
The Entomopathogenic Fungus Beauveria bassiana Shows Its Toxic Side within Insects: Expression of Genes Encoding Secondary Metabolites during Pathogenesis


Abstract
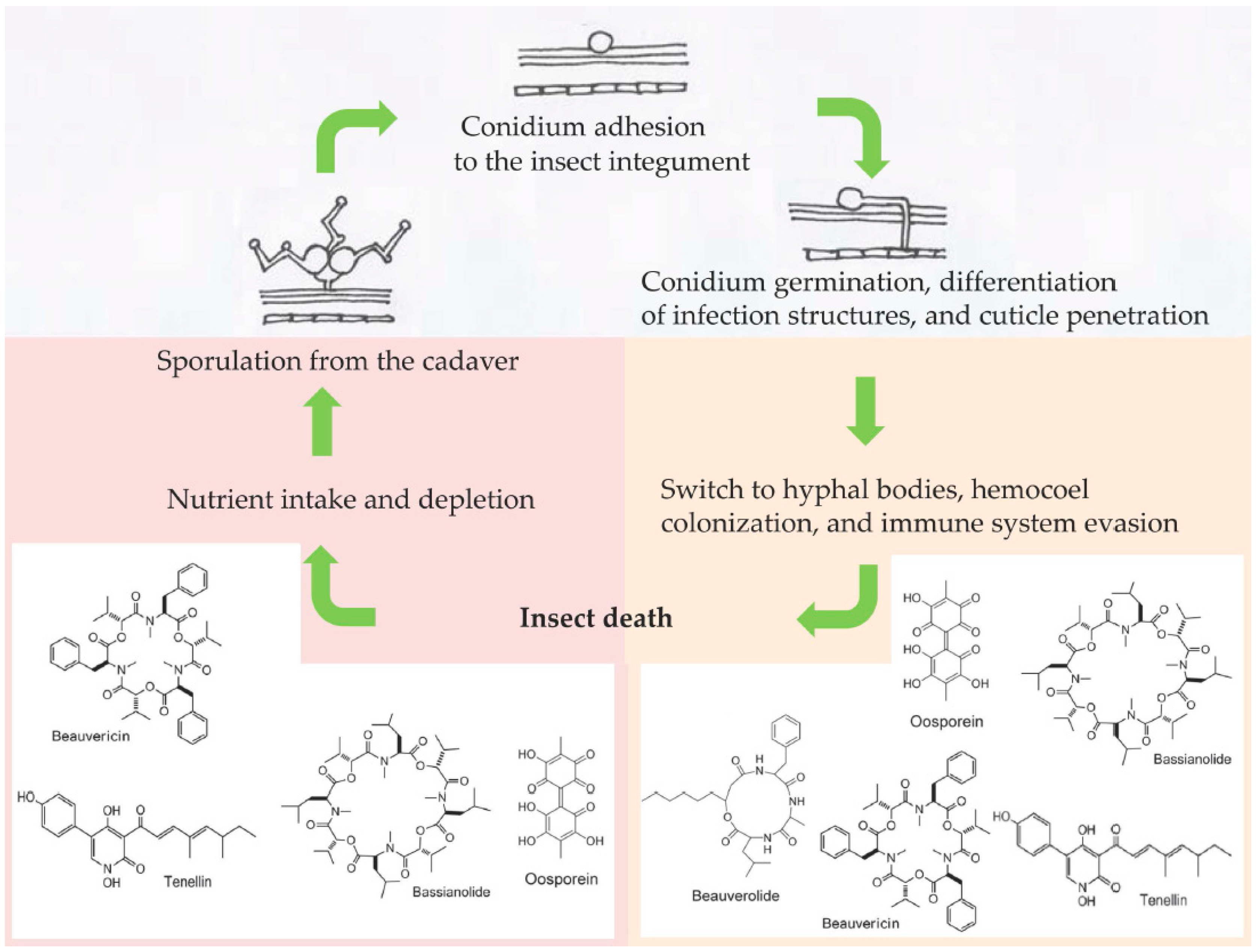
:1. Introduction
2. Role of Secondary Metabolites as Virulence Factors: What We Know and What We Ignore
3. Expression of Nonribosomal Peptide Synthetases (NRPSs) and Polyketide Synthetases (PKSs) during Pathogenesis
4. Dimorphic Transition, Fungal Secondary Metabolites, and the Insect Immune System: A Crosstalk Waiting for Elucidation
5. Immune Response of Insect Hosts
6. Concluding Remarks and Future Directions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Araújo, J.P.; Hughes, D.P. Diversity of entomopathogenic fungi: Which groups conquered the insect body? Adv. Genet. 2016, 94, 1–39. [Google Scholar] [PubMed]
- Pedrini, N. Molecular interactions between entomopathogenic fungi (Hypocreales) and their insect host: Perspectives from stressful cuticle and hemolymph battlefields and the potential of dual RNA sequencing for future studies (Review). Fungal Biol. 2018, 122, 538–545. [Google Scholar] [CrossRef] [PubMed]
- Pedrini, N.; Crespo, R.; Juárez, M.P. Biochemistry of insect epicuticle degradation by entomopathogenic fungi. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2007, 146, 124–137. [Google Scholar] [CrossRef] [PubMed]
- Butt, T.M.; Coates, C.J.; Dubovskiy, I.M.; Ratcliffe, N.A. Entomopathogenic fungi: New insights into host-pathogen interactions. Adv. Genet. 2016, 94, 307e364. [Google Scholar]
- Zhang, L.; Fasoyin, O.E.; Molnár, I.; Xu, Y. Secondary metabolites from hypocrealean entomopathogenic fungi: Novel bioactive compounds. Nat. Prod. Rep. 2020, 37, 1181–1206. [Google Scholar] [CrossRef]
- Altimira, F.; Arias-Aravena, M.; Jian, L.; Real, N.; Correa, P.; González, C.; Godoy, S.; Castro, J.F.; Zamora, O.; Vergara, C.; et al. Genomic and experimental analysis of the insecticidal factors secreted by the entomopathogenic fungus Beauveria pseudobassiana RGM 2184. J. Fungi 2022, 8, 253. [Google Scholar] [CrossRef]
- Lu, H.-L.; St Leger, R.J. Insect Immunity to Entomopathogenic Fungi. Adv. Genet. 2016, 94, 251–285. [Google Scholar] [CrossRef]
- de Bekker, C.; Smith, P.B.; Patterson, A.D.; Hughes, D.P. Metabolomics reveals the heterogeneous secretome of two ento-mopathogenic fungi to ex vivo cultured insect tissues. PLoS ONE 2013, 8, e70609. [Google Scholar] [CrossRef]
- Brewer, D.; Jen, W.-C.; Jones, G.A.; Taylor, A. The antibacterial activity of some naturally occurring 2,5-dihydroxy-l,4-benzoquinones. Can. J. Microbiol. 1984, 30, 1068–1072. [Google Scholar] [CrossRef]
- Mc Namara, L.; Dolan, S.K.; Walsh, J.M.D.; Stephens, J.C.; Glare, T.R.; Kavanagh, K.; Griffin, C.T. Oosporein, an abundant metabolite in Beauveria caledonica, with a feedback induction mechanism and a role in insect virulence. Fungal Biol. 2019, 123, 601–610. [Google Scholar] [CrossRef]
- Gupta, S.; Montllor, C.; Hwang, Y.-S. Isolation of Novel Beauvericin Analogues from the Fungus Beauveria bassiana. J. Nat. Prod. 1995, 58, 733–738. [Google Scholar] [CrossRef]
- Xu, Y.; Zhan, J.; Wijeratne, E.M.K.; Burns, A.M.; Gunatilaka, A.A.L.; Molnár, I. Cytotoxic and Antihaptotactic Beauvericin Analogues from Precursor-Directed Biosynthesis with the Insect Pathogen Beauveria bassiana ATCC 7159. J. Nat. Prod. 2007, 70, 1467–1471. [Google Scholar] [CrossRef] [PubMed]
- Amiri-Besheli, B.; Khambay, B.P.S.; Cameron, S.; Deadman, M.; Butt, T.M. Inter and intra-specific variation in destruxin production by insect pathogenic Metarhizium spp., and its significance to pathogenesis. Mycol. Res. 2000, 104, 447–452. [Google Scholar] [CrossRef]
- Skrobek, A.; Shah, F.A.; Butt, T.M. Destruxin production by the entomogenous fungus Metarhizium anisopliae in insects and factors influencing their degradation. BioControl 2008, 53, 361–373. [Google Scholar] [CrossRef]
- Strasser, H.; Abendstein, D.; Stuppner, H.; Butt, T.M. Monitoring the distribution of secondary metabolites produced by the entomogenous fungus Beauveria brongniartii with particular reference to oosporein. Mycol. Res. 2000, 104, 1227–1233. [Google Scholar] [CrossRef]
- Molnár, I.; Gibson, D.M.; Krasnoff, S.B. Secondary metabolites from entomopathogenic Hypocrealean fungi. Nat. Prod. Rep. 2010, 27, 1241–1275. [Google Scholar] [CrossRef]
- Süssmuth, R.; Muller, J.; von Dohren, H.; Molnar, I. Fungal cyclooligomer depsipeptides: From classical biochemistry to com-binatorial biosynthesis. Nat. Prod. Rep. 2011, 28, 99–124. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Yue, Q.; Wang, C.; Xu, Y.; Molnár, I. Secondary metabolites from hypocrealean entomopathogenic fungi: Genomics as a tool to elucidate the encoded parvome. Nat. Prod. Rep. 2020, 37, 1164–1180. [Google Scholar] [CrossRef]
- Gibson, D.M.; Donzelli, B.G.G.; Krasnoff, S.B.; Keyhani, N.O. Discovering the secondary metabolite potential encoded within entomopathogenic fungi. Nat. Prod. Rep. 2014, 31, 1287–1305. [Google Scholar] [CrossRef]
- Meng, X.; Liao, Z.; Liu, T.; Hussain, K.; Chen, J.; Fang, Q.; Wang, J. Vital roles of Pks11, a highly reducing polyketide synthase, in fungal conidiation, antioxidant activity, conidial cell wall integrity, and UV tolerance of Beauveria bassiana. J. Invertebr. Pathol. 2021, 181, 107588. [Google Scholar] [CrossRef]
- Udompaisarn, S.; Toopaang, W.; Sae-Ueng, U.; Srisuksam, C.; Wichienchote, N.; Wasuwan, R.; Nahar, N.A.; Tanticharoen, M.; Amnuaykanjanasin, A. The polyketide synthase PKS15 has a crucial role in cell wall formation in Beauveria bassiana. Sci. Rep. 2020, 10, 12630. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Orozco, R.; Wijeratne, E.M.K.; Espinosa-Artiles, P.; Gunatilaka, A.A.L.; Stock, S.P.; Molnar, I. Biosynthesis of the cy-clooligomer depsipeptide bassianolide, an insecticidal virulence factor of Beauveria bassiana. Fungal Genet. Biol. 2009, 46, 353–364. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Orozco, R.; Wijeratne, E.M.K.; Gunatilaka, A.A.L.; Stock, S.P.; Molnár, I. Biosynthesis of the Cyclooligomer Depsipeptide Beauvericine, a Virulence Factor of the Entomopathogenic Fungus Beauveria bassiana. Chem. Biol. 2008, 15, 898–907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eley, K.L.; Halo, L.M.; Song, Z.; Powles, H.; Cox, R.J.; Bailey, A.M.; Lazarus, C.M.; Simpson, T.J. Biosynthesis of the 2-Pyridone Tenellin in the Insect Pathogenic FungusBeauveria bassiana. ChemBioChem 2007, 8, 289–297. [Google Scholar] [CrossRef]
- Feng, P.; Shang, Y.; Cen, K.; Wang, C. Fungal biosynthesis of the bibenzoquinone oosporein to evade insect immunity. Proc. Natl. Acad. Sci. USA 2015, 112, 11365–11370. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.; Liu, X.; Keyhani, N.O.; Tang, G.; Pei, Y.; Zhang, W.; Tong, S. Regulatory cascade and biological activity of Beauveria bassiana oosporein that limits bacterial growth after host death. Proc. Natl. Acad. Sci. USA 2017, 114, E1578–E1586. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.-F.; Liu, Y.; Tang, G.-R.; Jin, D.; Chen, X.; Pei, Y.; Fan, Y.-H. The secondary metabolite regulator, BbSmr1, is a central regulator of conidiation via the BrlA-AbaA-WetA pathway in Beauveria bassiana. Environ. Microbiol. 2021, 23, 810–825. [Google Scholar] [CrossRef] [PubMed]
- Rohlfs, M.; Churchill, A.C.L. Fungal secondary metabolites as modulators of interactions with insects and other arthropods. Fungal Genet. Biol. 2011, 48, 23–34. [Google Scholar] [CrossRef]
- Liu, H.; Xie, L.; Wang, J.; Guo, Q.; Yang, S.; Liang, P.; Wang, C.; Lin, M.; Xu, Y.; Zhang, L. The Stress-Responsive and Host-Oriented Role of Nonribosomal Peptide Synthetases in an Entomopathogenic Fungus, Beauveria bassiana. J. Microbiol. Biotechnol. 2017, 27, 439–449. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Lovett, B.; Fang, W. Genetically Engineering Entomopathogenic Fungi. Adv. Genet. 2016, 94, 137–163. [Google Scholar] [CrossRef]
- Lovett, B.; Leger, R.J.S. Genetically engineering better fungal biopesticides. Pest Manag. Sci. 2018, 74, 781–789. [Google Scholar] [CrossRef] [PubMed]
- Xiao, G.; Ying, S.H.; Zheng, P.; Wang, Z.L.; Zhang, S.; Xie, X.Q.; Feng, M.G. Genomic perspectives on the evolution of fungal entomopatho-genicity in Beauveria bassiana. Sci. Rep. 2012, 2, 483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valero-Jiménez, C.A.; Faino, L.; Veld, D.; Smit, S.; Zwaan, B.J.; Van Kan, J.A.L. Comparative genomics of Beauveria bassiana: Uncovering signatures of virulence against mosquitoes. BMC Genom. 2016, 17, 986. [Google Scholar] [CrossRef] [Green Version]
- Chu, Z.-J.; Wang, Y.-J.; Ying, S.-H.; Wang, X.-W.; Feng, M.-G. Genome-Wide Host-Pathogen Interaction Unveiled by Transcriptomic Response of Diamondback Moth to Fungal Infection. PLoS ONE 2016, 11, e0152908. [Google Scholar] [CrossRef]
- Dong, W.-X.; Ding, J.-L.; Gao, Y.; Peng, Y.-J.; Feng, M.-G.; Ying, S.-H. Transcriptomic insights into the alternative splic-ing-mediated adaptation of the entomopathogenic fungus Beauveria bassiana to host niches: Autophagy-related gene 8 as an example. Environ. Microbiol. 2017, 19, 4126–4139. [Google Scholar] [CrossRef] [PubMed]
- Lobo, L.S.; Luz, C.; Fernandes, É.K.; Juárez, M.P.; Pedrini, N. Assessing gene expression during pathogenesis: Use of qRT-PCR to follow toxin production in the entomopathogenic fungus Beauveria bassiana during infection and immune response of the insect host Triatoma infestans. J. Invertebr. Pathol. 2015, 128, 14–21. [Google Scholar] [CrossRef]
- Baldiviezo, L.V.; Pedrini, N.; Santana, M.; Mannino, M.C.; Nieva, L.B.; Gentile, A.; Cardozo, R.M. Isolation of Beauveria bassiana from the Chagas Disease Vector Triatoma infestans in the Gran Chaco Region of Argentina: Assessment of Gene Expression during Host–Pathogen Interaction. J. Fungi 2020, 6, 219. [Google Scholar] [CrossRef]
- Gauthier, G.M. Dimorphism in Fungal Pathogens of Mammals, Plants, and Insects. PLOS Pathog. 2015, 11, e1004608. [Google Scholar] [CrossRef]
- Klein, B.S.; Tebbets, B. Dimorphism and virulence in fungi. Curr. Opin. Microbiol. 2007, 10, 314–319. [Google Scholar] [CrossRef] [Green Version]
- Naruzawa, E.S.; Bernier, L. Control of yeast-mycelium dimorphism in vitro in Dutch elm disease fungi by manipulation of specific external stimuli. Fungal Biol. 2014, 118, 872–884. [Google Scholar] [CrossRef]
- Berrocal, A.; Navarrete, J.; Oviedo, C.; Nickerson, K.W. Quorum sensing activity in Ophiostoma ulmi: Effects of fusel oils and branched chain amino acids on yeast-mycelial dimorphism. J. Appl. Microbiol. 2012, 113, 126–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boucias, D.; Liu, S.; Meagher, R.; Baniszewski, J. Fungal dimorphism in the entomopathogenic fungus Metarhizium rileyi: Detection of an in vivo quorum-sensing system. J. Invertebr. Pathol. 2016, 136, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.-X.; Mouhoumed, A.-Z.; Tong, S.-M.; Ying, S.-H.; Feng, M.-G. BrlA and AbaA Govern Virulence-Required Dimorphic Switch, Conidiation, and Pathogenicity in a Fungal Insect Pathogen. mSystems 2019, 4, e00140-19. [Google Scholar] [CrossRef] [Green Version]
- Mascarin, G.M.; Jackson, M.A.; Kobori, N.N.; Behle, R.W.; Dunlap, C.A.; Júnior, Í. Glucose concentration alters dissolved oxygen levels in liquid cultures of Beauveria bassiana and affects formation and bioefficacy of blastospores. Appl. Microbiol. Biotechnol. 2015, 99, 6653–6665. [Google Scholar] [CrossRef]
- Iwanicki, N.S.A.; Júnior, I.D.; Eilenberg, J.; De Fine Licht, H.H. Comparative RNAseq analysis of the in-sect-pathogenic fungus Metarhizium anisopliae reveals specific transcriptome signatures of filamentous and yeast-like development. G3 2020, 10, 2141–2157. [Google Scholar] [CrossRef] [PubMed]
- Dufour, N.; Rao, R.P. Secondary metabolites and other small molecules as intercellular pathogenic signals. FEMS Microbiol. Lett. 2011, 314, 10–17. [Google Scholar] [CrossRef]
- MacAlpine, J.; Daniel-Ivad, M.; Liu, Z.; Yano, J.; Revie, N.M.; Todd, R.T.; Cowen, L.E. A small molecule produced by Lactobacillus species blocks Candida albicans fila-mentation by inhibiting a DYRK1-family kinase. Nat. Commun. 2021, 12, 6151. [Google Scholar] [CrossRef]
- Lee, S.C.; Li, A.; Calo, S.; Heitman, J. Calcineurin Plays Key Roles in the Dimorphic Transition and Virulence of the Human Pathogenic Zygomycete Mucor circinelloides. PLoS Pathog. 2013, 9, e1003625. [Google Scholar] [CrossRef] [Green Version]
- Cooper, D.; Eleftherianos, I. Memory and Specificity in the Insect Immune System: Current Perspectives and Future Challenges. Front. Immunol. 2017, 8, 539. [Google Scholar] [CrossRef]
- de Oliveira Barbosa Bitencourt, R.; Salcedo-Porras, N.; Umaña-Diaz, C.; Da Costa Angelo, I.; Lowenberger, C. Antifungal immune responses in mosquitoes (Diptera: Culicidae): A review. J. Invertebr. Pathol. 2020, 178, 107505. [Google Scholar] [CrossRef]
- Strand, M.R. The insect cellular immune response. Insect Sci. 2008, 15, 1–14. [Google Scholar] [CrossRef]
- Lavine, M.D.; Strand, M.R. Insect hemocytes and their role in immunity. Insect Biochem. Mol. Biol. 2002, 32, 1295–1309. [Google Scholar] [CrossRef]
- Bulet, P.; Hetru, C.; Dimarcq, J.-L.; Hoffmann, D. Antimicrobial peptides in insects; structure and function. Dev. Comp. Immunol. 1999, 23, 329–344. [Google Scholar] [CrossRef]
- Hultmark, D. Drosophila immunity: Paths and patterns. Curr. Opin. Immunol. 2003, 15, 12–19. [Google Scholar] [CrossRef]
- Pal, S.; Wu, L.P. Lessons from the Fly: Pattern Recognition in Drosophila melanogaster. In Target Pattern Recognition in Innate Immunity; Kishore, U., Ed.; Springer: New York, NY, USA, 2019; pp. 162–174. [Google Scholar]
- Mannino, M.C.; Paixão, F.R.; Pedrini, N. The limpet transcription factors of Triatoma infestans regulate the response to fungal infection and modulate the expression pattern of defensin genes. Insect Biochem. Mol. Biol. 2019, 108, 53–60. [Google Scholar] [CrossRef]
- Brown, S.E.; Howard, A.; Kasprzak, A.B.; Gordon, K.H.; East, P.D. The discovery and analysis of a diverged family of novel antifungal moricin-like peptides in the wax moth Galleria mellonella. Insect Biochem. Mol. Biol. 2008, 38, 201–212. [Google Scholar] [CrossRef]
- Schuhmann, B.; Seitz, V.; Vilcinskas, A.; Podsiadlowski, L. Cloning and expression of gallerimycin, an antifungal peptide expressed in immune response of greater wax moth larvae, Galleria mellonella. Arch. Insect Biochem. Physiol. 2003, 53, 125–133. [Google Scholar] [CrossRef]
- Steiner, H.; Hultmark, D.; Engström, Å.; Bennich, H.; Boman, H.G. Sequence and specificity of two antibacterial proteins involved in insect immunity. Nature 1981, 292, 246–248. [Google Scholar] [CrossRef]
- Lopez-Lastra, C.; Boucias, D.G. Studies on the cellular reactions of Spodoptera exigua larvae infected with the fungus Nomuraea rileyi. J. Invertebr. Pathol. 1994, 63, 101–102. [Google Scholar] [CrossRef]
- Steinchen, W.; Lackner, G.; Yasmin, S.; Schrettl, M.; Dahse, H.-M.; Haas, H.; Hoffmeister, D. Bimodular Peptide Synthetase SidE Produces Fumarylalanine in the Human Pathogen Aspergillus fumigatus. Appl. Environ. Microbiol. 2013, 79, 6670–6676. [Google Scholar] [CrossRef] [Green Version]
- Ding, J.-L.; Hou, J.; Feng, M.-G.; Ying, S.-H. Transcriptomic analyses reveal comprehensive responses of insect hemocytes to mycopathogen Beauveria bassiana, and fungal virulence-related cell wall protein assists pathogen to evade host cellular defense. Virulence 2020, 11, 1352–1365. [Google Scholar] [CrossRef] [PubMed]
- Long, Y.; Gao, T.; Liu, S.; Zhang, Y.; Li, X.; Zhou, L.; Su, Q.; Xu, L.; Yang, Y. Analysis of the humoral immunal response tran-scriptome of Ectropis obliqua infected by Beauveria bassiana. Insects 2022, 13, 225. [Google Scholar] [CrossRef] [PubMed]
- Xiong, G.-H.; Xing, L.-S.; Lin, Z.; Saha, T.T.; Wang, C.; Jiang, H.; Zou, Z. High throughput profiling of the cotton bollworm Helicoverpa armigera immunotranscriptome during the fungal and bacterial infections. BMC Genom. 2015, 16, 321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, D.; Liu, Y.; Zhou, F.; Wang, G.; An, C. Identification of immunity-related genes in Ostrinia furnacalis against ento-mopathogenic fungi by RNA-seq analysis. PLoS ONE 2014, 9, e86436. [Google Scholar]
- Lai, Y.; Chen, H.; Wei, G.; Wang, G.; Li, F.; Wang, S. In vivo gene expression profiling of the entomopathogenic fungus Beauveria bassiana elucidates its infection stratagems in Anopheles mosquito. Sci. China Life Sci. 2017, 60, 839–851. [Google Scholar] [CrossRef]


{kind=link}
| Gene Detected (Protein encoded, Family’s Acronym 1) | Insect Host | Technique Used [Reference] |
|---|---|---|
| BbbeaS (beauvericin synthetase, NRPS) | Triatoma infestans (Hemiptera: Reduviidae) | qPCR [36,37] |
| BbbeaS (beauvericin synthetase, NRPS) | Plutella xylostella (Lepidoptera: Plutellidae) | qPCR [29] |
| BbbslS (bassianolide synthetase, NRPS) | Triatoma infestans (Hemiptera: Reduviidae) | qPCR [36,37] |
| BbbslS (bassianolide synthetase, NRPS) | Plutella xylostella (Lepidoptera: Plutellidae) | qPCR [29] |
| BbtenS (tenellin synthetase, NRPS) | Triatoma infestans (Hemiptera: Reduviidae) | qPCR [36,37] |
| BbtenS (tenellin synthetase, NRPS) | Plutella xylostella (Lepidoptera: Plutellidae) | qPCR [29] |
| BbopS1 (oosporein synthetase, PKS) | Galleria mellonella (Lepidoptera: Pyralidae) | qPCR [26] |
| BBA_08222 (putative beauverolide synthetase, NRPS) | Plutella xylostella (Lepidoptera: Plutellidae) | qPCR [29] |
| BBA_07611 (not detailed, NRPS) | Plutella xylostella (Lepidoptera: Plutellidae) | qPCR [29] |
| BBA_07611 (not detailed, NRPS) | Galleria mellonella (Lepidoptera: Pyralidae) | RNA-seq [35] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pedrini, N. The Entomopathogenic Fungus Beauveria bassiana Shows Its Toxic Side within Insects: Expression of Genes Encoding Secondary Metabolites during Pathogenesis. J. Fungi 2022, 8, 488. https://doi.org/10.3390/jof8050488
Pedrini N. The Entomopathogenic Fungus Beauveria bassiana Shows Its Toxic Side within Insects: Expression of Genes Encoding Secondary Metabolites during Pathogenesis. Journal of Fungi. 2022; 8(5):488. https://doi.org/10.3390/jof8050488
Chicago/Turabian StylePedrini, Nicolás. 2022. "The Entomopathogenic Fungus Beauveria bassiana Shows Its Toxic Side within Insects: Expression of Genes Encoding Secondary Metabolites during Pathogenesis" Journal of Fungi 8, no. 5: 488. https://doi.org/10.3390/jof8050488
APA StylePedrini, N. (2022). The Entomopathogenic Fungus Beauveria bassiana Shows Its Toxic Side within Insects: Expression of Genes Encoding Secondary Metabolites during Pathogenesis. Journal of Fungi, 8(5), 488. https://doi.org/10.3390/jof8050488