
- Song, Y.; Zhang, J.B.; Li, J.W.; Qiu, L.H. Lactifluus sinensis sp. nov. and L. sinensis var. reticulatus var. nov. (Russulaceae) from southern China. Nova Hedwigia 2018, 107, 91–103. [Google Scholar]
- Montoya, L.; Caro, A.; Ramos, A.; Bandala, V. Two new species of Lactifluus (Fungi, Russulales) from tropical Quercus forest in Eastern Mexico. MycoKeys 2019, 59, 27–45. [Google Scholar] [CrossRef]
- Lee, H.; Wissitrassameewong, K.; Park, M.S.; Fong, J.J.; Verbeken, A.; Kim, C.; Lim, Y.W. Taxonomic revision of the genus Lactifluus (Russulales, Basidiomycota) of South Korea. Mycobiology 2021, 49, 308–345. [Google Scholar] [CrossRef] [PubMed]
- Maba, D.L.; Guelly, A.K.; Yorou, N.S.; Agerer, R. Diversity of Lactifluus (Basidiomycota, Russulales) in West Africa: 5 new species described and some considerations regarding their distribution and ecology. Mycosphere 2015, 6, 737–759. [Google Scholar] [CrossRef]
- Houdanon, R.D.; Furneaux, B.; Yorou, N.S.; Ryberg, M. Phylogenetic diversity and affiliation of tropical African ectomycorrhizal fungi. Mycosphere 2020. [Google Scholar] [CrossRef]
- Silva, A.G.S.; Sa, M.C.A.; Komura, D.L.; Baseia, L.G.; Marinho, P.; Moncalvo, J.M.; Roy, M.; Wartchow, F. Novelties in Lactifluus subg. Gymnocarpi (Russulales, Basidiomycota) from Brazilian tropical forests. Mycol. Prog. 2021, 20, 549–565. [Google Scholar] [CrossRef]
- Hesler, L.R.; Smith, A.H. North American Species of Lactarius. Ph.D. Thesis, University of Michigan, Ann Arbor, MI, USA, 1979. [Google Scholar]
- Comandini, O.; Marco, C.; Rinaldi, A.C. An overview of Cistus ectomycorrhizal fungi. Mycorrhiza 2006, 16, 381–395. [Google Scholar] [CrossRef]
- Leonardi, M.; Comandini, O.; Rinaldi, A.C. Peering into the Mediterranean black box: Lactifluus rugatus ectomycorrhizas on Cistus. IMA Fungus 2016, 7, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Leonardi, M.; Furtado, A.N.M.; Comandini, O.; Geml, J.; Rinaldi, A.C. Halimium as an ectomycorrhizal symbiont: New records and an appreciation of known fungal diversity. Mycol. Prog. 2020, 19, 1495–1509. [Google Scholar] [CrossRef]
- De Crop, E.; Hampe, F.; Wisitrassameewong, K.; Stubbe, D.; Nuytinck, J.; Verbeken, A. Novel diversity in Lactifluus section Gerardii from Asia: Five new species with pleurotoid or small agaricoid basidiocarps. Mycologia 2018, 110, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Zhou, L.W.; Yang, Z.L.; Bau, T.; Li, T.H.; Dai, Y.C. Resource diversity of Chinese macrofungi: Edible, medicinal and poisonous species. Fungal Divers. 2019, 98, 1–76. [Google Scholar] [CrossRef]
- Li, H.J.; Zhang, H.S.; Zhang, Y.Z.; Zhang, K.P.; Zhou, J.; Yin, Y.; Jiang, S.F.; Ma, P.B.; He, Q.; Zhang, Y.T.; et al. Mushroom poisoning outbreaks—China, 2019. China CDC Wkly. 2020, 2, 19–27. [Google Scholar] [CrossRef]
- Boa, E. Wild Edible Fungi. A Global Overview of Their Use and Importance to People; FAO: Rome, Italy, 2004; p. 17. [Google Scholar]
- Colak, A.; Faiz, O.; Sesli, E. Nutritional composition of some wild edible mushrooms. Turk. J. Biochem. 2009, 34, 25–31. [Google Scholar]
- Garibay-Orijel, R.; Caballero, J.; Estrada-Toress, A.; Cifuentes, J. Understanding cultural significance, the edible mushroom case. J. Ethnobiol. Ethnomed. 2007, 3, 1–18. [Google Scholar] [CrossRef]
- Christensen, M.; Bhattarai, S.; Devkota, S.; Larsen, H.O. Collection and use of wild edible fungi in Nepal. Econ. Bot. 2008, 62, 12–23. [Google Scholar] [CrossRef]
- Dell, B.; Sanmee, R.; Lumyong, P. Ectomycorrhizal fungi in dry and wet dipterocarp forests in northern Thailand—Diversity and use as food; Asia Pacific Association of Forestry Research Institutions (APAFRI). In Proceedings of the 8th Round-Table Conference on Dipterocarps, Ho Chi Minh, Vietnam, 15–17 November 2005. [Google Scholar]
- Wang, X.H.; Liu, P.; Yu, F. Color Atlas of Wild Commercial Mushrooms in Yunnan; Yunnan Science and Technology Press: Kunming, China, 2004. (In Chinese) [Google Scholar]
- Pérez-Moreno, J.; Guerin-Laguette, A.; Rinaldi, A.C.; Yu, F.; Verbeken, A.; Hernàndez-Santiago, F.; Martìnez-Reyes, M. Edible ectomycorrhizal fungi: How can they contribute to forest sustainability, food security, biocultural conservation and mitigation of hunger and climate change? Plants People Planet 2021, 3, 471–490. [Google Scholar] [CrossRef]
- Russell, B. Field Guide to Wild Mushrooms of Pennsylvania and the Mid-Atlantic; Pennsylvania State University Press: University Park, PA, USA, 2006. [Google Scholar]
- Wang, L.; Yang, Z.L. Wild edible fungi of the Hengduan mountains, Southwestern China. In Proceedings of the Sino-German Symposium 2006—The Sustainable Harvest of Non-Timber Forest Products in China: Strategies to Balance Economic Benefits and Biodiversity Conservation, Göttingen, Germany, 13–17 March 2006; pp. 58–65. [Google Scholar]
- Le, H.T. Biodiversity of the Genus Lactarius (Basidiomycota) in Northern Thailand. Ph.D.Thesis, Chiang Mai University, Chiang Mai, Thailand, 2007. [Google Scholar]
- Liu, P.G.; Yu, F.Q.; Wang, X.H.; Zheng, H.D.; Chen, J.Y.; Tian, X.F.; Xie, X.D.; Shi, X.F.; Deng, X.J. The cultivation of Lactarius volemus in China. Acta Bot. Yunnanica 2009, XVI, 115–116. [Google Scholar]
- Lincoff, G. The Complete Mushroom Hunter: An Illustrated Guide to Finding, Harvesting, and Enjoying Wild Mushrooms; Quarry Books: Gloucester, MA, USA, 2010. [Google Scholar]
- Nuytinck, J.; De Crop, E.; Delgat, L.; Bafort, Q.; Rivas Ferreiro, M.; Verbeke, A.; Wang, X.H. Recent insights in the phylogeny, species diversity and culinary uses of milkcap genera Lactarius and Lactifluus. In Mushrooms, Humans and Nature in a Changing World: Perspectives from Ecological, Agricultural and Social Sciences; Perez, J., Guerin, A., Flores, R., Eds.; Springer: Cham, Switzerland, 2020; pp. 273–286. [Google Scholar] [CrossRef]
- Kobata, K.; Wada, T.; Hayashi, Y.; Shibata, H. Volemolide, a novel norsterol from the fungus Lactarius volemus. Biosci. Biotechnol. Biochem. 1994, 58, 1542–1544. [Google Scholar] [CrossRef]
- Daniewski, W.M.; Gumułka, M.; Przesmycka, D.; Ptaszyńska, K.; Błoszyk, E.; Drożdż, B. Sesquiterpenes of Lactarius origin, antifeedant structure-activity relationships. Phytochemistry 1995, 38, 1161–1168. [Google Scholar] [CrossRef]
- Sivakumar, M.; Bhat, S.V.; Nagasampagi, B.A. Chemistry of Natural Products; Springer: Berlin, Germany, 2005; p. 495. [Google Scholar]
- Luo, D.Q.; Zhao, L.Y.; Shi, Y.L.; Tang, H.L.; Li, Y.Y.; Yang, L.M.; Zheng, Y.T.; Zhu, H.J.; Liu, J.K. Velleratretraol, an unusual highly functionalized lactarane sesquiterpene from Lactarius vellereus. J. Antibiot. 2009, 62, 129–132. [Google Scholar] [CrossRef] [PubMed]
- Nowacka, N.; Nowak, R.; Drozd, M.; Olech, M.; Los, R.; Malm, A. Antibacterial, antiradical potential and phenolic compounds of thirty-one polish mushrooms. PLoS ONE 2015, 10, e0140355. [Google Scholar] [CrossRef]
- Shen, Q.L.; Sun, L.P.; He, Z.D.; Xie, J.X.; Zhuang, Y.L. Isolation, taste characterization and molecular docking study of novel umami peptides from Lactarius volemus (Fr.). Food Chem. 2022, 401, 134137. [Google Scholar] [CrossRef]
- Lu, S.F.; Yin, X.J.; Wei, Q.W.; Zhang, C.; Ma, D.X.; Liu, X.L. The geographical distribution response of plant functional types to climate change in southwestern China. Acta Ecol. Sin. 2020, 40, 310–324. [Google Scholar] [CrossRef]
- Zheng, Z.J.; Zeng, Y.; Zhao, Y.J.; Zhao, D.; Wu, B.F. Monitoring and dynamic analysis of fractional vegetation cover in southwestern China over the past 15 years based on MODIS data. Remote Sens. Land Resour. 2017, 29, 128–136. [Google Scholar] [CrossRef]
- Wang, R.; Herrera, M.; Xu, W.J.; Zhang, P.; Moreno, J.P.; Colinas, C.; Yu, F.Q. Ethnomycological study on wild mushrooms in Pu’er Prefecture, Southwest Yunnan, China. J. Ethnobiol. Ethnomed. 2022, 18, 55. [Google Scholar] [CrossRef]
- Fu, Z.Q.; Liu, G.; Wang, L.X. Assessment of potential human health risk of trace element in wild edible mushroom species collected from Yunnan province, China. Environ. Sci. Pollut. Res. 2020, 27, 29218–29227. [Google Scholar] [CrossRef]
- Leonardi, M.; Comandini, O.; Sanjust, E.; Rinaldi, A.C. Conservation status of milkcaps (Basidiomycota, Russulales, Russulaceae), with notes on poorly known species. Sustainability 2021, 13, 10365. [Google Scholar] [CrossRef]
- Dai, Y.C.; Yang, Z.L.; Cui, B.K.; Wu, G.; Yuan, H.S.; Zhou, L.W.; He, S.H.; Ge, Z.W.; Wu, F.; Wei, Y.L.; et al. Diversity and systematics of the important macrofungi in Chinese forests. Mycosystema. 2021, 40, 770–805. [Google Scholar] [CrossRef]
- Wang, K.; ChhhhDai, Y.C.; Jia, Z.F.; Li, T.H.; Liu, T.Z.; Phurbu, D.; Mamut, R.; Sun, G.Y.; Bau, T.; Wei, S.L.; et al. Overview of China’s nomenclature novelties of fungi in the new century (2000–2020). Mycosystema. 2021, 40, 822–833. [Google Scholar] [CrossRef]
- Shi, S.F.; Wang, X.H.; Bau, T. Three new species of Lactarius (Russulaceae, Russulales) from Northeast China. Mycoscience 2018, 59, 206–217. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Academic Press Inc.: Cambridge, MA, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Liu, Y.J.; Wehlen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerase II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef]
- Hall, T.; Biosciences, I.; Carlsbad, C. BioEdit: An important software for molecular biology. GERF Bull. Biosci. 2011, 2, 60–61. [Google Scholar]
- Benson, D.A.; Cavanaugh, M.; Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2017, 45, D37–D42. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Zhang, D.; Gao, F.L.; Jakovlić, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Minh, B.Q.; Nguyen, M.A.; von Haeseler, A. Ultrafast approximation for phylogenetic bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. Modelfinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Rambaut, A. FigTree version 1.4.0. 2012. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 1 November 2022).
- Yang, H.; Zhang, Z.C.; Deng, C.; Gu, C.H.; Li, J.L.; Deng, C.Y. Newly recorded species of wild macrofungi in Fanjingshan National Nature Re-serve. Guizhou Sci. 2021, 39, 13–22. [Google Scholar]
- Zhang, J. Study on Macrofungi Diversity in Fanjingshan. Master’s Thesis, Guizhou Normal University, Guizhou, China, 2021. [Google Scholar]
- Wang, X.H.; Stubbe, D.; Verbeken, A. Lactifluus parvigerardii sp. nov. a new link towards the pleurotoid habit in Lactifluus subgen. Gerardii (Russulaceae, Russulales). Cryptogam. Mycol. 2012, 33, 181–190. [Google Scholar] [CrossRef]
- Zhao, Y.Z.; Sun, X.G.; Feng, J.W.; Guo, Q.Q. Diversity of edible mycorrhizal fungi in Pinus massoniana forests of central Guizhou, southwest China. Mycosystema 2021, 40, 108–123. [Google Scholar] [CrossRef]
- Ran, L.; Zhang, G.L.; Shen, S.M.; Chen, W.; Li, S.L. Extraction and antioxidant activities of polysaccharides from Lactarius volemus in Guizhou province. Food Sci. Technol. 2007, 42, 194–198. [Google Scholar] [CrossRef]
- Zhou, Q.Z.; Su, W.C. Analysis of the nutritional composition of Lactarius volemus (Fr.) in Guizhou province. Acta Nutr. Sin. 2007, 2, 169–170. [Google Scholar]
- Wang, Z.; Wang, X.H.; Qin, W.Q.; Chen, Z.H.; Deng, W.Q.; Zhang, J.H.; Bau, T. Lactarius mirus sp. nov. (Russulaceae, Russulales), the first species of Lactarius subg. Plinthogalus with yellowing latex in China. Mycosystema 2021, 40, 1591–1604. [Google Scholar] [CrossRef]
- Wu, F.; Wang, X.H.; Qin, W.Q.; Hao, Y.J.; Guo, T.; Zhang, P.; Chen, Z.H. Five new species and one new record of Lactarius (Russulaceae, Russulales) from tropical and subtropical regions of China. Mycosystema 2022, 41, 1234–1253. [Google Scholar] [CrossRef]
- Cao, S.Q.; Wang, X.H.; Qin, W.Q.; Wu, F.; Liu, T.Z.; Bau, T.; Li, S.L.; Chen, Z.H.; Liu, D.M. New species Lactarius liyuanus in East Asia: A close relative of European L. acerrimus. Mycosystema 2023, 42. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Voucher Specimen No. | Locality | ITS | LSU | RPB2 |
|---|---|---|---|---|---|
| Lactifluus subgenus Lactifluus | |||||
| Lactifluus sect. Lactifluus | |||||
| Lf. acicularis | H.T. Le 265 (CMU, GENT, MFLU, SFSU) | Thailand | HQ318277 | HQ318196 | HQ328926 |
| Lf. acicularis | KVP08002 | Thailand | HQ318226 | HQ318132 | HQ328869 |
| Lf. aff. tenuicystidiatus | KUN:F75810 | China | KC154105 | KC154131 | KC154157 |
| Lf. aff. tenuicystidiatus | KUN:F75810 | China | KC154105 | KC154131 | KC154157 |
| Lf. aff. volemus var. flavus | KVP08023 | Thailand | HQ318227 | HQ318133 | HQ328870 |
| Lf. crocatus | H.T. Le 268 (CMU, GENT, MFLU, SFSU) | Thailand | HQ318266 | HQ318181 | HQ328917 |
| Lf. dissitus | AV-KD-KVP09-134 | India | JN388978 | JN389026 | JN375628 |
| Lf. distantifolius | H.T. Le 288 (CMU, GENT, MFLU, SFSU) | Thailand | HQ318274 | HQ318193 | |
| Lf. distantifolius | DS07-461 | Thailand | HQ318223 | HQ318124 | HQ328866 |
| Lf. indovolemus | IB 18-013 | India | MN005117 | ||
| Lf. indovolemus | IB 18-003 | India | MN005115 | ||
| Lf. jianbaensis | TB 4 | China | OL423565 | OL423578 | OM030355 |
| Lf. jianbaensis | TB 5 | China | OL423566 | OL423579 | OM030356 |
| Lf. jianbaensis | TB 6 | China | OL423567 | OL423580 | OM030357 |
| Lf. lamprocystidiatus | EH 72-195 | Papua New Guinea | KR364015 | ||
| Lf. leptomerus | AV-KD-KVP 09-131 (GENT) | India | JN388972 | JN389023 | JN375625 |
| Lf. longipilus | LTH273 | Thailand | HQ318276 | HQ318195 | HQ328925 |
| Lf. longipilus | LTH184 | Thailand | HQ318256 | HQ318169 | HQ328905 |
| Lf. longipilus | AV-RW04-160 | Thailand | HQ318235 | HQ318143 | HQ328880 |
| Lf. maenamensis | KD 16-008 | India | MF928075 | ||
| Lf. mexicanus | Montoya 5189 | Mexico | MK211179 | MK211188 | MK258869 |
| Lf. mexicanus | Montoya 5266 | Mexico | MK211180 | MK211189 | MK258870 |
| Lf. oedematopus | KVP12-001 GENT neotype | Germany | KJ210065 | KJ210066 | KJ210068 |
| Lf. oedematopus | AF 2386 (BR) | Belgium | JQ753876 | JQ348324 | JQ348260 |
| Lf. oedematopus | AV07-079 GENT | Belgium | JQ753835 | JQ348270 | JQ348131 |
| Lf. pallidilamellatus | M 4716 (XAL) | Mexico | JQ753824 | JQ348268 | |
| Lf. qinggangtangensis | TB 7 | China | OL423568 | OL423581 | OM030358 |
| Lf. qinggangtangensis | TB 8 | China | OL423569 | OL655455 | |
| Lf. pinguis | AV-RW04-162 | Thailand | HQ318221 | HQ318121 | HQ328863 |
| Lf. pinguis | H.T. Le 117—Type | Thailand | HQ318211 | HQ318111 | HQ328858 |
| Lf. pinguis | H.T. Le 255 (CMU, GENT, MFLU, SFSU) | Thailand | HQ318263 | HQ318178 | HQ328914 |
| Lf. rugiformis | SFC20150818-14 | South Korea | MN215387 | MN215343 | MN212835 |
| Lf. sect. Tenuicystidiati | |||||
| Lf. species 17 | LTH214 | Thailand | HQ318249 | HQ318158 | HQ328894 |
| Lf. species 21 | AV-KD-KVP09-137 | Sikkim | JN388958 | JN389027 | JN375629 |
| Lf. species 22 | AV-KD-KVP09-129 | Sikkim | JN388957 | JN389021 | JN375623 |
| Lf. species 8a | KVP08021 | Thailand | HQ318233 | HQ318140 | HQ328877 |
| Lf. species 8a | LTH170 | Thailand | HQ318252 | HQ318165 | HQ328902 |
| Lf. subpruinosus | X.H. Wang 3489 (KUN) | China | KC154110 | KC154136 | KC154162 |
| Lf. subvolemus | KVP 08-048 | Slovenia | JQ753927 | JQ348379 | JQ348241 |
| Lf. taibaiensis | TB 1 | China | OL423562 | OL423575 | OM030352 |
| Lf. taibaiensis | TB 2 | China | OL423563 | OL423576 | OM030353 |
| Lf. taibaiensis | TB 3 | China | OL423564 | OL423577 | OM030354 |
| Lf. versiformis | AV-KD-KVP09-002 | Sikkim | JN388966 | JN389030 | JN375631 |
| Lf. versiformis | AV-KD-KVP09-006 | Sikkim | JN388965 | JN389033 | JN375633 |
| Lf. vitellinus | H.T. Le 348 (CMU, GENT, MFLU, SFSU) | Thailand | HQ318251 | HQ318164 | HQ328900 |
| Lf. vitellinus | K. Van de Putte 08-024 (GENT, MFLU) | Thailand | HQ318236 | HQ318144 | HQ328881 |
| Lf. volemus | L. Pihlik et al.—TAAM095075 | Russia | JQ753905 | JQ348357 | JQ348219 |
| Lf. volemus | Walther—STU 406307 | Germany | JQ753909 | JQ348361 | JQ348223 |
| Lf. volemus | L. Tedersoo—TAAM182733 | Estonia | JQ753907 | JQ348359 | JQ348221 |
| Lf. volemus | LE 254509 | Russia | JQ753937 | JQ348388 | |
| Lf. volemus | L. Pihlik et al.—TAAM095097 | Russia | JQ753906 | JQ348358 | JQ348220 |
| Lf. volemus | Kobeke Van de Putte 08-45 | Slovenia | JQ753953 | ||
| Lf. sect. Allardii | |||||
| Lf. allardii | J. Nuytinck 2004-008 | USA | KF220016 | KF220125 | KF220217 |
| Lf. sect. Ambicystidiati | |||||
| Lf. ambicystidiatus | KUN:F57008—Type | China | NR_155311 | NG_060287 | KC154148 |
| Lf. sect. Gerardii | |||||
| Lf. atrovelutinus | D. Stubbe 06-003 | Malaysia | GU258231 | GU265588 | GU258325 |
| Lf. bicolor | DS06-247 | Malaysia | JN388955 | JN388987 | JN375590 |
| Lf. gerardii | A.Verbeken 05-375 | USA | GU258254 | GU265616 | GU258353 |
| Lf. sect. Piperati | |||||
| Lf. aff. glaucescens | AV 05-374 | North America | KF220049 | KF220150 | KF220236 |
| Lf. aff. piperatus | A.Verbeken 04-202 | USA | KF220021 | KF220127 | KF220220 |
| Outgroup | |||||
| Auriscalpium vulgare | PBM_944 | North America | DQ911613 | DQ911614 | AY218472 |
| Bondarzewia montana | AFTOL_452 | No data | DQ200923 | DQ234539 | AY218474 |
| Stereum hirsutum | AFTOL_492 | No data | AY854063 | AF393078 | AY218520 |
| Species | Lf. rugiformis | Lf. pinguis | Lf. acicularis |
|---|---|---|---|
| Location | Korea | Thailand | Thailand |
| Pileus length (mm) | 50–110 | 35–85 | 3.3–4.6 |
| Pileus color | Rusty orange (6C8–7C8) tinged with a more brownish color | Yellowish-orange-brown (5C7–5C8), brown (6D5) | Yellowish-orange-brown (5C7–5C8), brown (6D5) |
| Lamella breadth (mm) | Three broad, rarely furcate, with numerous lamellula of different length | 2–4 | Narrow to rather broad (1.5– 6 mm) |
| Lamella color | Cream to pale orange | Whitish to cream | Cream (4A3–4A4), discoloring to brown (6D5– 6E5) to grayish-brown (5C3–5C4) when damaged |
| Stipe (mm) | 30–70 × 15–20 | 40–95 × 10–15 | 45–85 × 5–15 |
| Stipe color | Concolorous with pileus | Concolorous with pileus | Yellowish-orange (4A5–5A5),brownish-orange (6C8–6D8) to grayish-brownish-orange (6C5–5C6–6B5–6B6–6C6), or brown (6D4–6D5) |
| Latex | Abundant, sticky, white turning dark brown | Copious, sticky, white, unchanging when isolated | White |
| Basidiospores (μm) | 7.1–8.4–9.6 × 6.7–7.9–9.2, Q = 1.01–1.05–1.09, globose to subglobose | 8.0–9.0–9.1–10.2(–10.5) × 7.4–8.3–8.4– 9.4(–9.6) | 7.0–7.9–8.5–9.1(–9.3) × 6.5–7.2–7.8– 8.5, subglobose (Q = 1.01–1.08–1.10–1.21) |
| Basidia (μm) | 49.5–60 × 9–12.5 | 40–65 × 11–14 | 40–60 × 9–12 |
| Pileipellis (μm) | 20–68 × 2.5–4.0, cell wall 0.5–1.5, thick, erect | 50–140 | 50–120, thick |
| References | [21] | [15] | [15] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, X.-H.; Chen, A.-M.; Yao, N.; Wen, T.-C.; Pei, Y.; Zhang, W.-P. Three New Species of Lactifluus (Basidiomycota, Russulaceae) from Guizhou Province, Southwest China. J. Fungi 2023, 9, 122. https://doi.org/10.3390/jof9010122
Xu X-H, Chen A-M, Yao N, Wen T-C, Pei Y, Zhang W-P. Three New Species of Lactifluus (Basidiomycota, Russulaceae) from Guizhou Province, Southwest China. Journal of Fungi. 2023; 9(1):122. https://doi.org/10.3390/jof9010122
Chicago/Turabian StyleXu, Xiu-Hong, A-Min Chen, Nan Yao, Ting-Chi Wen, Yun Pei, and Wan-Ping Zhang. 2023. "Three New Species of Lactifluus (Basidiomycota, Russulaceae) from Guizhou Province, Southwest China" Journal of Fungi 9, no. 1: 122. https://doi.org/10.3390/jof9010122
APA StyleXu, X.-H., Chen, A.-M., Yao, N., Wen, T.-C., Pei, Y., & Zhang, W.-P. (2023). Three New Species of Lactifluus (Basidiomycota, Russulaceae) from Guizhou Province, Southwest China. Journal of Fungi, 9(1), 122. https://doi.org/10.3390/jof9010122