
Structure of the Yeast Cell Wall Integrity Sensor Wsc1 Reveals an Essential Role of Surface-Exposed Aromatic Clusters

 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Yeast Strains and Growth Tests
2.2. Plasmids
2.3. Recombinant Production of CRD Proteins
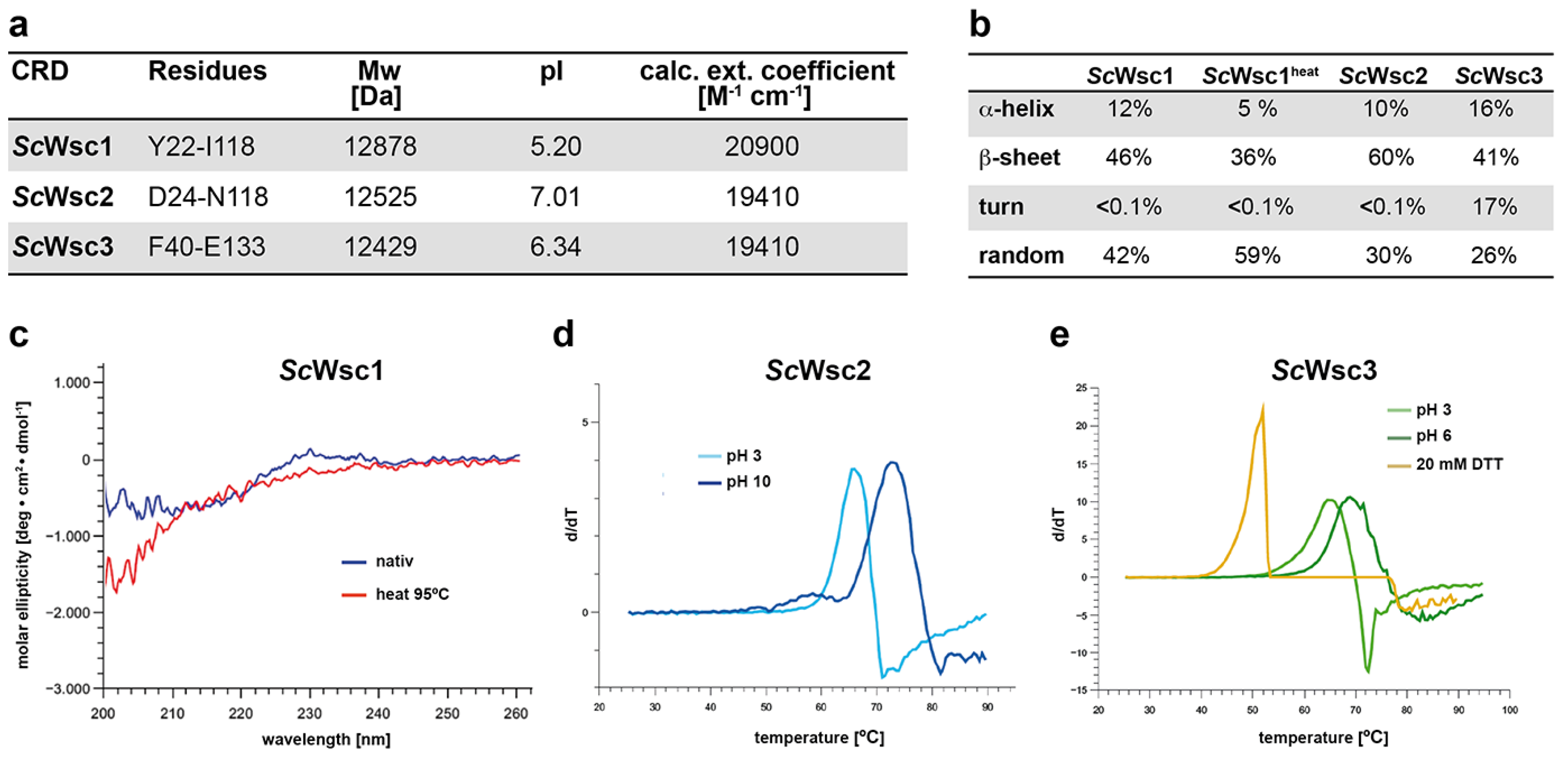
2.4. CD Spectroscopy and Thermal Shift Assays
2.5. Crystallization
2.6. X-ray Data Collection, Structure Solution, and Analysis
2.7. Western-Blot Analysis
2.8. Microscopy
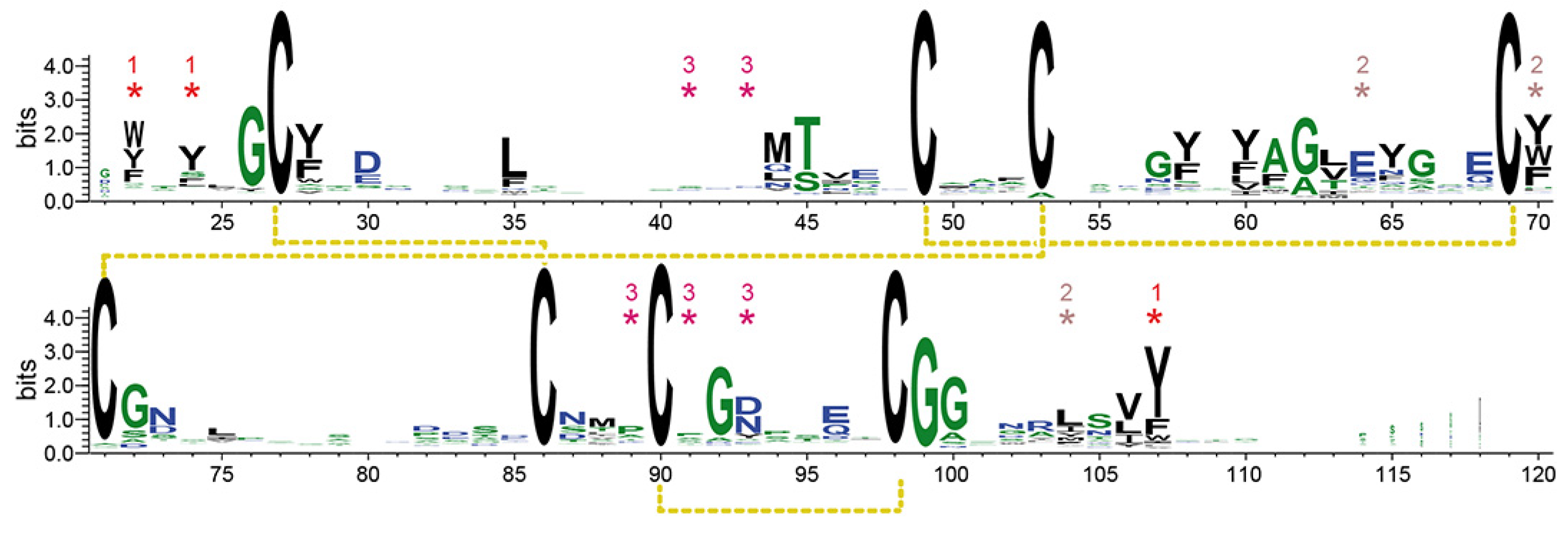
2.9. Bioinformatic Analysis
2.10. Data Availability
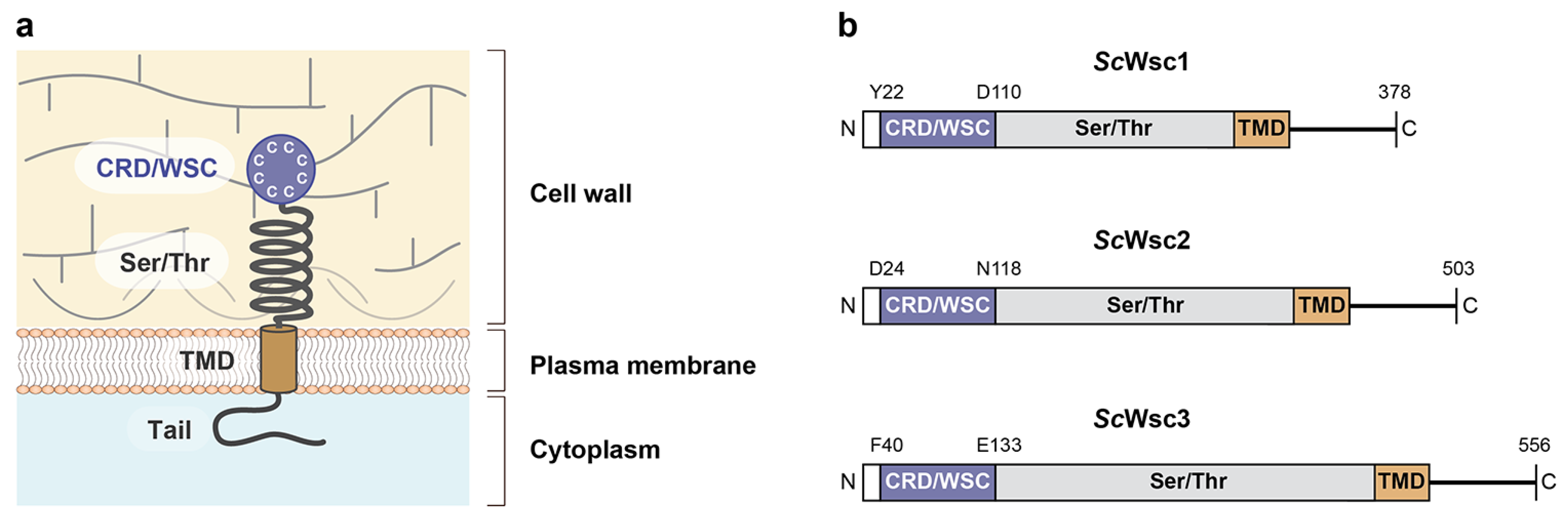
3. Results
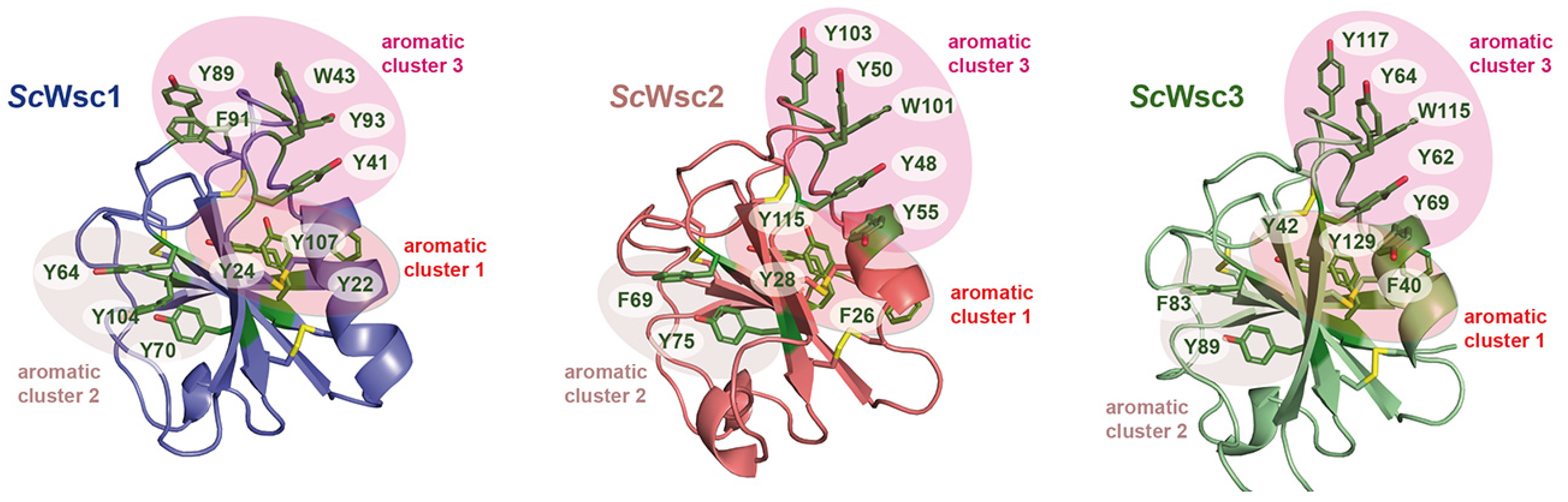
3.1. Characterization of Recombinant CRDs from S. cerevisiae Wsc1, Wsc2, and Wsc3
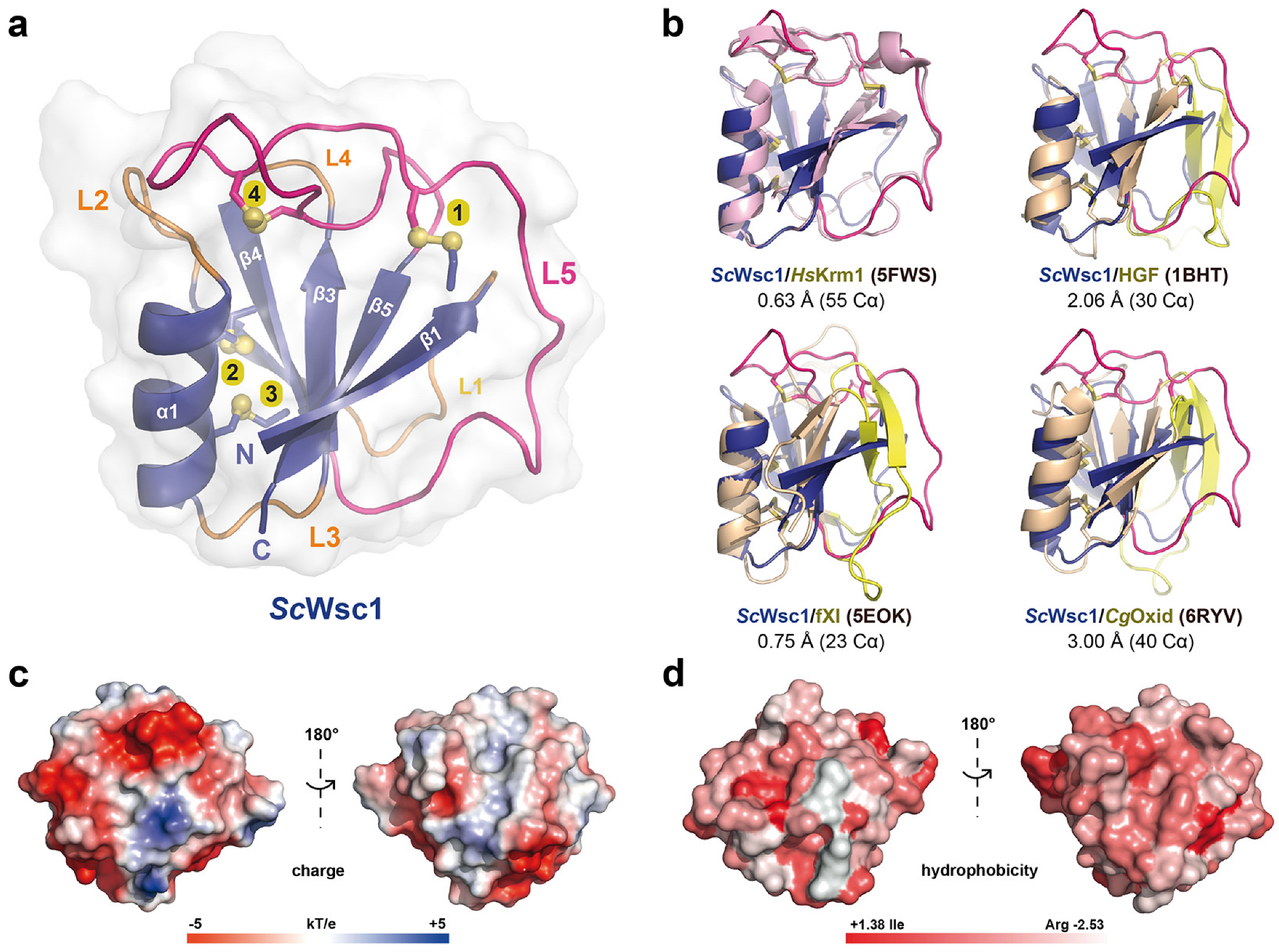
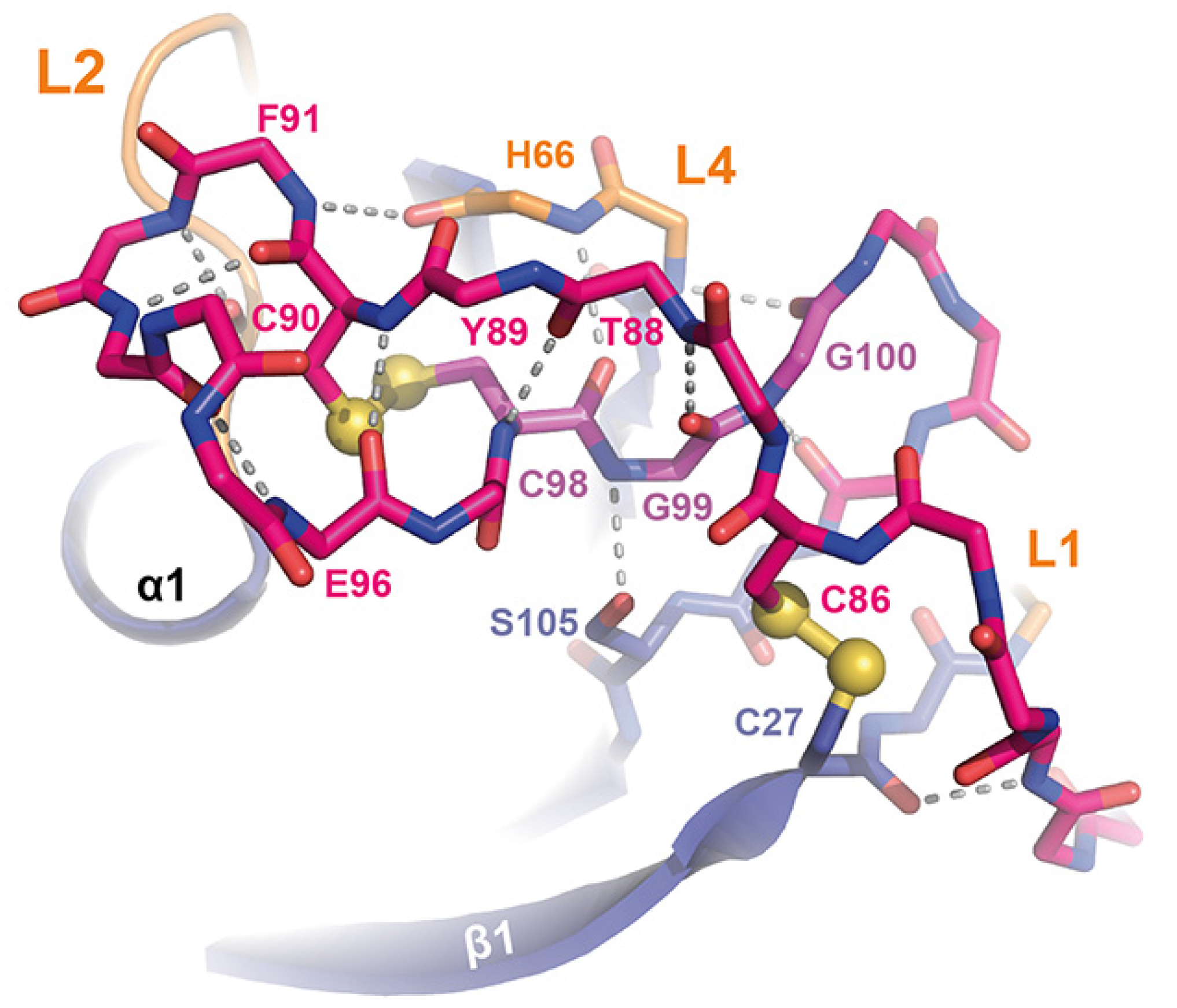
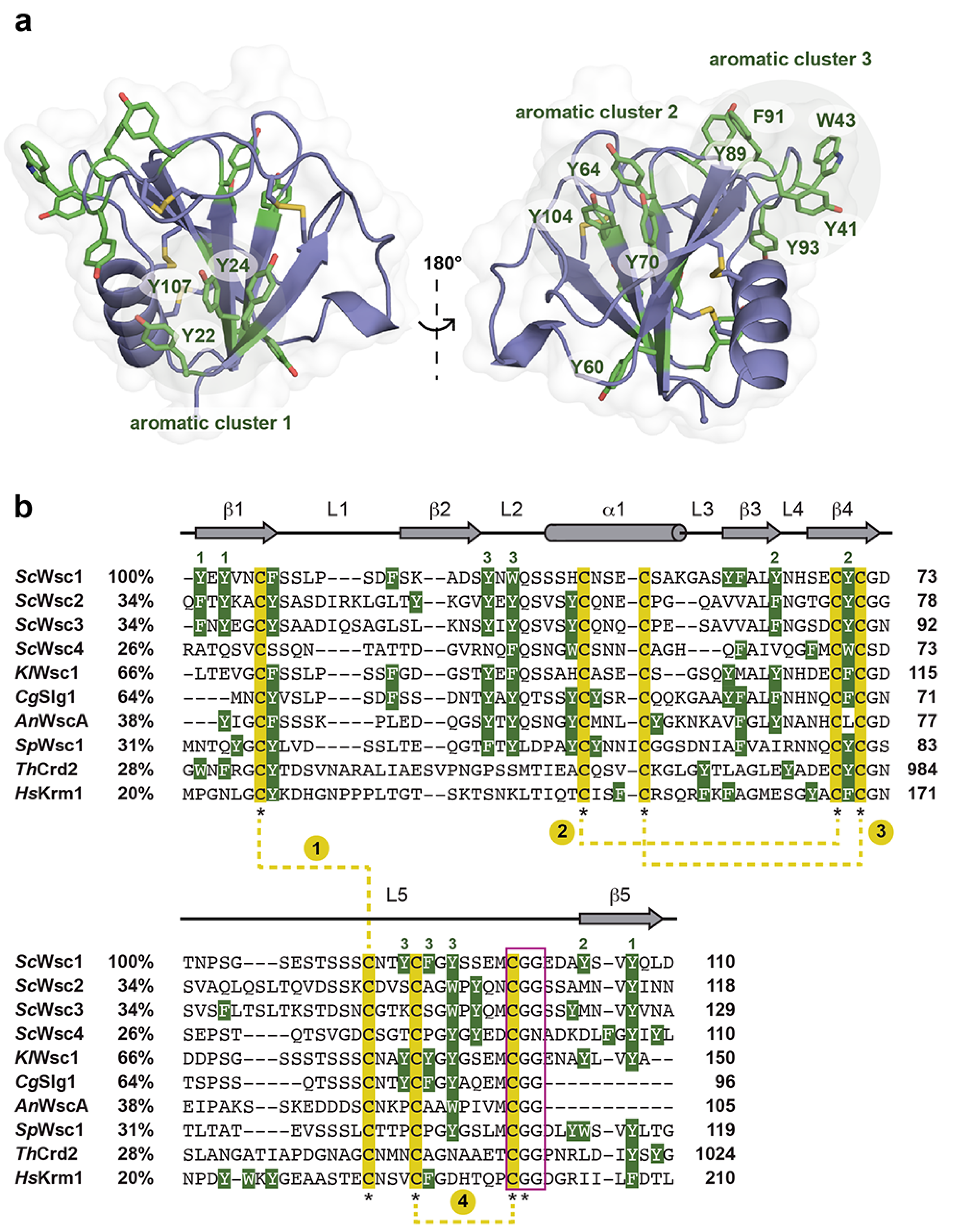
3.2. High-Resolution Structure of the ScWsc1 CRD
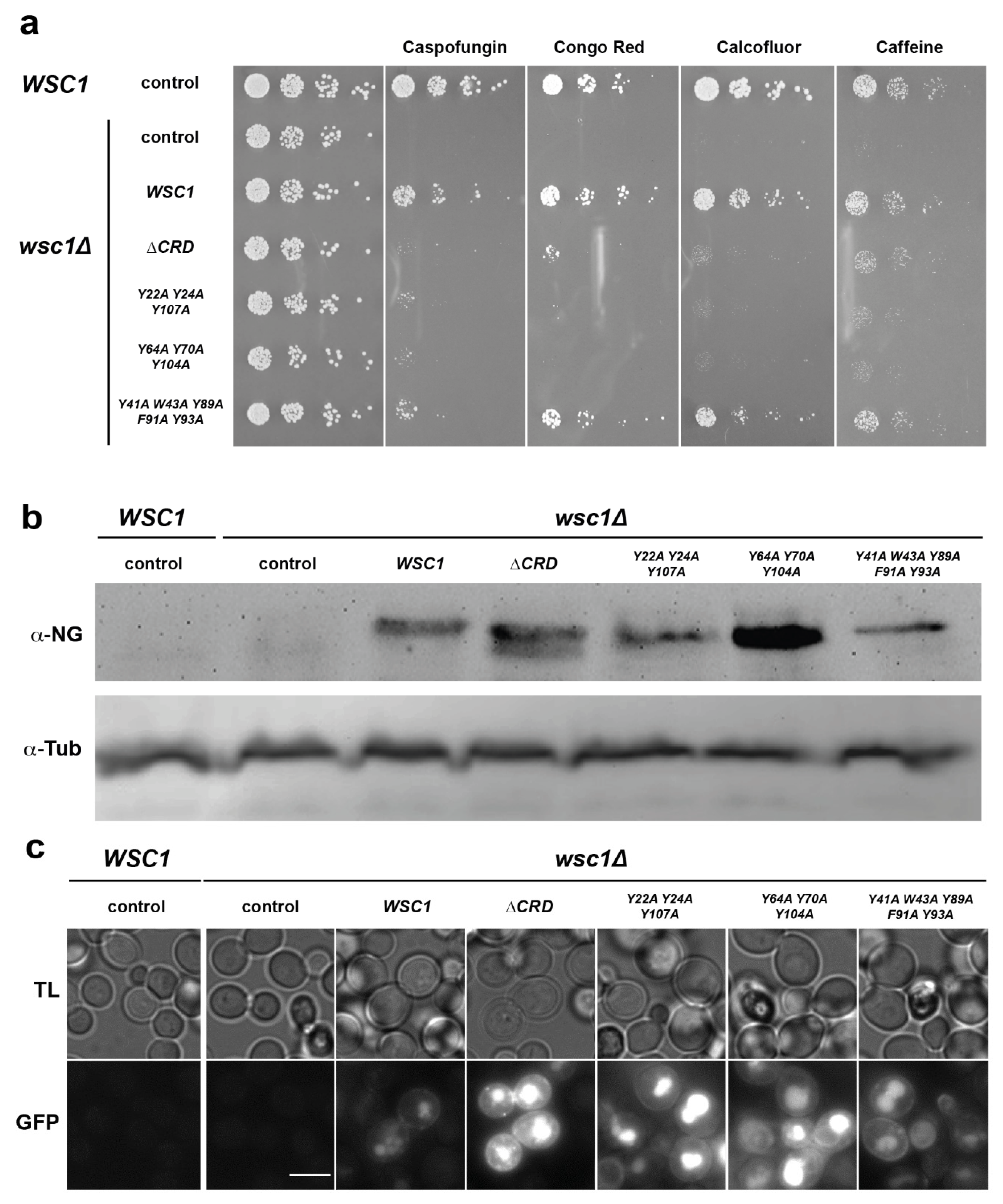
3.3. Functional Characterization of Wsc1 Surface-Exposed Aromatic Clusters
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Garcia-Rubio, R.; de Oliveira, H.C.; Rivera, J.; Trevijano-Contador, N. The Fungal Cell Wall: Candida, Cryptococcus, and Aspergillus Species. Front. Microbiol. 2019, 10, 2993. [Google Scholar] [CrossRef] [PubMed]
- Gow, N.A.R.; Latge, J.P.; Munro, C.A. The Fungal Cell Wall: Structure, Biosynthesis, and Function. Microbiol. Spectr. 2017, 5. [Google Scholar] [CrossRef] [PubMed]
- De Groot, P.W.; Hellingwerf, K.J.; Klis, F.M. Genome-wide identification of fungal GPI proteins. Yeast 2003, 20, 781–796. [Google Scholar] [CrossRef] [PubMed]
- Lesage, G.; Bussey, H. Cell wall assembly in Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 2006, 70, 317–343. [Google Scholar] [CrossRef]
- Orlean, P. Architecture and biosynthesis of the Saccharomyces cerevisiae cell wall. Genetics 2012, 192, 775–818. [Google Scholar] [CrossRef]
- Levin, D.E. Cell wall integrity signaling in Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 2005, 69, 262–291. [Google Scholar] [CrossRef]
- Elorza, M.V.; Rico, H.; Sentandreu, R. Calcofluor white alters the assembly of chitin fibrils in Saccharomyces cerevisiae and Candida albicans cells. J. Gen. Microbiol. 1983, 129, 1577–1582. [Google Scholar] [CrossRef][Green Version]
- Roncero, C.; Duran, A. Effect of Calcofluor white and Congo red on fungal cell wall morphogenesis: In vivo activation of chitin polymerization. J. Bacteriol. 1985, 163, 1180–1185. [Google Scholar] [CrossRef]
- Ketela, T.; Green, R.; Bussey, H. Saccharomyces cerevisiae mid2p is a potential cell wall stress sensor and upstream activator of the PKC1-MPK1 cell integrity pathway. J. Bacteriol. 1999, 181, 3330–3340. [Google Scholar] [CrossRef]
- Garcia, R.; Bermejo, C.; Grau, C.; Perez, R.; Rodriguez-Pena, J.M.; Francois, J.; Nombela, C.; Arroyo, J. The global transcriptional response to transient cell wall damage in Saccharomyces cerevisiae and its regulation by the cell integrity signaling pathway. J. Biol. Chem. 2004, 279, 15183–15195. [Google Scholar] [CrossRef]
- Imai, K.; Noda, Y.; Adachi, H.; Yoda, K. A novel endoplasmic reticulum membrane protein Rcr1 regulates chitin deposition in the cell wall of Saccharomyces cerevisiae. J. Biol. Chem. 2005, 280, 8275–8284. [Google Scholar] [CrossRef] [PubMed]
- Bermejo, C.; Garcia, R.; Straede, A.; Rodriguez-Pena, J.M.; Nombela, C.; Heinisch, J.J.; Arroyo, J. Characterization of sensor-specific stress response by transcriptional profiling of wsc1 and mid2 deletion strains and chimeric sensors in Saccharomyces cerevisiae. Omics J. Integr. Biol. 2010, 14, 679–688. [Google Scholar] [CrossRef] [PubMed]
- Rodicio, R.; Heinisch, J.J. Together we are strong—Cell wall integrity sensors in yeasts. Yeast 2010, 27, 531–540. [Google Scholar] [CrossRef]
- Dichtl, K.; Samantaray, S.; Wagener, J. Cell wall integrity signalling in human pathogenic fungi. Cell Microbiol. 2016, 18, 1228–1238. [Google Scholar] [CrossRef] [PubMed]
- Levin, D.E. Regulation of cell wall biogenesis in Saccharomyces cerevisiae: The cell wall integrity signaling pathway. Genetics 2011, 189, 1145–1175. [Google Scholar] [CrossRef]
- Heinisch, J.J.; Rodicio, R. Protein kinase C in fungi-more than just cell wall integrity. FEMS Microbiol. Rev. 2018, 42, fux051. [Google Scholar] [CrossRef] [PubMed]
- Kock, C.; Dufrene, Y.F.; Heinisch, J.J. Up against the wall: Is yeast cell wall integrity ensured by mechanosensing in plasma membrane microdomains? Appl. Environ. Microbiol. 2015, 81, 806–811. [Google Scholar] [CrossRef] [PubMed]
- Jacoby, J.J.; Nilius, S.M.; Heinisch, J.J. A screen for upstream components of the yeast protein kinase C signal transduction pathway identifies the product of the SLG1 gene. Mol. Gen. Genet. 1998, 258, 148–155. [Google Scholar] [CrossRef]
- Lodder, A.L.; Lee, T.K.; Ballester, R. Characterization of the Wsc1 protein, a putative receptor in the stress response of Saccharomyces cerevisiae. Genetics 1999, 152, 1487–1499. [Google Scholar] [CrossRef]
- Heinisch, J.J.; Dupres, V.; Wilk, S.; Jendretzki, A.; Dufrene, Y.F. Single-molecule atomic force microscopy reveals clustering of the yeast plasma-membrane sensor Wsc1. PLoS ONE 2010, 5, e11104. [Google Scholar] [CrossRef]
- Dupres, V.; Alsteens, D.; Wilk, S.; Hansen, B.; Heinisch, J.J.; Dufrene, Y.F. The yeast Wsc1 cell surface sensor behaves like a nanospring in vivo. Nat. Chem. Biol. 2009, 5, 857–862. [Google Scholar] [CrossRef] [PubMed]
- Kock, C.; Arlt, H.; Ungermann, C.; Heinisch, J.J. Yeast cell wall integrity sensors form specific plasma membrane microdomains important for signalling. Cell Microbiol. 2016, 18, 1251–1267. [Google Scholar] [CrossRef] [PubMed]
- Voskoboynikova, N.; Karlova, M.; Kurre, R.; Mulkidjanian, A.Y.; Shaitan, K.V.; Sokolova, O.S.; Steinhoff, H.J.; Heinisch, J.J. A Three-Dimensional Model of the Yeast Transmembrane Sensor Wsc1 Obtained by SMA-Based Detergent-Free Purification and Transmission Electron Microscopy. J. Fungi 2021, 7, 118. [Google Scholar] [CrossRef] [PubMed]
- Rodicio, R.; Buchwald, U.; Schmitz, H.P.; Heinisch, J.J. Dissecting sensor functions in cell wall integrity signaling in Kluyveromyces lactis. Fungal Genet. Biol. 2008, 45, 422–435. [Google Scholar] [CrossRef] [PubMed]
- Futagami, T.; Nakao, S.; Kido, Y.; Oka, T.; Kajiwara, Y.; Takashita, H.; Omori, T.; Furukawa, K.; Goto, M. Putative stress sensors WscA and WscB are involved in hypo-osmotic and acidic pH stress tolerance in Aspergillus nidulans. Eukaryot. Cell 2011, 10, 1504–1515. [Google Scholar] [CrossRef]
- Maddi, A.; Dettman, A.; Fu, C.; Seiler, S.; Free, S.J. WSC-1 and HAM-7 are MAK-1 MAP kinase pathway sensors required for cell wall integrity and hyphal fusion in Neurospora crassa. PLoS ONE 2012, 7, e42374. [Google Scholar] [CrossRef]
- Tong, S.M.; Chen, Y.; Zhu, J.; Ying, S.H.; Feng, M.G. Subcellular localization of five singular WSC domain-containing proteins and their roles in Beauveria bassiana responses to stress cues and metal ions. Environ. Microbiol. Rep. 2016, 8, 295–304. [Google Scholar] [CrossRef]
- Cohen-Kupiec, R.; Broglie, K.E.; Friesem, D.; Broglie, R.M.; Chet, I. Molecular characterization of a novel beta-1,3-exoglucanase related to mycoparasitism of Trichoderma harzianum. Gene 1999, 226, 147–154. [Google Scholar] [CrossRef]
- Wawra, S.; Fesel, P.; Widmer, H.; Neumann, U.; Lahrmann, U.; Becker, S.; Hehemann, J.H.; Langen, G.; Zuccaro, A. FGB1 and WSC3 are in planta-induced beta-glucan-binding fungal lectins with different functions. New Phytol. 2019, 222, 1493–1506. [Google Scholar] [CrossRef]
- Mao, B.; Wu, W.; Davidson, G.; Marhold, J.; Li, M.; Mechler, B.M.; Delius, H.; Hoppe, D.; Stannek, P.; Walter, C.; et al. Kremen proteins are Dickkopf receptors that regulate Wnt/beta-catenin signalling. Nature 2002, 417, 664–667. [Google Scholar] [CrossRef]
- Nakamura, T.; Nakamura, T.; Matsumoto, K. The functions and possible significance of Kremen as the gatekeeper of Wnt signalling in development and pathology. J. Cell Mol. Med. 2008, 12, 391–408. [Google Scholar] [CrossRef] [PubMed]
- Zebisch, M.; Jackson, V.A.; Zhao, Y.; Jones, E.Y. Structure of the Dual-Mode Wnt Regulator Kremen1 and Insight into Ternary Complex Formation with LRP6 and Dickkopf. Structure 2016, 24, 1599–1605. [Google Scholar] [CrossRef] [PubMed]
- Pereira, G.; Tanaka, T.U.; Nasmyth, K.; Schiebel, E. Modes of spindle pole body inheritance and segregation of the Bfa1p-Bub2p checkpoint protein complex. EMBO J. 2001, 20, 6359–6370. [Google Scholar] [CrossRef] [PubMed]
- Janke, C.; Magiera, M.M.; Rathfelder, N.; Taxis, C.; Reber, S.; Maekawa, H.; Moreno-Borchart, A.; Doenges, G.; Schwob, E.; Schiebel, E.; et al. A versatile toolbox for PCR-based tagging of yeast genes: New fluorescent proteins, more markers and promoter substitution cassettes. Yeast 2004, 21, 947–962. [Google Scholar] [CrossRef] [PubMed]
- Guthrie, C.; Fink, G.R. (Eds.) Guide to Yeast Genetics and Molecular Biology; Academic Press: San Diego, CA, USA, 1991; Volume 194, pp. 1–863. [Google Scholar]
- Lutz, A.P.; Schladebeck, S.; Renicke, C.; Spadaccini, R.; Mosch, H.U.; Taxis, C. Proteasome Activity Is Influenced by the HECT_2 Protein Ipa1 in Budding Yeast. Genetics 2018, 209, 157–171. [Google Scholar] [CrossRef] [PubMed]
- Sikorski, R.S.; Hieter, P. A system of shuttle vectors and yeast host strains designed for efficient manipulation of DNA in Saccharomyces cerevisiae. Genetics 1989, 122, 19–27. [Google Scholar] [CrossRef]
- Veelders, M.; Brückner, S.; Ott, D.; Unverzagt, C.; Mösch, H.U.; Essen, L.O. Structural basis of flocculin-mediated social behavior in yeast. Proc. Natl. Acad. Sci. USA 2010, 107, 22511–22516. [Google Scholar] [CrossRef]
- Kabsch, W. Xds. Acta Crystallogr. 2010, 66, 125–132. [Google Scholar] [CrossRef]
- Kabsch, W. Integration, scaling, space-group assignment and post-refinement. Acta Crystallogr. 2010, 66, 133–144. [Google Scholar] [CrossRef]
- Evans, P.R. An introduction to data reduction: Space-group determination, scaling and intensity statistics. Acta Crystallogr. 2011, 67, 282–292. [Google Scholar] [CrossRef]
- Winn, M.D.; Ballard, C.C.; Cowtan, K.D.; Dodson, E.J.; Emsley, P.; Evans, P.R.; Keegan, R.M.; Krissinel, E.B.; Leslie, A.G.; McCoy, A.; et al. Overview of the CCP4 suite and current developments. Acta Crystallogr. 2011, 67, 235–242. [Google Scholar] [CrossRef]
- McCoy, A.J.; Grosse-Kunstleve, R.W.; Adams, P.D.; Winn, M.D.; Storoni, L.C.; Read, R.J. Phaser crystallographic software. J. Appl. Crystallogr. 2007, 40, 658–674. [Google Scholar] [CrossRef] [PubMed]
- Adams, P.D.; Afonine, P.V.; Bunkoczi, G.; Chen, V.B.; Davis, I.W.; Echols, N.; Headd, J.J.; Hung, L.W.; Kapral, G.J.; Grosse-Kunstleve, R.W.; et al. PHENIX: A comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr. 2010, 66, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Emsley, P.; Lohkamp, B.; Scott, W.G.; Cowtan, K. Features and development of Coot. Acta Crystallogr. 2010, 66, 486–501. [Google Scholar] [CrossRef]
- Yaffe, M.P.; Schatz, G. Two nuclear mutations that block mitochondrial protein import in yeast. Proc. Natl. Acad. Sci. USA 1984, 81, 4819–4823. [Google Scholar] [CrossRef]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef]
- Dupres, V.; Heinisch, J.J.; Dufrene, Y.F. Atomic force microscopy demonstrates that disulfide bridges are required for clustering of the yeast cell wall integrity sensor Wsc1. Langmuir 2011, 27, 15129–15134. [Google Scholar] [CrossRef]
- Ultsch, M.; Lokker, N.A.; Godowski, P.J.; de Vos, A.M. Crystal structure of the NK1 fragment of human hepatocyte growth factor at 2.0 A resolution. Structure 1998, 6, 1383–1393. [Google Scholar] [CrossRef]
- Wong, S.S.; Ostergaard, S.; Hall, G.; Li, C.; Williams, P.M.; Stennicke, H.; Emsley, J. A novel DFP tripeptide motif interacts with the coagulation factor XI apple 2 domain. Blood 2016, 127, 2915–2923. [Google Scholar] [CrossRef]
- Mathieu, Y.; Offen, W.A.; Forget, S.M.; Ciano, L.; Viborg, A.H.; Blagova, E.; Henrissat, B.; Walton, P.H.; Davies, G.J.; Brumer, H. Discovery of a Fungal Copper Radical Oxidase with High Catalytic Efficiency toward 5-Hydroxymethylfurfural and Benzyl Alcohols for Bioprocessing. ACS Catal. 2020, 10, 3042–3058. [Google Scholar] [CrossRef]
- Crooks, G.E.; Hon, G.; Chandonia, J.M.; Brenner, S.E. WebLogo: A sequence logo generator. Genome Res. 2004, 14, 1188–1190. [Google Scholar] [CrossRef] [PubMed]
- Ovchinnikov, S.; Kamisetty, H.; Baker, D. Robust and accurate prediction of residue-residue interactions across protein interfaces using evolutionary information. eLife 2014, 3, e02030. [Google Scholar] [CrossRef] [PubMed]
- Kraushaar, T.; Brückner, S.; Veelders, M.; Rhinow, D.; Schreiner, F.; Birke, R.; Pagenstecher, A.; Mösch, H.-U.; Essen, L.-O. Interactions by the Fungal Flo11 Adhesin Depend on a Fibronectin Type III-like Adhesin Domain Girdled by Aromatic Bands. Structure 2015, 23, 1005–1017. [Google Scholar] [CrossRef] [PubMed]
- Reithofer, V.; Fernandez-Pereira, J.; Alvarado, M.; de Groot, P.; Essen, L.O. A novel class of Candida glabrata cell wall proteins with beta-helix fold mediates adhesion in clinical isolates. PLoS Pathog. 2021, 17, e1009980. [Google Scholar] [CrossRef] [PubMed]
- Lanzarotti, E.; Biekofsky, R.R.; Estrin, D.A.; Marti, M.A.; Turjanski, A.G. Aromatic-aromatic interactions in proteins: Beyond the dimer. J. Chem. Inf. Model 2011, 51, 1623–1633. [Google Scholar] [CrossRef] [PubMed]
- Lanzarotti, E.; Defelipe, L.A.; Marti, M.A.; Turjanski, A.G. Aromatic clusters in protein-protein and protein-drug complexes. J. Cheminform. 2020, 12, 30. [Google Scholar] [CrossRef] [PubMed]
- Brückner, S.; Schubert, R.; Kraushaar, T.; Hartmann, R.; Hoffmann, D.; Jelli, E.; Drescher, K.; Müller, D.J.; Oliver Essen, L.O.; Mösch, H.U. Kin discrimination in social yeast is mediated by cell surface receptors of the Flo11 adhesin family. eLife 2020, 9, e55587. [Google Scholar] [CrossRef]
- Maestre-Reyna, M.; Diderrich, R.; Veelders, M.S.; Eulenburg, G.; Kalugin, V.; Brückner, S.; Keller, P.; Rupp, S.; Mösch, H.U.; Essen, L.O. Structural basis for promiscuity and specificity during Candida glabrata invasion of host epithelia. Proc. Natl. Acad. Sci. USA 2012, 109, 16864–16869. [Google Scholar] [CrossRef]
- Diderrich, R.; Kock, M.; Maestre-Reyna, M.; Keller, P.; Steuber, H.; Rupp, S.; Essen, L.O.; Mösch, H.U. Structural Hot Spots Determine Functional Diversity of the Candida glabrata Epithelial Adhesin Family. J. Biol. Chem. 2015, 290, 19597–19613. [Google Scholar] [CrossRef]
- Kock, M.; Brückner, S.; Wozniak, N.; Maestre-Reyna, M.; Veelders, M.; Schlereth, J.; Mösch, H.U.; Essen, L.O. Structural and Functional Characterization of PA14/Flo5-Like Adhesins From Komagataella pastoris. Front. Microbiol. 2018, 9, 2581. [Google Scholar] [CrossRef]
- Hoffmann, D.; Diderrich, R.; Reithofer, V.; Friederichs, S.; Kock, M.; Essen, L.O.; Mösch, H.U. Functional reprogramming of Candida glabrata epithelial adhesins: The role of conserved and variable structural motifs in ligand binding. J. Biol. Chem. 2020, 295, 12512–12524. [Google Scholar] [CrossRef] [PubMed]
- Essen, L.O.; Vogt, M.S.; Mösch, H.U. Diversity of GPI-anchored fungal adhesins. Biol. Chem. 2020, 401, 1389–1405. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plasmid | Relevant Genotype | Source |
|---|---|---|
| pET-28(a)+ | PT7-6xHis lacI KanR | Merck, Germany |
| BHUM3120 | WSC1(22−118) in pET-28(a)+ | this study |
| BHUM3121 | WSC2(24−118) in pET-28(a)+ | this study |
| BHUM3122 | WSC3(40−133) in pET-28(a)+ | this study |
| pFA6a-natNT2 | PFA6a-NatNT2 AmpR | [34] |
| pCR95 | mNeonGreen::NatNT2 AmpR | [36] |
| pRS314 | TRP1 ARS CEN2 AmpR | [37] |
| BHUM3291 | WSC1 in pRS314 | this study |
| BHUM3293 | WSC1Y22A Y24A Y107A in pRS314 | this study |
| BHUM3295 | WSC1Y64A Y70A Y104A in pRS314 | this study |
| BHUM3297 | WSC1Y41A W43A Y89A F91A Y93A in pRS314 | this study |
| BHUM3301 | WSC1∆CRD in pRS314 | this study |
| BHUM3303 | WSC1-mNeonGreen in pRS314 | this study |
| BHUM3304 | WSC1Y22A Y24A Y107A-mNeonGreen in pRS314 | this study |
| BHUM3305 | WSC1Y64A Y70A Y104A-mNeonGreen in pRS314 | this study |
| BHUM3306 | WSC1Y41A W43A Y89A F91A Y93A-mNeonGreen in pRS314 | this study |
| BHUM3308 | WSC1∆CRD-mNeonGreen in pRS314 | this study |
| PDB Code | 7PZ2 |
|---|---|
| X-ray source | ESRF ID29 |
| Wavelength (Å) | 0.979 |
| Resolution range (Å) 1 | 26.75–1.58 (1.64–1.58) |
| Space group | P 1 21 1 |
| Unit cell | a = 31.82 Å, b = 53.5 Å, c = 51.93 Å α = 90°, β = 95.46°, γ = 90° |
| Total reflections 1 | 70,058 (6136) |
| Unique reflections 1 | 23,125 (2133) |
| Multiplicity 1 | 3.0 (2.8) |
| Completeness (%) 1 | 97.06 (91.42) |
| Mean I/sigma(I) 1 | 10.35 (2.17) |
| Wilson B-factor (Å2) | 16.72 |
| Rmerge 1 | 0.06339 (0.4653) |
| CC1/2 1 | 0.997 (0.756) |
| CC* 1 | 0.999 (0.928) |
| Reflections used in refinement 1 | 23,052 (2132) |
| Reflections used for Rfree 1 | 1113 (126) |
| Rwork 1 | 0.150 (0.258) |
| Rfree 1 | 0.184 (0.251) |
| CC (work, free) 1 | 0.973 (0.819), 0.965 (0.824) |
| Number of non-hydrogen atoms | 1731 |
| Macromolecules | 1527 |
| Ligands, solvent | 7197 |
| Protein residues | 197 |
| r.m.s.d. bonds (Å) | 0.005 |
| r.m.s.d. angles (°) | 0.99 |
| Ramachandran favored (%) | 92.23 |
| Ramachandran allowed (%) | 7.77 |
| Ramachandran outliers (%) | 0.00 |
| Rotamer outliers (%) | 0.56 |
| Clashscore 2 | 1.06 |
| Average B-factor (Å2) | 20.88 |
| Macromolecules (Å2) | 19.53 |
| Ligands, Solvent (Å2) | 25.84, 31.15 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schöppner, P.; Lutz, A.P.; Lutterbach, B.J.; Brückner, S.; Essen, L.-O.; Mösch, H.-U. Structure of the Yeast Cell Wall Integrity Sensor Wsc1 Reveals an Essential Role of Surface-Exposed Aromatic Clusters. J. Fungi 2022, 8, 379. https://doi.org/10.3390/jof8040379
Schöppner P, Lutz AP, Lutterbach BJ, Brückner S, Essen L-O, Mösch H-U. Structure of the Yeast Cell Wall Integrity Sensor Wsc1 Reveals an Essential Role of Surface-Exposed Aromatic Clusters. Journal of Fungi. 2022; 8(4):379. https://doi.org/10.3390/jof8040379
Chicago/Turabian StyleSchöppner, Philipp, Anne Pia Lutz, Bernard Johannes Lutterbach, Stefan Brückner, Lars-Oliver Essen, and Hans-Ulrich Mösch. 2022. "Structure of the Yeast Cell Wall Integrity Sensor Wsc1 Reveals an Essential Role of Surface-Exposed Aromatic Clusters" Journal of Fungi 8, no. 4: 379. https://doi.org/10.3390/jof8040379
APA StyleSchöppner, P., Lutz, A. P., Lutterbach, B. J., Brückner, S., Essen, L.-O., & Mösch, H.-U. (2022). Structure of the Yeast Cell Wall Integrity Sensor Wsc1 Reveals an Essential Role of Surface-Exposed Aromatic Clusters. Journal of Fungi, 8(4), 379. https://doi.org/10.3390/jof8040379