
Fusarium Species Associated with Maize Leaf Blight in Heilongjiang Province, China

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Fusarium Isolates Collection
2.2. Morphological Characterization
2.3. DNA Extraction and Sequence Analysis
2.4. Phylogenetic Relationships among Fusarium Isolates
2.5. Pathogenicity Tests
2.6. DNA Polymorphism
2.7. Haplotype Analysis
3. Results
3.1. Fungal Isolation and Morphological Characterization
3.2. Phylogenetic Analysis
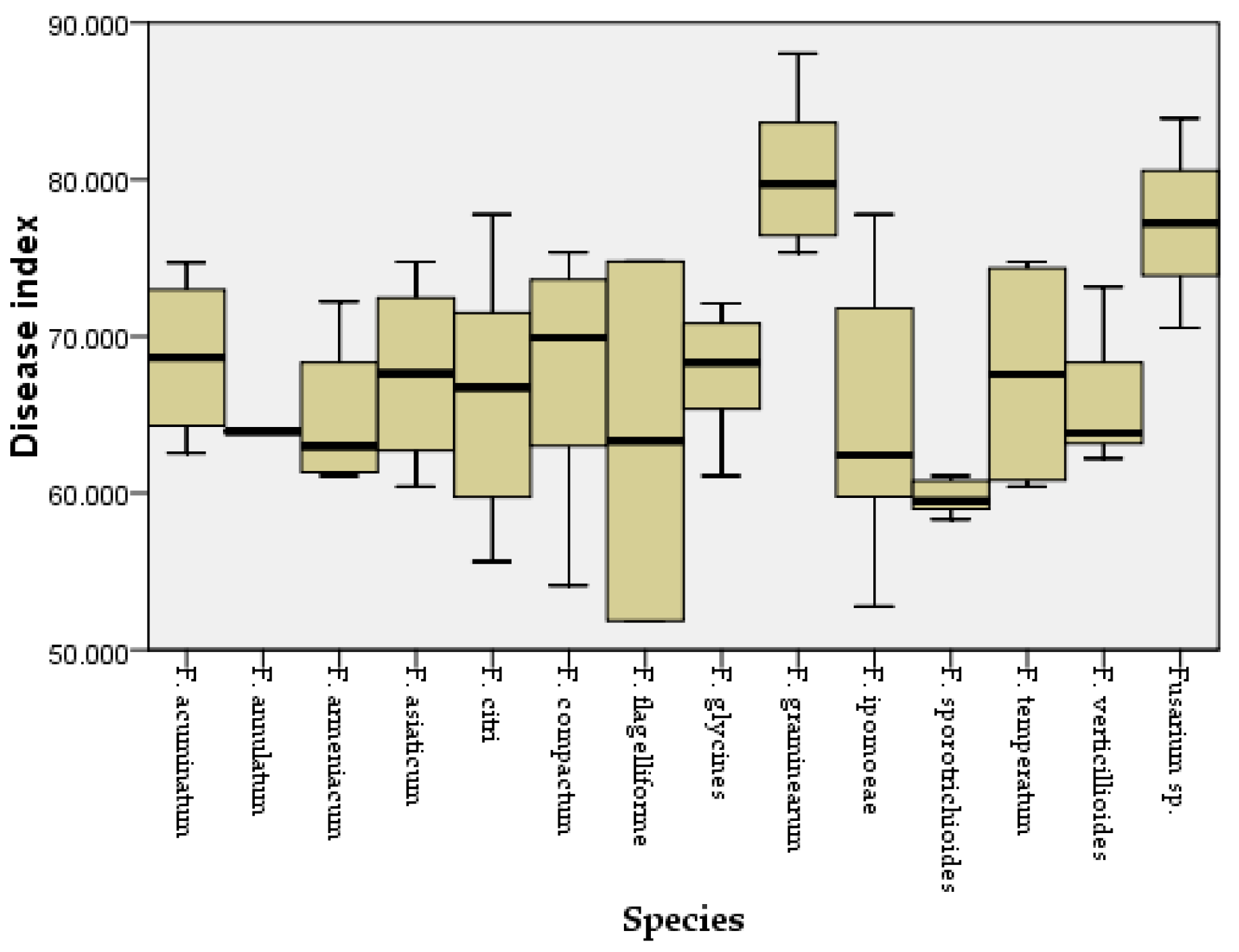
3.3. Pathogenicity Tests
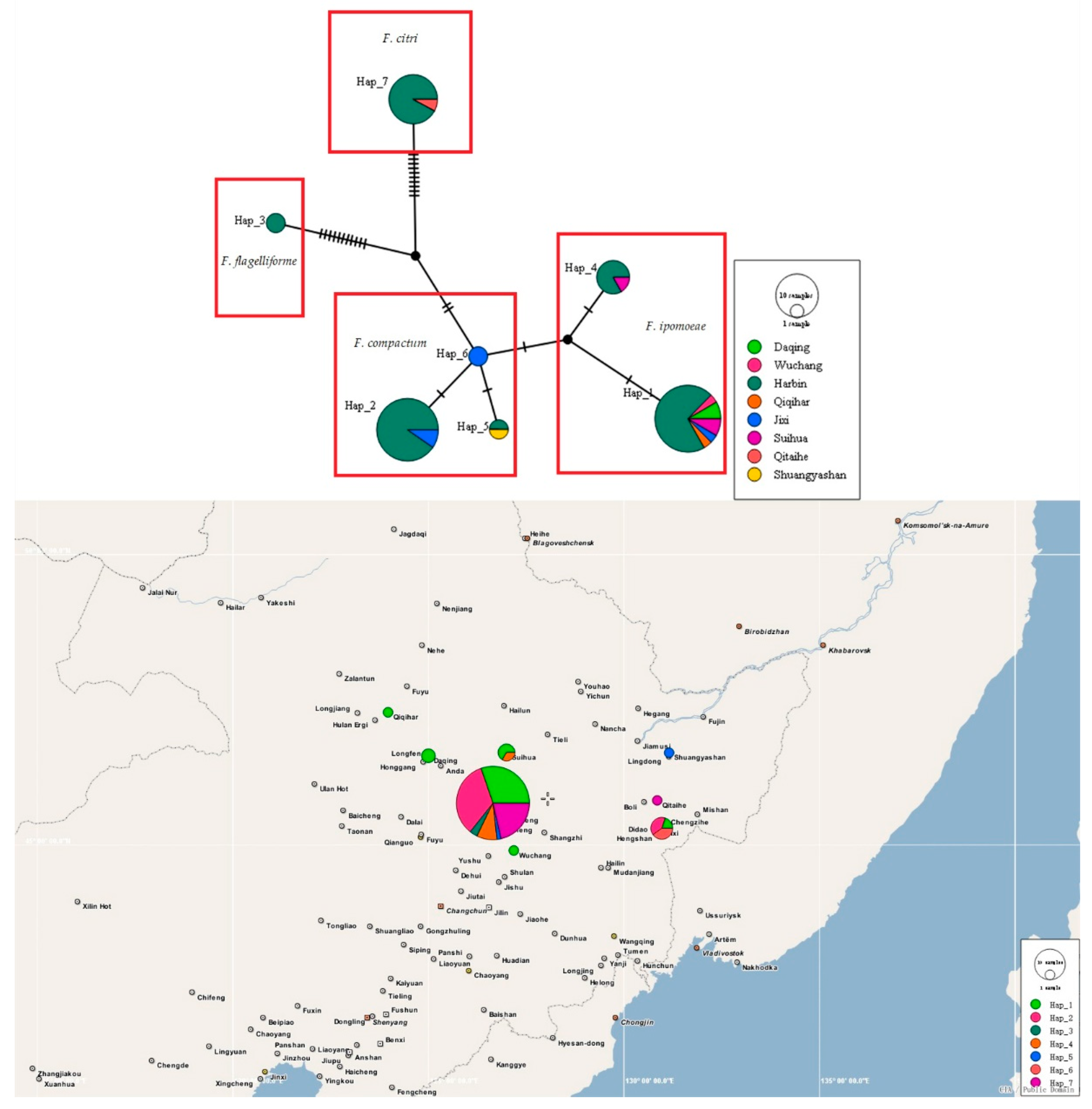
3.4. Haplotype Analyses and DNA Polymorphism
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Munkvold, G.P. Epidemiology of Fusarium diseases and their mycotoxins in maize ears. Eur. J. Plant Pathol. 2003, 109, 705–713. [Google Scholar] [CrossRef]
- Leslie, J.; Summerell, B.A. The Fusarium Laboratory Manual; Blackwell Publishing: Oxford, UK, 2006. [Google Scholar]
- Machado, F.J.; de Barros, A.V.; McMaster, N.; Schmale, D.G., 3rd; Vaillancourt, L.J.; Del Ponte, E.M. Aggressiveness and mycotoxin production by Fusarium meridionale compared with F. graminearum on maize ears and stalks in the field. Phytopathology 2022, 112, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Gai, X.T.; Yang, R.X.; Pan, X.J.; Yuan, Y.; Wang, S.N.; Liang, B.B.; Gao, Z.G. First report of Fusarium incarnatum causing stalk rot on maize in China. Plant Dis. 2016, 100, 1010. [Google Scholar] [CrossRef]
- Dong, H.; Qin, P.; Gao, Z.; Xu, J.; Xu, X. First report of seedling blight of maize caused by Fusarium asiaticum in Northeast China. Plant Dis. 2021, 105, 1206. [Google Scholar] [CrossRef]
- Schieber, R. A leaf blight of corn (Zea mays) incited by Fusarium moniliforme. Phytopathology 1968, 58, 554. [Google Scholar]
- Jacobs-Venter, A.; Laraba, I.; Geiser, D.M.; Busman, M.; Vaughan, M.M.; Proctor, R.H.; McCormick, S.P.; O’Donnell, K. Molecular systematics of two sister clades, the Fusarium concolor and F. babinda species complexes, and the discovery of a novel microcycle macroconidium-producing species from South Africa. Mycologia 2018, 110, 1189–1204. [Google Scholar] [CrossRef]
- Dean, R.; van Kan, J.A.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef]
- Ramdial, H.; Latchoo, R.K.; Hosein, F.N.; Rampersad, S.N. Phylogeny and haplotype analysis of fungi within the Fusarium incarnatum-equiseti species complex. Phytopathology 2017, 107, 109–120. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Academic Press: London, UK, 1990; pp. 315–322. [Google Scholar]
- O’Donnell, K.; Ward, T.J.; Robert, V.; Crous, P.W.; Geiser, D.M.; Kang, S. DNA sequence-based identification of Fusarium: Current status and future directions. Phytoparasitica 2015, 43, 583–595. [Google Scholar] [CrossRef]
- Geiser, D.M.; Jiménez-Gasco, M.D.M.; Kang, S.; Makalowska, I.; Veeraraghavan, N.; Ward, T.J.; Zhang, N.; Kuldau, G.A.; O’Donnell, K. Fusarium-ID v. 1.0: A DNA sequence database for identifying Fusarium. Eur. J. Plant Pathol. 2004, 110, 473–479. [Google Scholar] [CrossRef]
- O’Donnell, K.; Whitaker, B.K.; Laraba, I.; Proctor, R.H.; Brown, D.W.; Broders, K.; Kim, H.S.; McCormick, S.P.; Busman, M.; Aoki, T.; et al. DNA Sequence-Based Identification of Fusarium: A Work in Progress. Plant Dis. 2022, 106, 1597–1609. [Google Scholar] [CrossRef]
- Villani, A.; Moretti, A.; Saeger, S.D.; Han, Z.; Mavungu, J.; Soares, C.; Proctor, R.; Venâncio, A.; Limac, N.; Gaetano, S.; et al. A polyphasic approach for characterization of a collection of cereal isolates of the Fusarium incarnatum-equiseti species complex. Int. J. Food Microbiol. 2016, 234, 24–35. [Google Scholar] [CrossRef]
- O’Donnell, K.; Sutton, D.A.; Rinaldi, M.G.; Gueidan, C.; Crous, P.W.; Geiser, D.M. Novel multilocus sequence typing scheme reveals high genetic diversity of human pathogenic members of the Fusarium incarnatum-equiseti and F. chalmydosporum species complexes within the United States. J. Clin. Microbiol. 2009, 47, 3851–3861. [Google Scholar] [CrossRef]
- O’Donnell, K.; Humber, R.A.; Geiser, D.M.; Kang, S.; Park, B.; Robert, V.A.R.G.; Crous, P.W.; Johnston, P.R.; Aoki, T.; Rooney, A.P.; et al. Phylogenetic diversity of insecticolous Fusariam inferred from multilocus DNA sequence data and their molecular identification via FUSARIUM-ID and Fusarium MLST. Mycologia 2012, 104, 427–445. [Google Scholar] [CrossRef] [PubMed]
- Huson, D.H.; Bryant, D. Application of phylogenetic networks in evolutionary studies. Mol. Biol. Evol. 2006, 23, 254–267. [Google Scholar] [CrossRef] [PubMed]
- Blagojević, J.; Vukojevi, J.; Ivanovi, B.; Ivanovi, A. Characterization of Alternaria species associated with leaf spot disease of Armoracia rusticana in Serbia. Plant Dis. 2010, 104, 1378–1389. [Google Scholar] [CrossRef]
- Leigh, J.W.; Bryant, D. PopART: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef]
- Crous, P.; Lombard, L.; Sandoval-Denis, M.; Seifert, K.; Schroers, H.-J.; Chaverri, P.; Gené, J.; Guarro, J.; Hirooka, Y.; Bensch, K.; et al. Fusarium: More than a node or a foot-shaped basal cell. Stud. Mycol. 2021, 98, 100116. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Raza, M.; Zhang, Z.-F.; Hyde, K.D.; Diao, Y.-Z.; Cai, L. Culturable plant pathogenic fungi associated with sugarcane in southern China. Fungal Divers. 2019, 99, 1–104. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, L.; Yang, X.; Cao, H.; Li, J.; Cao, P.; Guo, L.; Wang, X.; Zhao, J.; Xiang, W. Alternaria spp. associated with leaf blight of maize in Heilongjiang Province, China. Plant Dis. 2022, 106, 572–584. [Google Scholar] [CrossRef] [PubMed]
- Vieira, R.A.; Mesquini, R.M.; Silva, C.N.; Hata, F.T.; Tessmann, D.J.; Scapim, C.A. A new diagrammatic scale for the assessment of northern corn leaf blight. Crop Prot. 2014, 56, 55–57. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA sequence polymorphism analysis of large datasets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Tajima, F. Statistical method for testing the neutral mutation hypothesis by DNA polymorphism. Genetics 1989, 123, 585–595. [Google Scholar] [CrossRef]
- Fu, Y.X.; Li, W.H. Statistical tests of neutrality of mutations. Genetics 1993, 133, 693–709. [Google Scholar] [CrossRef]
- Templeton, A.R.; Crandall, K.A.; Sing, C.F. A cladistics analysis of phenotypic associations with haplotypes inferred from restriction endo nuclease mapping and DNA sequence data III. Cladogram estimation. Genetics 1992, 132, 619–633. [Google Scholar] [CrossRef]
- Clement, M.; Snell, Q.; Walke, P.; Posada, D.; Crandall, K.A. TCS: Estimating gene genealogies. In Proceedings of the 16th International Parallel and Distributed Processing Symposium (IPDPS 2002), Fort Lauderdale, FL, USA, 15–19 April 2002. [Google Scholar]
- Summerell, B.A.; Salleh, B.; Leslie, J.F. A utilitarian approach to Fusarium identification. Plant Dis. 2003, 87, 117–128. [Google Scholar] [CrossRef]
- Jurado, M.; Vázquez, C.; Patiño, B.; González-Jaén, M.T. PCR detection assays for the trichothecene-producing species Fusarium graminearum, Fusarium culmorum, Fusarium poae, Fusarium equiseti and Fusarium sporotrichioides. Syst. Appl. Microbiol. 2005, 28, 562–568. [Google Scholar] [CrossRef]
- Wang, M.M.; Chen, Q.; Diao, Y.Z.; Duan, W.J.; Cai, L. Fusarium incarnatum-equiseti complex from China. Pers. Mol. Phylogeny Evol. Fungi 2019, 43, 70–89. [Google Scholar] [CrossRef] [PubMed]
- Marín, P.; Moretti, A.; Ritieni, A.; Jurado, M.; Vázquez, C.; González-Jaén, M.T. Phylogenetic analyses and toxigenic profiles of Fusarium equiseti and Fusarium acuminatum isolated from cereals from southern Europe. Food Microbiol. 2012, 31, 229–237. [Google Scholar] [CrossRef]
- Barros, G.; Zanon, M.S.; Palazzini, J.M.; Haidukowski, M.; Pascale, M.; Chulze, S. Trichothecenes and zearalenone production by Fusarium equiseti and Fusarium semitectum species isolated from Argentinean soybean. Food Addit. Contam.-Chem. Anal. Control Expo. Risk Assess. 2012, 29, 1436–1442. [Google Scholar] [CrossRef] [PubMed]
- Avila, C.F.; Moreira, G.M.; Nicolli, C.P.; Gomes, L.B.; Abreu, L.M.; Pfenning, L.H.; Haidukowski, M.; Moretti, A.; Logrieco, A.; Del Ponte, E.M. Fusarium incarnatum-equiseti species complex associated with Brazilian rice: Phylogeny, morphology and toxigenic potential. Int. J. Food Microbiol. 2019, 306, 108267. [Google Scholar] [CrossRef] [PubMed]
- Piacentini, K.C.; Rocha, L.O.; Savi, G.D.; Carnielli-Queiroz, L.; De Carvalho Fontes, L.; Correa, B. Assessment of toxigenic Fusarium species and their mycotoxins in brewing barley grains. Toxins 2019, 11, 31. [Google Scholar] [CrossRef] [PubMed]
- Castellá, G.; Cabañes, F.J. Phylogenetic diversity of Fusarium incarnatum-equiseti species complex isolated from Spanish wheat. Antonie Van Leeuwenhoek 2014, 106, 309–317. [Google Scholar] [CrossRef]
- Thirumalaisamy, P.P.; Dutta, R.; Jadon, K.S.; Nataraja, M.V.; Padvi, R.D.; Rajyaguru, R.; Yusufzai, S. Association and characterization of the Fusarium incarnatum-F. equiseti species complex with leaf blight and wilt of peanut in India. J. Gen. Plant Pathol. 2018, 85, 83–89. [Google Scholar] [CrossRef]
- Gupta, V.S.; Razdan, V.K.; John, D.T.; Sharma, B.C. First report of leaf blight of Cyperus iria caused by Fusarium equiseti in India. Plant Dis. 2013, 97, 838. [Google Scholar] [CrossRef]
- Posada, D.; Crandall, K.A. Evaluation of methods for detecting recombination from DNA sequences: Computer simulations. Proc. Natl. Acad. Sci. USA 2001, 98, 13757–13762. [Google Scholar] [CrossRef]
- Matić, S.; Tabone, G.; Garibaldi, A.; Gullino, M.L. Alternaria leaf spot caused by Alternaria species: An emerging problem on ornamental plants in Italy. Plant Dis. 2020, 104, 2275–2287. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolates. | Latitude and Longitude | Species | GenBank Accession Nos. | ||
|---|---|---|---|---|---|
| tef1 | rpb2 | Beta-Tubulin | |||
| HA-z142 | 126.738196, 45.753014 | F. ipomoeae | OM985077 | OP436018 | OP642121 |
| HA-z11 | 126.738196, 45.753014 | F. ipomoeae | OM985078 | OP436019 | OP642120 |
| HA-z12 | 126.738196, 45.753014 | F. ipomoeae | OM985079 | OP436020 | OP642119 |
| HA-z13 | 126.738196, 45.753014 | F. ipomoeae | OM985080 | OP436021 | OP642118 |
| HA-z14 | 126.738196, 45.753014 | F. ipomoeae | OM985081 | OP436022 | OP642117 |
| HA-z15 | 126.738196, 45.753014 | F. ipomoeae | OM985082 | OP436023 | OP642116 |
| HA-z16 | 126.738196, 45.753014 | F. ipomoeae | OM985083 | OP436024 | OP642115 |
| HA-z17 | 126.738196, 45.753014 | F. ipomoeae | OM985084 | OP436025 | OP642114 |
| HA-z18 | 126.738196, 45.753014 | F. ipomoeae | OM985085 | OP436026 | OP642113 |
| HA-z19 | 126.738196, 45.753014 | F. ipomoeae | OM985086 | OP436027 | OP642112 |
| HA-z20 | 126.738196, 45.753014 | F. ipomoeae | OM985087 | OP436028 | OP642111 |
| HA-z21 | 126.738196, 45.753014 | F. ipomoeae | OM985088 | OP436029 | OP642110 |
| HA-z22 | 126.738196, 45.753014 | F. ipomoeae | OM985089 | OP436030 | OP642109 |
| HA-x22 | 126.868024, 45.850128 | F. ipomoeae | OM985106 | OP436031 | OP642108 |
| HA-xy82 | 126.933932, 45.769353 | F. ipomoeae | OM985109 | OP436032 | OP642122 |
| HA-xy83 | 126.933932, 45.769353 | F. ipomoeae | OM985110 | OP436033 | OP642123 |
| HA-31 | 126.868024, 45.850128 | F. ipomoeae | OM985118 | OP436034 | OP642107 |
| SH-11 | 127.270457, 46.64457 | F. ipomoeae | OM985119 | OP436035 | OP642106 |
| SH-63 | 127.270457, 46.64457 | F. ipomoeae | OM985120 | OP436036 | OP642105 |
| WC-31 | 127.22506, 44.93996 | F. ipomoeae | OM985124 | OP436037 | OP642104 |
| QQ-41 | 124.340195, 47.29158 | F. ipomoeae | OM985125 | OP436038 | OP642103 |
| SH-62 | 127.270457, 46.64457 | F. ipomoeae | OM985126 | OP436039 | OP642124 |
| HA-z201 | 126.738196, 45.753014 | F. ipomoeae | OM985127 | OP436040 | OP642125 |
| HA-21 | 126.868024, 45.850128 | F. ipomoeae | OM985128 | OP436041 | OP642126 |
| HA-22 | 126.868024, 45.850128 | F. ipomoeae | OM985129 | OP436042 | OP642127 |
| HA-x21 | 126.868024, 45.850128 | F. ipomoeae | OM985130 | OP436043 | OP642102 |
| HA-212 | 126.868024, 45.850128 | F. ipomoeae | OM985140 | OP436044 | OP642101 |
| DQ-n22 | 125.835845, 46.329205 | F. ipomoeae | OM985182 | OP436045 | OP642100 |
| JX-21 | 132.477436, 46.339951 | F. ipomoeae | OM985183 | OP436046 | OP642098 |
| DQ-n31 | 125.835845, 46.329205 | F. ipomoeae | OM985184 | OP436047 | OP642099 |
| HA-61 | 126.868024, 45.850128 | F. compactum | OM985144 | OP435951 | OP642130 |
| HA-111 | 126.868024, 45.850128 | F. compactum | OM985102 | OP435952 | OP642131 |
| JX-y11 | 132.477436, 46.339951 | F. compactum | OM985123 | OP435953 | OP642132 |
| HA-621 | 126.868024, 45.850128 | F. compactum | OM985145 | OP435975 | OP642128 |
| SYS-31 | 132.768479, 46.215238 | F. compactum | OM985146 | OP435954 | OP642129 |
| HA-z152 | 126.738196, 45.753014 | F. compactum | OM985147 | OP435955 | OP642133 |
| HA-z31 | 126.738196, 45.753014 | F. compactum | OM985148 | OP435956 | OP642134 |
| HA-z32 | 126.738196, 45.753014 | F. compactum | OM985149 | OP435957 | OP642135 |
| HA-z33 | 126.738196, 45.753014 | F. compactum | OM985150 | OP435958 | OP642136 |
| HA-z34 | 126.738196, 45.753014 | F. compactum | OM985151 | OP435959 | OP642137 |
| HA-z35 | 126.738196, 45.753014 | F. compactum | OM985152 | OP435960 | OP642138 |
| HA-z36 | 126.738196, 45.753014 | F. compactum | OM985153 | OP435961 | OP642139 |
| HA-z37 | 126.738196, 45.753014 | F. compactum | OM985154 | OP435962 | OP642140 |
| HA-z38 | 126.738196, 45.753014 | F. compactum | OM985155 | OP435963 | OP642141 |
| HA-z39 | 126.738196, 45.753014 | F. compactum | OM985156 | OP435964 | OP642142 |
| HA-z310 | 126.738196, 45.753014 | F. compactum | OM985157 | OP435965 | OP642143 |
| HA-z311 | 126.738196, 45.753014 | F. compactum | OM985158 | OP435966 | OP642144 |
| HA-z312 | 126.738196, 45.753014 | F. compactum | OM985159 | OP435967 | OP642145 |
| HA-xy151 | 126.933932, 45.769353 | F. compactum | OM985160 | OP435968 | OP642146 |
| HA-xy31 | 126.933932, 45.769353 | F. compactum | OM985161 | OP435969 | OP642147 |
| HA-a11 | 126.868024, 45.850128 | F. compactum | OM985162 | OP435970 | OP642152 |
| HA-42 | 126.868024, 45.850128 | F. compactum | OM985163 | OP435971 | OP642148 |
| JX-52 | 132.477436, 46.339951 | F. compactum | OM985164 | OP435972 | OP642149 |
| JX-121 | 132.477436, 46.339951 | F. compactum | OM985165 | OP435973 | OP642150 |
| JX-31 | 132.477436, 46.339951 | F. compactum | OM985166 | OP435974 | OP642151 |
| HA-x12 | 126.868024, 45.850128 | F. citri | OM985167 | OP435950 | OP642166 |
| QTH-21 | 131.139405, 45.733699 | F. citri | OM985168 | OP435949 | OP642167 |
| HA-z1125 | 126.738196, 45.753014 | F. citri | OM985169 | OP435948 | OP642158 |
| HA-z171 | 126.738196, 45.753014 | F. citri | OM985170 | OP435947 | OP642165 |
| HA-z172 | 126.738196, 45.753014 | F. citri | OM985171 | OP435946 | OP642164 |
| HA-z173 | 126.738196, 45.753014 | F. citri | OM985172 | OP435945 | OP642163 |
| HA-z174 | 126.738196, 45.753014 | F. citri | OM985173 | OP435944 | OP642162 |
| HA-z175 | 126.738196, 45.753014 | F. citri | OM985174 | OP435943 | OP642161 |
| HA-z176 | 126.738196, 45.753014 | F. citri | OM985175 | OP435942 | OP642160 |
| HA-z177 | 126.738196, 45.753014 | F. citri | OM985176 | OP435941 | OP642159 |
| HA-z1126 | 126.738196, 45.753014 | F. citri | OM985177 | OP435940 | OP642157 |
| HA-xy141 | 126.933932, 45.769353 | F. citri | OM985178 | OP435939 | OP642156 |
| HA-z203 | 126.738196, 45.753014 | F. citri | OM985179 | OP435938 | OP642155 |
| HA-x11 | 126.868024, 45.850128 | F. flagelliforme | OM985104 | OP435921 | OP642153 |
| HA-x51 | 126.868024, 45.850128 | F. flagelliforme | OM985105 | OP435920 | OP642154 |
| HA-a31 | 126.868024, 45.850128 | F. graminearum | OM985090 | OP435980 | OP642200 |
| HG-11 | 130.440826, 47.312952 | F. graminearum | OM985091 | OP435981 | OP642201 |
| QTH-23 | 131.139405, 45.733699 | F. graminearum | OM985103 | OP435982 | OP642202 |
| SH-x72 | 127.270457, 46.64457 | F. graminearum | OM985108 | OP435983 | OP642203 |
| SYS-y21 | 132.768479, 46.215238 | F. graminearum | OM985111 | OP435984 | OP642204 |
| SYS-21 | 132.768479, 46.215238 | F. graminearum | OM985199 | OP435985 | OP642205 |
| SYS-141 | 132.768479, 46.215238 | F. graminearum | OM985200 | OP435986 | OP642206 |
| SYS-142 | 132.768479, 46.215238 | F. graminearum | OM985201 | OP435987 | OP642207 |
| SYS-143 | 132.768479, 46.215238 | F. graminearum | OM985202 | OP435988 | OP642208 |
| SYS-144 | 132.768479, 46.215238 | F. graminearum | OM985203 | OP435989 | OP642209 |
| SYS-145 | 132.768479, 46.215238 | F. graminearum | OM985204 | OP435990 | OP642210 |
| SYS-146 | 132.768479, 46.215238 | F. graminearum | OM985205 | OP435991 | OP642211 |
| SYS-147 | 132.768479, 46.215238 | F. graminearum | OM985206 | OP435992 | OP642212 |
| HA-a142 | 126.868024, 45.850128 | F. graminearum | OM985138 | OP435993 | OP642213 |
| SYS-x71 | 131.583118, 46.462499 | F. asiaticum | OM985092 | OP436053 | OP642088 |
| SYS-x91 | 131.583118, 46.462499 | F. asiaticum | OM985093 | OP436054 | OP642089 |
| HA-x72 | 126.868024, 45.850128 | F. asiaticum | OM985094 | OP436055 | OP642090 |
| HG-x62 | 130.440826, 47.312952 | F. asiaticum | OM985095 | OP436056 | OP642091 |
| SYS-x62 | 131.583118, 46.462499 | F. asiaticum | OM985096 | OP436057 | OP642092 |
| SYS-x131 | 131.583118, 46.462499 | F. asiaticum | OM985097 | OP436058 | OP642093 |
| SYS-x132 | 131.583118, 46.462499 | F. asiaticum | OM985098 | OP436059 | OP642094 |
| SYS-x133 | 131.583118, 46.462499 | F. asiaticum | OM985099 | OP436060 | OP642095 |
| SYS-x134 | 131.583118, 46.462499 | F. asiaticum | OM985100 | OP436061 | OP642096 |
| SYS-x135 | 131.583118, 46.462499 | F. asiaticum | OM985101 | OP436062 | OP642097 |
| HA-zh142 | 126.738196, 45.753014 | F. temperatum | OM985107 | OP436049 | OP642174 |
| QTH-X332 | 131.139405, 45.733699 | F. temperatum | OM985131 | OP436050 | OP642171 |
| QTH-X331 | 131.139405, 45.733699 | F. temperatum | OM985132 | OP436051 | OP642173 |
| QTH-X33 | 131.139405, 45.733699 | F. temperatum | OM985133 | OP436052 | OP642172 |
| HA-z113 | 126.738196, 45.753014 | Fusarium sp. | OM985112 | OP436063 | OP642168 |
| HA-b113 | 126.738196, 45.753014 | Fusarium sp. | OM985113 | OP436064 | OP642170 |
| HA-Z1131 | 126.738196, 45.753014 | Fusarium sp. | OM985143 | OP436065 | OP642169 |
| SYS-x11 | 131.583118, 46.462499 | F. sporotrichioides | OM985209 | OP436017 | OP642176 |
| SYS-x61 | 131.583118, 46.462499 | F. sporotrichioides | OM985210 | OP436016 | OP642177 |
| SYS-x1 | 131.583118, 46.462499 | F. sporotrichioides | OM985211 | OP436015 | OP642178 |
| SYS-x2 | 131.583118, 46.462499 | F. sporotrichioides | OM985212 | OP436014 | OP642179 |
| HG-12 | 130.440826, 47.312952 | F. sporotrichioides | OM985213 | OP436013 | OP642180 |
| SYS-33 | 132.768479, 46.215238 | F. sporotrichioides | OM985214 | OP436012 | OP642181 |
| SYS-101 | 132.768479, 46.215238 | F. sporotrichioides | OM985215 | OP436011 | OP642182 |
| SYS-102 | 132.768479, 46.215238 | F. sporotrichioides | OM985216 | OP436010 | OP642183 |
| SYS-103 | 132.768479, 46.215238 | F. sporotrichioides | OM985217 | OP436009 | OP642184 |
| SYS-104 | 132.768479, 46.215238 | F. sporotrichioides | OM985218 | OP436008 | OP642185 |
| SYS-105 | 132.768479, 46.215238 | F. sporotrichioides | OM985219 | OP436007 | OP642186 |
| SYS-51 | 132.768479, 46.215238 | F. sporotrichioides | OM985220 | OP436006 | OP642187 |
| HG-y102 | 130.440826, 47.312952 | F. sporotrichioides | OM985121 | OP436005 | OP642188 |
| HG-DBy101 | 130.440826, 47.312952 | F. sporotrichioides | OM985122 | OP436004 | OP642189 |
| SH-z61 | 127.270457, 46.64457 | F. acuminatum | OM985115 | OP435923 | OP642072 |
| SH-61 | 127.270457, 46.64457 | F. acuminatum | OM985116 | OP435922 | OP642073 |
| SH-41 | 127.270457, 46.64457 | F. acuminatum | OM985117 | OP435924 | OP642074 |
| HA-a72 | 126.868024, 45.850128 | F. acuminatum | OM985221 | OP435925 | OP642075 |
| HA-a161 | 126.868024, 45.850128 | F. acuminatum | OM985222 | OP435926 | OP642076 |
| HA-a162 | 126.868024, 45.850128 | F. acuminatum | OM985223 | OP435927 | OP642077 |
| HA-a163 | 126.868024, 45.850128 | F. acuminatum | OM985224 | OP435928 | OP642078 |
| HA-a164 | 126.868024, 45.850128 | F. acuminatum | OM985225 | OP435929 | OP642079 |
| HA-a1211 | 126.868024, 45.850128 | F. armeniacum | OM985134 | OP435979 | OP642214 |
| HA-13 | 126.868024, 45.850128 | F. armeniacum | OM985135 | OP435978 | OP642215 |
| HA-a121 | 126.868024, 45.850128 | F. armeniacum | OM985136 | OP435976 | OP642216 |
| HA-a122 | 126.868024, 45.850128 | F. armeniacum | OM985137 | OP435977 | OP642217 |
| HL-42 | 132.943466, 45.768947 | F. verticillioides | OM985139 | OP435994 | OP642190 |
| DQ-n32 | 125.835845, 46.329205 | F. verticillioides | OM985141 | OP435995 | OP642191 |
| SH-n12 | 127.270457, 46.64457 | F. verticillioides | OM985142 | OP435996 | OP642192 |
| JX-123 | 132.477436, 46.339951 | F. verticillioides | OM985181 | OP435997 | OP642193 |
| SH-n11 | 127.270457, 46.64457 | F. verticillioides | OM985187 | OP435998 | OP642194 |
| SH-n201 | 127.270457, 46.64457 | F. verticillioides | OM985188 | OP435999 | OP642195 |
| SH-n202 | 127.270457, 46.64457 | F. verticillioides | OM985189 | OP436000 | OP642197 |
| SH-n203 | 127.270457, 46.64457 | F. verticillioides | OM985190 | OP436001 | OP642197 |
| SH-n204 | 127.270457, 46.64457 | F. verticillioides | OM985191 | OP436002 | OP642199 |
| SH-n205 | 127.270457, 46.64457 | F. verticillioides | OM985192 | OP436003 | OP642199 |
| JX-3352 | 132.477436, 46.339951 | F. glycines | OM985193 | OP435937 | OP642080 |
| JX-335 | 132.477436, 46.339951 | F. glycines | OM985194 | OP435930 | OP642081 |
| HA-171 | 126.868024, 45.850128 | F. glycines | OM985195 | OP435936 | OP642082 |
| HA-172 | 126.868024, 45.850128 | F. glycines | OM985196 | OP435935 | OP642083 |
| HA-173 | 126.868024, 45.850128 | F. glycines | OM985197 | OP435934 | OP642084 |
| HA-174 | 126.868024, 45.850128 | F. glycines | OM985198 | OP435933 | OP642085 |
| WC-b53 | 127.22506, 44.93996 | F. glycines | OM985208 | OP435932 | OP642086 |
| HA-z1412 | 126.738196, 45.753014 | F. glycines | OM985180 | OP435931 | OP642087 |
| WC-22 | 127.22506, 44.93996 | F. annulatum | OM985207 | OP436048 | OP642175 |
| NRRL 34034 | - | F. ipomoeae | GQ505636 | GQ505814 | - |
| LC0455 | - | F. ipomoeae | MK289580 | MK289734 | - |
| NRRL 45996 | - | F. ipomoeae | GQ505671 | GQ505849 | - |
| CBS 140909 | - | F. ipomoeae | MN170479 | MN170412 | - |
| NRRL 28029 | - | F. compactum | GQ505602 | GQ505780 | - |
| NRRL 36318 | - | F. compactum | GQ505646 | GQ505824 | - |
| NRRL 6548 | - | F. flagelliforme | GQ505589 | GQ505767 | - |
| CBS 731.87 | - | F. flagelliforme | GQ505600 | GQ505778 | - |
| LC12147 | - | F. arcuatisporum | MK289584 | MK289739 | - |
| NRRL 32997 | - | F. arcuatisporum | GQ505624 | GQ505802 | - |
| NRRL 45997 | - | F. clavus | GQ505672 | GQ505850 | - |
| NRRL 34037 | - | F. clavus | GQ505638 | GQ505638 | - |
| LC7937 | - | F. citri | MK289640 | GQ505816 | - |
| LC7922 | - | F. citri | MK289634 | MK289788 | - |
| NRRL 66939 | - | Fusarium sp. | MW233217 | MW233561 | - |
| FRC R-9121 | - | Fusarium sp. | MW233213 | MW233557 | - |
| CBS 462.94 | - | F. sporotrichioides | MN120771 | MN120750 | - |
| NRRL 53430 | - | F. sibiricum | HM744684 | MW233474 | - |
| NRRL 6227 | - | F. armeniacum | HM744692 | JX171560 | - |
| FRC R-09335 | - | F. armeniacum | GQ915501 | GQ915485 | - |
| NRRL 13818 | - | F. asiaticum | AF212451 | MW233412 | - |
| NRRL 46738 | - | F. asiaticum | FJ240299 | - | - |
| NL19-100008 | - | F. graminearum | MZ921906 | MZ921775 | - |
| CBS 136009 | - | F. graminearum | MW928838 | MW928826 | - |
| NRRL 54216 | - | F. acuminatum | HM068314 | HM068334 | - |
| JW 289003 | - | F. acuminatum | MZ921908 | MZ921777 | - |
| CBS 130180 | - | F. verticillioides | MW402024 | MW402740 | - |
| CBS 131389 | - | F. verticillioides | MN534047 | MN534288 | - |
| CBS 135541 | - | F. temperatum | MW402051 | KU604284 | - |
| CBS 130323 | - | Fusarium sp. | MH485018 | MH484927 | - |
| CBS 214.49 | - | F. glycines | MH484960 | MH484869 | - |
| CBS 127316 | - | F. annulatum | MW402021 | MW402738 | - |
| CBS 100001 | - | Macroconia leptosphaeriae | KM231959 | HQ728164 | - |
| Geographic Origins | Number of Fusarium Isolates | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| FIESC | F. sporotrichioides | F. armeniacum | F. asiaticum | F. graminearum | Fusarium sp. | F. acuminatum | F. glycines | F. annulatum | F. temperatum | F. verticillioides | |
| Daqing city | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| Harbin city | 56 | 0 | 4 | 1 | 2 | 3 | 5 | 5 | 0 | 1 | 0 |
| Hegang city | 0 | 3 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Jixi city | 5 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 1 |
| Qiqihar city | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Qitaihe city | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 3 | 0 |
| Shuangyashan city | 1 | 11 | 0 | 8 | 9 | 0 | 0 | 0 | 0 | 0 | 0 |
| Suihua city | 3 | 0 | 0 | 0 | 1 | 0 | 3 | 0 | 0 | 0 | 7 |
| Hulin country | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| Wuchang city | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 |
| Total | 70 | 14 | 4 | 10 | 14 | 3 | 8 | 8 | 1 | 4 | 10 |
| Percentage a | 47.95 | 9.59 | 2.74 | 6.85 | 9.59 | 2.05 | 5.48 | 5.48 | 0.68 | 2.74 | 6.85 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, X.; Zhang, L.; Yang, X.; Shen, G.; Wang, S.; Teng, H.; Yang, C.; Liu, X.; Wang, X.; Zhao, J.; et al. Fusarium Species Associated with Maize Leaf Blight in Heilongjiang Province, China. J. Fungi 2022, 8, 1170. https://doi.org/10.3390/jof8111170
Xu X, Zhang L, Yang X, Shen G, Wang S, Teng H, Yang C, Liu X, Wang X, Zhao J, et al. Fusarium Species Associated with Maize Leaf Blight in Heilongjiang Province, China. Journal of Fungi. 2022; 8(11):1170. https://doi.org/10.3390/jof8111170
Chicago/Turabian StyleXu, Xi, Li Zhang, Xilang Yang, Guijin Shen, Shuo Wang, Haolin Teng, Chunbo Yang, Xueyan Liu, Xiangjing Wang, Junwei Zhao, and et al. 2022. "Fusarium Species Associated with Maize Leaf Blight in Heilongjiang Province, China" Journal of Fungi 8, no. 11: 1170. https://doi.org/10.3390/jof8111170
APA StyleXu, X., Zhang, L., Yang, X., Shen, G., Wang, S., Teng, H., Yang, C., Liu, X., Wang, X., Zhao, J., & Xiang, W. (2022). Fusarium Species Associated with Maize Leaf Blight in Heilongjiang Province, China. Journal of Fungi, 8(11), 1170. https://doi.org/10.3390/jof8111170