
Systemic Infection by Non-albicans Candida Species Affects the Development of a Murine Model of Multiple Sclerosis

 ,
,  , , , ,
, , , ,  , and
, and 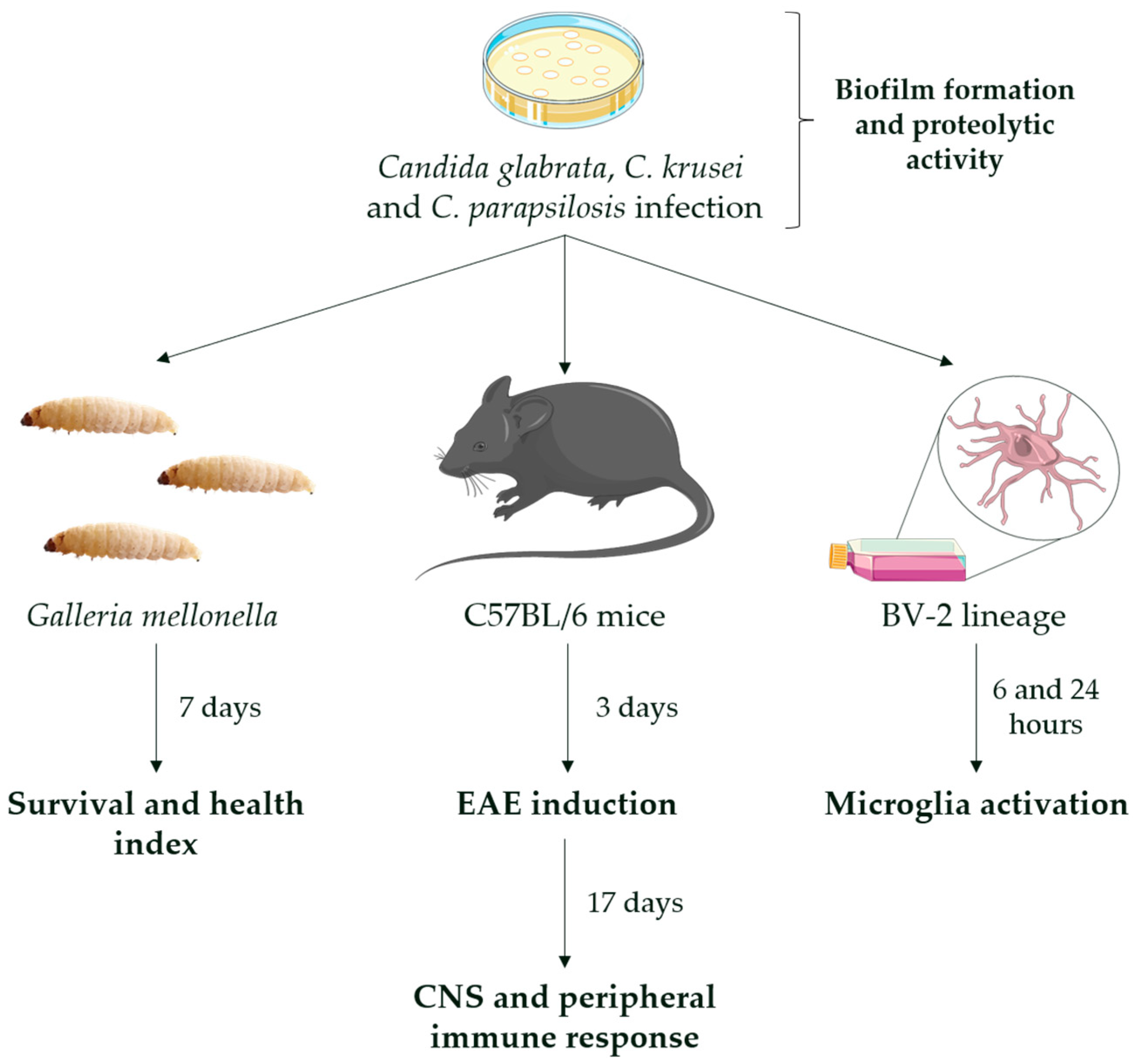{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Non-albicans Candida Strains
2.2. Animal Models
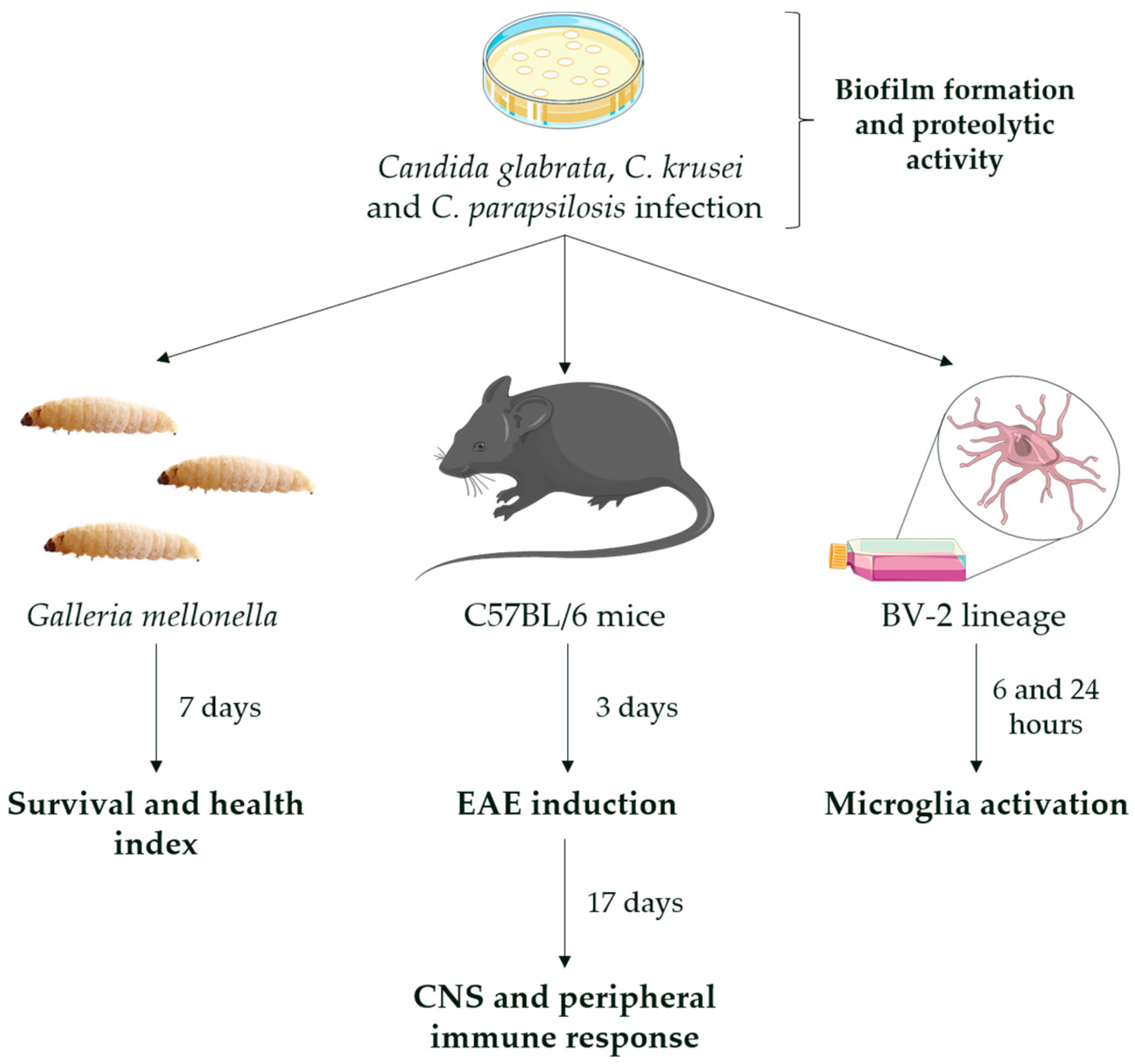
2.3. Experimental Design
2.4. Biofilm and Proteolytic Activity
2.5. G. mellonella Assay
2.6. Candida Infections in Mice
2.7. EAE Induction and Clinical Evaluation
2.8. Fungal Load Determination
2.9. Histopathological Evaluation
2.10. RT-PCR
2.11. Splenic Cell Cultures
2.12. Isolation of CNS Mononuclear Cells
2.13. Microglial Cell Culture
2.14. Cytokine Quantification
2.15. Flow Cytometry
2.16. Statistical Analysis
3. Results
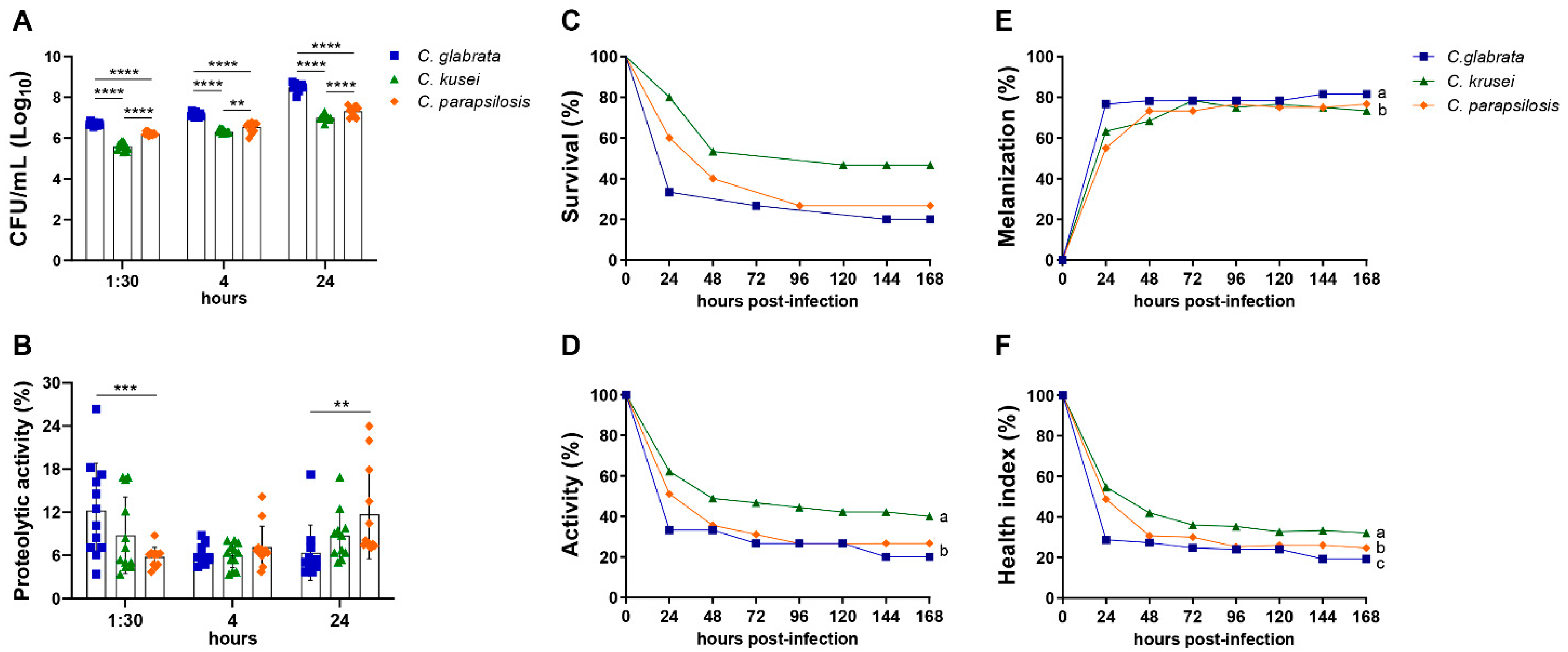
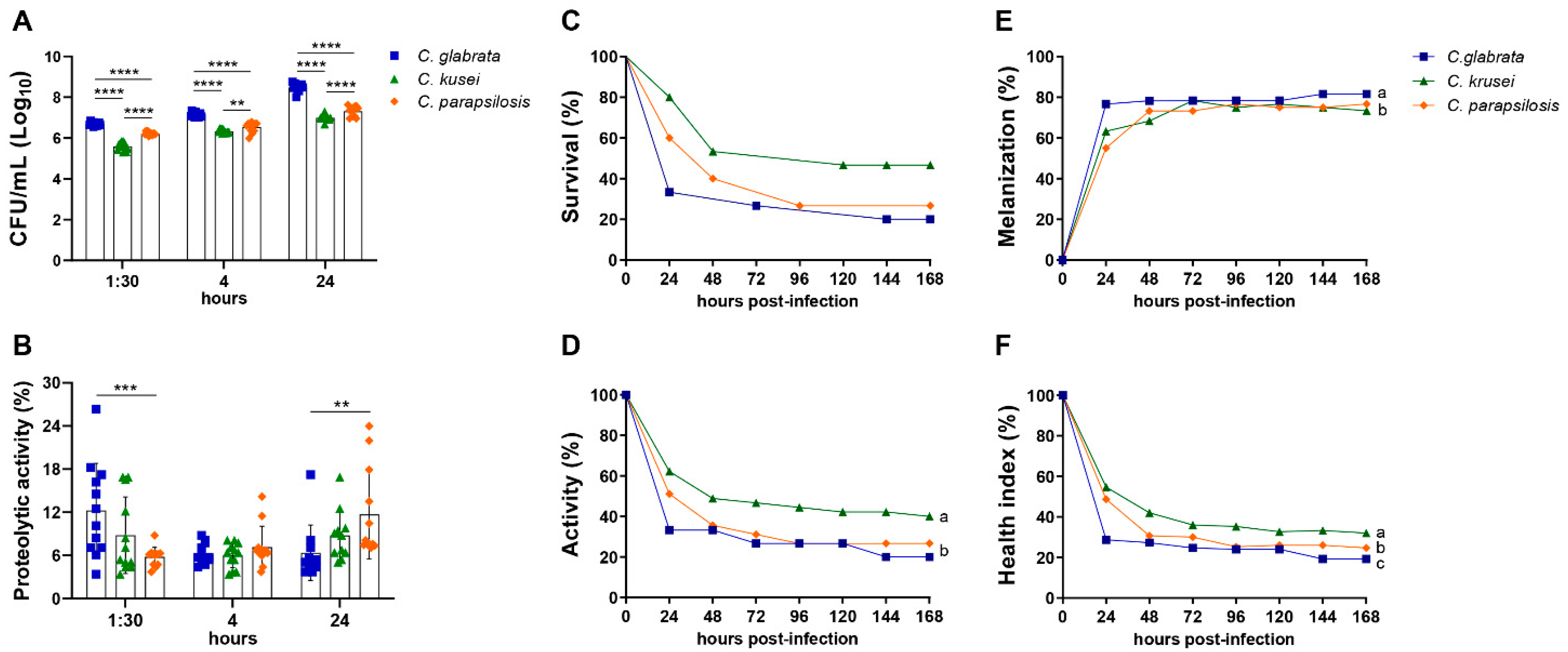
3.1. Biofilm Formation, Proteolytic Activity, and Infection of G. mellonella Showed Variable Virulence Profiles of Candida Strains
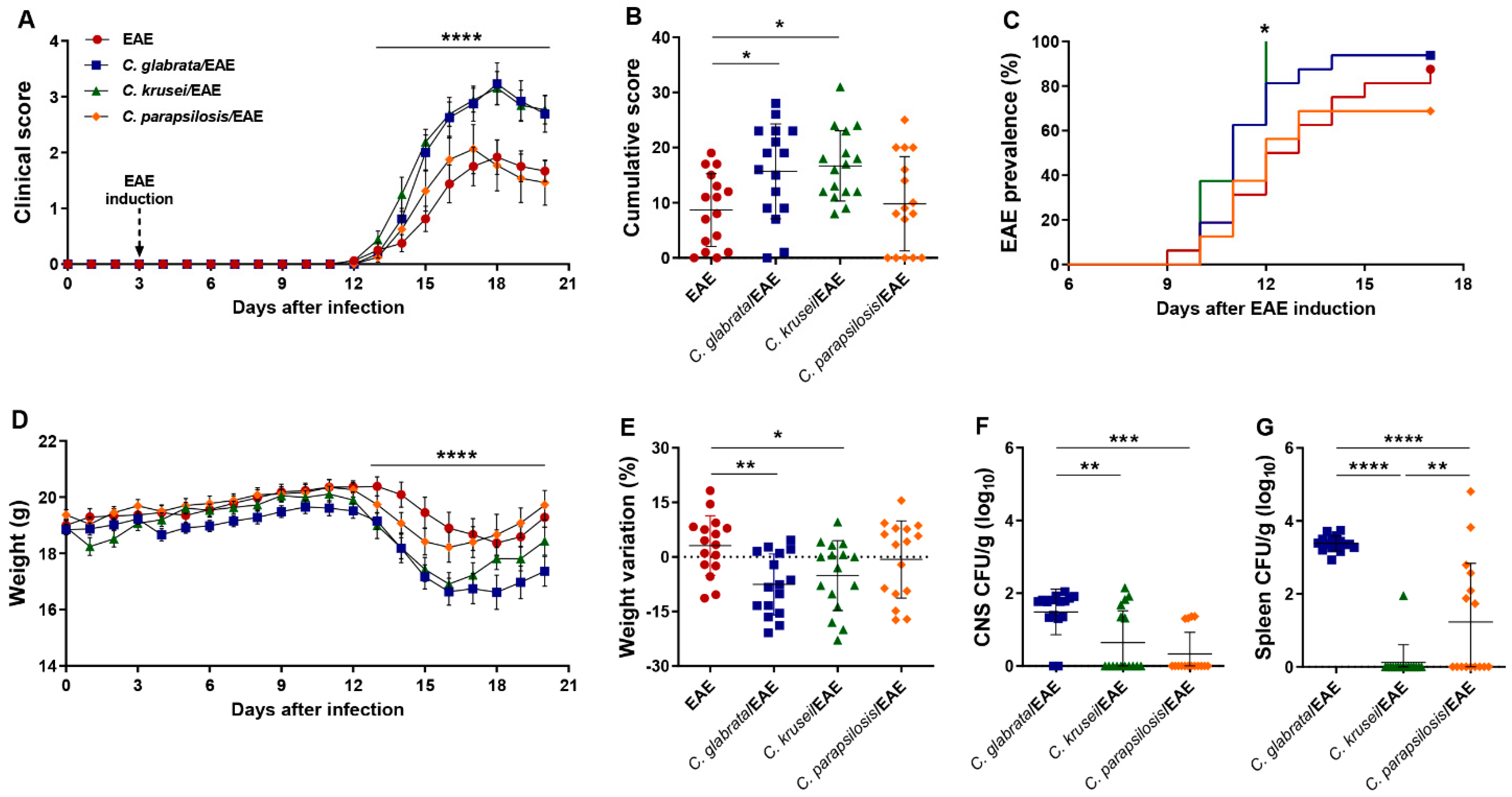
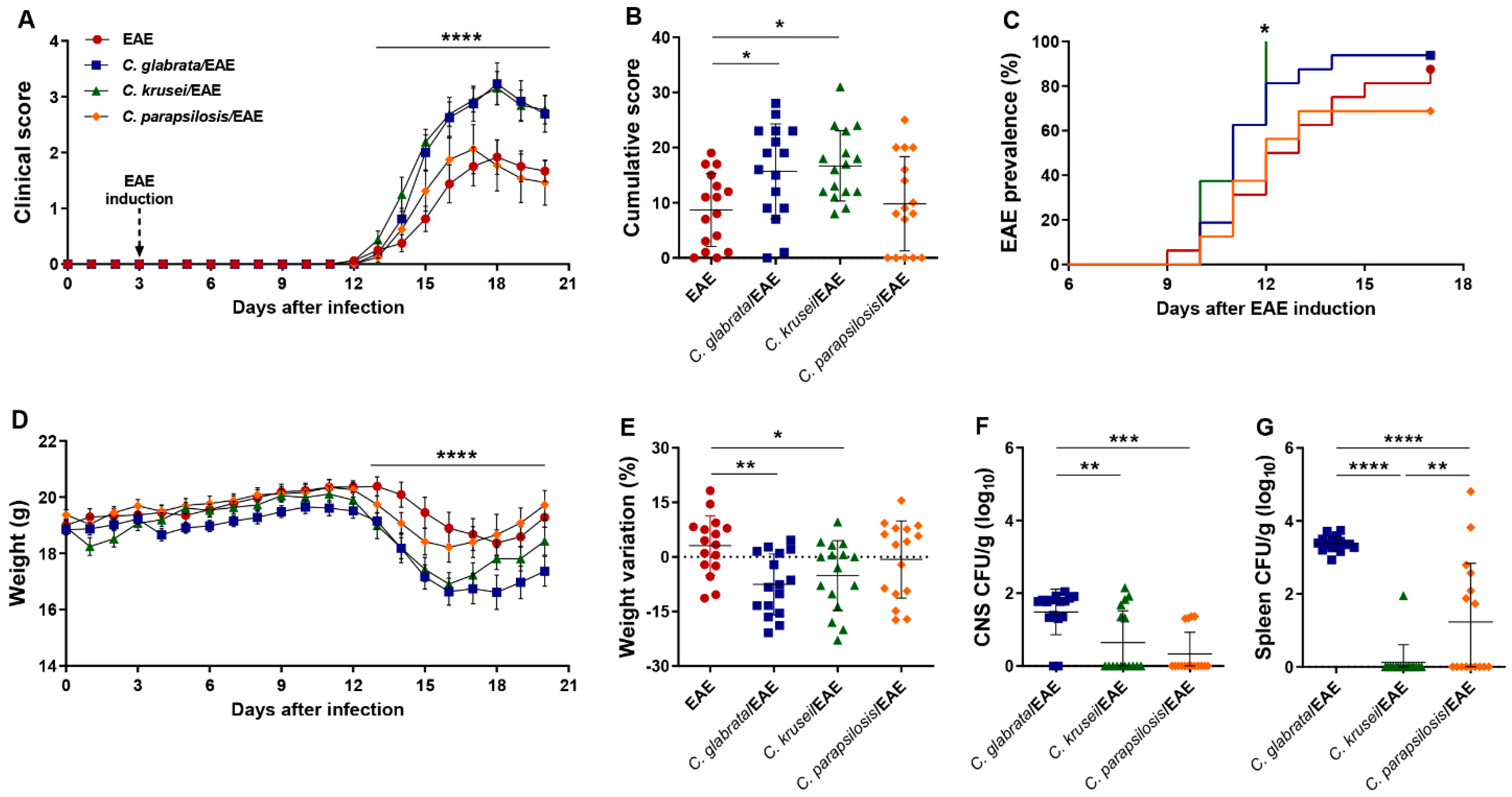
3.2. Previous Infection with C. glabrata and C. krusei Strains Aggravated EAE Clinical Signs
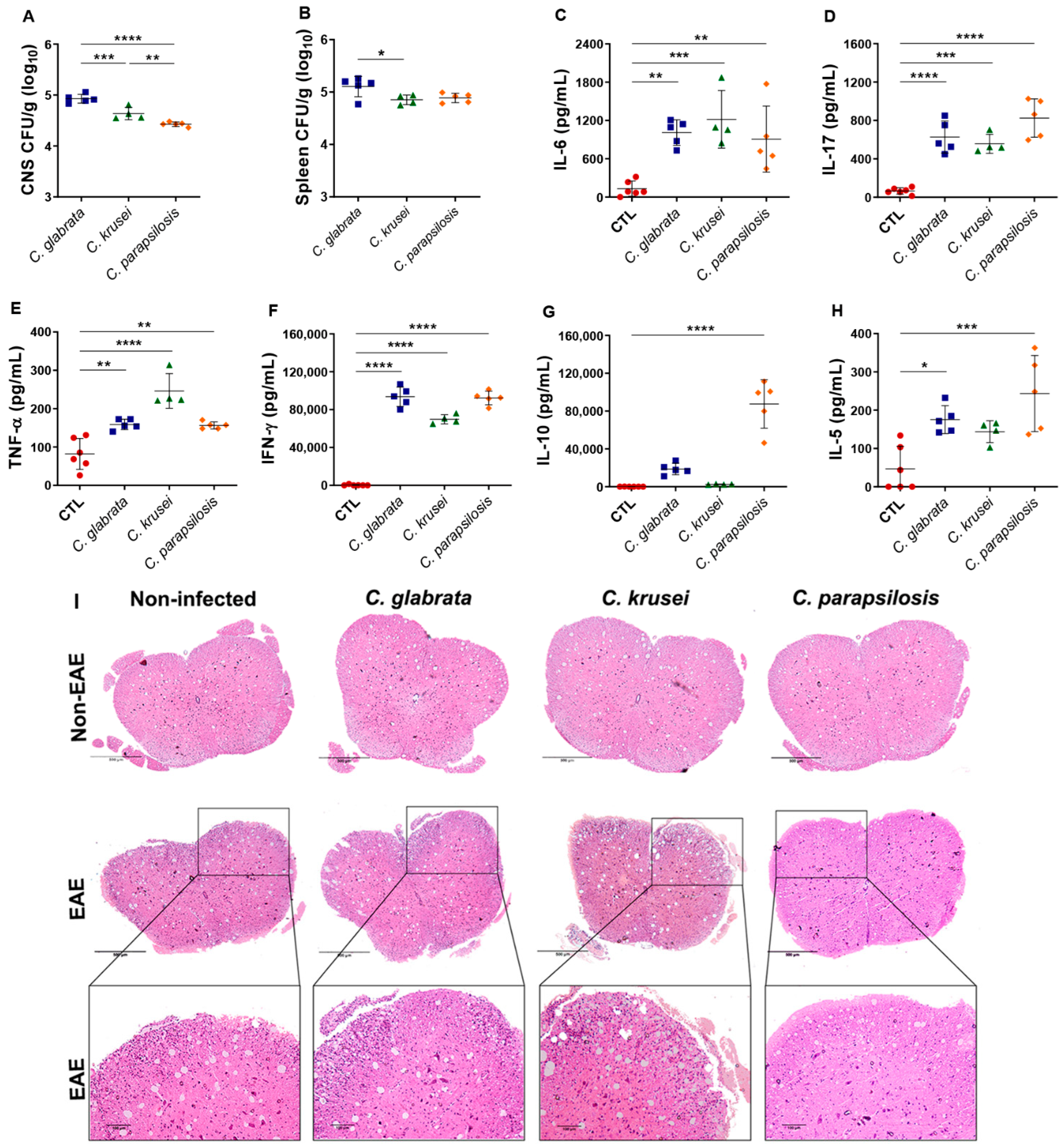
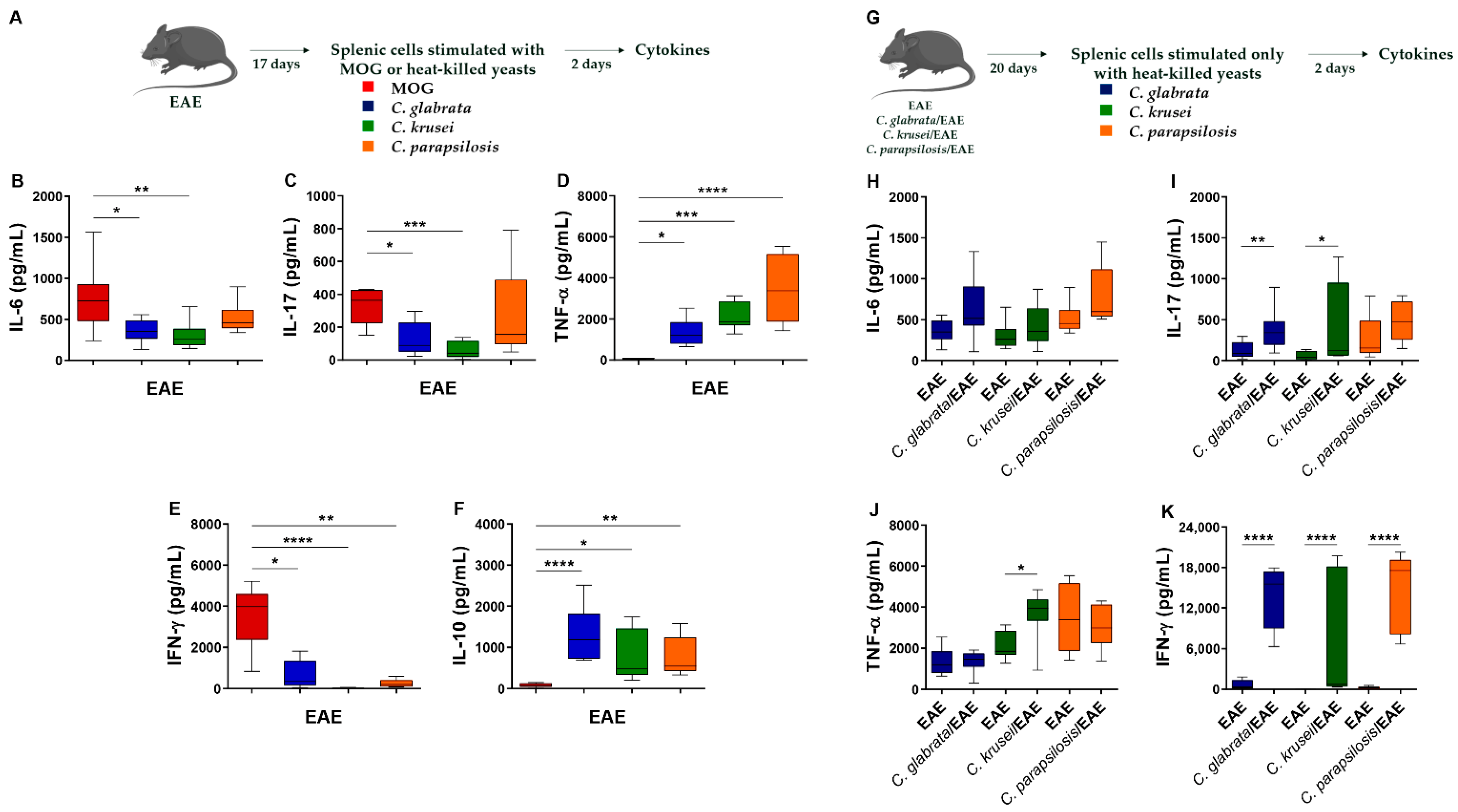
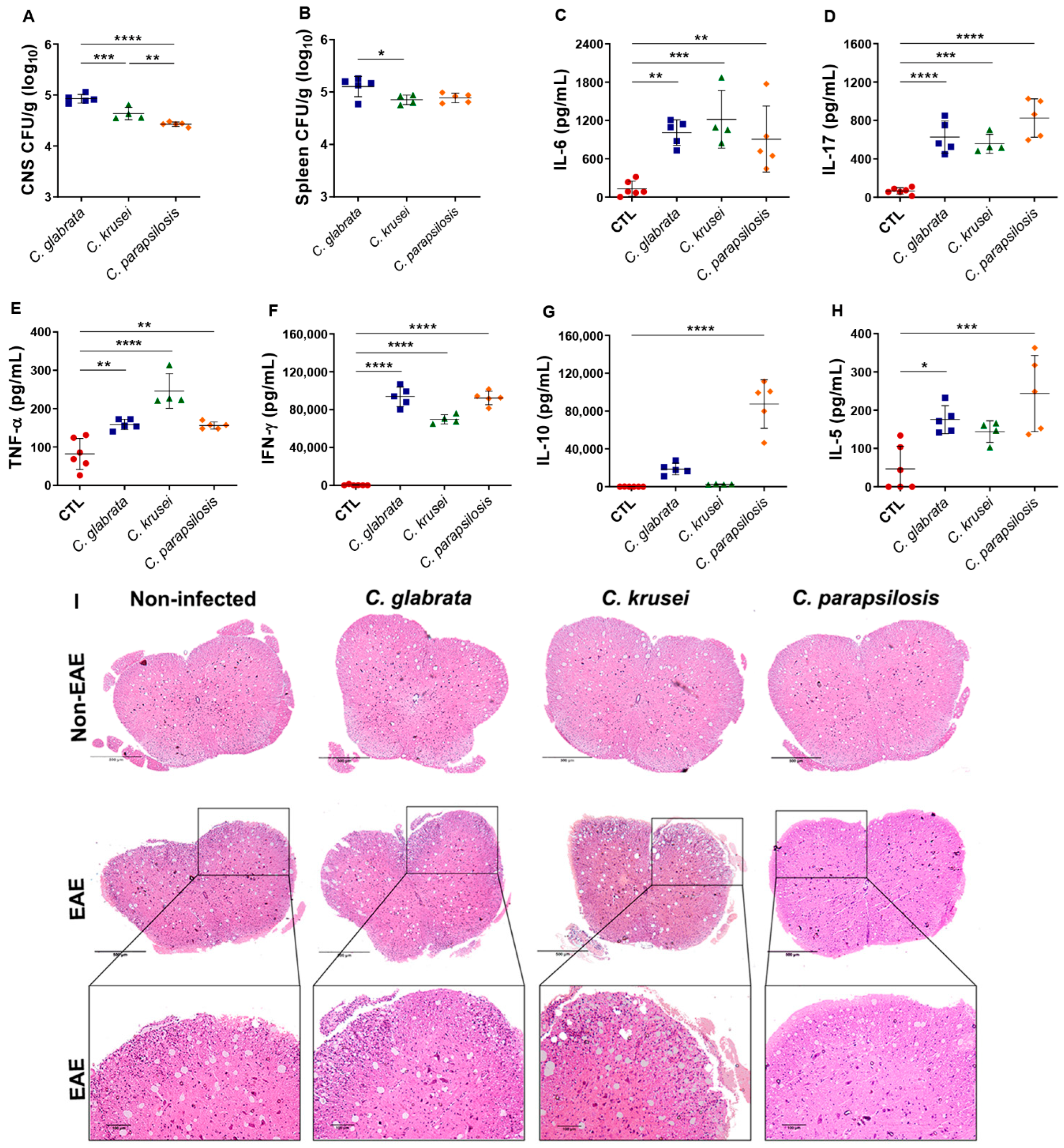
3.3. Th1/Th17 Profiles Induced by C. glabrata and C. krusei Strains Persisted during EAE Development
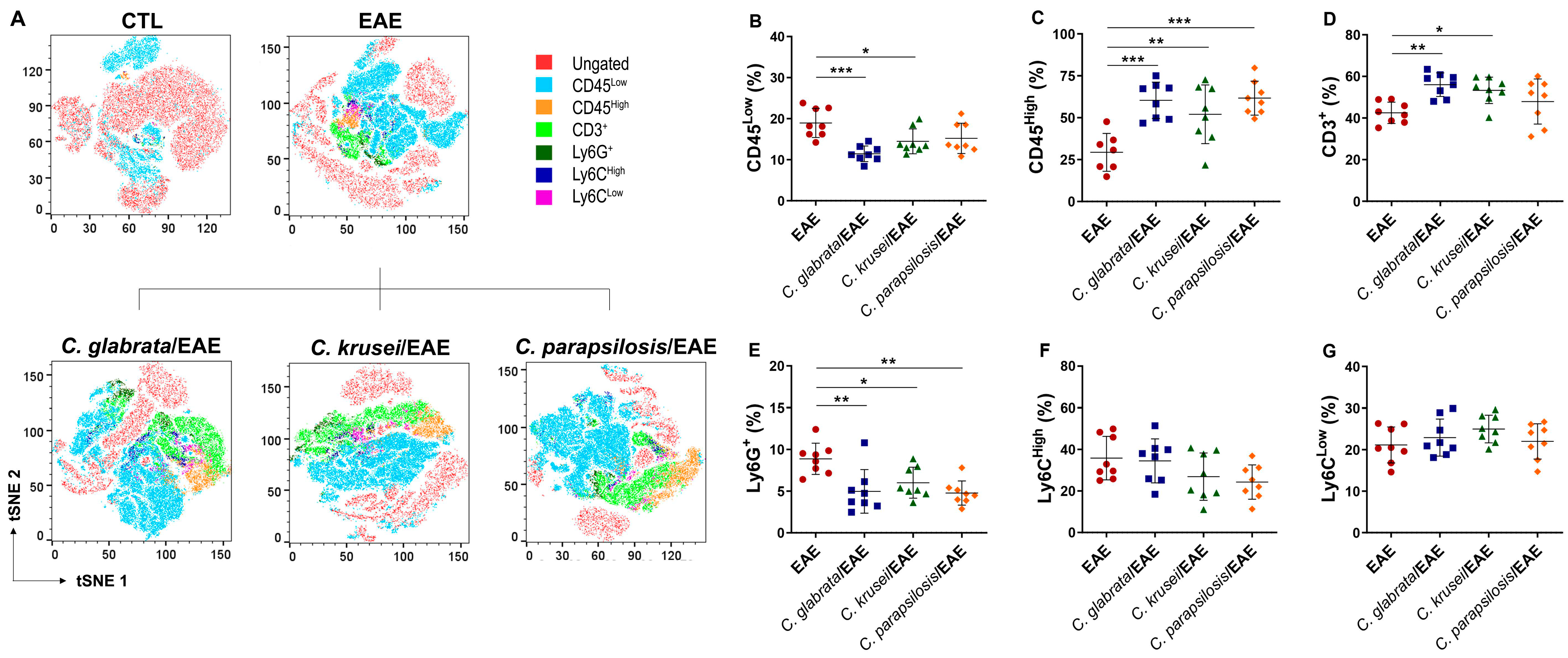
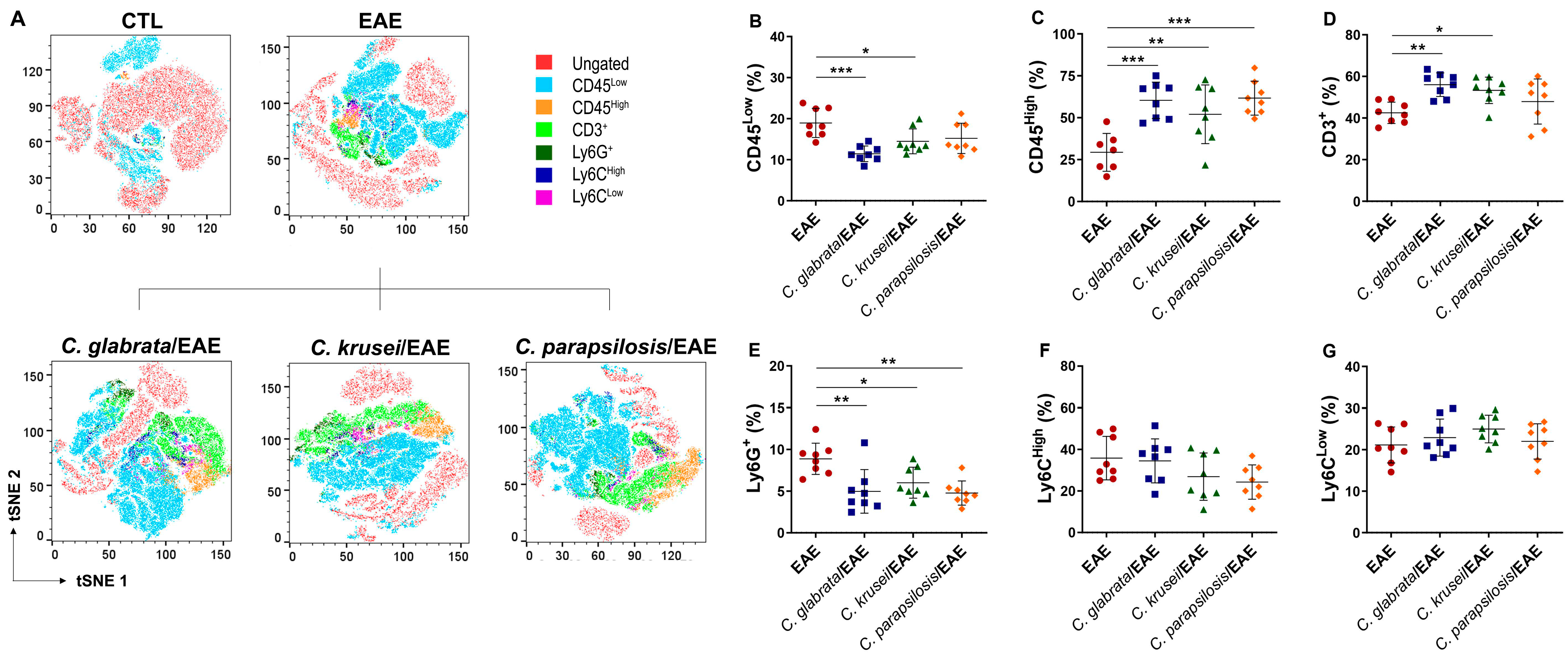
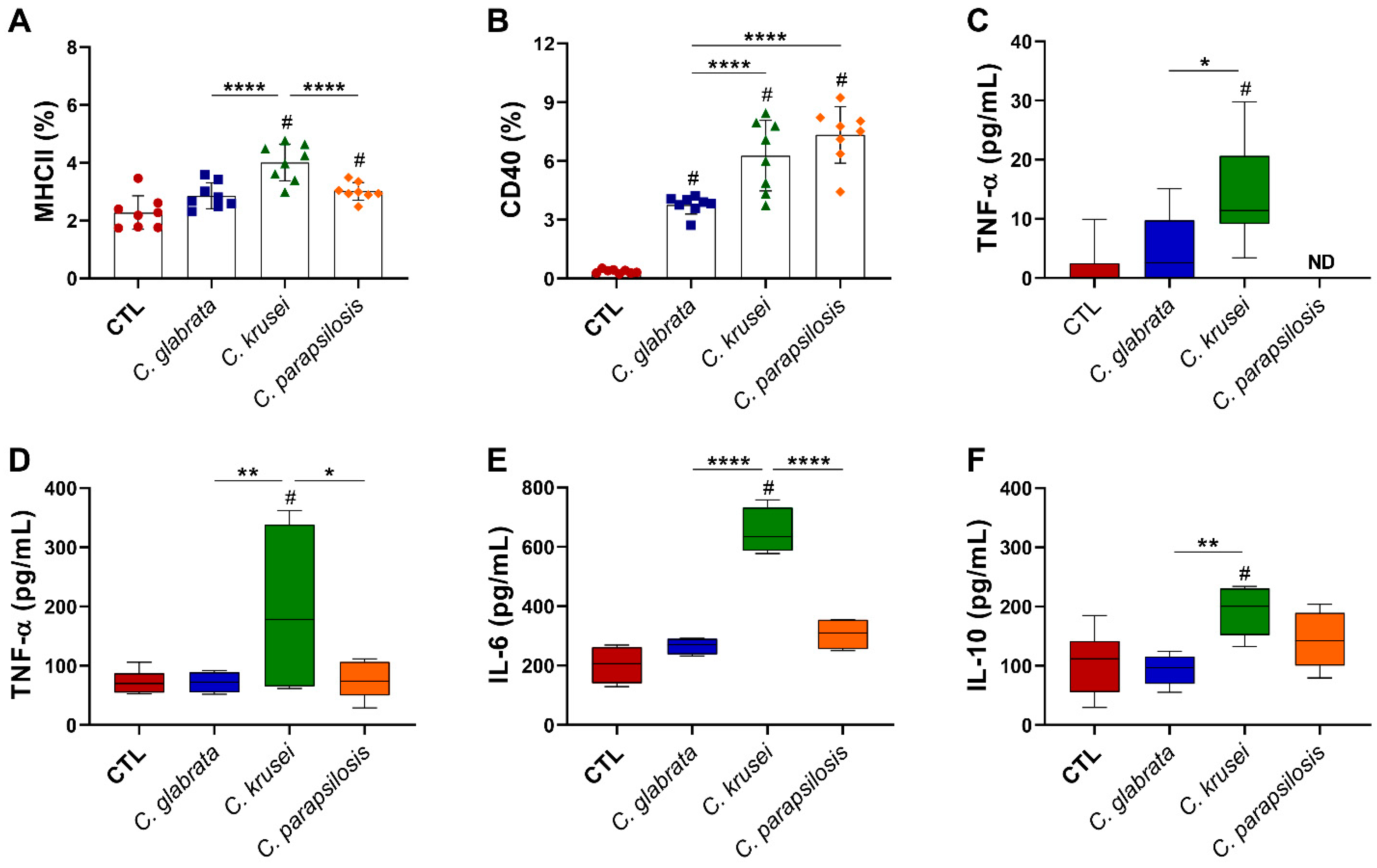
3.4. Previous Candida spp. Infection Increased Leukocyte Infiltration in the CNS of EAE Mice
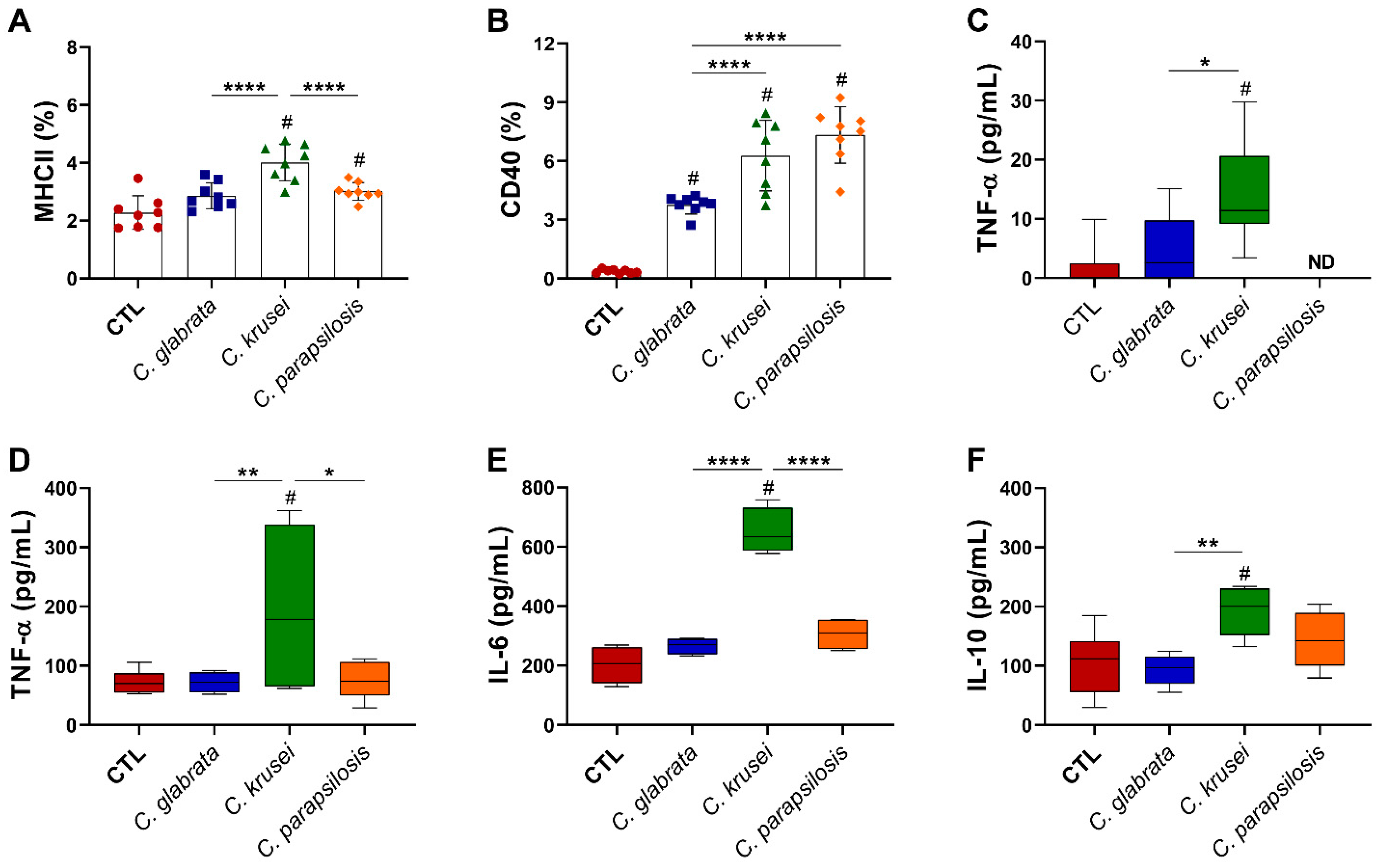
3.5. C. krusei Strain Was Highly Pro-Inflammatory for Microglia
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Romani, L. Immunity to fungal infections. Nat. Rev. Immunol. 2011, 11, 275–288. [Google Scholar] [CrossRef] [PubMed]
- Papon, N.; Courdavault, V.; Clastre, M.; Bennett, R.J. Emerging and Emerged Pathogenic Candida Species: Beyond the Candida albicans Paradigm. PLoS Pathog. 2013, 9, e1003550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadeghi, G.; Ebrahimi-Rad, M.; Mousavi, S.F.; Shams-Ghahfarokhi, M.; Razzaghi-Abyaneh, M. Emergence of non- Candida albicans species: Epidemiology, phylogeny and fluconazole susceptibility profile. J. Mycol. Med. 2018, 28, 51–58. [Google Scholar] [CrossRef] [PubMed]
- McCarty, T.P.; Pappas, P.G. Invasive Candidiasis. Infect. Dis. Clin. N. Am. 2016, 30, 103–124. [Google Scholar] [CrossRef] [PubMed]
- Panackal, A.A.; Williamson, P.R. Fungal Infections of the Central Nervous System. Contin. Lifelong Learn. Neurol. 2015, 21, 1662–1678. [Google Scholar] [CrossRef] [PubMed]
- Pfaller, M.A.; Andes, D.R.; Diekema, D.J.; Horn, D.L.; Reboli, A.C.; Rotstein, C.; Franks, B.; Azie, N.E. Epidemiology and Outcomes of Invasive Candidiasis Due to Non-albicans Species of Candida in 2496 Patients: Data from the Prospective Antifungal Therapy (PATH) Registry 2004–2008. PLoS ONE 2014, 9, e101510. [Google Scholar] [CrossRef] [Green Version]
- Pfaller, M.A.; Moet, G.J.; Messer, S.A.; Jones, R.N.; Castanheira, M. Geographic Variations in Species Distribution and Echinocandin and Azole Antifungal Resistance Rates among Candida Bloodstream Infection Isolates: Report from the SENTRY Antimicrobial Surveillance Program (2008 to 2009). J. Clin. Microbiol. 2011, 49, 396–399. [Google Scholar] [CrossRef] [Green Version]
- Nucci, M.; Queiroz-Telles, F.; Tobón, A.M.; Restrepo, A.; Colombo, A.L. Epidemiology of Opportunistic Fungal Infections in Latin America. Clin. Infect. Dis. 2010, 51, 561–570. [Google Scholar] [CrossRef]
- De Medeiros, M.A.P.; de Melo, A.P.V.; de Oliveira Bento, A.; de Souza, L.B.F.C.; de Assis Bezerra Neto, F.; Garcia, J.B.-L.; Zuza-Alves, D.L.; Francisco, E.C.; de Azevedo Melo, A.S.; Chaves, G.M. Epidemiology and prognostic factors of nosocomial candidemia in Northeast Brazil: A six-year retrospective study. PLoS ONE 2019, 14, e0221033. [Google Scholar] [CrossRef]
- Da Matta, D.A.; Souza, A.C.R.; Colombo, A.L. Revisiting Species Distribution and Antifungal Susceptibility of Candida Bloodstream Isolates from Latin American Medical Centers. J. Fungi 2017, 3, 24. [Google Scholar] [CrossRef] [Green Version]
- Yapar, N. Epidemiology and risk factors for invasive candidiasis. Ther. Clin. Risk Manag. 2014, 10, 95–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamoth, F.; Lockhart, S.R.; Berkow, E.L.; Calandra, T. Changes in the epidemiological landscape of invasive candidiasis. J. Antimicrob. Chemother. 2018, 73, i4–i13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pappas, P.G.; Lionakis, M.S.; Arendrup, M.C.; Ostrosky-Zeichner, L.; Kullberg, B.J. Invasive candidiasis. Nat. Rev. Dis. Prim. 2018, 4, 18026. [Google Scholar] [CrossRef] [PubMed]
- Romani, L. Parasites and autoimmunity: The case of fungi. Autoimmun. Rev. 2008, 8, 129–133. [Google Scholar] [CrossRef]
- Roe, K. How major fungal infections can initiate severe autoimmune diseases. Microb. Pathog. 2021, 161, 105200. [Google Scholar] [CrossRef]
- Wootla, B.; Eriguchi, M.; Rodriguez, M. Is Multiple Sclerosis an Autoimmune Disease? Autoimmune Dis. 2012, 2012, 969657. [Google Scholar] [CrossRef]
- Rodriguez, M. Effectors of Demyelination and Remyelination in the CNS: Implications for Multiple Sclerosis. Brain Pathol. 2007, 17, 219–229. [Google Scholar] [CrossRef]
- Sospedra, M.; Martin, R. Immunology of Multiple Sclerosis. Semin. Neurol. 2005, 36, 115–127. [Google Scholar] [CrossRef]
- Vojdani, A.; Rahimian, P.; Kalhor, H.; Mordechai, E. Immunological cross reactivity between Candida albicans and human tissue. J. Clin. Lab. Immunol. 1996, 48, 1–15. [Google Scholar]
- Purzycki, C.B.; Shain, D.H. Fungal toxins and multiple sclerosis: A compelling connection. Brain Res. Bull. 2010, 82, 4–6. [Google Scholar] [CrossRef]
- Lionakis, M.S.; Lim, J.K.; Lee, C.-C.R.; Murphy, P.M. Organ-Specific Innate Immune Responses in a Mouse Model of Invasive Candidiasis. J. Innate Immun. 2011, 3, 180–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Campos Fraga-Silva, T.F.; Mimura, L.A.N.; de Campos Toledo Leite, L.; Borim, P.A.; Ishikawa, L.L.W.; Venturini, J.; de Arruda, M.S.P.; Sartori, A. Gliotoxin Aggravates Experimental Autoimmune Encephalomyelitis by Triggering Neuroinflammation. Toxins 2019, 11, 443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benito-León, J.; Pisa, D.; Alonso, R.; Calleja, P.; Díaz-Sánchez, M.; Carrasco, L. Association between multiple sclerosis and Candida species: Evidence from a case-control study. Eur. J. Clin. Microbiol. Infect. Dis. 2010, 29, 1139–1145. [Google Scholar] [CrossRef] [PubMed]
- Benito-León, J.; Laurence, M. The role of fungi in the etiology of multiple sclerosis. Front. Neurol. 2017, 8, 535. [Google Scholar] [CrossRef]
- Pisa, D.; Alonso, R.; Carrasco, L. Fungal infection in a patient with multiple sclerosis. Eur. J. Clin. Microbiol. Infect. Dis. 2011, 30, 1173–1180. [Google Scholar] [CrossRef]
- Pisa, D.; Alonso, R.; Jiménez-Jiménez, F.J.; Carrasco, L. Fungal infection in cerebrospinal fluid from some patients with multiple sclerosis. Eur. J. Clin. Microbiol. Infect. Dis. 2013, 32, 795–801. [Google Scholar] [CrossRef] [Green Version]
- Da Cunha, E.T.S.; Figueiredo-Godoi, L.M.A.; Santos, D.H.; Carneiro, R.P.C.D.; do Olival, G.S.; de Barros, P.P.; Narimatsu, K.; Tilbery, C.P.; Junqueira, J.C. Oral Colonization by Candida Species in Patients with Multiple Sclerosis. Mycopathologia 2020, 185, 983–991. [Google Scholar] [CrossRef]
- Saroukolaei, S.A.; Ghabaee, M.; Shokri, H.; Badiei, A.; Ghourchian, S. The role of Candida albicans in the severity of multiple sclerosis. Mycoses 2016, 59, 697–704. [Google Scholar] [CrossRef]
- Baxter, A.G. The origin and application of experimental autoimmune encephalomyelitis. Nat. Rev. Immunol. 2007, 7, 904–912. [Google Scholar] [CrossRef]
- Constantinescu, C.S.; Farooqi, N.; O’Brien, K.; Gran, B. Experimental autoimmune encephalomyelitis (EAE) as a model for multiple sclerosis (MS). Br. J. Pharmacol. 2011, 164, 1079–1106. [Google Scholar] [CrossRef]
- Lavi, E.; Constantinescu, C.S. Experimental Models of Multiple Sclerosis. Am. J. Neuroradiol. 2007, 28, 190–191. [Google Scholar]
- Fraga-Silva, T.F.C.; Mimura, L.A.N.; Marchetti, C.M.; Chiuso-Minicucci, F.; França, T.G.D.; Zorzella-Pezavento, S.F.G.; Venturini, J.; Arruda, M.S.P.; Sartori, A. Experimental autoimmune encephalomyelitis development is aggravated by Candida albicans infection. J. Immunol. Res. 2015, 2015, 635052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fraga-Silva, T.F.C.; Venturini, J.; de Arruda, M.S.P. Trafficking of phagocytic peritoneal cells in hypoinsulinemic-hyperglycemic mice with systemic candidiasis. BMC Infect. Dis. 2013, 13, 147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanches, M.D.; Mimura, L.A.N.; Oliveira, L.R.C.; Ishikawa, L.L.W.; Garces, H.G.; Bagagli, E.; Sartori, A.; Kurokawa, C.S.; Fraga-Silva, T.F.C. Differential Behavior of Non-albicans Candida Species in the Central Nervous System of Immunocompetent and Immunosuppressed Mice. Front. Microbiol. 2019, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Almeida, N.L.M.; Godoi, L.M.A.F.; Venturini, J.; Leonardo, L.; Saldanha; Dokkedal, A.L.; Junqueira, J.C.; Lara, V.S. Antimicrobial and immunomodulatory activity of pomegranate in the systemic candidiasis on Galleria mellonella. J. Clin. Exp. Toxicol. 2018, 2, 8–17. [Google Scholar] [CrossRef]
- Loh, J.M.; Adenwalla, N.; Wiles, S.; Proft, T. Galleria mellonella larvae as an infection model for group A streptococcus. Virulence 2013, 4, 419–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saroukolaei, S.A.; Ghabaee, M.; Shokri, H.; Khosravi, A.; Badiei, A. Evaluation of APR1 Gene Expression in Candida albicans Strains Isolated From Patients With Multiple Sclerosis. Jundishapur J. Microbiol. 2016, 9, e60074. [Google Scholar] [CrossRef] [Green Version]
- Kadosh, D.; Mundodi, V. A Re-Evaluation of the Relationship between Morphology and Pathogenicity in Candida Species. J. Fungi 2020, 6, 13. [Google Scholar] [CrossRef] [Green Version]
- Calderone, R.A.; Fonzi, W.A. Virulence factors of Candida albicans. Trends Microbiol. 2001, 9, 327–335. [Google Scholar] [CrossRef]
- Priest, S.J.; Lorenz, M.C. Characterization of Virulence-Related Phenotypes in Candida Species of the CUG Clade. Eukaryot. Cell 2015, 14, 931–940. [Google Scholar] [CrossRef] [Green Version]
- Krcmery, V.; Barnes, A.J. Non-albicans Candida spp. causing fungaemia: Pathogenicity and antifungal resistance. J. Hosp. Infect. 2002, 50, 243–260. [Google Scholar] [CrossRef] [PubMed]
- Butler, G.; Rasmussen, M.D.; Lin, M.F.; Santos, M.A.S.; Sakthikumar, S.; Munro, C.A.; Rheinbay, E.; Grabherr, M.; Forche, A.; Reedy, J.L.; et al. Evolution of pathogenicity and sexual reproduction in eight Candida genomes. Nature 2009, 459, 657–662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nouraei, H.; Pakshir, K.; ZareShahrabadi, Z.; Zomorodian, K. High detection of virulence factors by Candida species isolated from bloodstream of patients with candidemia. Microb. Pathog. 2020, 149, 104574. [Google Scholar] [CrossRef] [PubMed]
- Cavalheiro, M.; Teixeira, M.C. Candida Biofilms: Threats, Challenges, and Promising Strategies. Front. Med. 2018, 5, 28. [Google Scholar] [CrossRef] [Green Version]
- Eix, E.F.; Nett, J.E. How Biofilm Growth Affects Candida-Host Interactions. Front. Microbiol. 2020, 11, 1437. [Google Scholar] [CrossRef]
- Naglik, J.R.; Challacombe, S.J.; Hube, B. Candida albicans Secreted Aspartyl Proteinases in Virulence and Pathogenesis. Microbiol. Mol. Biol. Rev. 2003, 67, 400–428. [Google Scholar] [CrossRef] [Green Version]
- Swidergall, M.; Filler, S.G. Oropharyngeal Candidiasis: Fungal Invasion and Epithelial Cell Responses. PLOS Pathog. 2017, 13, e1006056. [Google Scholar] [CrossRef]
- Villar, C.C.; Kashleva, H.; Nobile, C.J.; Mitchell, A.P.; Dongari-Bagtzoglou, A. Mucosal Tissue Invasion by Candida albicans Is Associated with E-Cadherin Degradation, Mediated by Transcription Factor Rim101p and Protease Sap5p. Infect. Immun. 2007, 75, 2126–2135. [Google Scholar] [CrossRef] [Green Version]
- Mroczyńska, M.; Brillowska-Dąbrowska, A. Virulence of Clinical Candida Isolates. Pathogens 2021, 10, 466. [Google Scholar] [CrossRef]
- Firacative, C.; Khan, A.; Duan, S.; Ferreira-Paim, K.; Leemon, D.; Meyer, W. Rearing and Maintenance of Galleria mellonella and Its Application to Study Fungal Virulence. J. Fungi 2020, 6, 130. [Google Scholar] [CrossRef]
- Ramarao, N.; Nielsen-Leroux, C.; Lereclus, D. The Insect Galleria mellonella as a Powerful Infection Model to Investigate Bacterial Pathogenesis. J. Vis. Exp. 2012, e4392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cirasola, D.; Sciota, R.; Vizzini, L.; Ricucci, V.; Morace, G.; Borghi, E. Experimental biofilm-related Candida infections. Future Microbiol. 2013, 8, 799–805. [Google Scholar] [CrossRef] [PubMed]
- Rossoni, R.D.; Barbosa, J.O.; Vilela, S.F.G.; Dos Santos, J.D.; Jorge, A.O.C.; Junqueira, J.C. Correlation of phospholipase and proteinase production of Candida with in vivo pathogenicity in Galleria mellonella. Braz. J. Oral Sci. 2013, 12, 199–204. [Google Scholar] [CrossRef] [Green Version]
- Göbel, K.; Ruck, T.; Meuth, S.G. Cytokine signaling in multiple sclerosis: Lost in translation. Mult. Scler. J. 2018, 24, 432–439. [Google Scholar] [CrossRef]
- Minagar, A.; Alexander, J.S. Blood-brain barrier disruption in multiple sclerosis. Mult. Scler. J. 2003, 9, 540–549. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, G.G.; Pacheco-Moisés, F.P.; Macías-Islas, M.Á.; Flores-Alvarado, L.J.; Mireles-Ramírez, M.A.; González-Renovato, E.D.; Hernández-Navarro, V.E.; Sánchez-López, A.L.; Alatorre-Jiménez, M.A. Role of the Blood–Brain Barrier in Multiple Sclerosis. Arch. Med. Res. 2014, 45, 687–697. [Google Scholar] [CrossRef] [PubMed]
- Banks, W. Blood-Brain Barrier Transport of Cytokines: A Mechanism for Neuropathology. Curr. Pharm. Des. 2005, 11, 973–984. [Google Scholar] [CrossRef]
- Berghmans, N.; Dillen, C.; Heremans, H. Exogenous IL-12 suppresses experimental autoimmune encephalomyelitis (EAE) by tuning IL-10 and IL-5 levels in an IFN-γ-dependent way. J. Neuroimmunol. 2006, 176, 63–75. [Google Scholar] [CrossRef]
- Netea, M.G.; Joosten, L.A.B.; van der Meer, J.W.M.; Kullberg, B.-J.; van de Veerdonk, F.L. Immune defence against Candida fungal infections. Nat. Rev. Immunol. 2015, 15, 630–642. [Google Scholar] [CrossRef]
- Dendrou, C.A.; Fugger, L.; Friese, M.A. Immunopathology of multiple sclerosis. Nat. Rev. Immunol. 2015, 15, 545–558. [Google Scholar] [CrossRef]
- Balasa, R.; Barcutean, L.; Balasa, A.; Motataianu, A.; Roman-Filip, C.; Manu, D. The action of TH17 cells on blood brain barrier in multiple sclerosis and experimental autoimmune encephalomyelitis. Hum. Immunol. 2020, 81, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Murphy, Á.C.; Lalor, S.J.; Lynch, M.A.; Mills, K.H.G. Brain, Behavior, and Immunity Infiltration of Th1 and Th17 cells and activation of microglia in the CNS during the course of experimental autoimmune encephalomyelitis. Brain Behav. Immun. 2010, 24, 641–651. [Google Scholar] [CrossRef] [PubMed]
- Cassone, A. Development of vaccines for Candida albicans: Fighting a skilled transformer. Nat. Rev. Microbiol. 2013, 11, 884–891. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fraga-Silva, T.F.d.C.; Munhoz-Alves, N.; Mimura, L.A.N.; Oliveira, L.R.C.d.; Figueiredo-Godoi, L.M.A.; Garcia, M.T.; Oliveira, E.S.; Ishikawa, L.L.W.; Zorzella-Pezavento, S.F.G.; Bonato, V.L.D.; et al. Systemic Infection by Non-albicans Candida Species Affects the Development of a Murine Model of Multiple Sclerosis. J. Fungi 2022, 8, 386. https://doi.org/10.3390/jof8040386
Fraga-Silva TFdC, Munhoz-Alves N, Mimura LAN, Oliveira LRCd, Figueiredo-Godoi LMA, Garcia MT, Oliveira ES, Ishikawa LLW, Zorzella-Pezavento SFG, Bonato VLD, et al. Systemic Infection by Non-albicans Candida Species Affects the Development of a Murine Model of Multiple Sclerosis. Journal of Fungi. 2022; 8(4):386. https://doi.org/10.3390/jof8040386
Chicago/Turabian StyleFraga-Silva, Thais Fernanda de Campos, Natália Munhoz-Alves, Luiza Ayumi Nishiyama Mimura, Larissa Ragozo Cardoso de Oliveira, Lívia Mara Alves Figueiredo-Godoi, Maíra Terra Garcia, Evelyn Silva Oliveira, Larissa Lumi Watanabe Ishikawa, Sofia Fernanda Gonçalves Zorzella-Pezavento, Vânia Luiza Deperon Bonato, and et al. 2022. "Systemic Infection by Non-albicans Candida Species Affects the Development of a Murine Model of Multiple Sclerosis" Journal of Fungi 8, no. 4: 386. https://doi.org/10.3390/jof8040386
APA StyleFraga-Silva, T. F. d. C., Munhoz-Alves, N., Mimura, L. A. N., Oliveira, L. R. C. d., Figueiredo-Godoi, L. M. A., Garcia, M. T., Oliveira, E. S., Ishikawa, L. L. W., Zorzella-Pezavento, S. F. G., Bonato, V. L. D., Junqueira, J. C., Bagagli, E., & Sartori, A. (2022). Systemic Infection by Non-albicans Candida Species Affects the Development of a Murine Model of Multiple Sclerosis. Journal of Fungi, 8(4), 386. https://doi.org/10.3390/jof8040386