
Squalene Monooxygenase Gene SsCI80130 Regulates Sporisorium scitamineum Mating/Filamentation and Pathogenicity

Abstract
:1. Introduction
2. Materials and Methods
2.1. Characterization of the SsCI80130 Gene Sequence
2.2. Strains and Growth Conditions
2.3. Nucleic Acid Manipulation
2.4. Construction of the SsCI80130 Gene Knockout Mutants and Complementary Mutants
2.5. Squalene Monooxygenase (SQE) Activity Assay
2.6. Determination of Squalene Content and Ergosterol Content
2.7. Analysis of Gene Expression
2.8. Assay of the Pathogenicity of the SsCI80130 Gene Knockout Mutants and Complementary Mutants
2.9. Microscopy
2.10. Statistic Analysis
3. Results
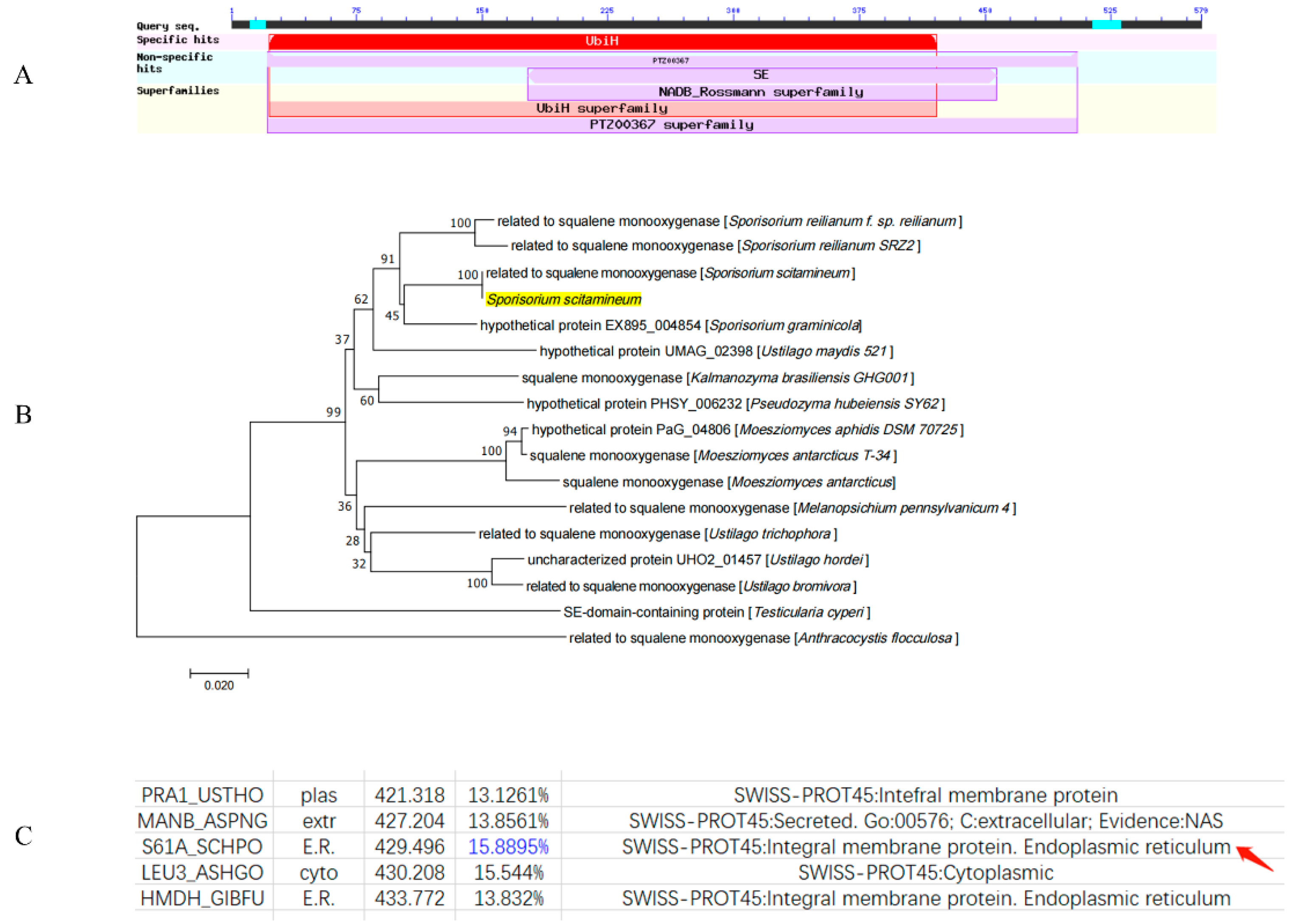
3.1. Identification and Characterization of the SsCI80130 Gene
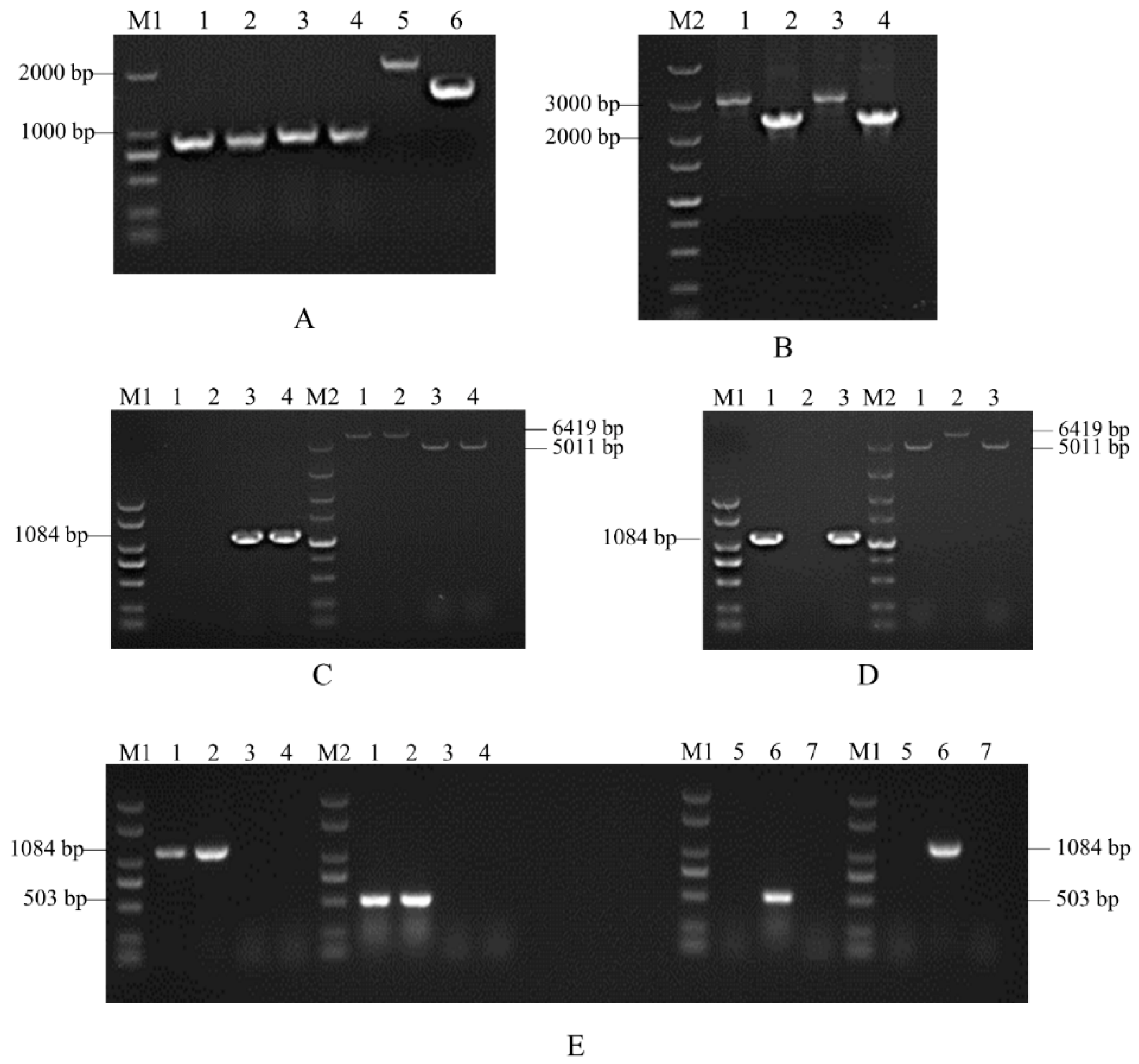
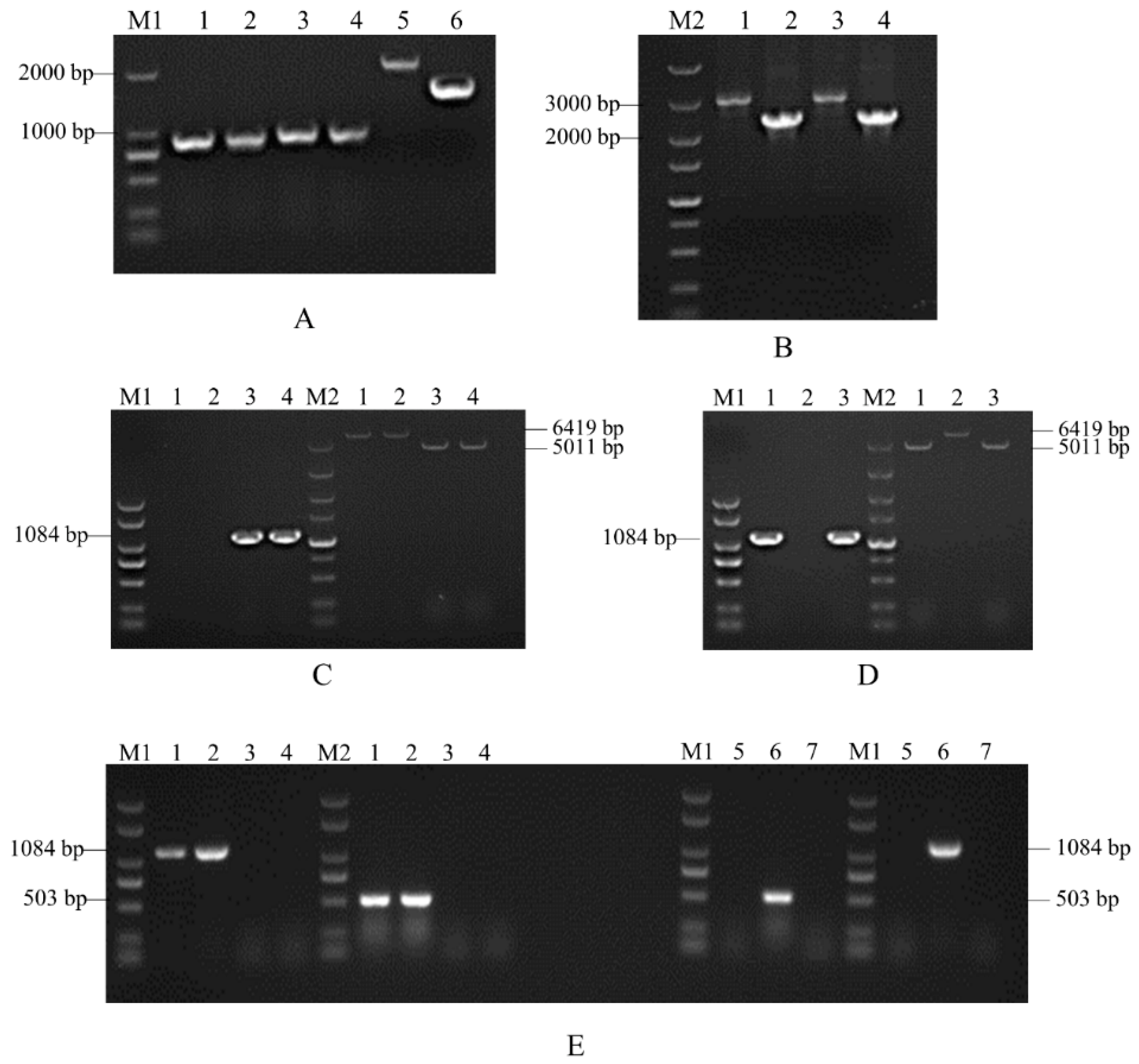
3.2. Molecular Construction of Knockout Mutants and Complementary Mutants
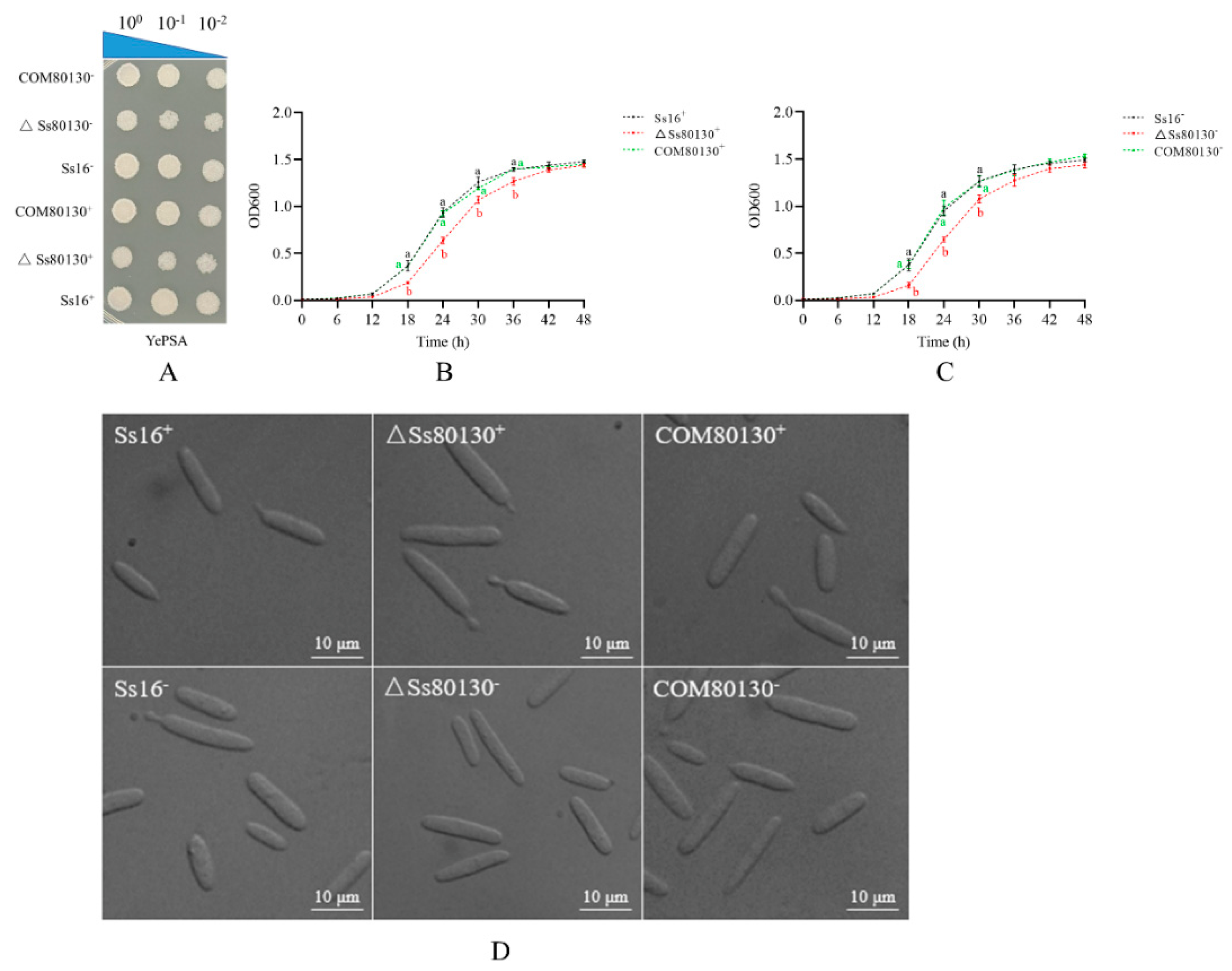
3.3. Effects of the SsCI80130 Gene on Morphology and Growth of S. scitamineum
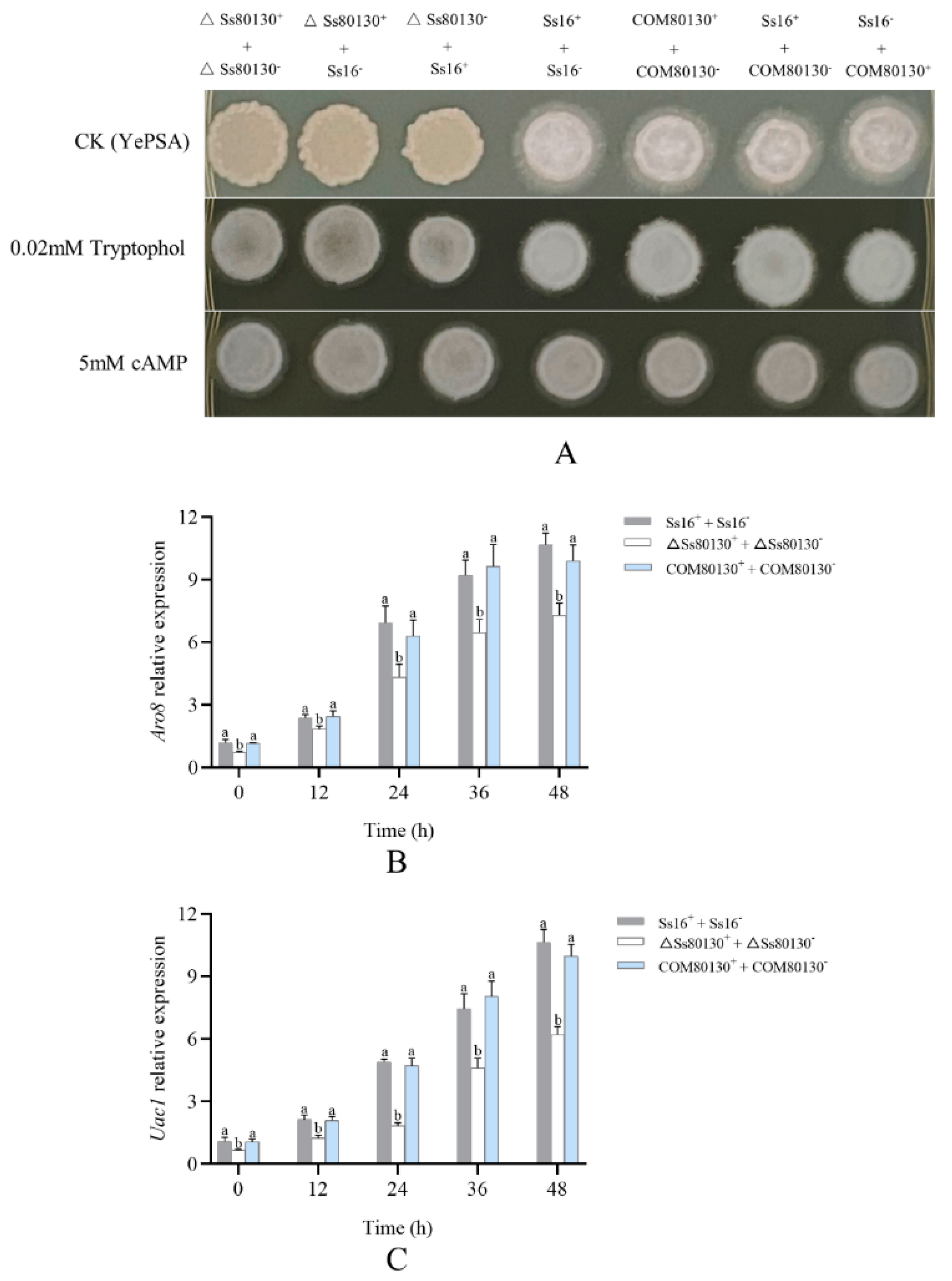
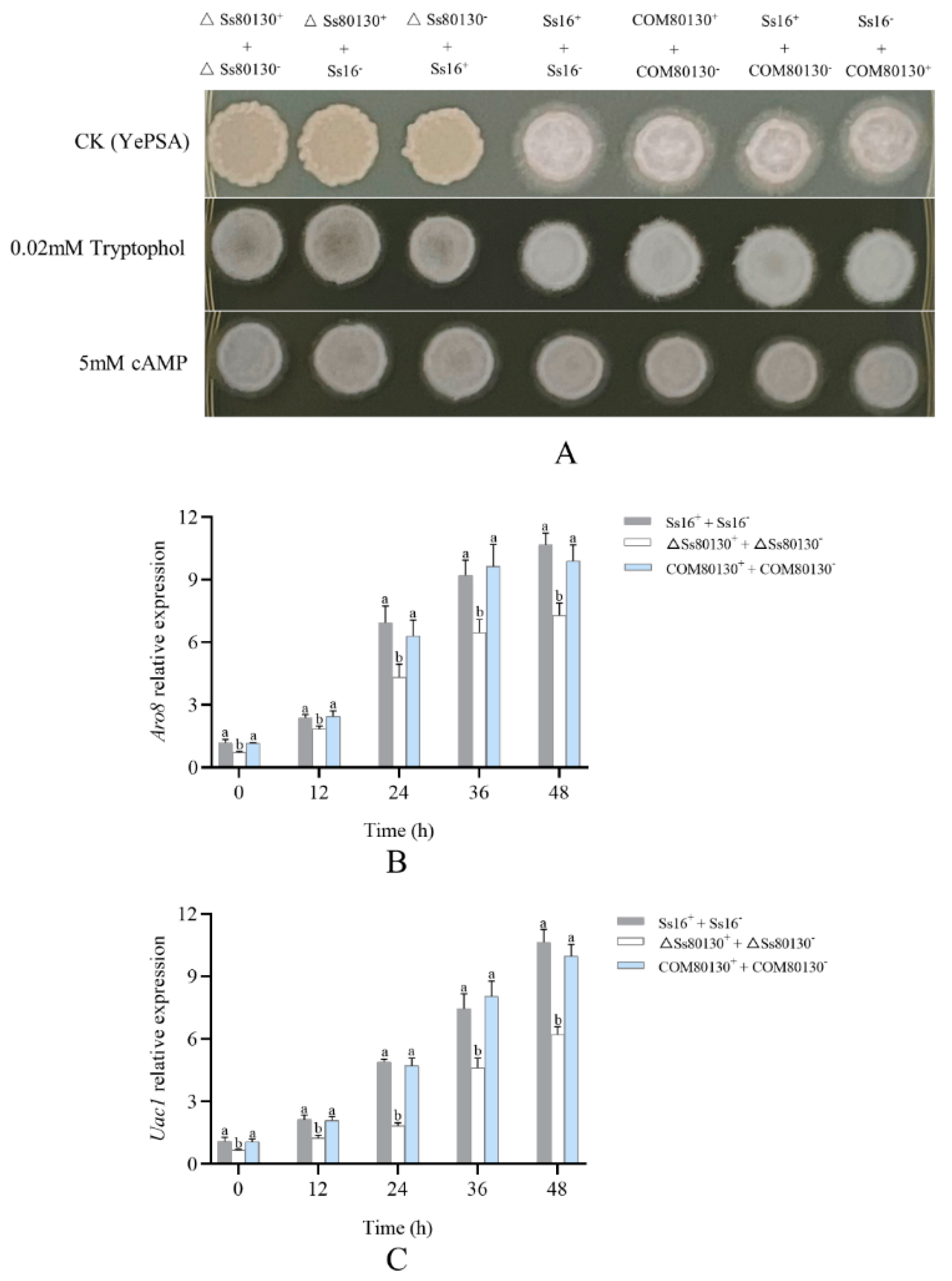
3.4. Effects of the SsCI80130 Gene on Sexual Mating of S. scitamineum
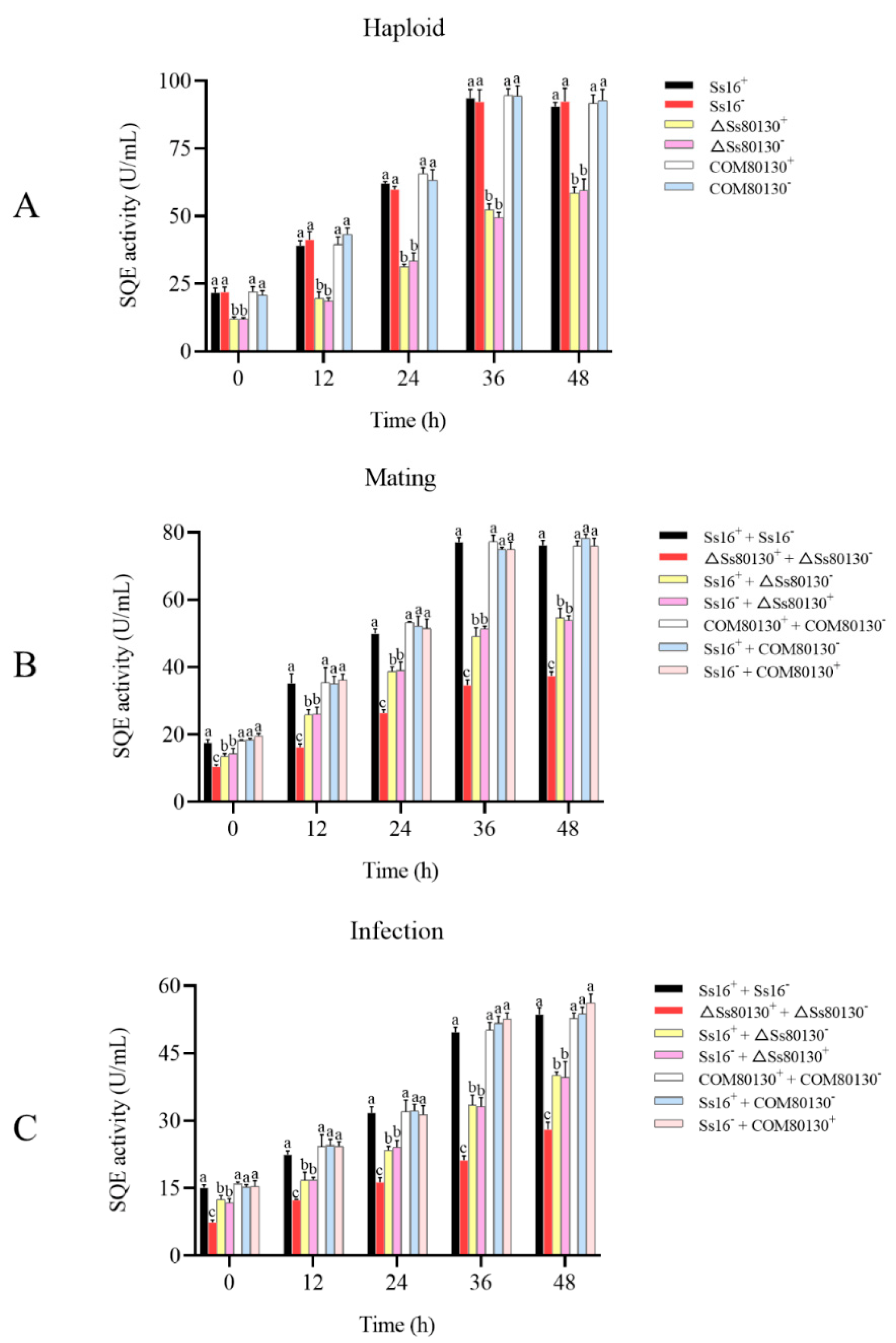
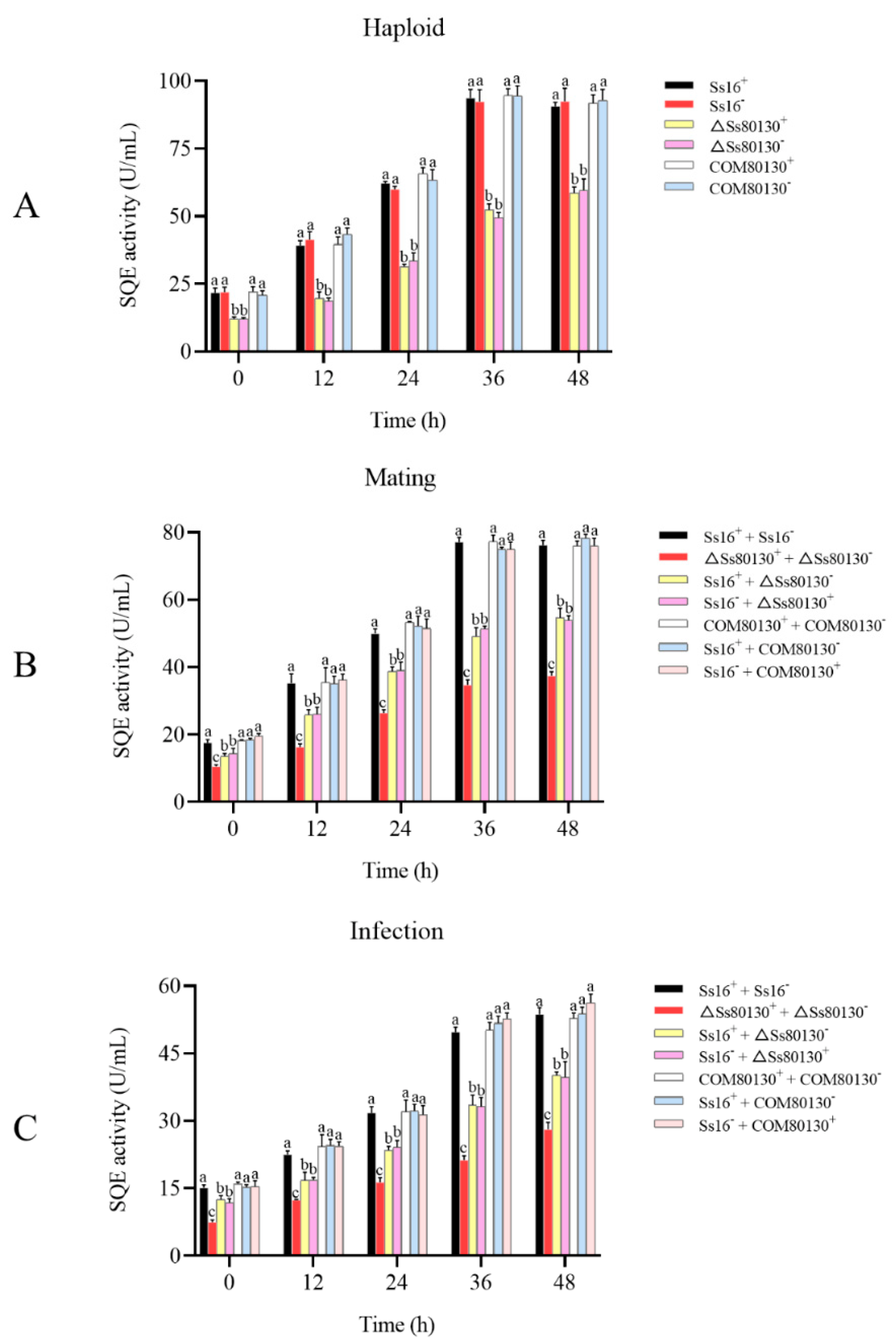
3.5. Effects of the SsCI80130 Gene on the SQE Activity of S. scitamineum
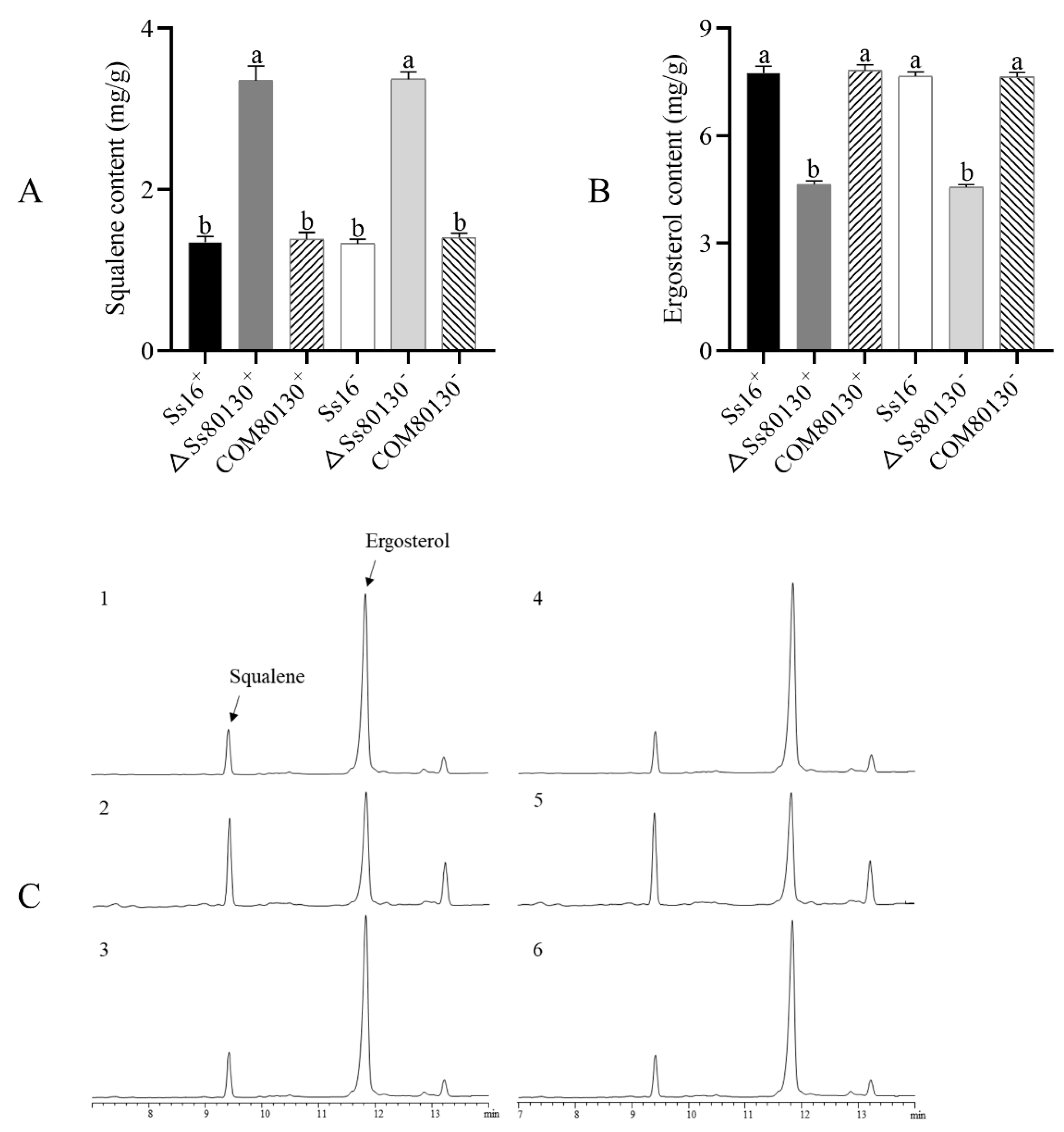
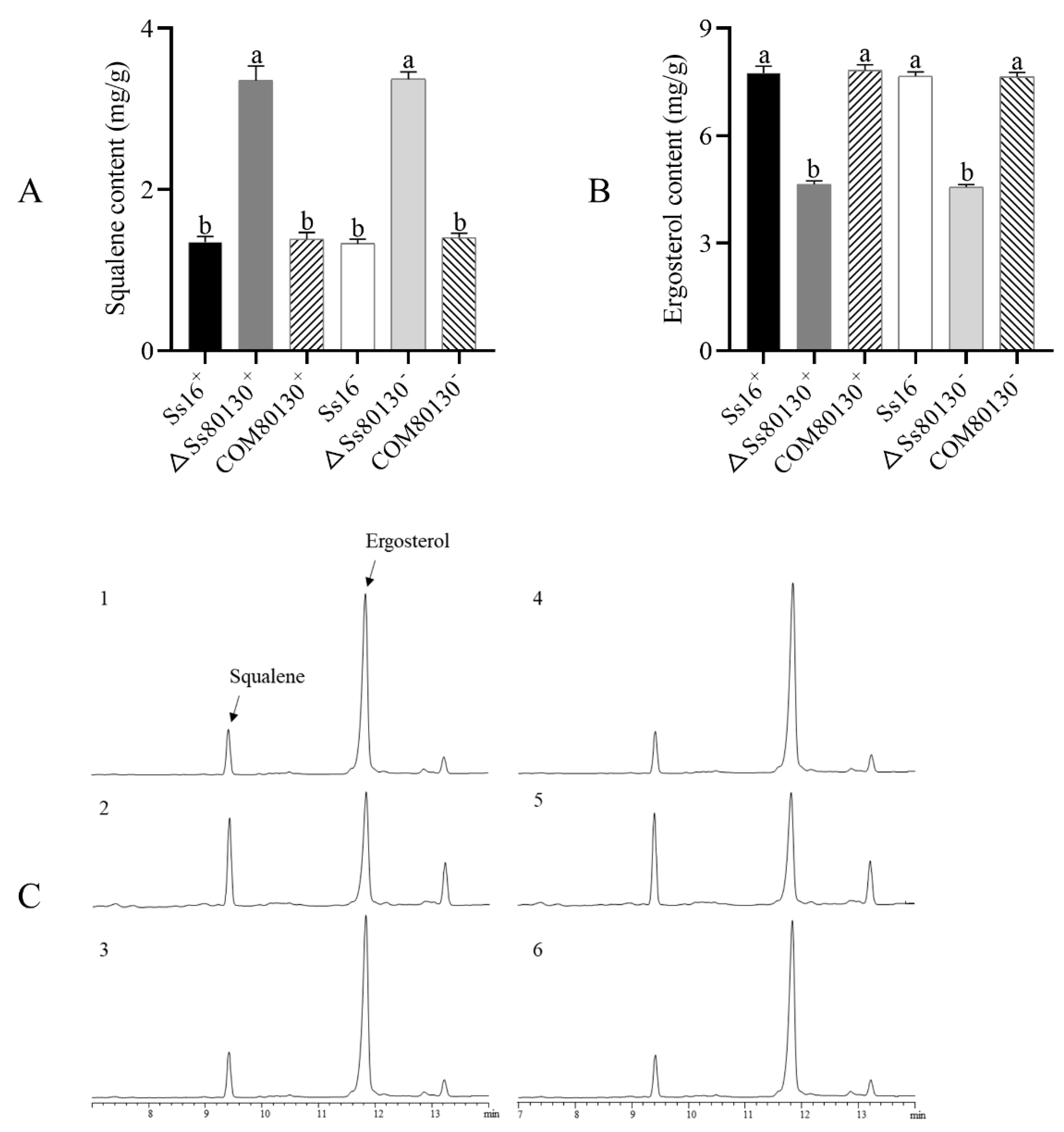
3.6. SsCI80130 Gene Is Involved in Ergosterol Biosynthesis of S. scitamineum
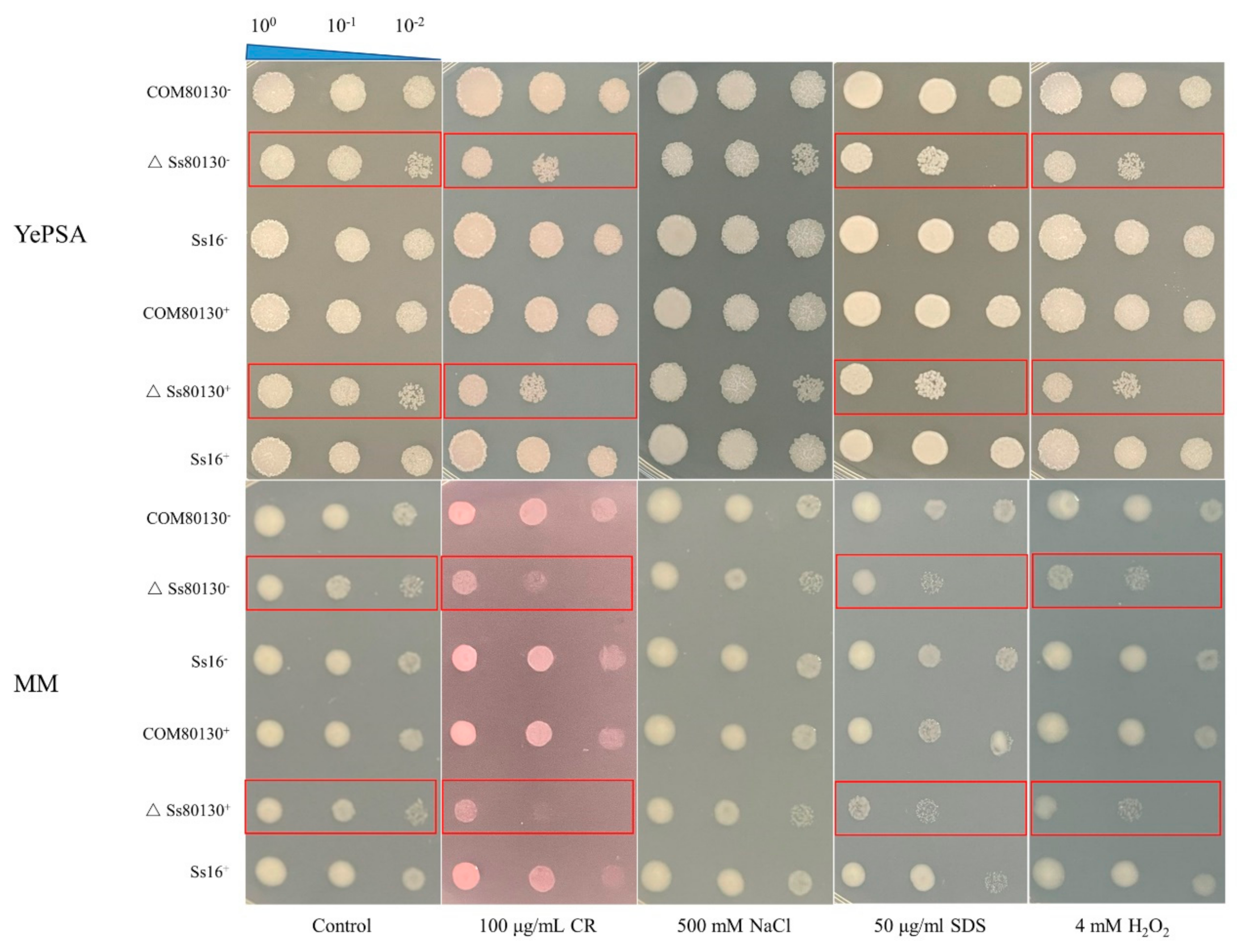
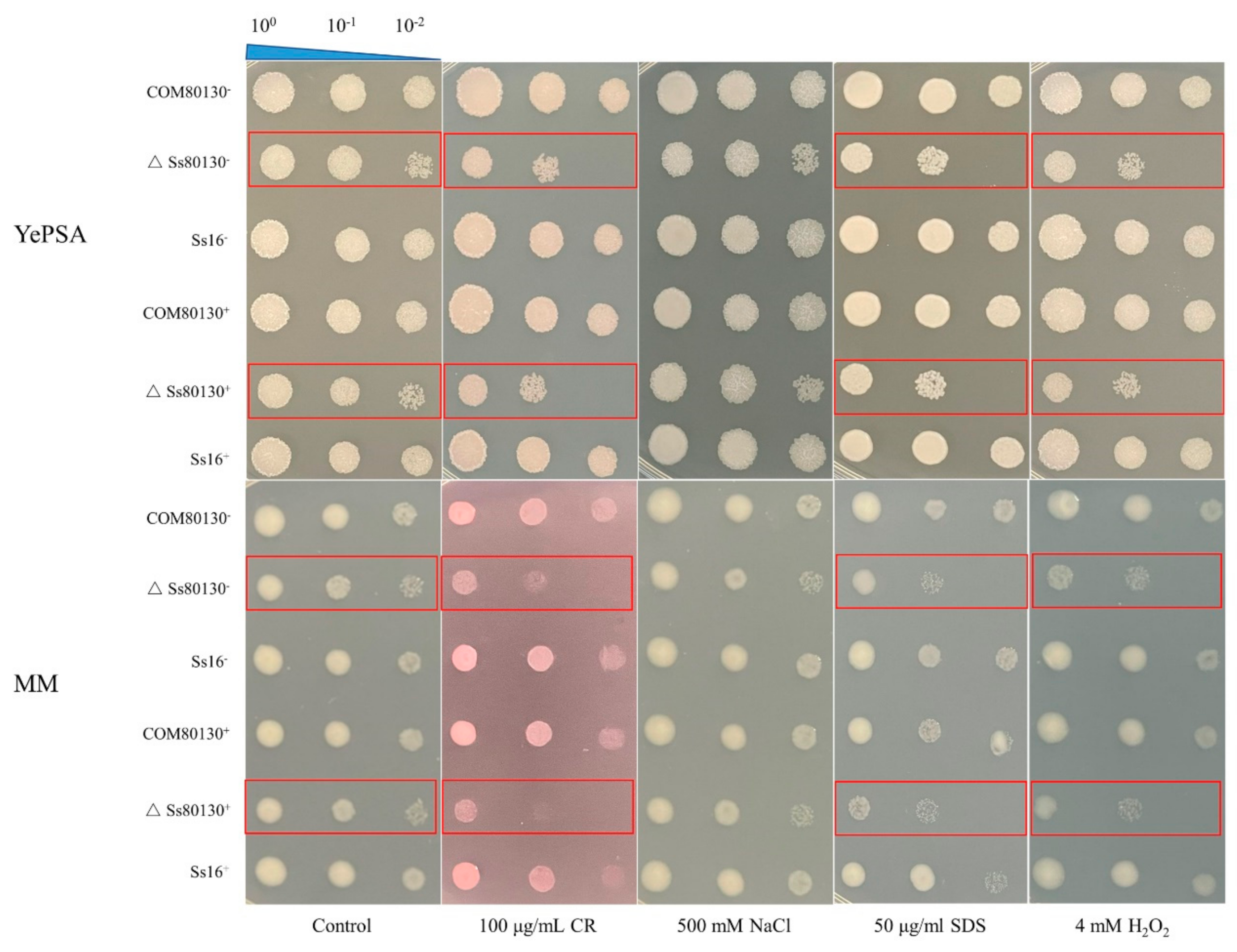
3.7. Effects of the SsCI80130 Gene on the Stress Tolerance of S. scitamineum
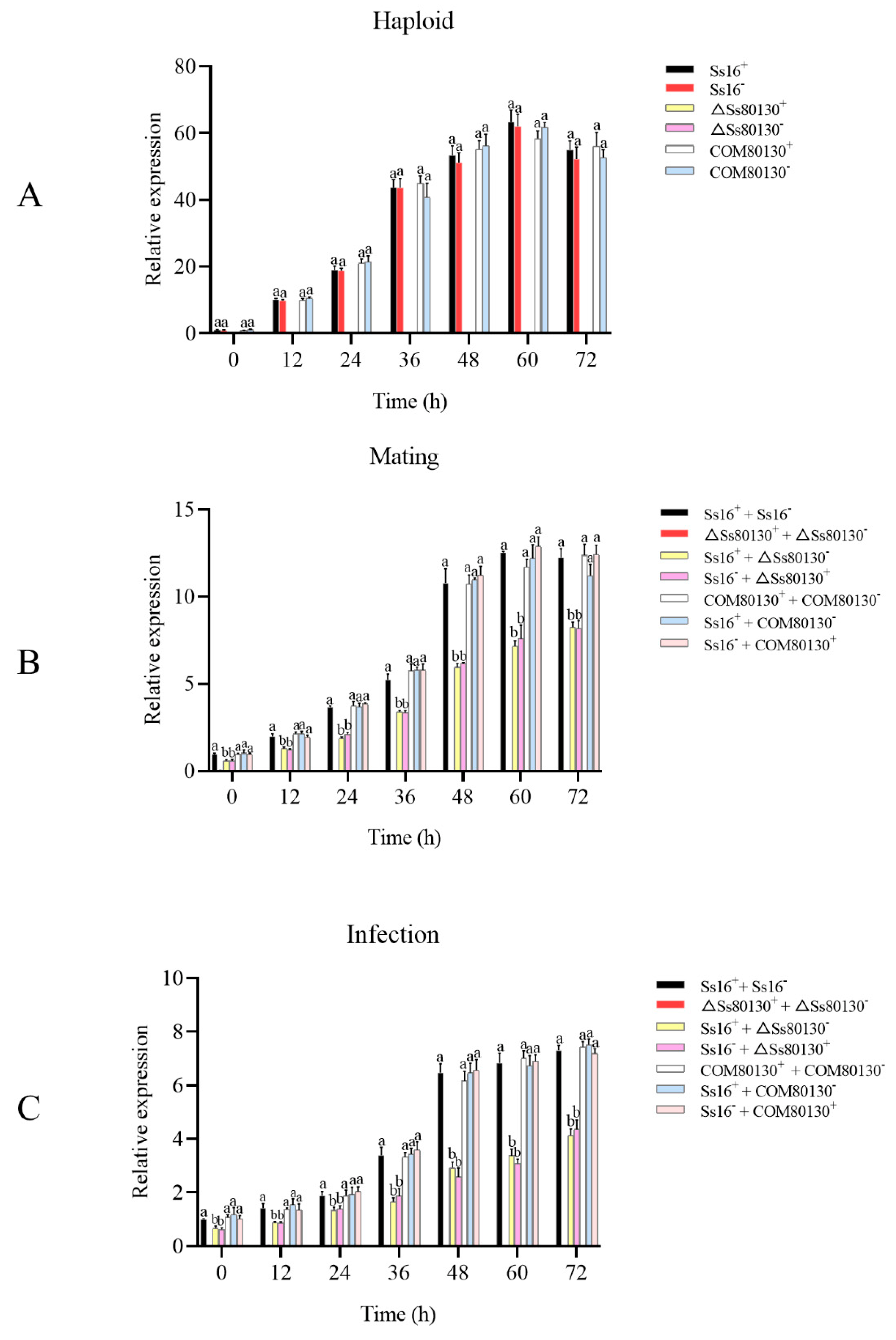
3.8. SsCI80130 Gene Expression Level
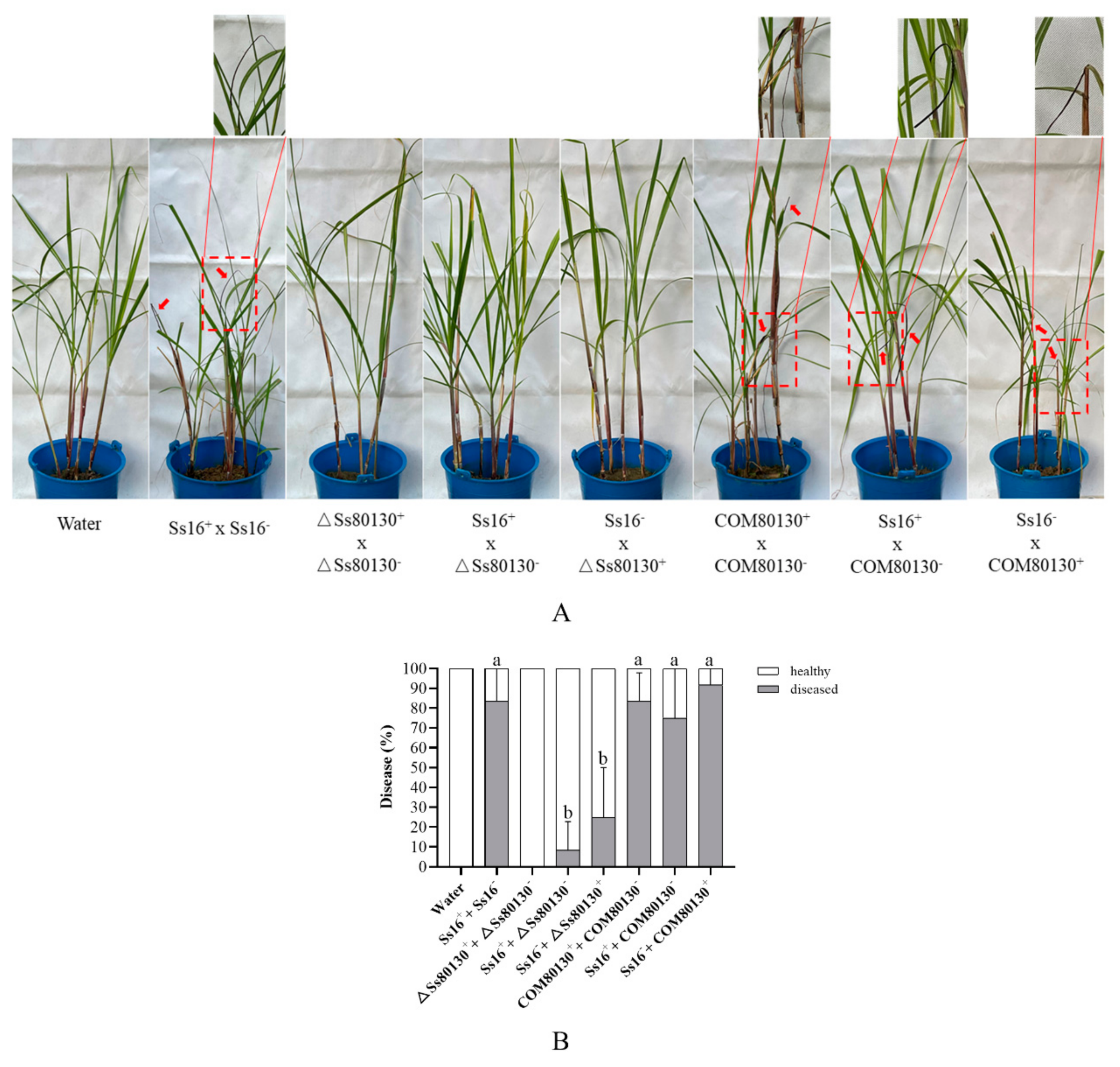
3.9. SsCI80130 Gene Is Required for the Pathogenicity of S. scitamineum
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lam, E.; Shine, J.R.; Silva, J.D.; Lawton, M.; Bonos, S.; Calvino, M.; Carrer, H.; Silver, M.; Glynn, N.; Heisel, Z.; et al. Improving sugarcane for biofuel: Engineering for an even better feedstock. Gcb. Bioenergy 2009, 1, 251–255. [Google Scholar] [CrossRef]
- Liu, Y.Q.; Li, Y.P.; Liang, W.H.; Song, Q.D.; Qin, X.L.; Ye, L. Development status of foreign sugarcane industry. World Agric. 2015, 8, 147–152. [Google Scholar]
- Wang, C.M.; Li, J.; Zhang, R.Y.; Wang, X.Y.; Shan, H.L.; Cang, X.Y.; Yin, J.; Luo, Z.M.; Huang, Y.K. Research progress of sugarcane smut disease. Sugar Crops China 2021, 43, 65–70. [Google Scholar]
- Shen, W.K.; Jiang, Z.D.; Deng, H.H.; Liu, R. Research progress on sugarcane smut disease and Sporisorium scitaminea. Chin. J. Trop Crops 2013, 34, 2063–2068. [Google Scholar]
- Tan, Z.S.; Li, D.Y. Occurrence status and control countermeasures of sugarcane smut. Chin. Hor. Abs. 2015, 31, 225–226. [Google Scholar]
- Wei, J.J.; Song, X.P.; Wei, C.Y.; Zhang, X.Q.; Huang, W.H.; Yan, M.X. Research progress on sugarcane smut and its control. Guangdong Agric. Sci. 2019, 6, 81–88. [Google Scholar]
- Dutheil, J.Y.; Mannhaupt, G.; Schweizer, G.; Sieber, C.M.; Martin, M.; Ulrich, G.; Jan, S.; Regine, K. A tale of genome compartmentalization: The evolution of virulence clusters in smut fungi. Genome Biol. Evol. 2016, 8, 681–704. [Google Scholar] [CrossRef] [Green Version]
- Taniguti, L.M.; Schaker, P.D.; Benevenuto, J.; Peters, L.P.; Carvalho, G.; Palhares, A.; Quecine, M.C.; Nunes, F.R.; Kmit, M.C.; Wai, A.; et al. Complete genome sequence of Sporisorium scitamineum and biotrophic interaction transcriptome with sugarcane. PLoS ONE 2015, 10, e0129318. [Google Scholar] [CrossRef]
- Albert, H.H.; Schenck, S. PCR amplification from a homolog86 of the bE mating-type gene as a sensitive assay for the presence of U. scitaminea DNA. Plant Dis. 1996, 80, 1189–1192. [Google Scholar] [CrossRef]
- Barnabas, L.; Ashwin, N.; Kaverinathan, K.; Trentin, A.R.; Pivato, M.; Sundar, A.R.; Malathi, R.; Viswanathan, O.B.; Rosana, K.; Neethukrishna, P.; et al. Proteomic analysis of a compatible interaction between sugarcane and Sporisorium scitamineum. Proteomics 2016, 16, 1111–1122. [Google Scholar] [CrossRef]
- Yan, M.; Zhu, G.; Lin, S.; Xian, X.; Zhang, L.H. The mating-type locus b of the sugarcane smut Sporisorium scitamineum is essential for mating, filamentous growth and pathogenicity. Fungal Genet. Biol. 2016, 86, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, S.Q.; Deng, Y.Z.; Cai, E.P.; Yan, M.X.; Li, Y.L.; Chen, B.S.; Chang, C.Q.; Jing, Z.D. The Farnesyltransferase β-Subunit Ram1 regulates Sporisorium scitamineum mating, pathogenicity and cell wall integrity. Front. Microbiol. 2019, 10, 976. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.Z.; Zhang, B.; Chang, C.Q.; Wang, Y.X.; Lu, S.; Sun, S.Q.; Zhang, X.M.; Chen, B.S.; Jiang, Z.D. The MAP kinase SsKpp2 is required for mating/fifilamentation in Sporisorium scitamineum. Front. Microbiol. 2018, 9, 2555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.X.; Deng, Y.Z.; Cui, G.B.; Huang, C.W.; Zhang, B.; Chang, C.Q.; Jiang, Z.D.; Zhang, L.H. The AGC Kinase SsAgc1 regulates Sporisorium scitamineum mating/fifilamentation and pathogenicity. mSphere 2019, 4, e00259-19. [Google Scholar] [CrossRef] [Green Version]
- Zhu, G.N.; Deng, Y.Z.; Cai, E.P.; Yan, M.X.; Cui, G.B.; Wang, Z.Q.; Zou, C.W.; Zhang, B.; Xi, P.G.; Chang, C.Q.; et al. Identification and functional analysis of the pheromone response factor gene of Sporisorium scitamineum. Front. Microbiol. 2019, 10, 2115. [Google Scholar] [CrossRef]
- Li, H.Z.; Cai, Y.C.; Deng, Q.Q.; Bao, H.; Chen, J.W.; Shen, W.K. Cytochrome P450 Sterol 14 Alpha-Demethylase Gene SsCI72380 Is Required for Mating/Filamentation and Pathogenicity in Sporisorium scitamineum. Front. Microbiol. 2021, 12, 696117. [Google Scholar] [CrossRef]
- Van, B.W.; Kamerbeek, N.M.; Fraaije, M.W. Flavoprotein monooxygenases, a diverse class of oxidative biocatalysts. J. Biotechnol. 2006, 124, 670–689. [Google Scholar]
- Joosten, V.; Van, B.W. Flavoenzymes. Cur. Opin. Chem. Biol. 2007, 11, 195. [Google Scholar] [CrossRef]
- Rasbery, J.M.; Shan, H.; LeClair, R.J.; Michael, N.; Seiichi, P.T.; Matsuda, B.B. Arabidopsis thaliana squalene epoxidase 1 is essential for root and seed development. J. Biol. Chem. 2007, 282, 17002–17013. [Google Scholar] [CrossRef] [Green Version]
- He, F.M.; Zhu, Y.P.; He, M.X.; Zhang, Y.Z. Molecular cloning and characterization of the gene encoding squalene epoxidase in Panax notogineseng. DNA Seq. 2009, 19, 270–273. [Google Scholar] [CrossRef]
- Gao, L.; Zhang, C.; Lu, L. Advances on genetic regulation nechanism of ergosterol synthesis pathway in opportunistic pathogen Aspergillus fumigatus. J. Fungal Res. 2019, 17, 60–66. [Google Scholar]
- Matyash, V.; Entchev, E.V.; Mende, F.; Michaela, W.B.; Christoph, T.; Arndt, W.S.; Knolker, H.J.; Samuel, W.; Teymuras, V.K. Sterol-derived hormone(s) controls entry into diapause in Caenorhabditis elegans by consecutive activation of DAF-12 and DAF-16. PLoS Biol. 2004, 2, e280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, J.X.; Zhai, P.F.; Zhang, Y.W.; Zhang, C.Y.; Sang, H.; Han, G.Z.; Keller, N.P.; Lu, L. The Aspergillus fumigatus damage resistance protein family coordinately regulates ergosterol biosynthesis and azole susceptibility. mBio 2016, 7, e01919-15. [Google Scholar] [CrossRef] [Green Version]
- Campagnac, E.; Fontaine, J.A.; Lounès, H.S.; Laruelle, F.; Durand, R.; Grandmougin, F. Fenpropimorph slows down the sterol pathway and the development of the arbuscular mycorrhizal fungus Glomus intraradices. Mycorrhiza 2009, 19, 365–374. [Google Scholar] [CrossRef]
- Ryder, N.S. Terbinafine: Mode of action and properties of the squalene epoxidase inhibition. Brit. J. Dermatol. 1992, 39, 2–7. [Google Scholar] [CrossRef] [PubMed]
- Ritu, P.; Shankarling, K.; Tulika, P.; Ernst, J.F.; Prasad, R. Squalene epoxidase encoded by erg1 affects morphogenesis and drug susceptibilities of Candida albicans. J. Antimicrob. Chemoth. 2005, 55, 905–913. [Google Scholar]
- Wu, J.; Li, H.Z.; Deng, Q.Q.; Chen, J.W.; Shen, W.K. Transcriptomic analysis of Sporisorium scitamineum isolates with difffferent pathogenicity. J. Huazhong Agric. Univ. 2020, 39, 40–44. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Deng, Q.Q.; Xu, G.H.; Dou, Z.M.; Shen, W.K. Identification of three Sporisorium scitamineum pathogenic races in mainland China. Int. J. Agric. Biol. 2018, 20, 799–802. [Google Scholar]
- Shen, W.K.; Zhou, G.H.; Deng, H.H.; Zhou, L.Y. Detection of sugarcane ratoon stunting disease pathogen with polymerase chain reaction (PCR) and nucleotide sequence analysis. Chin. Agric. Sci. Bull. 2006, 22, 413. [Google Scholar]
- Cai, E.P.; Mei, D.; Zhang, X.M.; Sun, X.; Li, L.Y.; Wu, R.R.; Deng, Y.Z.; Jiang, Z.D.; Chang, C.Q. A gene knockout method based on protoplast transformation with two PCR fragments in Sporisorium scitamineum. Mycosystema 2020, 39, 14. [Google Scholar]
- Chang, C.Q.; Cai, E.P.; Deng, Y.Z.; Mei, D.; Qiu, S.X.; Chen, B.S.; Zhang, L.H.; Jiang, Z. cAMP/PKA signalling pathway regulates redox homeostasis essential for Sporisorium scitamineum mating/filamentation and virulence. Environ. Microbiol. 2019, 21, 959–971. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; He, X.; Li, W.; Lu, Y.; Zhang, B. Regulation role of sterol C-24 methyltransferase and sterol C-8 isomerase in the ergosterol biosynthesis of Saccharomyces cerevisiae. Acta Microbiol. Sin. 2009, 49, 1063–1068. [Google Scholar]
- Chen, H.; Fujita, M.; Feng, Q.; Clardy, J.; Fink, G.R. Tyrosol is a quorum-sensing molecule in Candida albicans. Proc. Natl. Acad. Sci. USA 2004, 101, 5048–5052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using realtime quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Li, H.Z.; Cai, Y.C.; Deng, Q.Q.; Bao, H.; Chen, J.W.; Shen, W.K. Ovarian tumor domain-containing proteases-deubiquitylation enzyme gene SsCI33130 involved in the regulation of mating/filamentation and pathogenicity in Sporisorium scitamineum. Front. Microbiol. 2021, 12, 2922. [Google Scholar] [CrossRef]
- Petranyi, G.; Ryder, N.S.; Stuetz, A. Allylamine derivatives: New class of synthetic antifungal agents inhibiting fungal squalene epoxidase. Science 1984, 224, 1239–1241. [Google Scholar] [CrossRef]
- He, X.P.; Zhang, B.R. Advances in microbial ergosterol. Microbiol. China 1998, 3, 166–169. [Google Scholar]
- Leber, R.; Zenz, R.; Schröttner, K.; Fuchsbichler, S.; Pühringer, B.; Turnowsky, F. A novel sequence element is involved in the transcriptional regulation of expression of the erg1 (squalene epoxidase) gene in Saccharomyces cerevisiae. FEBS. J. 2010, 268, 914–924. [Google Scholar]
- Espenshade, P.J.; Hughes, A.L. Regulation of sterol synthesis in eukaryotes. Annu. Rev. Genet. 2007, 41, 401–427. [Google Scholar] [CrossRef] [PubMed]
- Sturley, S.L. Conservation of eukaryotic sterol homeo stasis: New insights from studies in budding yeast. BBA-Rev. Cancer 2000, 1529, 155–163. [Google Scholar]
- Jordá, T.; Puig, S. Regulation of ergosterol biosynthesis in Saccharomyces cerevisiae. Genes 2020, 11, 795. [Google Scholar] [CrossRef] [PubMed]
- Alcazar, L.; Mellado, E. Ergosterol biosynthesis in Aspergillus fumigatus: Its relevance as an antifungal target and role in antifungal drug resistance. Front. Microbiol. 2012, 3, 439. [Google Scholar] [CrossRef] [Green Version]
- Jin, H.; Michael, J.M.; Grote, E. Ergosterol promotes pheromone signaling and plasma membrane fusion in mating yeast. J. Cell Biol. 2008, 180, 813–826. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.; Jung, W.H.; Kronstad, J.W. The cAMP/protein kinase a signaling pathway in pathogenic basidio mycete fungi: Connections with iron homeostasis. J. Microbiol. 2015, 53, 579–587. [Google Scholar] [CrossRef] [Green Version]
- Hogan, D.A.; Sundstrom, P. The Ras/cAMP/PKA signaling pathway and virulence in Candida albicans. Future Microbiol. 2009, 4, 1263–1270. [Google Scholar] [CrossRef]
- Klosterman, S.J.; Perlin, M.H.; Garcia, P.M.; Covert, S.F.; Gold, S.E. Genetics of morphogenesis and pathogenic development of ustilago maydis. Adv. Genet. 2007, 57, 1–47. [Google Scholar]
- Davis, A.; Piispanen, A.E.; Stateva, L.I.; Hogan, D.A. Farnesol and dodecanol effects on the Candida albicans Ras1-cAMP signalling pathway and the regulation of morphogenesis. Mol. Microbiol. 2008, 67, 47–62. [Google Scholar] [CrossRef] [Green Version]
- Wongsuk, T.; Pumeesat, P.; Luplertlop, N. Fungal quorum sensing molecules: Role in fungal morphogenesis and pathogenicity. J. Basic Microbiol. 2016, 56, 440–447. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Primer Sequences (5′-3′) | Description |
|---|---|---|
| SsCI80130-LB-F | CTCAAAGCGGCCATCCTTG | |
| SsCI80130-LB-R | GTCGTGACTGGGAAAACCCTGAGACTTGGCGG GATTGTCAC | |
| SsCI80130-RB-F | GGTCATAGCTGTTTCCTGTGTGAGTGCGAGATT GGTGTCGGTAG | Deletion |
| SsCI80130-RB-R | TGAAGATGAGCTCGTTGGGC | construction |
| Hpt-LB-F | CAGGGTTTTCCCAGTCACGAC | |
| Hpt-LB-R | GGTCAAGACCAATGCGGAGC | |
| Hpt-RB-F | GCAAGACCTGCCTGAAACCG | |
| Hpt-RB-R | TCACACAGGAAACAGCTATGACC | |
| SsCI80130-IN-F SsCI80130-IN-R SsCI80130-OU-F SsCI80130-OU-R | TGGACTTCATCTCTGTGAACAC CGAGATCTACTCAAGGACGAAC TCTCAACAAGGGGGGCTACT GTTTTCGCGTGTGCTGATC | PCR verification |
| 80130COM-F 80130COM-R COM-HPT-LB-F Zeocin-R Situ-F COM-HPT-RB-R | ATCCAAGCTCAAGCTAAGCTTATACACTCGCCAC ATCGGTGC CAGCAAGATCTAATCAAGCTTGGGACAGAGCATA GGACAGCC GCGCGCGTAATACGACTCAC GAAGTGCACGCAGTTGCCG CTCCGTGTTGATGCTGGGAC CGAGCATTCACTAGGCAACCA | Complementation construction |
| Zeocin-IN-F Zeocin-IN-R | CTGTGATCAGCAGCCAAT GTCAACTTGGCCATGGTG | PCR verification |
| SsCI80130-qF | GATCTCATGATGCCGGCAGA | |
| SsCI80130-qR Aro8-qF Aro8-qR Uac1-qF Uac1-qR | TGCTTCGTAGGGTGTGACAC CCTGGTGTTGCGTTCATTCC CAAGCTCGGGCATCGTCTTA CTGACGGAGATGTAGCCAAAG AACGAGACAAGGAGGGAGTA | qRT-PCR |
| Actin-qF | ACAGGACGGCCTGGATAG | |
| Actin-qR | TCACCAACTGGGACGACA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cai, Y.; Zhang, Y.; Bao, H.; Chen, J.; Chen, J.; Shen, W. Squalene Monooxygenase Gene SsCI80130 Regulates Sporisorium scitamineum Mating/Filamentation and Pathogenicity. J. Fungi 2022, 8, 470. https://doi.org/10.3390/jof8050470
Cai Y, Zhang Y, Bao H, Chen J, Chen J, Shen W. Squalene Monooxygenase Gene SsCI80130 Regulates Sporisorium scitamineum Mating/Filamentation and Pathogenicity. Journal of Fungi. 2022; 8(5):470. https://doi.org/10.3390/jof8050470
Chicago/Turabian StyleCai, Yichang, Yi Zhang, Han Bao, Jiaoyun Chen, Jianwen Chen, and Wankuan Shen. 2022. "Squalene Monooxygenase Gene SsCI80130 Regulates Sporisorium scitamineum Mating/Filamentation and Pathogenicity" Journal of Fungi 8, no. 5: 470. https://doi.org/10.3390/jof8050470
APA StyleCai, Y., Zhang, Y., Bao, H., Chen, J., Chen, J., & Shen, W. (2022). Squalene Monooxygenase Gene SsCI80130 Regulates Sporisorium scitamineum Mating/Filamentation and Pathogenicity. Journal of Fungi, 8(5), 470. https://doi.org/10.3390/jof8050470