
In Vitro Characterization and Identification of Potential Probiotic Yeasts Isolated from Fermented Dairy and Non-Dairy Food Products

 , , , and
, , , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Isolation of Yeasts
2.3. Acid Tolerance: Preliminary Probiotic Investigation
2.4. Tolerance to In Vitro Digestion Conditions
2.5. Bile Salt Tolerance
2.6. Cholesterol Removal
2.7. Bile Salt Hydrolysis (BSH) Activity
2.8. Autoaggregation
2.9. Hydrophobicity
2.10. Coaggregation
2.11. Antimicrobial Activity
2.12. Antibiotic Susceptibility
2.13. Adhesion to the HT-29 Cell Line
2.14. EPS Production
2.15. Heat Resistance
2.16. Molecular Identification of the Selected Yeast Isolates
2.17. Statistical Analysis
3. Results and Discussion
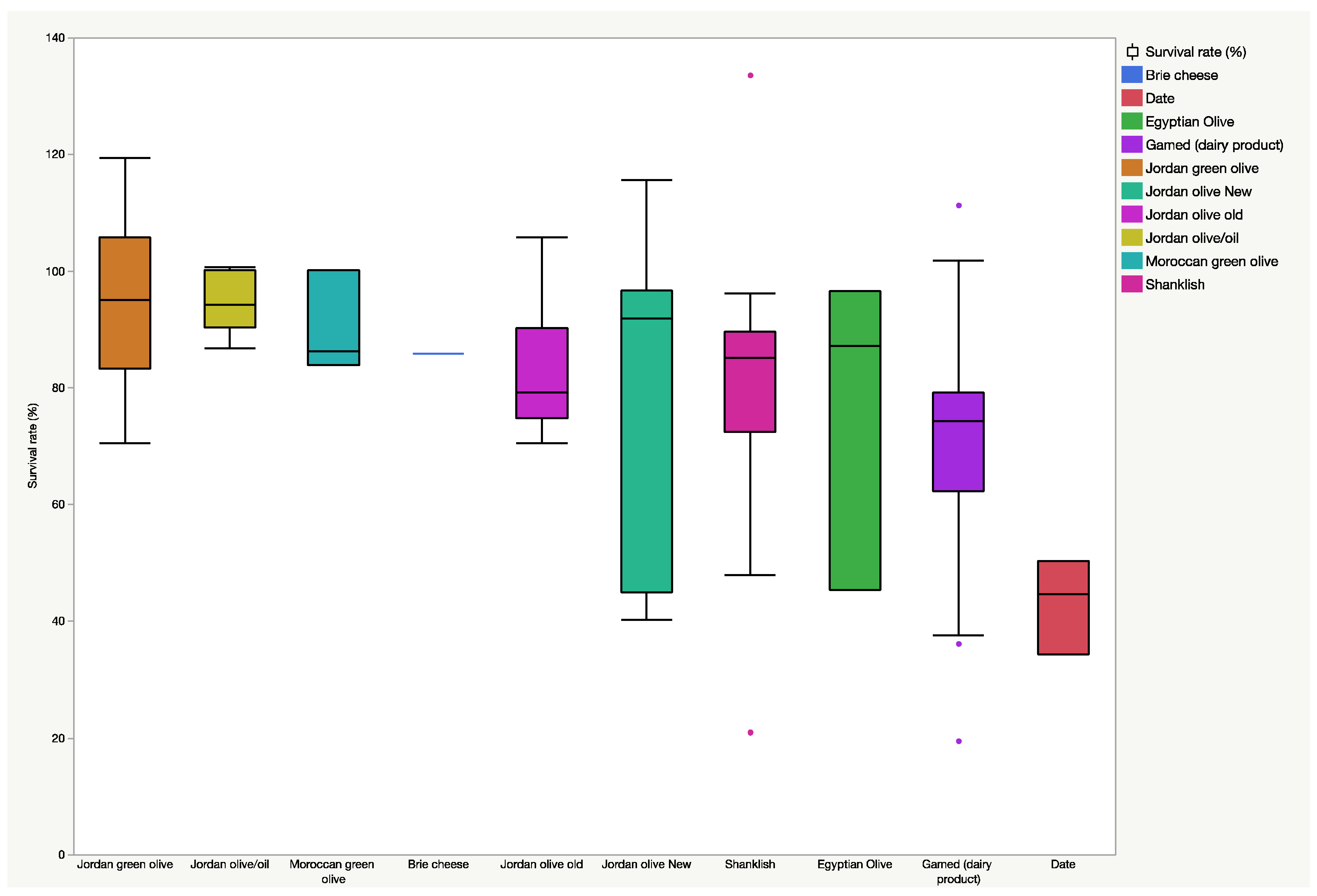
3.1. Preliminary Acid Tolerance
3.2. Tolerance to In Vitro Digestion Conditions and Bile Salts
3.3. Cholesterol Removal and Bile Salt Hydrolysis (BSH)
3.4. Autoaggregation and Hydrophobicity
3.5. Coaggregation and Antimicrobial Activity
3.6. Antibiotic Susceptibility and Attachment to the HT-29 Cell Line
3.7. EPS Production and Heat Resistance
3.8. Molecular Identification of Selected Yeast Isolates
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAO/WHO. Report of a Joint FAO/WHO Working Group on Drafting Guidelines for the Evaluation of Probiotics in Food; WHO: Geneva, Switzerland; Ottawa, ON, Canada, 2002; Available online: https://www.mhlw.go.jp/file/05-Shingikai-11121000-Iyakushokuhinkyoku-Soumuka/0000197343.pdf (accessed on 23 April 2022).
- Perricone, M.; Bevilacqua, A.; Corbo, M.R.; Sinigaglia, M. Technological characterization and probiotic traits of yeasts isolated from Altamura sourdough to select promising microorganisms as functional starter cultures for cereal-based products. Food Microbiol. 2014, 38, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Doron, S.; Snydman, D.R. Risk and Safety of Probiotics. Clin. Infect. Dis. 2015, 60, S129–S134. [Google Scholar] [CrossRef] [PubMed]
- Picard, C.; Fioramonti, J.; Francois, A.; Robinson, T.; Neant, F.; Matuchansky, C. Bifidobacteria as probiotic agents–Physiological effects and clinical benefits. Aliment. Pharmacol. Ther. 2005, 22, 495–512. [Google Scholar] [CrossRef] [PubMed]
- Satyanarayana, T.; Kunze, G. Yeast Diversity in Human Welfare; Springer: Singapore, 2017. [Google Scholar]
- Kelesidis, T.; Pothoulakis, C. Efficacy and safety of the probiotic Saccharomyces boulardii for the prevention and therapy of gastrointestinal disorders. Ther. Adv. Gastroenterol. 2012, 5, 111–125. [Google Scholar] [CrossRef] [PubMed]
- Amorim, J.C.; Piccoli, R.H.; Duarte, W.F. Probiotic potential of yeasts isolated from pineapple and their use in the elaboration of potentially functional fermented beverages. Food Res. Int. 2018, 107, 518–527. [Google Scholar] [CrossRef]
- Oliveira, T.; Ramalhosa, E.; Nunes, L.; Pereira, J.A.; Colla, E.; Pereira, E.L. Probiotic potential of indigenous yeasts isolated during the fermentation of table olives from Northeast of Portugal. Innov. Food Sci. Emerg. Technol. 2017, 44, 167–172. [Google Scholar] [CrossRef]
- Gil-Rodríguez, A.M.; Carrascosa, A.V.; Requena, T. Yeasts in foods and beverages: In vitro characterisation of probiotic traits. LWT Food Sci. Technol. 2015, 64, 1156–1162. [Google Scholar] [CrossRef]
- Lane, M.M.; Morrissey, J.P. Kluyveromyces marxianus: A yeast emerging from its sister’s shadow. Fungal Biol. Rev. 2010, 24, 17–26. [Google Scholar] [CrossRef]
- Foligne, B.; Dewulf, J.; Vandekerckove, P.; Pignede, G.; Pot, B. Probiotic yeasts: Anti-inflammatory potential of various non-pathogenic strains in experimental colitis in mice. World J. Gastroenterol. 2010, 16, 2134–2145. [Google Scholar] [CrossRef]
- Czerucka, D.; Piche, T.; Rampal, P. Review article: Yeast as probiotics—Saccharomyces boulardii. Aliment. Pharmacol. Ther. 2007, 26, 767–778. [Google Scholar] [CrossRef]
- Arévalo-Villena, M.; Fernandez-Pacheco, P.; Castillo, N.; Bevilacqua, A.; Briones Pérez, A. Probiotic capability in yeasts: Set-up of a screening method. LWT 2018, 89, 657–665. [Google Scholar] [CrossRef]
- Fadda, M.E.; Mossa, V.; Deplano, M.; Pisano, M.B.; Cosentino, S. In vitro screening of Kluyveromyces strains isolated from Fiore Sardo cheese for potential use as probiotics. LWT 2017, 75, 100–106. [Google Scholar] [CrossRef]
- Abdel-Rahman, M.A.; Tashiro, Y.; Sonomoto, K. Recent advances in lactic acid production by microbial fermentation processes. Biotechnol. Adv. 2013, 31, 877–902. [Google Scholar] [CrossRef] [PubMed]
- Fleet, G.H. Chapter 5—Yeast Spoilage of Foods and Beverages. In The Yeasts, 5th ed.; Kurtzman, C.P., Fell, J.W., Boekhout, T., Eds.; Elsevier: London, UK, 2011; pp. 53–63. [Google Scholar]
- Joshi, V.S.; Thorat, B.N. Formulation and Cost-Effective Drying of Probiotic Yeast. Dry Technol. 2011, 29, 749–757. [Google Scholar] [CrossRef]
- Hatoum, R.; Labrie, S.; Fliss, I. Antimicrobial and probiotic properties of yeasts: From fundamental to novel applications. Front. Microbiol. 2012, 3, 421. [Google Scholar] [CrossRef] [PubMed]
- Arroyo López, F.N.; Romero Gil, V.; Bautista Gallego, J.; Rodriguez Gomez, F.; Jimenez Diaz, R.; García García, P.; Querol Simon, A.; Garrido Fernandez, A. Potential benefits of the application of yeast starters in table olive processing. Front. Microbiol. 2012, 3, 161. [Google Scholar] [CrossRef] [PubMed]
- Moreira, N.; Pina, C.; Mendes, F.; Couto, J.A.; Hogg, T.; Vasconcelos, I. Volatile compounds contribution of Hanseniaspora guilliermondii and Hanseniaspora uvarum during red wine vinifications. Food Control 2011, 22, 662–667. [Google Scholar] [CrossRef]
- Ogunremi, O.R.; Agrawal, R.; Sanni, A.I. Development of cereal-based functional food using cereal-mix substrate fermented with probiotic strain—Pichia kudriavzevii OG32. Food Sci. Nutr. 2015, 3, 486–494. [Google Scholar] [CrossRef]
- Sarwar, A.; Aziz, T.; Al-Dalali, S.; Zhao, X.; Zhang, J.; Ud Din, J.; Chen, C.; Cao, Y.; Yang, Z. Physicochemical and microbiological properties of synbiotic yogurt made with probiotic yeast saccharomyces boulardii in combination with inulin. Foods 2019, 8, 468. [Google Scholar] [CrossRef]
- Binetti, A.; Carrasco, M.; Reinheimer, J.; Suárez, V. Yeasts from autochthonal cheese starters: Technological and functional properties. J. Appl. Microbiol. 2013, 115, 434–444. [Google Scholar] [CrossRef]
- Lowes, K.F.; Shearman, C.A.; Payne, J.; MacKenzie, D.; Archer, D.B.; Merry, R.J.; Gasson, M.J. Prevention of Yeast Spoilage in Feed and Food by the Yeast Mycocin HMK. Appl. Environ. Microbiol. 2000, 66, 1066–1076. [Google Scholar] [CrossRef] [PubMed]
- Antunes, J.; Aguiar, C. Search for killer phenotypes with potential for biological control. Ann. Microbiol. 2012, 62, 427–433. [Google Scholar] [CrossRef]
- Vera-Pingitore, E.; Jimenez, M.E.; Dallagnol, A.; Belfiore, C.; Fontana, C.; Fontana, P.; von Wright, A.; Vignolo, G.; Plumed-Ferrer, C. Screening and characterization of potential probiotic and starter bacteria for plant fermentations. LWT Food Sci. Technol. 2016, 71, 288–294. [Google Scholar] [CrossRef]
- Silambarasan, S.; Logeswari, P.; Cornejo, P.; Kannan, V.R. Evaluation of the production of exopolysaccharide by plant growth promoting yeast Rhodotorula sp. strain CAH2 under abiotic stress conditions. Int. J. Biol. Macromol. 2019, 121, 55–62. [Google Scholar] [CrossRef] [PubMed]
- De Melo Pereira, G.V.; de Oliveira Coelho, B.; Magalhães Júnior, A.I.; Thomaz-Soccol, V.; Soccol, C.R. How to select a probiotic? A review and update of methods and criteria. Biotechnol. Adv. 2018, 36, 2060–2076. [Google Scholar] [CrossRef] [PubMed]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assunção, R.; Ballance, S.; Bohn, T.; Bourlieu-Lacanal, C.; Boutrou, R.; Carrière, F. INFOGEST static in vitro simulation of gastrointestinal food digestion. Nat. Protoc. 2019, 14, 991–1014. [Google Scholar] [CrossRef]
- AlKalbani, N.S.; Turner, M.S.; Ayyash, M.M.J.M.C.F. Isolation, identification, and potential probiotic characterization of isolated lactic acid bacteria and in vitro investigation of the cytotoxicity, antioxidant, and antidiabetic activities in fermented sausage. Microb. Cell Factories 2019, 18, 188. [Google Scholar] [CrossRef]
- Alameri, F.; Tarique, M.; Osaili, T.; Obaid, R.; Abdalla, A.; Masad, R.; Al-Sbiei, A.; Fernandez-Cabezudo, M.; Liu, S.Q.; Al-Ramadi, B.; et al. Lactic Acid Bacteria Isolated from Fresh Vegetable Products: Potential Probiotic and Postbiotic Characteristics Including Immunomodulatory Effects. Microorganisms 2022, 10, 389. [Google Scholar] [CrossRef]
- Bonatsou, S.; Karamouza, M.; Zoumpopoulou, G.; Mavrogonatou, E.; Kletsas, D.; Papadimitriou, K.; Tsakalidou, E.; Nychas, G.E.; Panagou, E. Evaluating the probiotic potential and technological characteristics of yeasts implicated in cv. Kalamata natural black olive fermentation. Int. J. Food Microbiol. 2018, 271, 48–59. [Google Scholar] [CrossRef]
- Andrade, G.C.; Andrade, R.P.; Oliveira, D.R.; Quintanilha, M.F.; Martins, F.S.; Duarte, W.F. Kluyveromyces lactis and Torulaspora delbrueckii: Probiotic characterization, anti-Salmonella effect, and impact on cheese quality. LWT 2021, 151, 112240. [Google Scholar] [CrossRef]
- Hossain, M.N.; Afrin, S.; Humayun, S.; Ahmed, M.M.; Saha, B.K. Identification and Growth Characterization of a Novel Strain of Saccharomyces boulardii Isolated From Soya Paste. Front. Nutr. 2020, 7, 27. [Google Scholar] [CrossRef] [PubMed]
- Tarique, M.; Abdalla, A.; Masad, R.; Al-Sbiei, A.; Kizhakkayil, J.; Osaili, T.; Olaimat, A.; Liu, S.-Q.; Fernandez-Cabezudo, M.; al-Ramadi, B.; et al. Potential probiotics and postbiotic characteristics including immunomodulatory effects of lactic acid bacteria isolated from traditional yogurt-like products. LWT 2022, 159, 113207. [Google Scholar] [CrossRef]
- Charteris, W.P.; Kelly, P.M.; Morelli, L.; Collins, J.K. Antibiotic susceptibility of potentially probiotic Lactobacillus species. J. Food Prot. 1998, 61, 1636–1643. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.Y.; Son, S.H.; Hong, S.P.; Yi, S.H.; Kang, S.H.; Lee, N.K.; Paik, H.D. Production of β-glucan, glutathione, and glutathione derivatives by probiotic Saccharomyces cerevisiae isolated from cucumber jangajji. LWT 2019, 100, 114–118. [Google Scholar] [CrossRef]
- Angmo, K.; Kumari, A.; Savitri; Bhalla, T.C. Probiotic characterization of lactic acid bacteria isolated from fermented foods and beverage of Ladakh. LWT Food Sci. Technol. 2016, 66, 428–435. [Google Scholar] [CrossRef]
- Teles Santos, T.; Santos Ornellas, R.M.; Borges Arcucio, L.; Messias Oliveira, M.; Nicoli, J.R.; Villela Dias, C.; Trovatti Uetanabaro, A.P.; Vinderola, G. Characterization of lactobacilli strains derived from cocoa fermentation in the south of Bahia for the development of probiotic cultures. LWT 2016, 73, 259–266. [Google Scholar] [CrossRef]
- Lara-Hidalgo, C.; Dorantes-Álvarez, L.; Hernández-Sánchez, H.; Santoyo-Tepole, F.; Martínez-Torres, A.; Villa-Tanaca, L.; Hernández-Rodríguez, C. Isolation of yeasts from guajillo pepper (Capsicum annuum L.) fermentation and study of some probiotic characteristics. Probiotics Antimicrob. Proteins 2019, 11, 748–764. [Google Scholar] [CrossRef]
- Franco-Duarte, R.; Mendes, I.; Gomes, A.C.; Santos, M.A.; de Sousa, B.; Schuller, D. Genotyping of Saccharomyces cerevisiae strains by interdelta sequence typing using automated microfluidics. Electrophoresis 2011, 32, 1447–1455. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Kapteyn, J.; Ter Riet, B.; Vink, E.; Blad, S.; De Nobel, H.; Van Den Ende, H.; Klis, F.J.M.M. Low external pH induces HOG1-dependent changes in the organization of the Saccharomyces cerevisiae cell wall. Mol. Microbiol. 2001, 39, 469–480. [Google Scholar] [CrossRef] [PubMed]
- Lucena, R.M.; Dolz-Edo, L.; Brul, S.; de Morais, M.A.; Smits, G.J.G. Extreme Low Cytosolic pH Is a Signal for Cell Survival in Acid Stressed Yeast. Genes 2020, 11, 656. [Google Scholar] [CrossRef] [PubMed]
- Santos, C.C.A.D.A.; Almeida, E.G.D.; Melo, G.V.P.D.; Schwan, R.F. Microbiological and physicochemical characterisation of caxiri, an alcoholic beverage produced by the indigenous Juruna people of Brazil. Int. J. Food Microbiol. 2012, 156, 112–121. [Google Scholar] [CrossRef] [PubMed]
- Moreira, I.M.D.V.; Miguel, M.G.D.C.P.; Duarte, W.F.; Dias, D.R.; Schwan, R.F. Microbial succession and the dynamics of metabolites and sugars during the fermentation of three different cocoa (Theobroma cacao L.) hybrids. Food Res. Int. 2013, 54, 9–17. [Google Scholar] [CrossRef]
- Şanlidere Aloğlu, H.; Demir Özer, E.; Öner, Z. Assimilation of cholesterol and probiotic characterisation of yeast strains isolated from raw milk and fermented foods. Int. J. Dairy Technol. 2016, 69, 63–70. [Google Scholar] [CrossRef]
- Uymaz Tezel, B.; Şanlıbaba, P.; Akçelik, N.; Akçelik, M. Chapter 2—Selection Criteria for Identifying Putative Probiont. In Advances in Probiotics; Dhanasekaran, D., Sankaranarayanan, A., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 23–35. [Google Scholar]
- Walker, G.M. Yeasts. In Encyclopedia of Microbiology, 3rd ed.; Schaechter, M., Ed.; Academic Press: Oxford, UK, 2009; pp. 478–491. [Google Scholar]
- Ayyash, M.M.; Abdalla, A.K.; AlKalbani, N.S.; Baig, M.A.; Turner, M.S.; Liu, S.-Q.; Shah, N.P.J.J.o.D.S. Invited review: Characterization of new probiotics from dairy and nondairy products—Insights into acid tolerance, bile metabolism and tolerance, and adhesion capability. J. Dairy Sci. 2021, 104, 8363–8379. [Google Scholar] [CrossRef] [PubMed]
- Zamith-Miranda, D.; Palma, M.L.; Matos, G.S.; Schiebel, J.G.; Maya-Monteiro, C.M.; Aronovich, M.; Bozza, P.T.; Bozza, F.A.; Nimrichter, L.; Montero-Lomeli, M.; et al. Lipid droplet levels vary heterogeneously in response to simulated gastrointestinal stresses in different probiotic Saccharomyces cerevisiae strains. J. Funct. Foods 2016, 21, 193–200. [Google Scholar] [CrossRef]
- Palma, M.L.; Zamith-Miranda, D.; Martins, F.S.; Bozza, F.A.; Nimrichter, L.; Montero-Lomeli, M.; Marques, E.T.A.; Douradinha, B. Probiotic Saccharomyces cerevisiae strains as biotherapeutic tools: Is there room for improvement? Appl. Microbiol. Biotechnol. 2015, 99, 6563–6570. [Google Scholar] [CrossRef]
- Greppi, A.; Saubade, F.; Botta, C.; Humblot, C.; Guyot, J.P.; Cocolin, L. Potential probiotic Pichia kudriavzevii strains and their ability to enhance folate content of traditional cereal-based African fermented food. Food Microbiol. 2017, 62, 169–177. [Google Scholar] [CrossRef]
- Goktas, H.; Dertli, E.; Sagdic, O. Comparison of functional characteristics of distinct Saccharomyces boulardii strains isolated from commercial food supplements. LWT 2021, 136, 110340. [Google Scholar] [CrossRef]
- Chen, L.S.; Ma, Y.; Maubois, J.L.; Chen, L.J.; Liu, Q.H.; Guo, J.P. Identifcation of yeasts from raw milk and selection for some specific antioxidant properties. Int. J. Dairy Technol. 2010, 63, 47–54. [Google Scholar] [CrossRef]
- Menezes, A.G.T.; Ramos, C.L.; Cenzi, G.; Melo, D.S.; Dias, D.R.; Schwan, R.F. Probiotic Potential, Antioxidant Activity, and Phytase Production of Indigenous Yeasts Isolated from Indigenous Fermented Foods. Probiotics Antimicrob. Proteins 2020, 12, 280–288. [Google Scholar] [CrossRef] [PubMed]
- Ayyash, M.; Abushelaibi, A.; Al-Mahadin, S.; Enan, M.; El-Tarabily, K.; Shah, N. In-vitro investigation into probiotic characterisation of Streptococcus and Enterococcus isolated from camel milk. LWT Food Sci. Technol. 2018, 87, 478–487. [Google Scholar] [CrossRef]
- Ishimwe, N.; Daliri, E.B.; Lee, B.H.; Fang, F.; Du, G. The perspective on cholesterol-lowering mechanisms of probiotics. Mol. Nutr. Food Res. 2015, 59, 94–105. [Google Scholar] [CrossRef]
- Chen, L.S.; Ma, Y.; Maubois, J.L.; He, S.H.; Chen, L.J.; Li, H.M. Screening for the potential probiotic yeast strains from raw milk to assimilate cholesterol. Dairy Sci. Technol. 2010, 90, 537–548. [Google Scholar] [CrossRef]
- Porru, C.; Rodriguez-Gomez, F.; Benitez-Cabello, A.; Jimenez-Diaz, R.; Zara, G.; Budroni, M.; Mannazzu, I.; Arroyo-Lopez, F.N. Genotyping, identification and multifunctional features of yeasts associated to Bosana naturally black table olive fermentations. Food Microbiol. 2018, 69, 33–42. [Google Scholar] [CrossRef]
- Zullo, B.; Ciafardini, G.J.F.m. Evaluation of physiological properties of yeast strains isolated from olive oil and their in vitro probiotic trait. Food Microbiol. 2019, 78, 179–187. [Google Scholar] [CrossRef]
- Allain, T.; Chaouch, S.; Thomas, M.; Vallée, I.; Buret, A.G.; Langella, P.; Grellier, P.; Polack, B.; Bermúdez-Humarán, L.G.; Florent, I. Bile-Salt-Hydrolases from the probiotic strain Lactobacillus johnsonii La1 mediate anti-giardial activity in vitro and in vivo. Front. Microbiol. 2018, 8, 2707. [Google Scholar] [CrossRef]
- Ruiz, L.; Margolles, A.; Sánchez, B. Bile resistance mechanisms in Lactobacillus and Bifidobacterium. Front. Microbiol. 2013, 4, 396. [Google Scholar] [CrossRef]
- Merchán, A.V.; Benito, M.J.; Galván, A.I.; Ruiz-Moyano Seco de Herrera, S. Identification and selection of yeast with functional properties for future application in soft paste cheese. LWT 2020, 124, 109173. [Google Scholar] [CrossRef]
- Abushelaibi, A.; Al-Mahadin, S.; El-Tarabily, K.; Shah, N.P.; Ayyash, M. Characterization of potential probiotic lactic acid bacteria isolated from camel milk. LWT Food Sci. Technol. 2017, 79, 316–325. [Google Scholar] [CrossRef]
- Verstrepen, K.J.; Klis, F.M. Flocculation, adhesion and biofilm formation in yeasts. Mol. Microbiol. 2006, 60, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Kumura, H.; Tanoue, Y.; Tsukahara, M.; Tanaka, T.; Shimazaki, K. Screening of dairy yeast strains for probiotic applications. J. Dairy Sci. 2004, 87, 4050–4056. [Google Scholar] [CrossRef]
- Burns, P.; Reinheimer, J.; Vinderola, G. Impact of bile salt adaptation of Lactobacillus delbrueckii subsp. lactis 200 on its interaction capacity with the gut. Res. Microbiol. 2011, 162, 782–790. [Google Scholar] [CrossRef]
- Pericolini, E.; Gabrielli, E.; Ballet, N.; Sabbatini, S.; Roselletti, E.; Cayzeele Decherf, A.; Pélerin, F.; Luciano, E.; Perito, S.; Jüsten, P.; et al. Therapeutic activity of a Saccharomyces cerevisiae-based probiotic and inactivated whole yeast on vaginal candidiasis. Virulence 2017, 8, 74–90. [Google Scholar] [CrossRef]
- Millsap, K.W.; Van Der Mei, H.C.; Bos, R.; Busscher, H.J. Adhesive interactions between medically important yeasts and bacteria. FEMS Microbiol. Rev. 1998, 21, 321–336. [Google Scholar] [CrossRef]
- Gedek, B.R. Adherence of Escherichia coli serogroup 0 157 and the Salmonella Typhimurium mutant DT 104 to the surface of Saccharomyces boulardii. Mycoses 1999, 42, 261–264. [Google Scholar] [CrossRef]
- Pontier-Bres, R.; Munro, P.; Boyer, L.; Anty, R.; Imbert, V.; Terciolo, C.; André, F.; Rampal, P.; Lemichez, E.; Peyron, J.F.; et al. Saccharomyces boulardii modifies Salmonella typhimurium traffic and host immune responses along the intestinal tract. PLoS ONE 2014, 9, e103069. [Google Scholar] [CrossRef]
- Caruffo, M.; Navarrete, N.C.; Salgado, O.A.; Faúndez, N.B.; Gajardo, M.C.; Feijóo, C.G.; Reyes-Jara, A.; García, K.; Navarrete, P. Protective yeasts control V. anguillarum pathogenicity and modulate the innate immune response of challenged zebrafish (Danio rerio) larvae. Front. Cell. Infect. Microbiol. 2016, 6, 127. [Google Scholar] [CrossRef]
- Fijan, S.J.P. Antimicrobial effect of probiotics against common pathogens. In Probiotics and Prebiotics in Human Nutrition Health; Rao, V.R.A.L.G., Ed.; IntechOpen: London, UK, 2016; Volume 10, p. 5772. [Google Scholar]
- Golubev, W. Antagonistic interactions among yeasts. In Biodiversity and Ecophysiology of Yeasts; Springer: Cham, Switzerland, 2006; pp. 197–219. [Google Scholar]
- Young, T.W.; Yagiu, M. A comparison of the killer character in different yeasts and its classification. Antonie Leeuwenhoek 1978, 44, 59–77. [Google Scholar] [CrossRef]
- Pais, P.; Almeida, V.; Yılmaz, M.; Teixeira, M.C. Saccharomyces boulardii: What makes it tick as successful probiotic? J. Fungi 2020, 6, 78. [Google Scholar] [CrossRef] [PubMed]
- Belda, I.; Ruiz, J.; Alonso, A.; Marquina, D.; Santos, A. The biology of pichia membranifaciens killer toxins. Toxins 2017, 9, 122. [Google Scholar] [CrossRef] [PubMed]
- Blum, S.; Reniero, R.; Schiffrin, E.J.; Crittenden, R.; Mattila-Sandholm, T.; Ouwehand, A.C.; Salminen, S.; Von Wright, A.; Saarela, M.; Saxelin, M.; et al. Adhesion studies for probiotics: Need for validation and refinement. Trends Food Sci. Technol. 1999, 10, 405–410. [Google Scholar] [CrossRef]
- Isolauri, E.; Salminen, S.; Mattila-Sandholm, T. New functional foods in the treatment of food allergy. Ann. Med. 1999, 31, 299–302. [Google Scholar] [CrossRef] [PubMed]
- Åvall-Jääskeläinen, S.; Lindholm, A.; Palva, A. Surface display of the receptor-binding region of the Lactobacillus brevis S-layer protein in Lactococcus lactis provides nonadhesive lactococci with the ability to adhere to intestinal epithelial cells. Appl. Environ. Microbiol. 2003, 69, 2230–2236. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Simões, L.A.; Cristina de Souza, A.; Ferreira, I.; Melo, D.S.; Lopes, L.A.A.; Magnani, M.; Schwan, R.F.; Dias, D.R. Probiotic properties of yeasts isolated from Brazilian fermented table olives. J. Appl. Microbiol. 2021, 131, 1983–1997. [Google Scholar] [CrossRef] [PubMed]
- Schmid, J.; Fariña, J.; Rehm, B.; Sieber, V. Editorial: Microbial exopolysaccharides: From genes to applications. Front. Microbiol. 2016, 7, 308. [Google Scholar] [CrossRef]
- Costa, O.Y.A.; Raaijmakers, J.M.; Kuramae, E.E. Microbial Extracellular Polymeric Substances: Ecological Function and Impact on Soil Aggregation. Front. Microbiol. 2018, 9, 1636. [Google Scholar] [CrossRef]
- Rahbar Saadat, Y.; Yari Khosroushahi, A.; Movassaghpour, A.A.; Talebi, M.; Pourghassem Gargari, B. Modulatory role of exopolysaccharides of Kluyveromyces marxianus and Pichia kudriavzevii as probiotic yeasts from dairy products in human colon cancer cells. J. Funct. Foods 2020, 64, 103675. [Google Scholar] [CrossRef]
- Fekri, A.; Torbati, M.; Yari Khosrowshahi, A.; Bagherpour Shamloo, H.; Azadmard-Damirchi, S. Functional effects of phytate-degrading, probiotic lactic acid bacteria and yeast strains isolated from Iranian traditional sourdough on the technological and nutritional properties of whole wheat bread. Food Chem. 2020, 306, 125620. [Google Scholar] [CrossRef]
- Singer, M.A.; Lindquist, S. Thermotolerance in Saccharomyces cerevisiae: The Yin and Yang of trehalose. Trends Biotechnol. 1998, 16, 460–468. [Google Scholar] [CrossRef]
- Gut, A.M.; Vasiljevic, T.; Yeager, T.; Donkor, O.N. Characterization of yeasts isolated from traditional kefir grains for potential probiotic properties. J. Funct. Foods 2019, 58, 56–66. [Google Scholar] [CrossRef]
- Parafati, L.; Palmeri, R.; Pitino, I.; Restuccia, C. Killer yeasts isolated from olive brines: Technological and probiotic aptitudes. Food Microbiol. 2022, 103, 103950. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.Q.; Liu, Q.; Hu, J.P.; Zhou, J.J.; Zhang, X.; Peng, S.Y.; Peng, L.J.; Wang, X.D. Identification and characterization of probiotic yeast isolated from digestive tract of ducks. Poult. Sci. 2018, 97, 2902–2908. [Google Scholar] [CrossRef]
- Kalyuzhin, V.A. Heat resistance in Saccharomyces cerevisiae yeast. Biol. Bull. Rev. 2011, 1, 207–213. [Google Scholar] [CrossRef]
- Franco-Duarte, R.; Mendes, I.; Umek, L.; Drumonde-Neves, J.; Zupan, B.; Schuller, D. Computational models reveal genotype-phenotype associations in Saccharomyces cerevisiae. Yeast 2014, 31, 265–277. [Google Scholar] [CrossRef]
- Franco-Duarte, R.; Bigey, F.; Carreto, L.; Mendes, I.; Dequin, S.; Santos, M.A.; Pais, C.; Schuller, D. Intrastrain genomic and phenotypic variability of the commercialSaccharomyces cerevisiaestrain Zymaflore VL1 reveals microevolutionary adaptation to vineyard environments. FEMS Yeast Res. 2015, 15, fov063. [Google Scholar] [CrossRef][Green Version]



{kind=link}
{kind=link}
| Isolate | Tolerance to GIT | Bile Salt Tolerances (%) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 6 h | 24 h | ||||||||
| Before | After | Log Reduction | 0.3 CA | 1.0 TA | 1.0 OX | 0.3 CA | 1.0 TA | 1.0 OX | |
| G.1 | 7.3 ± 0.01 | 5.5 ± 0.03 | 1.8 | 54.2 | 36.9 | 74.1 | 68.3 | 81.4 | 89.1 |
| G.2 | 7.5 ± 0.09 | 5.4 ± 0.02 | 2.1 | 53.7 | 36.1 | 74.8 | 66.7 | 73.5 | 88.1 |
| G.3 | 7.6 ± 0.24 | 6.2 ± 0.12 | 1.4 | 71.6 | 52.5 | 70.7 | 84.3 | 82.1 | 83.3 |
| G.6 | 7.4 ± 0.09 | 6.3 ± 0.11 | 1.1 | 73.8 | 61.6 | 77.2 | 78.9 | 81.1 | 87.5 |
| G.7 | 7.4 ± 0.13 | 6.1 ± 0.12 | 1.3 | 66.4 | 68.6 | 79.7 | 80.8 | 83.4 | 88.0 |
| G.8 | 7.5 ± 0.11 | 6.1 ± 0.06 | 1.4 | 71.0 | 81.7 | 79.6 | 83.8 | 85.6 | 88.3 |
| G.9 | 7.5 ± 0.07 | 6.2 ± 0.10 | 1.3 | 67.8 | 76.8 | 80.0 | 79.9 | 81.5 | 84.5 |
| G.10 | 7.5 ± 0.02 | 6.2 ± 0.02 | 1.4 | 70.5 | 64.7 | 80.8 | 83.0 | 80.7 | 87.6 |
| O.12 | 7.3 ± 0.06 | 6.3 ± 0.04 | 0.9 | 80.5 | 78.9 | 81.0 | 87.4 | 84.1 | 87.4 |
| O.13 | 7.5 ± 0.06 | 6.2 ± 0.03 | 1.3 | 72.8 | 68.2 | 79.2 | 80.5 | 81.1 | 87.1 |
| O.18 | 7.5 ± 0.01 | 6.3 ± 0.02 | 1.2 | 67.0 | 80.0 | 84.8 | 79.9 | 81.4 | 88.7 |
| O.19 | 7.4 ± 0.04 | 6.1 ± 0.03 | 1.3 | 69.0 | 45.9 | 85.8 | 78.2 | 58.0 | 86.2 |
| O.20 | 7.5 ± 0.02 | 6.3 ± 0.01 | 1.2 | 79.8 | 42.4 | 80.6 | 86.1 | 81.5 | 87.1 |
| O.21 | 7.6 ± 0.19 | 6.4 ± 0.20 | 1.3 | 82.2 | 59.2 | 86.3 | 86.9 | 80.7 | 89.1 |
| O.22 | 7.5 ± 0.04 | 6.3 ± 0.05 | 1.2 | 82.9 | 76.7 | 84.2 | 87.5 | 79.4 | 87.9 |
| O.23 | 7.4 ± 0.08 | 6.2 ± 0.03 | 1.2 | 73.7 | 67.8 | 83.8 | 79.8 | 75.6 | 88.0 |
| O.24 | 7.5 ± 0.03 | 6.5 ± 0.09 | 1.0 | 84.5 | 70.3 | 86.7 | 87.9 | 83.7 | 91.1 |
| O.26 | 7.4 ± 0.06 | 6.2 ± 0.09 | 1.2 | 80.3 | 69.5 | 82.0 | 83.4 | 77.9 | 88.2 |
| O.30 | 7.5 ± 0.01 | 6.3 ± 0.10 | 1.2 | 67.0 | 61.6 | 79.7 | 78.7 | 75.4 | 86.0 |
| O.33 | 7.4 ± 0.03 | 6.4 ± 0.06 | 0.9 | 73.9 | 62.7 | 80.5 | 83.1 | 69.9 | 83.9 |
| O.36 | 7.4 ± 0.05 | 6.2 ± 0.01 | 1.3 | 84.3 | 79.2 | 84.3 | 87.7 | 85.7 | 89.8 |
| SH.40 | 7.4 ± 0.08 | 6.6 ± 0.09 | 0.9 | 73.7 | 63.4 | 81.3 | 81.4 | 77.9 | 86.9 |
| SH.45 | 7.1 ± 0.02 | 6.1 ± 0.04 | 1.0 | 65.1 | 62.7 | 84.0 | 84.6 | 74.4 | 86.1 |
| SH.46 | 7.2 ± 0.10 | 6.3 ± 0.11 | 0.9 | 70.5 | 63.4 | 82.2 | 85.0 | 76.6 | 90.7 |
| SH.55 | 7.0 ± 0.24 | 6.0 ± 0.16 | 1.0 | 73.2 | 68.2 | 72.7 | 86.0 | 80.9 | 84.4 |
| O.63 | 7.1 ± 0.12 | 6.4 ± 0.08 | 0.7 | 64.2 | 64.8 | 81.4 | 66.9 | 65.9 | 86.4 |
| O.65 | 7.2 ± 0.00 | 6.3 ± 0.04 | 1.0 | 68.0 | 62.7 | 81.4 | 77.1 | 75.1 | 86.7 |
| G.69 | 7.3 ± 0.06 | 6.3 ± 0.06 | 1.0 | 43.8 | 17.4 | 68.6 | 48.9 | 26.5 | 69.8 |
| O.66 | 7.2 ± 0.15 | 5.4 ± 0.07 | 1.8 | 53.1 | 51.7 | 69.2 | 57.5 | 66.0 | 69.9 |
| G.75 | 7.4 ± 0.20 | 5.9 ± 0.11 | 1.5 | 52.5 | 57.0 | 69.4 | 57.5 | 61.3 | 70.3 |
| G.71 | 7.5 ± 0.13 | 6.2 ± 0.16 | 1.3 | 76.4 | 70.4 | 72.7 | 86.6 | 82.7 | 83.3 |
| G.77 | 7.2 ± 0.17 | 5.8 ± 0.12 | 1.4 | 78.5 | 75.7 | 76.6 | 87.2 | 87.3 | 85.6 |
| G.78 | 7.1 ± 0.06 | 6.1 ± 0.04 | 1.0 | 81.7 | 72.7 | 77.3 | 88.1 | 85.5 | 86.1 |
| G.80 | 7.2 ± 0.21 | 5.9 ± 0.10 | 1.3 | 67.2 | 78.0 | 76.5 | 80.3 | 84.3 | 86.4 |
| G.82 | 7.4 ± 0.09 | 5.9 ± 0.06 | 1.5 | 78.1 | 70.1 | 75.5 | 85.9 | 81.1 | 84.8 |
| G.84 | 7.2 ± 0.08 | 5.9 ± 0.04 | 1.3 | 78.9 | 70.7 | 72.0 | 87.8 | 80.8 | 86.0 |
| SH.96 | 7.4 ± 0.17 | 5.8 ± 0.14 | 1.6 | 86.8 | 84.8 | 75.7 | 89.7 | 89.2 | 90.1 |
| SH.97 | 7.4 ± 0.08 | 6.2 ± 0.04 | 1.3 | 85.9 | 84.8 | 78.1 | 90.2 | 89.3 | 89.8 |
| SH.98 | 7.2 ± 0.23 | 5.7 ± 0.09 | 1.5 | 87.3 | 82.4 | 71.9 | 89.5 | 88.8 | 81.9 |
| SH.99 | 7.3 ± 0.32 | 6.3 ± 0.27 | 1.0 | 84.9 | 77.3 | 68.4 | 89.6 | 89.0 | 69.2 |
| SH.100 | 7.4 ± 0.13 | 5.9 ± 0.18 | 1.5 | 84.9 | 83.0 | 81.1 | 89.8 | 89.1 | 90.3 |
| SH.102 | 7.3 ± 0.13 | 5.6 ± 0.07 | 1.7 | 83.9 | 85.7 | 73.3 | 89.4 | 88.7 | 89.4 |
| SH.103 | 7.1 ± 0.19 | 5.5 ± 0.17 | 1.6 | 87.9 | 81.7 | 79.9 | 90.8 | 89.5 | 90.3 |
| SH.104 | 7.4 ± 0.30 | 6.2 ± 0.22 | 1.2 | 86.6 | 79.8 | 80.0 | 90.5 | 89.4 | 90.6 |
| SH.105 | 7.4 ± 0.14 | 6.1 ± 0.08 | 1.3 | 86.3 | 70.5 | 74.8 | 89.4 | 87.8 | 89.4 |
| Isolate | CR (%) | BSH | |||||
|---|---|---|---|---|---|---|---|
| Na-SG | SA | Na-TA | SA | Bile salt mixture | SA | ||
| G1 | 47.98 ± 7.55 ab | 1.79 ± 0.05 abc | 3.70 | 1.83 ± 0.07 bc | 3.79 | 1.72 ± 0.05 a | 3.56 |
| O12 | 50.16 ± 8.68 ab | 1.80 ± 0.07 bc | 3.68 | 1.72 ± 0.07 ab | 3.52 | 1.84 ± 0.07 bc | 3.77 |
| O13 | 71.96 ± 5.20 d | 2.13 ± 0.10 e | 4.46 | 1.88 ± 0.04 c | 3.93 | 1.73 ± 0.07 a | 3.62 |
| O18 | 62.31 ± 2.35 cd | 1.90 ± 0.06 d | 3.85 | 1.72 ± 0.04 ab | 3.49 | 2.11 ± 0.08 d | 4.27 |
| O21 | 95.02 ± 1.43 e | 1.87 ± 0.03 cd | 4.01 | 1.70 ± 0.06 a | 3.65 | 2.22 ± 0.05 e | 4.77 |
| O26 | 91.59 ± 2.47 e | 2.17 ± 0.03 ef | 4.55 | 1.91 ± 0.02 c | 4.01 | 2.26 ± 0.04 e | 4.73 |
| O36 | 53.58 ± 1.08 bc | 1.89 ± 0.02 d | 3.95 | 1.82 ± 0.05 abc | 3.81 | 1.92 ± 0.05 c | 4.02 |
| O63 | 47.98 ± 1.95 ab | 1.74 ± 0.04 ab | 3.57 | 1.71 ± 0.05 ab | 3.50 | 1.89 ± 0.04 bc | 3.87 |
| O66 | 65.42 ± 2.80 cd | 1.94 ± 0.04 d | 4.04 | 1.88 ± 0.02 c | 3.90 | 2.04 ± 0.05 d | 4.25 |
| SH40 | 39.56 ± 2.86 a | 1.76 ± 0.02 ab | 3.48 | 1.71 ± 0.02 ab | 3.40 | 1.81 ± 0.02 ab | 3.59 |
| SH45 | 59.81 ± 1.87 bc | 1.71 ± 0.05 a | 3.48 | 1.71 ± 0.02 ab | 3.48 | 1.90 ± 0.09 bc | 3.86 |
| SH55 | 91.90 ± 2.35 e | 2.23 ± 0.03 f | 4.62 | 1.83 ± 0.03 bc | 3.79 | 1.86 ± 0.07 bc | 3.84 |
| Isolate | Autoaggregation (%) | Hydrophobicity (%) | ||||
|---|---|---|---|---|---|---|
| 3 h | 6 h | 24 h | n-Hexane | Octane | Xylene | |
| G1 | 42.3 ± 0.28 b | 56.7 ± 1.13 b | 69.8 ± 1.57 b | 36.8 ± 3.04 bcde | 42.31 ± 1.85 fg | 6.51 ± 2.21 a |
| O12 | 58.9 ± 0.55 cd | 73.6 ± 0.60 c | 80.7 ± 0.32 c | 32.6 ± 5.71 abcd | 36.7 ± 5.24 cde | 25.16 ± 2.55 bcde |
| O13 | 60.7 ± 0.44 de | 75.8 ± 1.14 c | 83.2 ± 0.75 de | 30.1 ± 1.15 ab | 40.65 ± 0.86 efg | 13.08 ± 7.56 ab |
| O18 | 64.1 ± 0.51 fg | 78.4 ± 0.46 c | 82.8 ± 1.00 d | 31.5 ± 1.95 abc | 43.46 ± 3.02 g | 24.86 ± 4.20 bcde |
| O21 | 65.1 ± 0.21 gh | 77.0 ± 2.41 c | 83.7 ± 0.13 de | 41.9 ± 1.45 de | 35.21 ± 1.07 bcd | 20.73 ± 2.72 abcd |
| O26 | 65.6 ± 0.35 gh | 77.5 ± 0.75 c | 84.4 ± 1.11 def | 37.6 ± 2.76 bcde | 34.46 ± 1.47 abcd | 37.72 ± 3.31 e |
| O36 | 59.2 ± 2.49 cd | 75.0 ± 2.64 c | 84.8 ± 1.01 ef | 42.9 ± 1.11 e | 42.27 ± 2.68 fg | 15.62 ± 2.98 abc |
| O63 | 37.7 ± 0.75 a | 47.0 ± 2.53 a | 51.0 ± 0.28 a | 30.7 ± 2.36 ab | 30.67 ± 1.27 a | 23.71 ± 4.37 bcde |
| O66 | 62.6 ± 0.34 ef | 77.8 ± 0.22 c | 82.9 ± 1.15 de | 24.9 ± 1.12 a | 31.84 ± 3.67 ab | 18.03 ± 1.78 abc |
| SH40 | 42.8 ± 1.38 b | 57.2 ± 0.49 b | 83.7 ± 0.05 de | 41.2 ± 3.61 cde | 44.98 ± 1.57 g | 29.55 ± 8.17 cde |
| SH45 | 66.6 ± 0.31 h | 75.3 ± 4.86 c | 86.1 ± 0.55 f | 33.6 ± 1.84 abcde | 38.51 ± 3.84 def | 21.19 ± 3.46 abcd |
| SH55 | 58.5 ± 0.06 c | 71.3 ± 0.51 c | 80.3 ± 1.43 c | 28.3 ± 1.72 ab | 32.85 ± 1.14 abc | 33.11 ± 9.87 de |
| Isolate | S. Typhimurium | E. coli O157:H7 | S. aureus | L. monocytogenes | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 3 h | 6 h | 24 h | A.M | 3 h | 6 h | 24 h | A.M | 3 h | 6 h | 24 h | A.M | 3 h | 6 h | 24 h | A.M | |
| G1 | 12.2 ± 1.53 b | 23.9 ± 0.46 a | 42.7 ± 1.79 a | +++ | 12.8 ± 0.55 h | 16.5 ± 0.97 a | 38.3 ± 0.6 2 a | +++ | 18.0 ± 0.82 f | 26.8 ± 0.97 f | 48.3 ± 0.98 e | + | 23.8 ± 0.65 a | 33.7 ± 0.12 a | 52.1 ± 0.20 a | +++ |
| O12 | 17.3 ± 0.01 c | 46.7 ± 0.54 cd | 59.7 ± 1.18 cd | +++ | 46.1 ± 1.04 a | 51.3 ± 0.07 f | 64.2 ± 0.08 ij | +++ | 23.0 ± 0.62 d | 48.7 ± 0.33 a | 62.4 ± 1.76 a | +++ | 38.5 ± 0.45 d | 45.9 ± 1.15 b | 61.9 ± 0.83 c | +++ |
| O13 | 25.8 ± 0.61 e | 52.9 ± 1.33 ef | 65.3 ± 0.15 e | + | 38.9 ± 0.26 cd | 46.2 ± 0.09 def | 62.0 ± 0.85 gh | +++ | 26.8 ± 0.78 c | 37.4 ± 0.46 cd | 53.8 ± 0.46 cd | + | 28.9 ± 0.21 b | 40.1 ± 0.59 ab | 57.2 ± 0.14 b | +++ |
| O18 | 35.3 ± 0.93 g | 58.6 ± 1.55 g | 65.4 ± 2.67 e | +++ | 37.2 ± 1.04 d | 47.4 ± 2.45 def | 59.9 ± 0.86 ef | +++ | 21.9 ± 0.08 de | 31.9 ± 1.16 e | 48.3 ± 1.06 e | +++ | 49.9 ± 1.08 f | 60.2 ± 0.95 d | 68.9 ± 2.07 d | +++ |
| O21 | 21.5 ± 0.37 d | 50.1 ± 0.42 de | 62.2 ± 0.59 de | +++ | 43.5 ± 1.05 ab | 51.7 ± 0.83 f | 65.4 ± 0.45 j | +++ | 19.5 ± 0.78 ef | 37.6 ± 0.10 cd | 49.8 ± 1.50 de | +++ | 47.1 ± 0.26 e | 57.6 ± 2.25 cd | 69.6 ± 1.19 d | +++ |
| O26 | 21.9 ± 0.84 d | 50.8 ± 1.08 de | 62.6 ± 1.03 de | +++ | 41.1 ± 0.73 bc | 50.3 ± 0.38 f | 63.1 ± 0.11 hi | +++ | 21.0 ± 0.70 def | 46.8 ± 1.94 ab | 60.1 ± 0.91 ab | + | 45.7 ± 1.09 e | 52.9 ± 0.30 c | 66.8 ± 0.62 d | +++ |
| O36 | 12.0 ± 0.95 b | 43.4 ± 1.25 bc | 57.4 ± 0.06 bcd | +++ | 22.6 ± 0.54 f | 35.0 ± 0.42 c | 52.1 ± 1.21 c | +++ | 14.1 ± 0.12 g | 27.3 ± 0.49 f | 48.5 ± 1.28 e | +++ | 48.0 ± 0.10 ef | 55.2 ± 3.36 cd | 68.5 ± 1.12 d | +++ |
| O63 | 36.0 ± 2.60 gh | 42.6 ± 0.16 bc | 54.2 ± 0.36 b | +++ | 17.2 ± 1.15 g | 26.9 ± 1.09 b | 44.4 ± 0.72 b | +++ | 32.7 ± 0.68 b | 37.6 ± 1.66 cd | 52.7 ± 0.86 cde | +++ | 33.0 ± 0.45 c | 40.0 ± 0.93 ab | 53.4 ± 3.19 ab | +++ |
| O66 | 31.6 ± 0.63 f | 56.0 ± 0.39 fg | 65.0 ± 1.23 e | +++ | 32.4 ± 0.88 e | 42.9 ± 3.03 de | 58.5 ± 1.68 e | +++ | 40.6 ± 1.41 a | 48.0 ± 0.52 a | 62.1 ± 0.82 a | +++ | 52.5 ± 0.30 g | 60.1 ± 2.06 d | 70.0 ± 0.42 d | +++ |
| SH40 | 37.9 ± 0.00 h | 55.8 ± 1.41 fg | 67.3 ± 2.02 e | +++ | 33.5 ± 0.24 e | 41.9 ± 1.92 d | 54.6 ± 1.45 d | +++ | 23.1 ± 0.56 d | 35.3 ± 0.68 de | 33.3 ± 1.00 f | +++ | 55.5 ± 0.71 h | 59.7 ± 2.94 d | 69.3 ± 2.19 d | +++ |
| SH45 | 9.30 ± 0.31 a | 40.2 ± 0.04 b | 55.4 ± 1.49 bc | +++ | 30.4 ± 0.49 e | 44.3 ± 1.41 de | 60.4 ± 0.97 fg | +++ | 34.0 ± 1.06 b | 43.2 ± 0.23 b | 58.6 ± 1.71 ab | +++ | 47.0 ± 0.20 e | 59.2 ± 1.07 cd | 69.8 ± 2.24 d | +++ |
| SH55 | 18.8 ± 0.36 c | 48.1 ± 1.93 d | 58.3 ± 0.04 bcd | +++ | 39.4 ± 0.80 cd | 47.9 ± 0.66 ef | 61.6 ± 0.07 fgh | +++ | 34.6 ± 1.61 b | 39.2 ± 1.07 c | 56.4 ± 0.39 bc | +++ | 29.0 ± 0.97 b | 37.5 ± 0.42 a | 57.5 ± 4.19 b | +++ |
| Isolate | Antibiotic Resistance | Attachment to HT-29 Cells | |||||
|---|---|---|---|---|---|---|---|
| CLI | AMP | SXT | PEN | VAN | ERY | Log10 CFU | |
| G1 | MS | MS | MS | R | MS | S | 6.66 ± 0.06 e |
| O12 | MS | S | MS | MS | R | S | 6.82 ± 0.17 e |
| O13 | MS | MS | MS | R | R | MS | 6.65 ± 0.06 e |
| O18 | S | S | S | S | S | S | 6.27 ± 0.06 bcd |
| O21 | MS | MS | MS | S | MS | MS | 6.00 ± 0.06 a |
| O26 | S | R | R | MS | S | S | 6.23 ± 0.26 bcd |
| O36 | MS | MS | MS | MS | MS | MS | 6.15 ± 0.04 abc |
| O63 | S | MS | MS | MS | S | S | 6.16 ± 0.19 abc |
| O66 | MS | MS | MS | MS | MS | MS | 6.37 ± 0.04 cd |
| SH40 | MS | MS | S | S | MS | MS | 6.36 ± 0.17 cd |
| SH45 | MS | S | S | S | S | MS | 6.41 ± 0.02 d |
| SH55 | MS | S | S | MS | S | S | 6.06 ± 0.03 ab |
| Isolate | EPS Production | Heat Resistance (Log10 CFU/mL) | |
|---|---|---|---|
| Before | After | ||
| G1 | + | 6.6 ± 0.01 a | 4.4 ± 0.02 a |
| O12 | + | 7.5 ± 0.13 efg | 5.2 ± 0.17 c |
| O13 | + | 7.7 ± 0.03 g | 5.3 ± 0.00 cd |
| O18 | + | 7.3 ± 0.05 bcd | 5.6 ± 0.06 f |
| O21 | + | 7.3 ± 0.02 bcd | 5.5 ± 0.02 ef |
| O26 | + | 7.3 ± 0.07 bcd | 5.3 ± 0.07 cd |
| O36 | + | 7.5 ± 0.00 def | 5.4 ± 0.03 cde |
| O63 | + | 7.2 ± 0.06 bcd | 5.3 ± 0.17 cde |
| O66 | + | 7.3 ± 0.04 cde | 4.7 ± 0.10 b |
| SH40 | + | 7.1 ± 0.02 b | 5.4 ± 0.02 def |
| SH45 | + | 7.6 ± 0.07 fg | 5.3 ± 0.13 cd |
| SH55 | + | 7.2 ± 0.03 bc | 4.6 ± 0.24 ab |
| Isolate | Microorganism | Accession No | Source |
|---|---|---|---|
| G1 | Candida sp. | OK441052 | Gamed (traditional fermented dairy product) |
| O12 | Pichia kudriavzevii | OK441055 | Jordanian Olive |
| O13 | Pichia kudriavzevii | OK441056 | Jordanian Olive |
| O18 | Pichia kudriavzevii | OK441057 | Jordanian Olive in oil |
| O21 | Pichia kudriavzevii | OK441060 | Jordanian Olive in oil |
| O26 | Pichia kudriavzevii | OK441064 | Moroccan green olives |
| O36 | Pichia kudriavzevii | OK441067 | Jordanian green olives |
| O63 | Pichia sp. | OK441068 | Jordanian green olives |
| O66 | Saccharomyces cerevisiae | OK441070 | Jordanian green olives |
| SH40 | Pichia kudriavzevii | OK441071 | Shanklish (traditional fermented dairy product) |
| SH45 | Pichia kudriavzevii | OK441072 | Shanklish (traditional fermented dairy product) |
| SH55 | Pichia kudriavzevii | OK441073 | Shanklish (traditional fermented dairy product) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alkalbani, N.S.; Osaili, T.M.; Al-Nabulsi, A.A.; Obaid, R.S.; Olaimat, A.N.; Liu, S.-Q.; Ayyash, M.M. In Vitro Characterization and Identification of Potential Probiotic Yeasts Isolated from Fermented Dairy and Non-Dairy Food Products. J. Fungi 2022, 8, 544. https://doi.org/10.3390/jof8050544
Alkalbani NS, Osaili TM, Al-Nabulsi AA, Obaid RS, Olaimat AN, Liu S-Q, Ayyash MM. In Vitro Characterization and Identification of Potential Probiotic Yeasts Isolated from Fermented Dairy and Non-Dairy Food Products. Journal of Fungi. 2022; 8(5):544. https://doi.org/10.3390/jof8050544
Chicago/Turabian StyleAlkalbani, Nadia S., Tareq M. Osaili, Anas A. Al-Nabulsi, Reyad S. Obaid, Amin N. Olaimat, Shao-Quan Liu, and Mutamed M. Ayyash. 2022. "In Vitro Characterization and Identification of Potential Probiotic Yeasts Isolated from Fermented Dairy and Non-Dairy Food Products" Journal of Fungi 8, no. 5: 544. https://doi.org/10.3390/jof8050544
APA StyleAlkalbani, N. S., Osaili, T. M., Al-Nabulsi, A. A., Obaid, R. S., Olaimat, A. N., Liu, S.-Q., & Ayyash, M. M. (2022). In Vitro Characterization and Identification of Potential Probiotic Yeasts Isolated from Fermented Dairy and Non-Dairy Food Products. Journal of Fungi, 8(5), 544. https://doi.org/10.3390/jof8050544